

Растительные ресурсы, 2019, T. 55, № 4, стр. 473-489
Пространственное распределение запасов напочвенного покрова и лесной подстилки в средневозрастных сосновых лесах Кольского полуострова
И. В. Лянгузова 1, *, П. А. Примак 2
1 Ботанический институт им. В.Л. Комарова РАН
г. Санкт-Петербург, Россия
2 Санкт-Петербургский государственный лесотехнический университет
г. Санкт-Петербург, Россия
* E-mail: ILyanguzova@binran.ru
Поступила в редакцию 25.07.2019
После доработки 10.08.2019
Принята к публикации 28.08.2019
Аннотация
Приведены результаты исследования внутриценотической неоднородности почвенно-растительного покрова в средневозрастных сосновых лесах разной типологической принадлежности на фоновой территории Кольского полуострова. Установлены различия в запасе биомассы растений травяно-кустарничкового и мохово-лишайникового ярусов, а также запасов растительного опада и лесной подстилки на приствольных и подкроновых участках деревьев Pinus sylvestris L. и Betula pubescens Ehrh. и в межкроновом пространстве в лишайниковом сосновом редколесье, лишайниково-зеленомошном и зеленомошном фитоценозах. Сопоставление запасов биомассы компонентов напочвенного покрова (кустарничков, мхов и лишайников) в различных микроместообитаниях выявило существенное внутриэкосистемное разнообразие этого параметра. Общие запасы надземной биомассы и всего живого напочвенного покрова (с учетом подземных частей) в лишайниковом сосновом редколесье и сосняке лишайниково-зеленомошном существенно увеличиваются от приствольных к межкроновым микроместообитаниям, в сосняке зеленомошном достоверные различия отсутствуют. Противоположная закономерность обнаружена для запаса растительного опада независимо от типа растительного сообщества. Наиболее равномерно по площади биогеоценоза распределен запас лесной подстилки во всех исследуемых сообществах. В целом микромозаика почвенно-растительного покрова наиболее ярко выражена в сосновом редколесье, что обусловлено контрастным гидротермическим режимом экотопа и разреженностью древостоя. В более густых и высокополнотных сосняках зеленомошных запасы органического вещества в почвенно-растительном покрове распределены достаточно равномерно.
Для северных лесов характерна ярко выраженная неоднородность ценотической среды и мозаичность напочвенного покрова, обусловленная влиянием древесного яруса. Деревья – эдификаторы фитоценоза вызывают пространственную неоднородность и временнýю изменчивость освещенности, микроклимата, распределения атмосферных осадков, эвапотранспирации, водного, температурного и питательного режимов почвы. Микромозаика растительности в лесных экосистемах определяет пространственную гетерогенность лесных почв, что наиболее четко отражается на составе их верхних горизонтов, формирующихся из растительного опада. Изучение микромозаики напочвенного покрова и верхних горизонтов почв в пределах лесных биогеоценозов наиболее часто проводилось в северотаежных еловых лесах или в темнохвойных лесах средней тайги, где внутриценотическая мозаичность выражена наиболее контрастно [1–10]. Значительно меньше работ посвящено изучению пространственной гетерогенности напочвенного покрова и верхних горизонтов почв в сосновых биогеоценозах, что, по-видимому, связано с тем, что фитоценозы сосновых лесов отличаются более равномерной освещенностью [9, 11–14].
Для исследования элементов внутриценотической неоднородности растительности и почвенного покрова обычно используют такие понятия, как “парцелла” [12, 15], “фитогенное поле” [16, 17], “зоны или пространства” [7, 12], “микросайты” [4, 5, 18, 19], “тессеры” [1, 20, 21], “элементарный биогеоареал” [2, 3, 8, 9, 22]. Определение “микроместообитания” или “микросайты” применимы для сопряженной оценки неоднородности нижних ярусов лесного сообщества и почвенного покрова [23]. В наших исследованиях мы придерживаемся определения “микроместообитания”.
Одним из наиболее важных параметров, определяющих продуктивность лесных сообществ, является накопление органического вещества в надземной биомассе высших растений, мхов и лишайников, а также в растительном опаде и лесной подстилке. В лесах Кольского полуострова биомасса нижних ярусов может достигать 25% и более от ее общего запаса, а годичная продукция и участие в биогеохимическом круговороте – 50–60% [11].
Растительный опад как источник органического вещества почв и элементов питания для биоты является одним из ключевых компонентов биогеохимических циклов в лесных сообществах. Размеры, качество и структура опада зависят от видового состава растительных сообществ, вклада различных видов растений в структуру и запас биомассы и определяют характеристики лесной подстилки [7, 8, 11, 14, 24]. Кроме того, растительный опад является источником питания и местом обитания большинства беспозвоночных животных и микроорганизмов, обеспечивающих его минерализацию [25]. Опад хвои и некоторых видов мхов отличается замедленным разложением и затрудняет образование гумуса. Противоположные свойства имеют листья березы, скручивающиеся при опадании, что создает аэробные условия, благоприятствующие разложению опада. Примесь березы в хвойных лесах усиливает нитрификацию, которой способствуют также многие травянистые растения, за исключением злаков [26].
Хорошо известно, что лесная подстилка в значительной степени определяет химические свойства, водный и тепловой режим верхнего корнеобитаемого слоя почв, является областью концентрации сосущих корней, банка семян, средой обитания большинства видов грибов, микроорганизмов и беспозвоночных. Скорость минерализации органического вещества почв и высвобождение минеральных форм азота определяют продуктивность лесных экосистем [27]. Прямые корреляции были установлены между объемом древесины, видовым разнообразием напочвенного покрова и почвенным плодородием в ряду от бореальных до неморальных типов лесов южной Эстонии [28].
В связи с этим представляет интерес провести сравнительный анализ накопления органического вещества в живом напочвенном покрове, растительном опаде и лесной подстилке сосновых лесов при совместном произрастании Pinus sylvestris L. и Betula pubescens Ehrh.
Цель данного исследования – оценка накопления надземной и подземной биомассы компонентами напочвенного покрова (лишайники, мхи, кустарнички), а также запасов растительного опада и лесной подстилки в разных микроместообитаниях средневозрастных сосновых лесов северной тайги (Кольский полуостров).
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Исследования проводили в средневозрастных сосновых лесах, расположенных в центральной части Кольского полуострова в среднем течении р. Ливы (67°49′ с.ш., 31°20′ в.д.), произрастающих на Al-Fe-гумусовых подзолах (Albic Rustic Podzols, согласно WRB) [29].
Давность последнего пожара в исследованных сообществах составляет в среднем 90 лет. Эдификатором древесного яруса является Pinus sylvestris L., в составе древостоев присутствует Betula pubescens Ehrh. (табл. 1). В травяно-кустарничковом ярусе доминирующими видами являются кустарнички Vaccinium vitis-idaea, V. myrtillus, Empetrum hermaphroditum, в мохово-лишайниковом – зеленые мхи Pleurozium schreberi, Dicranum sp. и лишайники Cladonia rangiferina, Cl. stellaris, Cl. mitis.
Таблица 1.
Краткая таксационная характеристика древостоев на пробных площадях Table 1. Brief forest stand characteristic of the permanent sample plots (PSP)
| Номер ППП Number of permanent sample plot (PSP) |
Вид Species |
Возраст,
лет Age, years |
Диаметр
на высоте
1.3 м, см Diameter at breath height, cm |
Высота, м Нeight, m |
Плотность, экз./га Density, ind./ha |
Сумма
площадей
сечений, м2/га Basal area, m2/ha |
|---|---|---|---|---|---|---|
| Subpinetum cladinosum | ||||||
| 1 | С* | 188 | 34.2 | 12.0 | 13 | 1.25 |
| С | 72 | 11.8 | 9.2 | 453 | 5.0 | |
| Б (B) | Н.д. | 4.8 | 5.0 | 40 | 0.28 | |
| Pinetum cladinoso-hylocomiosum | ||||||
| 2 | С | 70 | 11.9 | 11.5 | 1156 | 12.9 |
| Б (B) | Н.д. | 7.9 | 7.7 | 478 | 2.30 | |
| Pinetum hylocomiosum | ||||||
| 3 | С | 72 | 13.0 | 12.4 | 2260 | 27.1 |
| Б (B) | Н.д. | 6.3 | 7.0 | 220 | 0.78 | |
Исследование проводили на постоянных пробных площадях (ППП) размером 30 × 50 м, заложенных в сосновых лесах разной типологической принадлежности: ППП 1 – лишайниковое редколесье (Subpinetum cladinosum), ППП 2 – сосняк лишайниково-зеленомошный (Pinetum cladinoso-hylocomiosum), ППП 3 – сосняк зеленомошный (Pinetum hylocomiosum).
На каждой ППП были заложены трансекты, на которых с интервалом 1 м размещали учетные площадки размером 10 × 10 см. Площадки располагались в следующих типах микроместообитаний: А – в приствольной зоне в 20 см от стволов деревьев сосны или березы с 4-х сторон света; Б – в подкроновых пространствах деревьев; В – на межкроновых участках в “окнах” полога древостоя. Всего заложено 170 учетных площадок.
В пределах каждой учетной площадки был срезан живой напочвенный покров и разобран на следующие компоненты: кустарнички, лишайники, мхи; была измерена толщина лесной подстилки, отобран растительный опад и органогенный горизонт (О), из которого были извлечены все подземные части растений. Образцы растительного материала и лесной подстилки высушены до воздушно-сухого состояния и взвешены. Общий запас надземной биомассы включает надземные части кустарничков, мхов и лишайников, а в общий запас живого напочвенного покрова включена биомасса подземных частей растений. Запас биомассы разных компонентов напочвенного покрова и запасы растительного опада и лесной подстилки рассчитаны в г/м2.
Проверка выборок исследуемых параметров на соответствие закону нормального распределения показала, что распределения большинства параметров значимо отличаются от нормального распределения. В связи с этим при оценке значимости различий использовали непараметрические критерии Краскела–Уоллиса (H) и Манна–Уитни (z), при уровне значимости р < 0.05 различия считали достоверными. На рисунках и в таблицах представлены средние значения и их стандартная ошибка.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
В сосновом лишайниковом редколесье (ППП 1) запас биомассы лишайников достоверно увеличивается от приствольных к межкроновым микроместообитаниям независимо от видовой принадлежности дерева, в то время как изменение запаса биомассы мхов от приствольных микроместообитаний Betula pubescens к межкроновым пространствам имеет противоположную закономерность (табл. 2). Запас надземной и подземной биомассы кустарничков в межкроновых пространствах существенно меньше по сравнению с приствольными и подкроновыми микроместообитаниями деревьев Pinus sylvestris. В приствольной части и под кронами деревьев Betula pubescens запас биомассы кустарничков достоверно не отличается от запаса в межкроновых пространствах.
Таблица 2.
Запас (г/м2) биомассы компонентов напочвенного покрова в разных микроместообитаниях исследуемых сосновых сообществ Table 2. Stock (g/m2) of ground cover phytomass components in different microhabitats of pine communities
| Компонент Component |
Микроместообитания Microhabitats |
Критерий
Краскела–Уоллиса (H) Kruskal–Wallis criteria (H) |
Уровень
значимости (p) Significance level (p) |
||
|---|---|---|---|---|---|
| приствольные at tree base |
подкроновые under crowns |
межкроновые in canopy gaps |
|||
| Subpinetum cladinosum (ППП 1, PSP 1) | |||||
| Лишайники Lichens |
$\frac{{238 \pm 76}}{{168 \pm 53}}$ | $\frac{{400 \pm 156}}{{397 \pm 32}}$ | 639 ± 52 | $\frac{{14.2}}{{20.7}}$ | $\frac{{{\mathbf{0}}{\mathbf{.001}}{\text{*}}}}{{{\mathbf{0}}.{\mathbf{000}}}}$ |
| Мхи Mosses |
$\frac{0}{{56 \pm 25}}$ | $\frac{0}{{13 \pm 7}}$ | 3.3 ± 2.3 | $\frac{{4.5}}{{13.6}}$ | $\frac{{0.104}}{{0.001}}$ |
| Кустарнички: Dwarf-shrubs |
|||||
| надземные части aboveground parts |
$\frac{{112 \pm 24}}{{66 \pm 43}}$ | $\frac{{154 \pm 56}}{{36 \pm 19}}$ | 50 ± 16 | $\frac{{9.8}}{{0.67}}$ | $\frac{{{\mathbf{0}}{\mathbf{.007}}}}{{0.72}}$ |
| подземные части belowground parts |
$\frac{{161 \pm 27}}{{123 \pm 41}}$ | $\frac{{191 \pm 35}}{{156 \pm 75}}$ | 97 ± 15 | $\frac{{6.2}}{{0.002}}$ | $\frac{{{\mathbf{0}}{\mathbf{.044}}}}{{0.99}}$ |
| Pinetum cladinoso-hylocomiosum (ППП 2, PSP 2) | |||||
| Лишайники Lichens |
$\frac{{46 \pm 20}}{{5 \pm 2}}$ | $\frac{{84 \pm 25}}{{48 \pm 39}}$ | 80 ± 24 | $\frac{{2.27}}{{7.18}}$ | $\frac{{0.32}}{{{\mathbf{0}}.{\mathbf{028}}}}$ |
| Мхи Mosses |
$\frac{{28 \pm 17}}{{59 \pm 14}}$ | $\frac{{22.6 \pm 81}}{{70 \pm 62}}$ | 148 ± 37 | $\frac{{13.7}}{{4.79}}$ | $\frac{{{\mathbf{0}}{\mathbf{.001}}}}{{0.09}}$ |
| Кустарнички: Dwarf-shrubs |
|||||
| надземные части aboveground parts | $\frac{{106 \pm 25}}{{69 \pm 8}}$ | $\frac{{91 \pm 18}}{{66 \pm 25}}$ | 68 ± 10 | $\frac{{2.31}}{{0.09}}$ | $\frac{{0.32}}{{0.96}}$ |
| подземные части belowground parts | $\frac{{289 \pm 32}}{{225 \pm 37}}$ | $\frac{{362 \pm 40}}{{248 \pm 67}}$ | 213 ± 25 | $\frac{{8.45}}{{0.51}}$ | $\frac{{{\mathbf{0}}{\mathbf{.015}}}}{{0.78}}$ |
| Pinetum hylocomiosum (ППП3, PSP3) | |||||
| Лишайники Lichens |
$\frac{{5 \pm 5}}{{7 \pm 7}}$ | $\frac{0}{0}$ | 51 ± 25 | $\frac{{11.2}}{{12.1}}$ | $\frac{{{\mathbf{0}}{\mathbf{.004}}}}{{{\mathbf{0}}.{\mathbf{002}}}}$ |
| Мхи Mosses |
$\frac{{114 \pm 12}}{{62 \pm 16}}$ | $\frac{{117 \pm 59}}{{58 \pm 12}}$ | 94 ± 26 | $\frac{{0.89}}{{1.66}}$ | $\frac{{0.64}}{{0.44}}$ |
| Кустарнички: Dwarf-shrubs |
|||||
| надземные части aboveground parts | $\frac{{44 \pm 8}}{{69 \pm 17}}$ | $\frac{{38 \pm 19}}{{59 \pm 10}}$ | 38 ± 27 | $\frac{{1.75}}{{2.58}}$ | $\frac{{0.42}}{{0.28}}$ |
| подземные части belowground parts | $\frac{{346 \pm 44}}{{360 \pm 77}}$ | $\frac{{406 \pm 7}}{{548 \pm 63}}$ | 346 ± 70 | $\frac{{0.89}}{{4.88}}$ | $\frac{{0.64}}{{0.09}}$ |
Примечание. * – Здесь и в табл. 3: жирным шрифтом выделены величины уровня значимости, указывающие на достоверные различия параметра в разных микроместообитаниях. Note. * – Here and in the tab. 3 in bold, are the values of significance level indicating significant differences of the parameter in different microhabitats.
На приствольных участках под деревьями Pinus sylvestris наблюдаются достоверно более высокие величины массы растительного опада, толщины и запаса лесной подстилки по сравнению с подкроновым и межкроновым пространством, в то время как с увеличением расстояния от стволов Betula pubescens достоверно снижается только толщина подстилки (табл. 3).
Таблица 3.
Запас (г/м2) массы опада и подстилки в разных микроместообитаниях исследуемых сосновых лесов Table 3. Stock (g/m2) of plant waste and forest litter in different microhabitats of pine forests
| Компонент Component |
Микроместообитания Microhabitats |
Критерий Краскела-Уоллиса (H) Kruskal–Wallis criteria | Уровень
значимости (p) Significance level (p) |
||
|---|---|---|---|---|---|
| приствольные at tree base |
подкроновые under crowns |
межкроновые in canopy gaps |
|||
| Subpinetum cladinosum (ППП 1, PSP 1) | |||||
| Масса опада Weight of plant waste |
$\frac{{1884 \pm 256}}{{1110 \pm 84}}$ | $\frac{{980 \pm 100}}{{796 \pm 100}}$ | 910 ± 86 | $\frac{{14.4}}{{5.6}}$ | $\frac{{{\mathbf{0}}{\mathbf{.001}}}}{{{\mathbf{0}}.{\mathbf{049}}}}$ |
| Масса подстилки Weight of forest litter |
$\frac{{2469 \pm 272}}{{1653 \pm 294}}$ | $\frac{{1582 \pm 254}}{{1305 \pm 221}}$ | 1513 ± 192 | $\frac{{8.3}}{{0.30}}$ | $\frac{{{\mathbf{0}}{\mathbf{.016}}}}{{0.86}}$ |
| Толщина подстилки, см Thickness of forest litter, cm |
$\frac{{4.5 \pm 0.4}}{{3.1 \pm 0.6}}$ | $\frac{{3.3 \pm 0.7}}{{2.1 \pm 0.4}}$ | 1.2 ± 0.2 | $\frac{{25.0}}{{13.1}}$ | $\frac{{{\mathbf{0}}{\mathbf{.000}}}}{{{\mathbf{0}}.{\mathbf{001}}}}$ |
| Pinetum cladinoso-hylocomiosum (ППП 2, PSP 2) | |||||
| Масса опада Weight of plant waste |
$\frac{{1653 \pm 176}}{{1020 \pm 69}}$ | $\frac{{1446 \pm 168}}{{798 \pm 287}}$ | 734 ± 65 | $\frac{{20.6}}{{9.7}}$ | $\frac{{{\mathbf{0}}{\mathbf{.000}}}}{{{\mathbf{0}}.{\mathbf{008}}}}$ |
| Масса подстилки Weight of forest litter |
$\frac{{3827 \pm 640}}{{3824 \pm 332}}$ | $\frac{{4814 \pm 472}}{{2823 \pm 511}}$ | 3890 ± 316 | $\frac{{3.90}}{{2.77}}$ | $\frac{{0.14}}{{0.25}}$ |
| Толщина подстилки, см Thickness of forest litter, cm |
$\frac{{5.0 \pm 0.6}}{{4.5 \pm 0.4}}$ | $\frac{{4.4 \pm 0.3}}{{4.3 \pm 0.7}}$ | 3.9 ± 0.5 | $\frac{{3.78}}{{1.60}}$ | $\frac{{0.15}}{{0.45}}$ |
| Сосняк зеленомошный (ППП 3, PSP 3) | |||||
| Масса опада Weight of plant waste |
$\frac{{1607 \pm 121}}{{1700 \pm 322}}$ | $\frac{{1239 \pm 262}}{{950 \pm 120}}$ | 863 ± 82 | $\frac{{10.24}}{{8.83}}$ | $\frac{{{\mathbf{0}}{\mathbf{.006}}}}{{{\mathbf{0}}.{\mathbf{012}}}}$ |
| Масса подстилки Weight of forest litter |
$\frac{{4912 \pm 391}}{{5816 \pm 847}}$ | $\frac{{3488 \pm 208}}{{4842 \pm 525}}$ | 4552 ± 795 | $\frac{{1.96}}{{1.63}}$ | $\frac{{0.86}}{{0.44}}$ |
| Толщина подстилки, см Thickness of forest litter, cm |
$\frac{{7.1 \pm 0.5}}{{6.8 \pm 0.7}}$ | $\frac{{3.5 \pm 1.5}}{{7.8 \pm 0.7}}$ | 4.0 ± 0.4 | $\frac{{9.27}}{{11.1}}$ | $\frac{{{\mathbf{0}}{\mathbf{.010}}}}{{{\mathbf{0}}.{\mathbf{004}}}}$ |
Запас биомассы надземных частей кустарничков, масса растительного опада, запас и толщина подстилки на приствольных участках под деревьями Pinus sylvestris достоверно выше, чем в тех же микроместообитаниях под деревьями Betula pubescens (z = 2.12–2.56, p < 0.05), в то время как запас биомассы мхов является достоверно более низким (z = –3.64, p < 0.001). В подкроновых микроместообитаниях сравниваемых видов запасы органического вещества статистически не различаются, за исключением запаса надземной биомассы кустарничков, который достоверно выше под кронами Pinus sylvestris (z = 2.00, p < 0.05).
В сосняке лишайниково-зеленомошном (ППП 2) запас биомассы мхов на приствольных и подкроновых участках деревьев Pinus sylvestris значительно ниже, чем в межкроновых микроместообитаниях (табл. 2). От приствольных участков деревьев Betula pubescens к межкроновым пространствам наблюдается достоверное снижение запаса биомассы мхов.
Запас надземной и подземной биомассы кустарничков на приствольных и покроновых участках деревьев Pinus sylvestris достоверно выше по сравнению с межкроновыми микроместообитаниями (z = 2.39–2.44, p = 0.015–0.017). В то же время в приствольном и подкроновом пространстве деревьев Betula pubescens эти показатели идентичны наблюдаемым в межкроновом пространстве.
В рассматриваемом сообществе масса растительного опада в приствольной зоне и в подкроновом пространстве деревьев Pinus sylvestris, а также на приствольных участках деревьев Betula pubescens достоверно выше, чем в “окнах древостоя” (табл. 3). Толщина и запас лесной подстилки по типам микроместообитаний достоверно не изменяются.
Масса растительного опада достоверно выше (z = 2.92, p < 0.01), а запас биомассы мхов достоверно ниже (z = –2.27, p = 0.02) на приствольных участках деревьев Pinus sylvestris по сравнению с аналогичными микроместообитаниями под деревьями Betula pubescens. В подкроновом пространстве сравниваемых видов деревьев достоверно различается только запас подстилки: он достоверно больше (z = 2.10, p = 0.04) под кронами Pinus sylvestris.
В сосняке зеленомошном (ППП 3) в приствольном и подкроновом пространстве лишайники практически отсутствуют, в небольшом количестве они присутствуют лишь в окнах полога древостоя (табл. 2). Биомасса мхов, надземных и подземных частей кустарничков более равномерно распределена в пространстве рассматриваемого биогеоценоза, чем в лишайниковом редколесье и сосняке лишайниково-зеленомошном и статистически одинакова в разных типах микроместообитаний.
На ППП 3 масса растительного опада достоверно больше у стволов и под кронами сосен по сравнению с межкроновыми участками, а также в приствольной части деревьев березы по сравнению с подкроновым и межкроновым пространством. Более высокие значения толщины подстилки характерны для приствольного пространства деревьев сосны, по сравнению с подкроновым и межкроновым, а также для приствольных и подкроновых участков деревьев березы по сравнению с межкроновым пространством (табл. 3).
Сопоставление запасов органического вещества вблизи стволов и под кронами деревьев Pinus sylvestris и Betula pubescens показало, что только запас биомассы мхов на приствольных участках деревьев Pinus sylvestris достоверно больше (z = 2.40, p = 0.016), чем в аналогичных микроместообитаниях деревьев Betula pubescens.
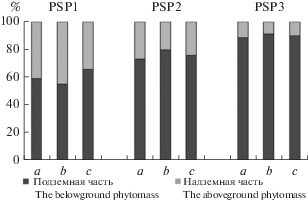
Анализ соотношения запасов биомассы надземных и подземных частей кустарничков в межкроновых пространствах, под кронами и в непосредственной близости от стволов Pinus sylvestris и Betula pubescens выявил сходный характер его изменения в зависимости от типа сообществ. На рис. 1 приведено соотношение указанных величин на примере Pinus sylvestris. Во всех случаях запас надземной биомассы кустарничков меньше, чем запас биомассы подземных частей, причем для каждого типа леса характерно свое соотношение, которое не зависит от типа микроместообитания: в сосновом редколесье (ППП 1) запас надземных частей кустарничков варьирует от 34 до 45%, в сосняке лишайниково-зеленомошном (ППП 2) – от 20 до 27%, в сосняке зеленомошном (ППП 3) – от 9 до 11% от общего запаса биомассы кустарничков.
Рис. 1.
Соотношение запасов биомассы надземных и подземных частей кустарничков в приствольном и подкроновом пространстве деревьев Pinus sylvestris и в межкроновых микроместообитаниях исследованных сосновых сообществ. Типы микроместообитаний (здесь и на рис. 2–4): a – приствольные, b – подкроновые, c – межкроновые. По вертикали: относительный запас биомассы, %; по горизонтали: тип микроместообитания. Fig. 1. Proportion of the stock of the above- and belowground shrub phytomass in microhabitats at tree base and under crown of Pinus sylvestris and in canopy gaps of the studied Scots pine communities. Microhabitat types (here and on Figs. 2–4): a – tree base; b – under crown; c – canopy gaps. X-axis: phytomass relative stock, %; y-axis: microhabitat type.
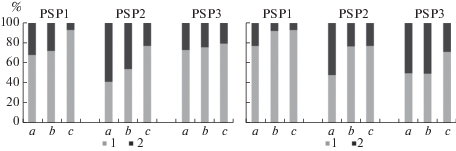
Сопоставление запасов надземной биомассы травяно-кустарничкового и мохово-лишайникового ярусов в межкроновых пространствах, под кронами и в непосредственной близости от стволов Pinus sylvestris и Betula pubescens показало, что в исследуемых сообществах основная часть органического вещества накапливается в мохово-лишайниковом ярусе (рис. 2). В приствольных микроместообитаниях запас надземной биомассы травяно-кустарничкового яруса варьирует от 23 до 59%, в подкроновых пространствах – от 8 до 50%. Наибольший вклад в общий запас надземной биомассы (45–60%) травяно-кустарничковый ярус вносит в приствольных и подкроновых микроместообитаниях деревьев сосны и в приствольном пространстве деревьев березы пушистой в сосняке лишайниково-зеленомошном (ППП 2), а также на приствольных и подкроновых участках деревьев березы пушистой в сосняке зеленомошном (ППП 3). Наиболее низкая доля растений травяно-кустарничкового яруса в общем запасе надземной биомассы (6–8%) наблюдается в межкроновом пространстве и под кронами березы в сосновом редколесье (ППП 1). Аналогичную закономерность преобладания вклада мохово-лишайникового яруса в общий запас надземной биомассы отмечали и другие исследователи при изучении микромозаики напочвенного покрова в сосновых лесах [11, 12]. Напротив, в темнохвойных лесах северной и средней тайги вклад сосудистых растений в общий запас надземной биомассы больше, чем вклад мхов [4, 5, 8, 11, 12].
Рис. 2.
Соотношение запасов биомассы мохово-лишайникового и травяно-кустарничкового ярусов в межкроновых, подкроновых и приствольных пространствах деревьев Pinus sylvestris (слева) и Betula pubescens (справа) в исследованных сосновых сообществах. 1 – травяно-кустарничковый ярус; 2 – мохово-лишайниковый ярус. По вертикали: относительный запас биомассы, %; по горизонтали: тип микроместообитания. Fig. 2. Proportion of the stock of lichen-moss and grass- dwarf shrub layers in canopy gaps, under crown and at tree base areas of Pinus sylvestris (left) and Betula pubescens (right) of the studied Scots pine communities. 1 – grass-dwarf shrub layer; 2 – lichen-moss layer. X-axis: phytomass relative stock, %; y-axis: microhabitat type.
Поскольку достоверные различия общего запаса надземной биомассы и общего запаса живого напочвенного покрова (с учетом подземных частей) в приствольных и подкроновых микроместообитаниях деревьев Pinus sylvestris и Betula pubescens отсутствуют, были проанализированы две соответствующие объединенные выборки. На рис. 3 и 4 представлены данные об общем запасе надземной биомассы и запасе живого напочвенного покрова в различных микроместообитаниях исследованных сообществ без учета видовой принадлежности деревьев. В сосновом редколесье (ППП 1) общий запас надземной биомассы (Н = 17.4, р < 0.001) и запас живого напочвенного покрова (Н = 13.0, р = 0.002) достоверно увеличиваются от приствольных к межкроновым микроместообитаниям. В сосняке лишайниково-зеленомошном (ППП 2) общий запас надземной биомассы (Н = 13.4, р < 0.001) и запас живого напочвенного покрова (Н = 8.1, p = 0.02) достоверно меньше в приствольных микроместообитаниях по сравнению с подкроновым и межкроновым пространством. В сосняке зеленомошном (ППП 3) различия этих показателей в разных типах микроместообитаний отсутствуют (рис. 3, 4).
Рис. 3.
Общий запас надземной биомассы напочвенного покрова в различных микроместообитаниях исследованных сосновых сообществ. По вертикали: общий запас надземной биомассы, г/м2; по горизонтали: номера пробных площадей. Fig. 3. Total stock of the aboveground phytomass in different microhabitats of the studied Scots pine communities. X-axis: total stock of the aboveground phytomass, g/m2; y-axis: sample plots.

Рис. 4.
Общий запас биомассы живого напочвенного покрова (с учетом подземных частей) в различных микроместообитаниях исследованных сосновых сообществ. По вертикали: общий запас биомассы живого напочвенного покрова, г/м2; по горизонтали: номера пробных площадей. Fig. 4. Total stock of the live ground cover phytomass (including below-ground parts) in different microhabitats of the studied Scots pine communities. X-axis: total stock of the live ground cover, g/m2; y-axis: sample plots.
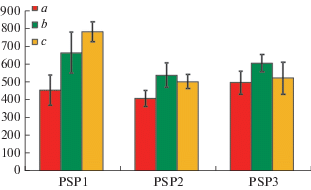
Интересно отметить, что в исследованных сосновых лесах характер изменения общего запаса надземной биомассы и общего запаса живого напочвенного покрова от приствольных участков к межкроновым пространствам различается в зависимости от типовой принадлежности сообществ. Среднее значение общего запаса надземной биомассы снижается от 504 до 157 г/м2, т.е. в 3.2 раза, в ряду сосновое редколесье –> ‒> сосняк лишайниково-зеленомошный –> сосняк зеленомошный, в то время как общий запас живого напочвенного покрова достоверно не различается в этом ряду, варьируя в пределах 490–642 г/м2. Такие различия обусловлены тем, что в сосновом редколесье запас надземной биомассы преимущественно слагается из биомассы лишайников, наиболее обильных в этом сообществе, а в сосняке зеленомошном в общей биомассе существенно увеличивается по сравнению с другими сообществами доля подземных частей растений, что приводит к выравниванию общих запасов живого напочвенного покрова в исследованных лесах и редколесье.
Хорошо известно, что на Кольском полуострове лишайниковые сосновые редколесья формируются на песчаных отложениях в наиболее сухих местообитаниях. Для лишайниково-зеленомошных и особенно зеленомошных сосновых лесов характерна более высокая степень почвенного увлажнения. Проведенные исследования выявили вполне логичные закономерности изменения запасов, как надземной биомассы компонентов напочвенного покрова, так и подземной биомассы кустарничков в ряду от лишайниковых редколесий до сосняков зеленомошных. Средние значения запаса биомассы лишайников снижаются более чем в 30 раз, а мхов, наоборот, увеличиваются более чем в 50 раз, в то время как различия в запасе надземной биомассы кустарничков составляют около 3 крат, а в запасе биомассы подземных органов растений не превышают 2.4 крат. Следует подчеркнуть, что во всех исследуемых типах сосновых сообществ запас биомассы подземных частей кустарничков превышает запас надземной биомассы кустарничков, причем эти различия увеличиваются от 1.1–1.4 раза в сосновом редколесье до 7–11 раз в сосняке зеленомошном.
В результате сравнительной оценки запасов биомассы компонентов напочвенного покрова в различных микроместообитаниях установлено существенное внутриэкосистемное разнообразие этого параметра и его закономерное изменение в зависимости от местоположения относительно стволов лесообразующих видов. Во всех исследуемых типах сосновых сообществ прослеживается общий тренд увеличения биомассы лишайников от приствольных участков деревьев Pinus sylvestris и Betula pubescens к межкроновым микроместообитаниям, но статистически эту тенденцию удалось подтвердить не во всех случаях.
Практически полное отсутствие мхов в сосновом редколесье обусловлено высокой сухостью данного экотопа, только в более влажных приствольных и подкроновых микроместообитаниях Betula pubescens поселяются зеленые мхи и формируется небольшой запас их биомассы. В более влажном сосняке лишайниково-зеленомошном запас биомассы мхов значительно выше и возрастает (в 2.5–5.3 раза) от приствольных микроместообитаний к межкроновым. Это обусловлено более высокой сухостью приствольных участков по сравнению с межкроновыми в связи с перехватом атмосферных осадков кронами Pinus sylvestris и Betula pubescens. В наиболее влажном экотопе сосняка зеленомошного наблюдаются максимальные значения запаса биомассы мхов, при этом различия между подкроновыми и межкроновыми микроместообитаниями отсутствуют, что связано с достаточной влагообеспеченностью экотопа и высокой общей затененностью нижних ярусов сообществ кронами высокополнотного древостоя. Такие условия благоприятствуют произрастанию зеленых мхов.
Закономерности изменения запасов надземной и подземной биомассы кустарничков по микроместообитаниям в исследуемых сосновых сообществах имеют различия. Во всех сообществах запас как надземной, так и подземной биомассы кустарничков в подкроновых микроместообитаниях Betula pubescens достоверно не отличается от запаса в межкроновых пространствах. В то же время, на приствольных участках и под кронами Pinus sylvestris в сосновых редколесьях и сосняках лишайниково-зеленомошных запас биомассы кустарничков достоверно больше, чем в межкроновых микроместообитаниях.
Проведенные исследования выявили закономерности изменения запасов лесной подстилки и массы опада в разных типах средневозрастных сосновых сообществ в зависимости от типа микроместообитаний. По градиенту почвенного увлажнения в ряду лишайниковое редколесье → лишайниково-зеленомошный сосняк → зеленомошный сосняк средние величины толщины и запаса лесной подстилки увеличиваются соответственно в 2 и 2.5 раза, а масса опада лишь в 1.3 раза. Согласно данным В.В. Горшкова и др. [30], стационарные значения толщины подстилки в северотаежных лишайниковых, лишайниково-зеленомошных и зеленомошных сосновых лесах составляют соответственно 2.5 ± 0.2, 5.0 ± 0.8 и 7.5 ± 0.6 см. В наших исследованиях наименьшая толщина подстилки наблюдается в лишайниковом редколесье, где средняя ее величина составляет 2.8 ± 0.4, а в двух других типах леса – соответственно 4.4 ± 0.3 и 5.8 ± 0.5 см. Более низкие по сравнению со стационарными значения толщины подстилки в сосняках лишайниково-зеленомошном и зеленомошном обусловлены давностью пожара, которая в наших исследованиях была равна 90 годам, а восстановление толщины подстилки до стационарных величин в этих типах сообществ происходит позже – через 120–190 лет [30]. Отметим, что высокая вариабельность толщины и запаса лесной подстилки отмечена в борах Марийского Заволжья, среднее значение толщины подстилки составляет 3.7 ± 0.3 см, варьируя от 1 до 15 см [13].
Согласно литературным данным, в лишайниковых сосновых лесах Европейского севера масса ежегодного опада в среднем составляет 1.5–2.0 т/га, в зеленомошных – 4.0–4.5 т/га [11, 31–33]. Поскольку зеленомошные сосновые леса распространены в местообитаниях с более высоким уровнем увлажнения по сравнению с лишайниковыми, формирующиеся в этих условиях моховой покров и, значительный по толщине, обладающий высокой порозностью и влагоемкостью, слой опада, характеризуются более высокими водоудерживающими и теплоизолирующими свойствами по сравнению с лишайниковым покровом и слоем опада в лишайниковых сообществах [34–36]. Из этого следует, что в зеленомошных сосновых лесах в условиях не только более высокой влажности, но и более низких температур по сравнению с лишайниковыми, процесс разложения растительных остатков происходит более медленно. Кроме того, сообщества зеленомошного типа характеризуются более высоким содержанием в опаде зольных элементов и азота [11, 31, 37–39]. По мнению ряда авторов [40, 41], минерализация органического вещества почвы – главный источник азота в лесных экосистемах России, поскольку низок уровень, как азотных выпадений, так и биологической фиксации азота. Снижение пула органического вещества в почве компенсируется растительным опадом и отпадом деревьев [41]. Кроме того, как уже отмечалось выше, опад хвои и растений мохового покрова замедляют разложение подстилки и затрудняют образование гумуса, а опад листьев березы, напротив, благоприятствует этому процессу. Таким образом, перечисленные факторы создают комплекс условий, с одной стороны, ускоряющих процесс разложения растительных остатков, с другой – тормозящих их полную минерализацию.
Логично было предположить, что в приствольных микроместообитаниях будет накапливаться максимальное количество растительного опада за счет высокой доли древесных фракций (веточек, коры) и шишек, и полученные данные подтвердили это положение. Во всех исследуемых типах сосновых лесов максимальные значения массы растительного опада наблюдаются в приствольных микроместообитаниях, а минимальные – в межкроновых пространствах, где она в среднем в 2 раза меньше, что согласуется с данными других исследователей [7].
Тренд примерно 2-кратного уменьшения толщины лесной подстилки от приствольных участков деревьев сосны и березы к межкроновым микроместообитаниям обнаруживается во всех исследованных типах сосновых сообществ, что в значительной степени связано с уменьшением массы растительного опада, из которого и формируется лесная подстилка. Однако достоверное уменьшение запаса лесной подстилки от приствольных микроместообитаний деревьев Pinus sylvestris к межкроновым наблюдается только в лишайниковом сосновом редколесье; в двух других типах сообществ пространственное распределение лесной подстилки является более равномерным и ее запас в разных микроместообитаниях достоверно не различается. Кроме того, во всех исследованных сосновых сообществах и во всех типах микроместообитаний отсутствует корреляция между толщиной и запасом лесной подстилки, что, возможно, объясняется большим интервалом варьирования значений обоих параметров. Высокую степень внутриценотического варьирования толщины и массы лесной подстилки в лесных сообществах разной типологической принадлежности отмечают и другие исследователи [1–4, 7, 8, 12, 13].
ЗАКЛЮЧЕНИЕ
Проведенные исследования позволили оценить различия в запасе биомассы растений травяно-кустарничкового и мохово-лишайникового ярусов, а также запасов растительного опада и лесной подстилки на приствольных и подкроновых участках деревьев Pinus sylvestris и Betula pubescens и в межкроновом пространстве в средневозрастных северотаежных сосновых сообществах разной типологической принадлежности.
Сравнительный анализ запасов биомассы компонентов напочвенного покрова (кустарничков, мхов и лишайников) в различных микроместообитаниях выявил существенное внутриэкосистемное разнообразие этих параметров. В лишайниково-зеленомошном и зеленомошном сосновых лесах и сосновом лишайниковом редколесье значимое увеличение биомассы лишайников от приствольных к межкроновым микроместообитаниям составляет от 2 до 15 крат. Запас биомассы зеленых мхов в сосняке лишайниково-зеленомошном возрастает в 2.5–5.3 раза при переходе от приствольных (28–59 г/м2) к межкроновым микроместообитаниям. В более влажном экотопе сосняка зеленомошного наблюдаются более высокие значения запаса биомассы мхов (60–115 г/м2), при этом различия между разными типами микроместообитаний отсутствуют.
Запасы биомассы надземных и подземных частей кустарничков в лишайниковом редколесье в приствольном и подкроновом пространствах деревьев Pinus sylvestris примерно в 2 раза выше, чем в межкроновом. В сосняке лишайниково-зеленомошном эти различия выражены гораздо слабее, а в сосняке зеленомошном запасы биомассы надземных и подземных частей кустарничков в разных типах микроместообитаний одинаковы. Деревья Betula pubescens, согласно полученным данным, не оказывают влияния на пространственное распределение биомассы травяно-кустарничкового яруса.
Общие запасы надземной биомассы и всего живого напочвенного покрова (с учетом подземных частей) в лишайниковом сосновом редколесье и сосняке лишайниково-зеленомошном достоверно увеличиваются от приствольных к межкроновым микроместообитаниям, в то время как в сосняке зеленомошном различия отсутствуют.
Масса растительного опада имеет незначительные различия в однотипных микроместообитаниях во всех растительных сообществах и сходный характер пространственного распределения: на приствольных и подкроновых участках она выше, чем в межкроновых пространствах. Во всех исследуемых типах сообществ наиболее равномерно по площади биогеоценоза распределен запас лесной подстилки. В отличие от запаса, толщина подстилки в большинстве случаев существенно (в ~2–4 раза) уменьшается от приствольных к межкроновым микроместообитаниям.
В целом можно констатировать, что наиболее неоднородное распределение запасов органического вещества по площади фитоценоза наблюдается в сосновом лишайниковом редколесье, а наиболее равномерное – в сосняке зеленомошном. Можно предположить, что различия, выявленные в редколесье, отличающемся значительной разреженностью древостоя, обусловлены большей контрастностью гидротермического режима в разных типах микроместообитаний по сравнению с исследуемыми лесами, особенно густыми и высокополнотными сосняками зеленомошными. В заключение отметим, что высокая степень внутриценотического варьирования исследуемых параметров требует продолжения исследований для более детального и доказательного выявления закономерностей формирования микромозаики почвенно-растительного покрова в северотаежных сосновых лесах.
Список литературы
Орлова М.А., Лукина Н.В., Камаев И.О., Смирнов В.Э., Кравченко Т.В. 2011. Мозаичность лесных биогеоценозов и продуктивность почв. – Лесоведение. 6: 39–48.
Орлова М.А., Лукина Н.В., Смирнов В.Э., Исаева Л.Г. 2014. Плодородие почв березовых лесов на северном пределе их распространения. – Почвоведение. 3: 327–339.
Орлова М.А., Лукина Н.В., Смирнов В.Э., Артемкина Н.А. 2016. Влияние ели на кислотность и содержании элементов питания в почвах северотаежных ельников кустарничково-зеленомошных – Почвоведение. 11: 1355–1367.
Смирнова О.В., Алейников А.А., Семиколенных А.А., Бовкунов А.Д., Запрудина М.В., Смирнов Н.С. 2011. Пространственная неоднородность почвенно-растительного покрова темнохвойных лесов в Печоро-Илычском заповеднике – Лесоведение. 6:67–78.
Луговая Д.Л., Смирнова О.В., Запрудина М.В., Алейников А.А., Смирнов В.Э. 2013. Микромозаичная организация и фитомасса напочвенного покрова в основных типах темнохвойных лесов Печоро-Илычского заповедника – Экология. 1: 3–10.
Лебедева В.Х., Ипатов В.С., Тиходеева М.Ю. 2015. Неоднородность пространственной структуры живого напочвенного покрова в лесных сообществах – Вестник СПбГУ. Сер. 3. Биология. 2: 32–46. https://biocomm.spbu.ru/article/view/859/750
Иванова Е.А., Лукина Н.В. 2017. Варьирование массы и фракционного состава древесного опада в сосняках кустарничково-лишайниковых при аэротехногенном загрязнении – Лесоведение. 5: 47–58. http://lesovedenie.ru/index.php/forestry/article/view/1049
Артемкина Н.А., Орлова М.А., Лукина Н.В. 2018. Микромозаика растительности и вариабельность химического состава L-горизонтов подстилки северотаежных ельников кустарничково-зеленомошных – Лесоведение. 2: 97–106. http://lesovedenie.ru/index.php/forestry/article/view/1091
Лукина Н.В., Ершов В.В., Горбачева Т.В., Орлова М.А., Исаева Л.Г., Тебенькова Д.Н. 2018. Оценка состава почвенных вод северотаежных хвойных лесов фоновых территорий индустриально развитого региона – Почвоведение. 3: 284–296. https://doi.org/10.7868/S0032180X18030036
Данилова М.А., Лукина Н.В., Кузнецова А.И., Смирнов В.Э. 2019. Влияние древесных растений на плодородие почв таежных лесов. В сб.: Экологические проблемы северных регионов и пути их решения: Тез. докл. VII Всеросс. научн. конф. с международ. участием. Апатиты. С. 18–19. http://inep.ksc.ru/documents/11_ecol_prob_19.pdf
Никонов В.В., Лукина Н.В. 1994. Биогеохимические функции лесов на северном пределе распространения. Апатиты. 315 с.
Никонов В.В., Лукина Н.В., Смирнова Е.В., Исаева Л.Г. 2002. Влияние Picea obovata и Pinus sylvestris на первичную продуктивность нижних ярусов хвойных лесов Кольского полуострова – Ботан. журн. 87(8):107–119.
Демаков Ю.П., Исаев А.В., Шарафутдинов Р.Н. 2017. Роль лесной подстилки в борах Марийского Заволжья и вариабельность ее параметров. В сб.: Научные Труды Государственного природного заповедника “Большая Кокшага”. Вып. 8. С. 15–43.
Иванова Е.А., Артемкина Н.А., Лукина Н.В. 2019. Разложение опада в сосновых лесах на северном пределе распространения на Кольском полуострове. В сб.: Экологические проблемы северных регионов и пути их решения: Тез. докл. VII Всеросс. научн. конф. с международ. участием. Апатиты. С. 30–32. http://inep.ksc.ru/documents/11_ecol_prob_19.pdf
Дылис Н.В. Структура лесного биогеоценоза. 1969. 28 с. https://www.booksite.ru/fulltext/rusles/dylis/text.pdf
Ипатов В.С., Журавлева Е.Н., Лебедева В.Х., Тиходеева М.Ю. 2009. Фитогенное поле Picea abies, P. obovata (Pinaceae). – Ботан. журн. 94(4): 558–568.
Журавлева Е.Н., Ипатов В.С., Лебедева В.Х., Тиходеева М.Ю. 2012. Изменение растительности на лугах под влиянием сосны обыкновенной (Pinus sylvestris L.) – Вестник Санкт-Петербургского университета. Сер. Биология. 2: 3–12. https://biocomm.spbu.ru/article/view/3842/3439
Tedersco L., Suvi T., Jairus T., Kôljalg U. 2008. Forest mircosite effects on community composition of ectomycorrhizal fungi on seedlings of Picea abies and Betula pendula – Environ. Microbiol. 10(5): 1189–1201. https://doi.org/10.1111/j.1462-2920.2007.01535.x
Sass E.M., D’Amato A.W., Foster D.R., Plotkin A.B., Fraver Sh., Schoonmaker P.K., Orwig D.A. 2018. Long-term influence of disturbance-generated microsites on forest structural and compositional development. – Can. J. of Forest Research. 48(8): 958–965. https://doi.org/10.1139/cjfr-2018-0097
Лукина Н.В., Орлова М.А., Исаева Л.Г. 2010. Плодородие лесных почв как основа взаимосвязи почва-растительность – Лесоведение. 5: 45–56.
Орлова М.А., Лукина Н.В., Смирнов В.Э., Краснов Д.А., Камаев И.О. 2012. Плодородие почв еловых лесов Хибинских гор – Почвоведение. 6:682–694.
Орлова М.А. 2013. Элементарная единица лесного биогеоценотического покрова для оценки экосистемных функций лесов – Труды Карельского научного центра. Серия Экологические исследования. 6: 126–132. http://resources.krc.karelia.ru/transactions/doc/trudy2013/trudy_2013_6_126-132.pdf
Сукцессионные процессы в заповедниках России и проблемы сохранения биологического разнообразия. 1999. СПб. 549 с.
Lang S.I., Cornelissen J.H.C., Klahn T., Van Logtestijn R.S.P., Broekman R., Schweikert W., Aerts R. 2009. An experimental comparison of chemical traits and litter decomposition rates in a diverse range of subarctic bryophyte, lichen and vascular plant species – Journal of Ecology. 97(5): 886–900. https://doi.org/10.1111/j.1365-2745.2009.01538.x
Berg B., Albrektson A., Berg M.P., Cortina J., Johansson M-B., Gallardo A., Mgadeira M., Pausas J., Kiratz W., Vallejo R., McClaugherty C. 1999. Amounts of litter fall in some pine forests in European transect, in particular Scots pine – Annals of Forest Science. 56: 625–639. https://doi.org/10.1051/forest:19990801
Формирование лесной подстилки. http://geolike.ru/page/gl_2090.htm.
Федорец Н.Г., Бахмет О.Н. 2003. Экологические особенности трансформации соединений углерода и азота в лесных почвах. Петрозаводск. 240 с.
Noreika N., Helm A., Opik M., Jairus T., Vasar M., Reier U., Kook E., Riibak K., Kasari L., Tullus H., Tullus T., Lutter R., Oja E., Saag A., Randlane T., Partel M. 2019. Forest biomass, soil and biodiversity relationships originate from biogeographic affinity and direct ecological effects. – Oikos. 00:1–13. https://doi.org/10.1111/oik.06693
Мировая реферативная база почвенных ресурсов 2014, исправленная и дополненная версия 2015. Международная система почвенной классификации для диагностики почв и создания легенд почвенных карт. Доклады о мировых почвенных ресурсах №106. ФАО, Рим. 203 с. http://www.fao.org/3/i3794ru/I3794RU.pdf
Горшков В.В., Ставрова Н.И., Баккал И.Ю. 2005. Динамика восстановления лесной подстилки в бореальных сосновых лесах после пожаров – Лесоведение. 3: 37–45.
Манаков К.Н. 1978. Продуктивность и биологический круговорот в сосновых лесах. В кн.: Биологическая продуктивность и обмен в лесных биогеоценозах Кольского полуострова. Апатиты. С. 3–18.
Молчанов А.А. 1974. Продуктивность органической и биологической массы леса. М. 250 с.
Чепурко Н.Л. 1971. Структура и годовой баланс биомассы в лесах Хибинского горного массива. В кн.: Почвы и продуктивность растительных сообществ. М. С. 94–116.
Семко А.П. 1982. Гидротермический режим почв лесной зоны Кольского полуострова. Апатиты. 142 с.
Ярмишко В.Т., Цветков В.Ф. 1987. Строение, запасы и распределение в почве корневых систем растений в сообществах сосновых молодняков Кольского полуострова – Ботан. журн. 72(4):496–505.
Ипатов B.C., Трофимец В.И. 1988. Влияние лишайниковых и зеленомошных ковров на водный режим верхнего корнеобитаемого слоя почвы в сухих сосняках – Экология. 1: 19–23.
Никонов В.В., Манаков К.Н. 1979. Экологические и биогеохимические особенности северотаежных лесов Кольского полуострова – Экология. 5:33–38.
Bormann B.T., Sidle R.C. 1990. Changes in productivity and distribution of nutrients in a chronosequence at Glacier Bay National Park, Alaska. – Journal of Ecology. 78(3): 561–578. https://www.jstor.org/stable/2260884
Cleve K., Viereck L.A. 1981. Forests succession in relation to nutrient cycling in the boreal forest of Alaska. In: Forest succession. Concepts and application. Berlin. P. 185–211. https://www.springer.com/gp/book/9780387905976
Разгулин С.М. 2008. Минерализация азота в почвах бореальных лесов – Лесоведение. 4: 57–62.
Меняйло О.В., Матвиенко А.И., Макаров М.И., Ченг Ш.-К. 2018. Роль азота в регуляции цикла углерода в лесных экосистемах. – Лесоведение. 2: 143–159. http://lesovedenie.ru/index.php/forestry/article/view/1095
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы