

Растительные ресурсы, 2020, T. 56, № 2, стр. 151-164
Особенности структуры сообществ экотонного комплекса ельник черничный–осинник злаково-разнотравный в условиях Республики Карелия
Н. В. Геникова 1, *, В. А. Харитонов 1, А. Н. Пеккоев 1, А. Ю. Карпечко 1, А. В. Кикеева 1, А. М. Крышень 1, Р. П. Обабко 1
1 Институт леса – обособленное подразделение Федерального государственного бюджетного учреждения науки Федерального исследовательского центра “Карельский научный центр Российской академии наук”
г. Петрозаводск, Россия
* E-mail: genikova@krc.karelia.ru
Поступила в редакцию 28.01.2020
После доработки 27.02.2020
Принята к публикации 18.03.2020
Аннотация
Приведены результаты исследования экотонного комплекса 90-летнего ельника черничного и 35-летнего осинника злаково-разнотравного, появившегося на месте сплошной рубки. Выявлено, что после 35 лет естественного восстановления леса на вырубке по ряду признаков определяется переходная зона, распространяющаяся примерно на 10 м в обе стороны от границы примыкания сообществ. Максимальные размеры переходной зоны – 10 м от границы сообществ – определены по изменению радиального прироста, который был в среднем выше, чем в центральной части ельника черничного в течение двух десятилетий после рубки. В переходной зоне, в отличие от других зон экотонного комплекса, отмечается максимальная густота подроста ели. Минимальная протяженность куртин Hylocomium splendens (Hedw.) Bruch et al. на стволах осины в ельнике черничном отмечается у границы с осинником, максимальная – на самом удаленном вглубь ельника дереве. Насыщенность еловыми корнями верхнего слоя почвы снижается в направлении от ельника черничного к осиннику злаково-разнотравному. Масса корней лиственных пород в среднем не различается по зонам экотонного комплекса и соответствует запасу древесины лиственных в составе древостоев. Через 35 лет после рубки древостоя переходная зона по видовому составу и по соотношению функциональных групп сосудистых растений обладает большим сходством с примыкающим к ней ельником черничным, чем с осинником злаково-разногравным. Однако в переходной зоне отмечается повышение проективного покрытия и средней высоты лесных кустарничков (черники и брусники) по сравнению с другими зонами экотонного комплекса.
Лесное законодательство накладывает ограничения на размеры лесосек [1]. В таежной зоне в эксплуатационных лесах предельная площадь сплошных рубок составляет 50, а чересполосно-постепенных – 30 га. Чересполосно-постепенные рубки, проводимые в защитных лесах, не должны превышать 15 га. Это создает особую мозаику лесных сообществ различного возраста с переходными зонами, характеристики которых отличаются от характеристик как лесного сообщества, так и вырубки [2–4]. В частности, известны работы, в которых подтверждено, что условия краевых участков леса способствуют развитию такого ресурсного вида как черника [5].
Пространственное размещение деревьев, их дифференциация по высоте и диаметру, а также процессы естественного отпада во многом определяются конкуренцией растений за световые и почвенные ресурсы [6, 7]. После проведения сплошной рубки происходит резкое изменение условий среды, вызванное увеличением интенсивности поступления света [8, 9]. Рубка приводит также к изменению роста корневых систем, к ослаблению конкуренции [10], что отражается на деревьях, произрастающих на границе с вырубкой. При изучении экотонных комплексов лес–вырубка объектами исследования преимущественно являются растительные сообщества на ранних стадиях восстановления [4, 11, 12], а предметом исследования – отдельные компоненты биогеоценоза [2, 13]. Сохраняются ли выделенные зоны экотонного комплекса при формировании древесного яруса на вырубке и если да, то как они проявляются? Ответы на эти вопросы могут быть получены только в комплексных исследованиях древесного яруса, напочвенного покрова, почвы.
Ранее нами был исследован 10-летний период изменений в сообществах экотонного комплекса ельник черничный–вырубка [14–16]. Цель настоящей работы – комплексная оценка влияния условий в зоне контакта ельника черничного и 35-летнего осинника злаково-разнотравного на радиальный прирост ели, количество и высоту подроста древесных растений, моховой эпифитный покров, видовой состав и проективное покрытие напочвенного покрова, высоту лесных кустарничков, корненасыщенность и микоризообразование.
МАТЕРИАЛ И МЕТОДЫ
В Прионежском районе Республики Карелия (подзона средней тайги) в условиях ельника черничного были проведены исследования влияния рубки на лесное сообщество. Опытный участок состоял из двух пробных площадей (ПП), заложенных в различных по составу и возрасту насаждениях: исходного ельника черничного и примыкающего к нему осинника злаково-разнотравного, возникшего на вырубке 35-летней давности. Координаты опытного участка: N 61.860975°, E 33.918453°. Размер пробной площади в спелом ельнике черничном – 30 × 30 м (0.09 га). В лиственном сообществе для описания сформировавшегося насаждения заложены учетные ленты 25 × 2 м.
В ельнике черничном (ПП №1) проведено картирование и сплошной перечет деревьев с измерением их высот и диаметров на высоте 1.3 м, выполнен учет подроста. Таксационная характеристика ельника и примыкающего к нему лиственного сообщества (ПП № 2), представлены в табл. 1.
Таблица 1.
Таксационная характеристика древостоев на пробных площадях Table 1. Taxational characteristics of stands on the sample plots
| № ПП Sample plot |
Возраст, лет Age, years |
Состав Composition | Густота, тыс.экз./га Density, 103ind. /ha |
Средние Average |
Полнота Density of stock |
Запас, м3/га Volume, m3/ha |
Класс бонитета Productivity class |
|
|---|---|---|---|---|---|---|---|---|
| диаметр, см diameter, cm |
высота, м height, m |
|||||||
| 1 | 90 | 6.7Pic | 1.00 | 16.7 | 17.2 | 1.2 | 376 | IV |
| 1.7B | 0.20 | 18.4 | 20.0 | |||||
| 1.6Pop | 0.09 | 30.4 | 23.7 | |||||
| ед.Pin | 0.02 | 36.8 | 23.8 | |||||
| 2 | 35 | 5.7Pop | 1.56 | 10.3 | 14.9 | 1.3 | 162 | IV |
| 2.8B | 1.20 | 5.7 | 8.4 | |||||
| 1.5Pic | 3.20 | 2.0 | 2.7 | |||||
Исходное насаждение – спелый, высокополнотный ельник с долей участия ели Picea abies (L.) H. Karst. в составе древостоя – 6.7 единицы, березы Betula sp. – 1.7 единицы и осины Populus tremula L. – 1.6 единицы (соответственно 251, 62 и 61 м3/га). В небольшом количестве встречается сосна Pinus sylvestris L. Количество елового подроста составляет 4.0, осинового – 1.0 тыс. шт./га. В подлеске представлена рябина Sorbus aucuparia L., имеющая густоту 1.1 тыс. шт./га. Напочвенный покров довольно хорошо развит, общее проективное покрытие травяно-кустарничкового яруса составляет 18%, в его составе преобладают черника Vaccinium myrtillus L., брусника V. vitis-idaea L., кислица Oxalis acetosella L. Покрытие мохового яруса, сформированного зелеными мхами, составляет 12%.
Осинник разнотравный, имеющий возраст 35 лет, с участием березы и культурами ели, возник после рубки в 1983 году. Подрост в осиннике представлен хвойными породами – елью (3.0 тыс. шт./га), сосной (0.07 тыс. шт./га) и лиственными – березой (0.7 тыс. шт./га) и осиной (0.5 тыс. шт./га). В подлеске рябина – 1.1 тыс. шт./га. Общее проективное покрытие травяно-кустарничкового яруса в среднем составляет 15%. Доминантами являются щитовник картузианский Dryopteris carthusiana (Vill.) H.P. Fuchs, вейник лесной Calamagrostis arundinacea (L.) Roth, черника и политрихум обыкновенный Polytrichum commune Hedw. Моховой покров выражен слабо, его общее покрытие составляет 6%.
Известно, что радиальный прирост может служить одним из количественных показателей конкурентных отношений, о чем свидетельствует его довольно тесная связь с различными индексами конкуренции [17–19]. Этот показатель, на наш взгляд, может также наглядно характеризовать продолжительность и силу влияния “краевого эффекта” после рубки древостоя в результате изменения светового режима. Поэтому одной из задач исследования было выявление наличия у ели реакции на сплошную рубку прилегающего участка, которая выражается в изменении радиального прироста, а также определить расстояние, на котором проявляется данная реакция, и продолжительность периода изменений. Для этого в спелом ельнике у модельных деревьев ели, расположенных на разном расстоянии от границы осинника, отбирались керны древесины на высоте 1.3 м для определения радиального прироста (всего 13 кернов). С целью уменьшения влияния конкуренции окружающих деревьев на ход роста по диаметру модельные деревья подбирались таким образом, чтобы минимальное расстояние до соседних деревьев составляло не менее 1.5 м. По кернам в лабораторных условиях, с точностью 0.01 мм, определялась ширина годичных слоев. Полученные данные радиального прироста группировались по 5-летиям и сравнивались между собой.
Эпифитный моховой покров регистрировался на каждом дереве осины, произрастающем в пределах ПП в ельнике черничном. Для каждого дерева была проведена оценка эпифитного покрова глазомерным методом, с четырех сторон света, от основания дерева до верхней границы встречаемости эпифитных растений. В данном исследовании в качестве индикаторного вида для оценки того, насколько удаление форофита от границы контакта двух лесных сообществ влияет на эпифитный покров, был использован Hylocomium splendens (Hedw.) Bruch et al. Данный вид характеризуется тем, что в основном произрастает на почве, но может расти и в нижней части стволов. Куртины мха с увеличением степени и продолжительности увлажнения способны разрастаться, поднимаясь на более значительную высоту по стволу. Этот показатель и был использован в анализе.
90-летний ельник черничный, 35-летний осинник злаково-разнотравный, сформировавшийся после рубки, и переходная зона между ними составляют единый экотонный комплекс, в котором были заложены три трансекты шириной 0.5 м и длиной 50 м (по 25 м в обе стороны от границы примыкания ельника и осинника). Ширина каждой зоны экотонного комплекса составляла 16 м (с учетом 1 м промежутков между зонами). Это обусловлено тем, что согласно нашим предыдущим исследованиям, ширина переходной зоны между двумя сообществами (ельником черничным и вырубкой 2–10 лет) составляет примерно 8 м от границы в обе стороны [14, 16].
Трансекты были разделены на учетные площадки размером 50 × 50 см, на которых определялись встречаемость (в процентах от общего числа учетных площадок) и проективное покрытие видов растений мохово-лишайникового и травяно-кустарничкового ярусов. На каждой учетной площадке была определена средняя высота лесных кустарничков (черники и брусники). Структуру доминирования видов сосудистых растений в напочвенном покрове оценивали по величине индекса выравненности Пиелу, принимающего значения от 0 до 1, где 1 означает равное участие всех видов [20].
Вдоль всех трех трансект проводился учет подроста на лентах шириной по 2 метра с каждой стороны трансекты. Общая площадь учета подроста составила 600 квадратных метров. При учете измерялась высота подроста с последующим распределением его на категории: мелкий (высота до 0.5 м), средний (от 0.5 до 1.5 м) и крупный (свыше 1.5 м).
С помощью люксметра вдоль трансекты с интервалом в 1 метр была измерена освещенность на высоте 0.5 м и 1.3 м над поверхностью земли.
Кроме того, по обеим сторонам одной из трансект на разном удалении от края спелого ельника для определения корненасыщенности были взяты образцы почвенных монолитов, размер которых составлял 10 × 10 × 20 см. Из монолита отбирались корни древесных пород диаметром до 3 мм, которые высушивались до абсолютно сухого состояния и взвешивались. Состояние тонких сосущих корней ели оценивалось с помощью измерения морфолого-анатомических параметров эктомикориз. Плотность эктомикориз оценивали в 5-кратной повторности на каждом участке. Поперечные срезы фиксированного материала эктомикориз толщиной 10–15 мкм изучали без окрашивания при помощи микроскопа. Определяли наличие грибного чехла, его толщину, на основе полученных данных рассчитывали долю чехла в объеме микоризного окончания [21].
Достоверность различий выборочных средних оценивалась на основе дисперсионного анализа с использованием непараметрического критерия Краскелла–Уоллиса.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
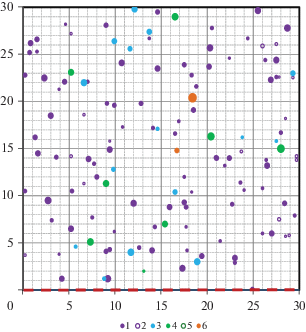
В ельнике черничном (ПП № 1) горизонтальная структура древостоя довольно однородна по составу (рис. 1). Анализ встречаемости с использованием критерия хи-квадрат показал, что расположение деревьев разных пород по пробной площади не зависело от их удаленности от края леса. В целом, обследованное насаждение типично для лесов таежной зоны, образовавшихся после вырубки продуктивных хвойных древостоев, имеющих достаточное количество подроста [22, 23].
Рис. 1.
Горизонтальная структура древостоя на ПП № 1 (ельник черничный). Пробная площадь разделена на квадраты 5 × 5 м2. 1 – ель живая, 2 – ель сухая, 3 – береза живая, 4 – осина живая, 5 – осина сухая, 6 – сосна живая. Красной пунктирной линией обозначена граница примыкания к осиннику. Fig. 1. The horizontal structure of the tree stand on the sample plot 1 (bilberry Norway spruce forest Vaccinium myrtillus–Picea abies). The sample plot is divided into quadrats of 5 × 5 m2. 1 – live Norway spruce trees, 2 – dead Norway spruce trees, 3 – live birch trees, 4 – live aspen trees, 5 – dead aspen trees, 6 – live Scots pine trees. Red dotted line shows the border with the aspen forest.
Возобновление ели. Значительные различия условий в зонах изученного экотонного комплекса оказали существенное влияние как на появление самосева ели, так и на дальнейшее развитие елового подроста. Наибольшее количество хвойного подроста в спелом ельнике наблюдается в 5‑метровой полосе от границы контакта двух сообществ, а в лиственном сообществе на расстоянии 10 м. Далее указанных границ общее количество подроста резко снижается (рис. 2). Увеличение густоты подроста ели в переходной зоне по сравнению с другими зонами экотонного комплекса наблюдалось нами при исследовании ельников черничных и прилегающих вырубок 5- и 10-летней давности [24].
Рис. 2.
Распределение елового и осинового подроста вдоль трансекты ельник–осинник. По горизонтали – расстояние от границы между двумя сообществами; по вертикали – количество подроста, шт. Fig. 2. The distribution of Norway spruce and aspen undergrowth along the Norway spruce forest–aspen forest transect. X-axis – the distance from the interface between two communities; y-axis – undergrowth number, inds.
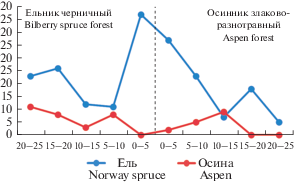
В целом можно утверждать, что в ельнике черничном условия для появления самосева ели более благоприятны, чем в осиннике благодаря особому микроклимату под материнским пологом (нет больших перепадов температур и влажности) и слабому развитию живого напочвенного покрова [25, 26]. Поэтому здесь отмечалась большая густота подроста – 3.1 тыс. шт./га (табл. 2). Однако, несмотря на высокую численность, преобладал мелкий подрост, высотой до 0.5 м, который составлял 92%. На долю среднего и крупного подроста приходилось только 8% от общего количества. При высокой полноте древостоя молодое возобновление ели испытывает сильное угнетение и погибает [27], поэтому в нашем случае, при полноте 1.2, из мелкого подроста в средний, а затем в крупный перешла лишь незначительная его часть. По результатам исследований Н.В. Беляевой [28] в условиях ельников черничных таежной зоны максимальное количество жизнеспособного подроста ели наблюдалось при относительной полноте 0.6–0.7, а при более высоких значениях данного показателя жизнеспособность подроста заметно снижалась.
Таблица 2.
Влияние условий произрастания на количество и распределение по крупности елового подроста Table 2. Effect of growing conditions on the number and size distribution of European spruce undergrowth
| Показатели Indicators |
Условия произрастания Growing conditions |
||
|---|---|---|---|
| Ельник черничный Bilberry spruce forest |
Переходная зона Transition zone |
Осинник злаково-разнотравный Aspen forest |
|
| Густота, тыс. экз./га Density, 103 ind./ha |
3.1 | 5.8 | 1.7 |
| Распределение подроста по крупности, % Size distribution of undergrowth, % |
|||
| Мелкий Seedlings |
92 | 60 | 58 |
| Средний и крупный Saplings |
8 | 40 | 42 |
В переходной зоне условия возобновления ели оказались наиболее благоприятными, что подтверждает максимальная густота подроста – 5.8 тыс. шт./га. Освещенность в первые годы после рубки древостоя была здесь значительно выше, чем в высокополнотном еловом насаждении, что способствовало не только появлению большого количества самосева, но и дальнейшему его росту и развитию. Поэтому доля среднего и крупного подроста в переходной зоне достигает 40%. В настоящее время измерения освещенности, проведенные в ясную безоблачную погоду на высоте 0.5 м и 1.3 м над поверхностью почвы, показали, что ее средние значения, составляющие от 1500 до 2000 люкс, в ельнике черничном и осиннике значимо не различались.
Осиннник злаково-разнотравный в своем развитии прошел стадию злаковой вырубки. Мощная дернина вейника и луговика препятствовала появлению елового самосева. В результате общее количество подроста здесь существенно ниже, чем в рассмотренных выше сообществах (1.7 тыс. шт./га), но доля среднего и крупного подроста сопоставима с таковой в переходной зоне и составляет 42%.
Выявить значимые различия в количестве подроста лиственных пород в зонах экотонного комплекса не удалось, вероятно, по причине низких показателей его густоты.
Радиальный прирост деревьев ели. В момент рубки смежного лесного участка (начало 80-х годов) средний возраст ели в ельнике черничном составлял примерно 55 лет. Представляет интерес проследить реакцию деревьев по радиальному приросту на протяжении последующих 35 лет с момента проведения рубки. Кроме того, по изменению ширины годичных слоев у деревьев, находящихся на разном удалении от границы контакта ельника и осинового насаждения, сформировавшегося на вырубке, можно выявить дальность распространения влияния рубки на лесное сообщество.
В первые пять лет после рубки смежного лесного участка радиальный прирост деревьев ели увеличивался, но значимо не отличался от средних значений до рубки (рис. 3). Значимые различия проявились через 5–10 лет после рубки у деревьев, расположенных на расстоянии до 10 м от границы вырубки (табл. 3). На расстоянии более 10 м достоверной реакции деревьев не обнаружено. У деревьев, находящихся вблизи края леса (в зоне 0–5 м), статистически значимое увеличение радиального прироста по сравнению с 5-летним периодом до рубки наблюдалось на протяжении 20–25 лет после сплошной рубки. Таким образом, достоверное влияние сплошной рубки на прирост древостоя по диаметру распространяется на расстоянии около 10 м от края вырубки, а период влияния составляет более двух десятилетий.
Рис. 3.
Средние значения радиального прироста деревьев ели европейской на разном расстоянии от края елового леса. По горизонтали – годы; по вертикали – ширина годичного слоя, мм. Цветом обозначены кривые радиального прироста деревьев на разном расстоянии от границы с осинником. Fig. 3. Average values of radial increment for Norway spruce trees at different distances from the edge of the spruce forest. X-axis – years; y-axis – width of annual increment layer, mm. The color indicates the radial increment curves of trees at different distances from the border with aspen forest.
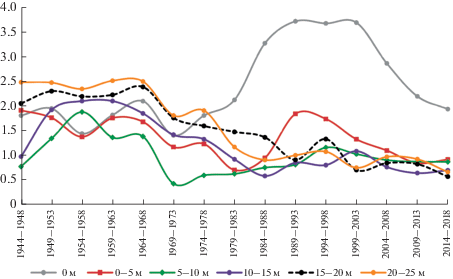
Таблица 3.
Средние значения радиального прироста деревьев ели европейской на разном расстоянии от края елового леса Table 3. Average values of radial increment for European spruce trees at different distances from the edge of the spruce forest
| Расстояние от края ельника, м Distance from the spruce forest border, m |
До рубки
(1979–1983) Before clearcut (1979–1983) |
Средняя ширина годичного слоя, мм Average values of radial increment, mm |
||||||
|---|---|---|---|---|---|---|---|---|
| 1984–1988 | 1989–1993 | 1994–1998 | 1999–2003 | 2004–2008 | 2009–2013 | 2014–2018 | ||
| 0 | 2.12 | 3.26/1.72 | 3.71/2.56* | 3.66/2.46* | 3.68/2.51* | 2.85/1.20 | 2.19/0.12 | 1.93/0.31 |
| 0–5 | 0.70 | 0.95/1.37 | 1.84/4.64*** | 1.73/4.61*** | 1.32/3.17** | 1.10/2.12* | 0.84/0.92 | 0.92/1.36 |
| 5–10 | 0.62 | 0.75/1.29 | 0.81/1.40 | 1.16/3.95*** | 1.03/4.09*** | 0.90/2.77* | 0.87/2.50* | 0.87/1.45 |
| 10–15 | 0.92 | 0.58/1.81 | 0.85/0.41 | 0.80/0.65 | 1.08/0.86 | 0.76/0.90 | 0.64/1.49 | 0.70/1.23 |
В последующий период прирост снижается, что, вероятно, связано с формированием лиственного молодняка и, следовательно, уменьшением доступности почвенных ресурсов и снижением освещенности. По литературным данным [29–33] после проведения рубок ухода в хвойных древостоях деревья, примыкающие к волокам, также увеличивали радиальный прирост, однако длительность периода увеличения прироста во многом зависела от состава древостоя и интенсивности рубки.
Следует отметить, что и до рубки наблюдались значимые различия годичных приростов у деревьев, произраставших на разном удалении от границы будущей вырубки, и между 5-летними периодами на протяжении роста деревьев на одинаковом расстоянии, однако после рубки эти различия большей частью нивелируются.
Эпифитный моховой покров на стволах осины. В ходе исследования эпифитного покрова было выявлено изменение встречаемости и величины куртин Hylocomium splendens на стволах осины в зависимости от удаления форофита от границы примыкания двух лесных сообществ. Так у самой границы ельника H. splendens либо совсем отсутствует на стволах осины, либо протяженность его куртин минимальна (1–2 см). Максимальная протяженность куртин мха на стволе осины достигается на самом удаленном вглубь леса дереве (29 метров от края ельника) и составляет 60 см от основания ствола.
Корненасыщенность верхнего горизонта почвы. Результаты исследования показали, что масса корней ели закономерно уменьшается (p < 0.01) в ряду ельник черничный–переходная зона–осинник злаково-разнотравный (табл. 4). В связи с тем, что корни разрастаются по площади, заполняя доступные места [34, 35], сложно провести строгую “подземную” границу между ельником и лиственным древостоем, в котором могут размещаться корни деревьев ели с прилегающих участков. Как известно, радиус корневой системы ели превышает радиус кроны в 2–3 раза [34]. Согласно К.С. Бобковой [36] в спелом ельнике чернично-зеленомошном корни ели распространяются в радиусе 8–10 м, отдельные корни простираются до 15 м. Этим можно объяснить достаточно высокое значение массы корней ели в лиственном насаждении (табл. 4) несмотря на ее угнетенное положение, о чем свидетельствуют таксационные показатели (табл. 1).
Таблица 4.
Масса корней древесных пород (диаметр до 3 мм) в верхнем 20-сантиметровом слое почвы Table 4. Weight of tree species roots (diameter up to 3 mm) in the upper 20-cm soil layer
| Зоны экотонного комплекса Ecotone complex zones |
Корни ели европейской, т/га Roots of European spruce, t/ ha |
Корни лиственных, т/га Hardwood roots, t/ ha |
Всего, т/га Total, t/ ha |
|---|---|---|---|
| Ельник черничный Bilberry spruce forest |
2.62 ± 0.27 | 1.57 ± 0.32 | 4.19 ± 0.53 |
| Переходная зона Transition zone |
1.73 ± 0.26 | 1.67 ± 0.26 | 3.39 ± 0.45 |
| Осинник злаково-разнотравный Aspen forest |
0.76 ± 0.13 | 1.96 ± 0.21 | 2.72 ± 0.14 |
Насыщенность верхнего слоя почвы корнями лиственных пород значимо не различается в разных зонах экотонного комплекса. Это, по-видимому, объясняется тем, что и запас древесины лиственных пород в ельнике черничном и осиннике злаково-разнотравном практически не различается. Корни лиственных характеризуются большей энергией роста по сравнению с корнями хвойных пород. По мере самоизреживания или проведения рубок, направленных на снижение доли лиственных пород, почвенные условия для ели будут улучшаться, что подтверждается литературными данными [37, 38].
Плотность эктомикориз ели. Оценка состояния тонких корней ели показала, что значимых различий морфолого-анатомических параметров микоризных окончаний в экотонном комплексе ельник–осинник не наблюдается (табл. 5).
Таблица 5.
Показатели состояния микоризных окончаний ели европейской в трех зонах экотонного комплекса Table 5. Indicators of the European spruce mycorrhizal endings state in three zones of the ecotone complex
| Зоны экотонного комплекса Ecotone complex zones |
Плотность микоризы, шт./10 см корня Mycorrhiza density, ex./10 cm of the root |
Общий радиус микоризного окончания, мкм Total radius of the mycorrhizal ending, µm |
Радиус корня в микоризном окончании, мкм Root radius in the mycorrhizal ending, µm |
Толщина мицелиального чехла, мкм Thickness of the mycelial sheath, µm |
Доля мицелиального чехла, % Proportion of the mycelial sheath, % |
|---|---|---|---|---|---|
| Ельник черничный Bilberry spruce forest |
80.4 ± 2.6 | 183.2 ± 9.7 | 170.6 ± 9.6 | 12.6 ± 0.6 | 13.4 ± 0.7 |
| Переходная зона Transition zone |
82.1 ± 3.1 | 186.4 ± 4.8 | 175.0 ± 4.8 | 11.4 ± 0.4 | 12.0 ± 0.5 |
| Осинник злаково-разнотравный Aspen forest |
83.7 ± 4.4 | 165.2 ± 3.9 | 153.4 ± 3.8 | 11.8 ± 0.3 | 13.8 ± 0.4 |
Некоторая тенденция увеличения количества микоризных окончаний при переходе от ельника к осиннику может рассматриваться как адаптивная реакция на неблагоприятные условия произрастания ели среди лиственных пород, обгоняющих ее в росте [39–41].
Нижние ярусы сообществ. По видовому составу сосудистых растений и мхов ельник черничный, сообщество переходной зоны и осинник злаково-разнотравный, составляющие экотонный комплекс, различаются незначительно (14, 14 и 17 видов соответственно). В осиннике отмечено несколько бóльшее количество видов сосудистых растений по сравнению с другими зонами экотонного комплекса (табл. 6).
Таблица 6.
Характеристики травяно-кустарничкового яруса в сообществах экотонного комплекса Table 6. Dwarf shrub and herb layer characteristics in the ecotone complex communities
| Виды Species |
Ельник черничный Bilberry spruce forest |
Сообщество переходной зоны Transition zone |
Осинник злаково-разнотравный Aspen forest |
|---|---|---|---|
| Общее проективное покрытие Total projective cover |
18 | 30 | 15 |
| Athyrium filix-femina (L.) Roth | – | – | 3* (+)** |
| Avenella flexuosa (L.) Drejer | 35 (+) |
43 (+) |
83 (+) |
| Calamagrostis arundinacea (L.) Roth | 3 (+) |
34 (1) |
62 (2) |
| Dryopteris carthusiana (Vill.) H.P. Fuchs | 3 (+) |
3 (+) |
51 (5) |
| Gymnocarpium dryopteris (L.) Newman | – | – | 16 (+) |
| Linnaea borealis L. | 44 (+) |
45 (+) |
– |
| Luzula pilosa (L.) Willd. | 4 (+) |
22 (+) |
30 (+) |
| Lycopodium annotinum L. | 6 (+) |
3 (+) |
– |
| Maianthemum bifolium (L.) F.W. Schmidt | 69 (+) |
42 (+) |
86 (1) |
| Melampyrum spp. | 19 (+) |
26 (+) |
47 (+) |
| Orthilia secunda (L.) House | 14 (+) |
8 (+) |
5 (+) |
| Oxalis acetosella L. | 85 (2) |
1 (+) |
44 (+) |
| Potentilla erecta (L.) Raeusch. | – | – | 2 (+) |
| Rubus saxatilis L. | – | – | 21 (+) |
| Solidago virgaurea L. | 3 (+) |
12 (+) |
67 (2) |
| Trientalis europaea L. | 18 (+) |
35 (+) |
68 (+) |
| Vaccinium myrtillus L. | 100 (12) |
98 (20) |
50 (2) |
| Vaccinium vitis-idaea L. | 66 (2) |
83 (7) |
34 (+) |
| Veronica chamaedrys L. | – | – | 3 (+) |
Примечание: * – среднее значение встречаемости (%); ** – среднее проективное покрытие (%); “+" – проективное покрытие вида меньше 1%; прочерк означает отсутствие вида. Note: * – the average value of the occurrence (%); ** – the average projective cover (%); “+”– projective cover is less than 1%; a dash indicates the absence of the species.
Заметные различия наблюдаются в структуре травяно-кустарничкового и мохового ярусов. В ельнике черничном основной вклад в сложение травяно-кустарничкового яруса вносят черника (покрытие 12%) и брусника (2%), среднее покрытие других видов на учетных площадках не превышает 1%. В напочвенном покрове доминируют зеленые мхи (Hylocomium splendens, Pleurozium schreberi, Dicranum scoparium).
Переходная зона (по 8 м в обе стороны от зоны контакта двух лесных сообществ) по участию видов сосудистых растений сохраняет основные черты структуры напочвенного покрова ельника. Здесь также доминируют кустарнички: черника и брусника, причем их проективное покрытие (соответственно 20 и 7%) выше, чем в среднем в ельнике, обилие остальных видов незначительно. В напочвенном покрове наряду с зелеными мхами участвуют мхи влажных местообитаний: политрихум обыкновенный и сфагновые мхи.
В осиннике наибольшим обилием обладает щитовник картузианский (5%), при этом несколько видов сосудистых растений также заметно участвуют в сложении напочвенного покрова со средним проективным покрытием 1–2% (вейник лесной, золотарник обыкновенный, черника и майник двулистный). Из-за большого объема лиственного опада и развития травянистой растительности в целом снижается обилие мхов и лесных кустарничков.
В исследованном экотонном комплексе для травяно-кустарничкового яруса ельника черничного и переходной зоны получены более низкие значения индекса Пиелу (0.65 и 0.54 соответственно), чем для осинника злаково-разнотравного (0.87). Это свидетельствует о различиях в структуре напочвенного покрова. В ельнике и в переходной зоне выраженным доминантом травяно-кустарничкового яруса является черника, в осиннике заметных доминантов нет, в сложении травяно-кустарничкового яруса участвуют несколько видов сосудистых растений с относительно небольшим, близким по величине (2–5%) проективным покрытием.
Наиболее заметные различия структуры напочвенного покрова наблюдаются по участию групп видов сосудистых растений. В ельнике черничном преобладают лесные кустарнички (черника и брусника), доля их участия в составе травяно-кустарничкового яруса составляет чуть менее 80% (рис. 4), в то время как в осиннике злаково-разнотравном – менее 15% от общей суммы проективных покрытий всех видов яруса. Переходная зона по доминированию лесных кустарничков очень схожа с ельником черничным.
Рис. 4.
Доля участия групп видов сосудистых растений в травяно-кустарничковом ярусе сообществ экотонного комплекса. Группы растений: 1 – кустарнички, 2 – травы, 3 – злаки. Fig. 4. The proportion of vascular plant species groups in the dwarf shrub-herb layer in the communities of the ecotone complex. Plant groups: 1 – dwarf shrubs, 2 – herbs, 3 – graminoids.
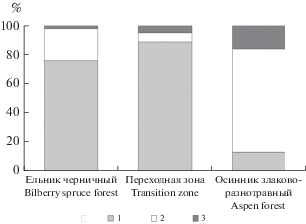
В 35-летнем лиственном насаждении сохраняется та же особенность сложения напочвенного покрова, что и в растительных сообществах более ранних стадий восстановления (2–10 лет после рубки ельника черничного) – примерно одинаковое участие нескольких видов в травяно-кустарничковом ярусе и низкая доля лесных кустарничков [14, 16].
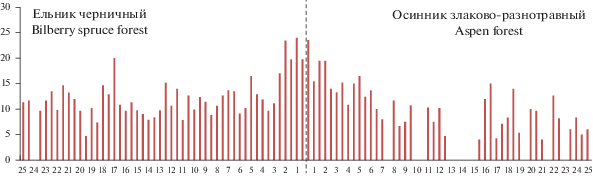
На графиках, отражающих изменение проективного покрытия кустарничков вдоль трансекты из ельника черничного до осинника, заметно повышение обилия черники (до 30–40%) в зоне шириной 4–5 м в обе стороны от границы ельник–осинник и брусники (до 10–20%) – в зоне шириной 2 м (рис. 5). Хорошо выделяется пе-реходная зона и по показателю высоты кустарничков (рис. 6, 7). Средние значения высоты побегов в ней оказались значимо выше по сравнению с другими зонами экотонного комплекса (P < 0.001). Ранее нами было показано, что в первые годы после рубки ельника черничного проективное покрытие и высота лесных кустарничков в экотонном комплексе снижаются от леса к вырубке [14, 16].
Рис. 5.
Средние значения проективного покрытия черники и брусники на трансекте ельник–осинник. По горизонтали – расстояние от края леса, м; по вертикали – проективное покрытие, %. Fig. 5. The average values of the projective cover of bilberry and cowberry along the Norway spruce forest–aspen forest transect. X-axis – distance from forest edge, m; y-axis – projective cover, %.
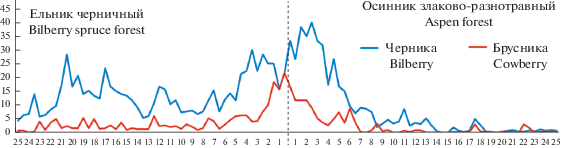
Рис. 6.
Высота побегов черники на трансекте ельник–осинник. По горизонтали – расстояние от края леса, м; по вертикали – средняя высота побегов, см. Fig. 6. The height of bilberry shoots along the Norway spruce forest–aspen forest transect. X-axis – distance from forest edge, m; y-axis – average shoot height, cm.
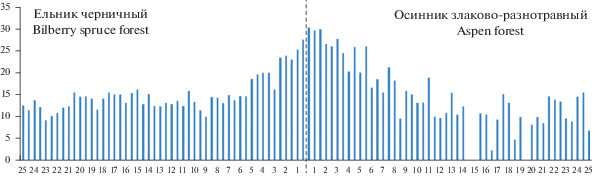
Рис. 7.
Высота побегов брусники вдоль трансекты. По горизонтали – расстояние от края леса, м; по вертикали – средняя высота побегов, см. Fig. 7. The height of cowberry shoots along the Norway spruce forest–aspen forest transect. X-axis – distance from forest edge, m; y-axis – average shoot height, cm.
ЗАКЛЮЧЕНИЕ И ВЫВОДЫ
После 35 лет естественного восстановления леса на вырубке в целом сохраняется структура экотонного комплекса: между ельником черничным и осинником злаково-разнотравным по ряду признаков выделяется переходная зона, занимающая примерно по 10 м в обе стороны от границы между сообществами.
Максимальные размеры переходной зоны – 10 м от границы сообществ – определены по изменению радиального прироста, который был в среднем выше, чем центральной части ельника черничного в течение двух десятилетий после рубки. Позднее различия уже не обнаруживались, что, вероятно, связано с формированием лиственного насаждения, которое привело к снижению доступности почвенных ресурсов и уменьшению освещенности.
Максимальная густота подроста ели наблюдалась в переходной зоне, здесь же наряду с более высокой густотой отмечено значительное повышение доли среднего и крупного подроста по сравнению с ельником черничным. В осиннике злаково-разнотравном появление самосева ели затруднено из-за развития травяно-злаковой растительности и плотного слоя опавших листьев осины, поэтому количество подроста ели значительно ниже.
Минимальная протяженность куртин Hylocomium splendens на стволах осины в ельнике черничном отмечается у границы с осинником, максимальная – на самом удаленном дереве. Это свидетельствует о более благоприятных условиях для формирования эпифитного покрова, то есть более высокой влажности воздуха и субстрата в глубине лесного сообщества по сравнению с границей примыкания к лиственному молодняку.
Насыщенность корнями ели верхнего слоя почвы снижается в направлении от ельника черничного к осиннику злаково-разнотравному. Довольно высокая масса корней ели в лиственном насаждении при относительно небольшом количестве елового подроста, вероятно, обусловлена их распространением с соседних участков. Масса корней лиственных пород, которые отличаются способностью к активному разрастанию, в среднем не различается в разных зонах экотонного комплекса и находится в соответствии с величиной запаса древесины лиственных в составе древостоев.
В данном исследовании не удалось выявить значимых различий параметров эктомикоризы ели в экотонном комплексе ельника черничного и осинника злаково-разнотравного. Поскольку подземная часть биогеоценоза подвержена влиянию факторов внешней среды в меньшей степени, чем надземная, то необходимы дальнейшие исследования для установления подземных границ взаимовлияния ельника и осинника.
Через 35 лет после рубки древостоя переходная зона по видовому составу и по соотношению функциональных групп растений обладает большим сходством с примыкающим к ней ельником, чем с лиственным сообществом, являющимся ранней стадией восстановления ельника черничного. Однако по таким показателям как проективное покрытие и средняя высота лесных кустарничков, среди которых доминируют такие важные ресурсные виды, как черника и брусника, переходная зона отличается от обоих лесных сообществ.
Список литературы
Правила заготовки древесины и особенностей заготовки древесины в лесничествах, лесопарках, указанных в статье 23 Лесного кодекса Российской Федерации. Утв. приказом Министерства природных ресурсов и экологии Российской Федерации № 474 от 13.09.2016 г.
Burton P.J. 2002. Effects of clearcut edges on trees in the sub-boreal spruce zone of Northwest-Central British Columbia. – Silva Fenn. 36(1): 329–352. https://doi.org/10.14214/sf.566
Pauchard A., Alaback P.B. 2006. Edge type defines alien plant species invasion along Pinus contorta burned, highway and clearcut forest edges. – Forest Ecol. Manag. 223(1–3): 327–335. https://doi.org/10.1016/j.foreco.2005.11.020
Бурова Н.В., Тараканов А.М., Дроздов И.И., Кононов О.Д., Гельфанд Е.Д. 2012. Влияние опушечного эффекта на состояние отдельных компонентов лесных биогеоценозов. – Вестн. МГУЛ. Лесной вестник. 4 (87): 19–22.
Торопова Е.В., Старицын В.В. 2019. Продуктивность черники обыкновенной (Vaccinium myrtillus L.) в экотонной зоне вырубки. – В сб.: Проблемы обеспечения экологической безопасности и устойчивое развитие арктических территорий: Сбор. матер. Всерос. конф. с межд. участием II Юдахинские чтения. Архангельск. С. 407–412.
Stoll P., Bergius E. 2005. Pattern and process: competition causes regular spacing of individuals within plant populations. – J. Ecol. 93(2): 395–403. https://doi.org/10.1111/j.0022-0477.2005.00989.x
Колобов А.Н. 2011. Моделирование процессов конкуренции за свет в смешанных разновозрастных древостоях. – Вестник Томского государственного университета. 351: 155–158. http://journals.tsu.ru/engine/download.php?id=18048&area=files
Иванов B.В. 2005. Экологические последствия механизированных лесозаготовок в южной тайге Красноярского края. – Лесоведение. 2: 3–8. http://lesovedenie.ru/index.php/forestry/article/view/690
Уланова Н.Г. 2007. Механизмы сукцессий растительности сплошных вырубок в ельниках южной тайги. – В сб.: Актуальные проблемы геоботаники: Матер. III Всерос. школы-конф. Петрозаводск. С. 198–211. http://resources.krc.karelia.ru/geobotany/doc/school2007-0/actual_probl_geobotany_lectures_198-211.pdf
Банева Н.А. 1986. Восстановление массы корней ели после рубок ухода. – Лесоведение. 3: 62–66.
Harper K.A., Lesieur D., Bergeron Y., Drapeau P. 2004. Forest structure and composition at young fire and cut edges in black spruce boreal forest. – Can. J. Forest Res. 34(2): 289–302. https://doi.org/10.1139/x03-279
Baker T., Jordan G., Baker S. 2016. Microclimatic edge effects in a recently harvested forest: Do remnant forest patches create the same impact as large forest areas? – Forest Ecol. Manag. 365: 128–136. https://doi.org/10.1016/j.foreco.2016.01.022
Pellissier V., Berges L., Nedeltcheva T., Schmitt M.-C., Avon C., Cluzeau C., Dupoey J.-L. 2013. Understorey plant species show long-range spatial patterns in forest patches according to distance-to-edge. – J. Veg. Sci. 24(1): 9–24. https://doi.org/10.1111/j.1654-1103.2012.01435.x
Геникова Н.В., Торопова Е.В., Крышень А.М. 2016. Реакция видов напочвенного покрова ельника черничного на рубку древостоя. – Труды КарНЦ РАН. Сер. Экологические исследования. 4: 92–99. https://doi.org/10.17076/eco292
Геникова Н.В., Крышень А.М. 2018а. Динамика напочвенного покрова северотаежного ельника черничного в первые годы после рубки. – Бот. журн. 103(3): 364–381. https://doi.org/10.1134/S0006813618030067
Геникова Н.В., Торопова Е.В., Крышень А.М., Мамонтов В.Н. 2018б. Изменение структуры напочвенного покрова в экотонном комплексе “лес–опушка–вырубка” в условиях ельника черничного через 10 лет после рубки. – Труды КарНЦ РАН. Сер. Экологические исследования. 10: 1–15. https://doi.org/10.17076/eco877
Кузьмичев В.В., Миндеева Т.Н., Черкашин В.П. 1989. Оценка взаимодействия деревьев в лесных фитоценозах. – Известия Сибирского отделения АН СССР. Серия биологических наук. 3: 133–139.
Ledermann T., Stage A.R. 2001. Effects of competitor spacing in individual-tree indices of competition. – Can. J. Forest Res. 31(12): 2143–2150. https://doi.org/10.1139/x01-153
Касаткин А.С., Бойко А.А., Колтунова А.И., Гаврилин Д.С. 2012. Радиальный прирост сосны обыкновенной в условиях конкуренции. – Известия Оренбургского государственного аграрного университета. 2(34): 50–54. https://orensau.ru/ru/component/docman/doc_download/2595-izvestiya–34
Количественные методы экологии и гидробиологии. Под ред. Розенберга Г.С. 2005. Тольятти. 404 с.
Веселкин Д.В. 2003. Изменчивость анатомических параметров эктомикоризных окончаний разного строения. – Микология и фитопатология. 37(1): 22–29. https://www.binran.ru/files/journals/MiF/MiF_2003_37_1.pdf
Данченко А.М., Бех И.А. 2009. Оценка типологического разнообразия лесных экосистем на основе данных таксации и ландшафтно-типологического анализа модельных территорий. – Вестник Томского государственного университета. Биология. 2: 70–74.
Жирин В.М. Князева С.В., Эйдлина С.П., Зукерт Н.В. 2012. Сезонная информативность многоспектральных космических снимков высокого разрешения при изучении породно-возрастной динамики лесов. – Современные проблемы дистанционного зондирования Земли из космоса. 9(1): 87–95. http://d33.infospace.ru/d33_conf/sb2012t1/87-94.pdf
Воспроизводство и повышение продуктивности лесов Восточной Фенноскандии на основе естественных процессов и интенсивных методов лесовыращивания: Промежуточный отчет о научно-исследовательской работе. 2017. Руководитель А.М. Крышень. Петрозаводск. 43 с.
Мелехов И.С. 1972. Лесоведение и лесоводство. М. 178 с.
Бобкова К.С., Бессонов И.М. 2009. Естественное возобновление в среднетаежных ельниках Европейского Северо-востока. – Лесоведение. 5: 10–16.
Ильчуков С.В. 2008. Горизонтальная структура подроста ели в спелых среднетаежных ельниках. – Известия высших учебных заведений. Лесной журнал. 1: 65–69. http://lesnoizhurnal.ru/issuesarchive/?ELEMENT_ID=675
Беляева Н.В. 2013. Закономерности изменения структуры и состояния молодого поколения ели в условиях интенсивного хозяйственного воздействия: Автореф. дис. … д-ра с.-х. наук. СПб. 43 с.
Сеннов С.Н. 2001. Проблемы лесоведения. СПб. 56 с.
Синькевич С.М. 2005. Влияние рубок ухода на рост сосновых насаждений. – В сб.: Лесоводственно-экологические аспекты хозяйственной деятельности в лесах Карелии. Петрозаводск. С. 101–122.
Vincent M., Krause C., Zhang S.Y. 2009. Radial growth response of black spruce roots and stems to commercial thinning in the boreal forest. – Forestry. 82(5): 557–571. https://doi.org/10.1093/forestry/cpp025
Карпечко А.Ю. 2013. Изменения среднепериодического прироста по диаметру и связь массы корней надземной части древостоя после разреживания. – Труды КарНЦ РАН. Сер. Экологические исследования. 6: 167–170. http://resources.krc.karelia.ru/transactions/doc/trudy2013/trudy_2013_6_167-170.pdf
Багаев С.С., Чудецкий А.И. 2018. Результаты рубок ухода в лиственно-еловых насаждениях Костромской области. – Лесохозяйственная информация. 1: 5–20. http://lhi.vniilm.ru/index.php/ru/bagaev-s-s-chudetskij-a-i-rezultaty-rubok-ukhoda-v-listvenno-elovykh-nasazhdeniyakh-kostromskoj-oblasti
Сеннов С.Н. 1980. Рубки ухода за лесом и внутривидовая конкуренция. – В сб.: Восстановление и мелиорация лесов Северо-Запада РСФСР. Л. С. 17–27.
Карпечко А.Ю. 2009. Влияние разреживания на корненасыщенность почвы еловых древостоев южной Карелии. – Известия вузов. Лесной журнал. 3: 19–25. http://lesnoizhurnal.ru/issuesarchive/?ELEMENT_ID=563
Бобкова К.С. 1987. Биологическая продуктивность хвойных лесов европейского Северо-Востока. Л. 156 с.
Чибисов Г.А. 1971. Рост корневых систем в березово-еловых насаждениях и влияние на них рубок ухода. – В сб. работ по лесному хозяйству и лесохимии. Архангельск. С. 121–130.
Зарубина Л.В., Коновалов В.Н. 2014. Эколого-физиологические особенности ели в березняках черничных. Архангельск. 378 с. https://rucont.ru/efd/557469
Шубин В.И. 1973. Микотрофность древесных пород, ее значение при разведении леса в таежной зоне. Л. 264 с.
Чумак Н.Ф. 1981. Микоризы сосны на песчаных почвах в связи с применением удобрений: Автореф. дис. … канд. биол. наук. Петрозаводск. 25 с.
Кикеева А.В., Чаженгина С.Ю., Чаженгина Е.А. 2019. Редкоземельные элементы в шунгитовых почвах: содержание, распределение и особенности микоризообразования. – Принципы экологии. 1(31): 32–46. https://doi.org/10.15393/j1.art.2019.8262
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы