

Растительные ресурсы, 2020, T. 56, № 3, стр. 221-240
Прирост кормовых видов лишайников рода Cladonia (Cladoniaceae) на пастбищах домашнего северного оленя
С. Ю. Соковнина (Абдульманова) 1, *, С. Н. Эктова 1
1 Институт экологии растений и животных УрО РАН
г. Екатеринбург, Россия
* E-mail: sokovnina_su@ipae.uran.ru
Поступила в редакцию 02.03.2020
После доработки 24.05.2020
Принята к публикации 10.06.2020
Аннотация
Кормовые виды лишайников – наиболее чувствительный компонент растительного покрова, особенно в условиях пастбищного использования обширных территорий в Арктике и на севере бореальной зоны. Объектами данного исследования являются кустисто-разветвленные лишайники – Cladonia arbuscula, C. rangiferina, C. stellaris, C. stygia, произрастающие в тундровой зоне полуостровов Ямал и Гыдан. Скорость роста лишайниковых подециев варьирует от 1.2 до 6.3 мм/год, живой части – 0.9–2.9 мм/год. Значимого снижения средних значений скорости роста модельных видов с юга на север, несмотря на значительное ужесточение гидротермических условий, не выявлено. В зональном градиенте наименьшие значения скорости роста изученных видов отмечены в сообществах южных субарктических тундр. Среди изученных типов фитоценозов наибольшие значения прироста выявлены в кустарниковых тундрах и сообществах бугристых болот. Биотическими факторами среды, оказывающими положительное влияние на рост лишайников на пастбищах, являются сомкнутость кустарникового яруса на юге и толщина мохово-лишайниковой покрова на севере. Вид с наибольшим восстановительным потенциалом в изученных сообществах – C. stygia. Наиболее чувствительный к выпасу вид – C. stellaris. Варьирование параметров подециев (высота и возраст подеция и его живой части) в изученных градиентах сходно. Наиболее стабильный параметр – возраст живой части.
Значительные территории в тундровой зоне Евразии и Северной Америки местное население активно использует в качестве пастбищных угодий для северных оленей [1, 2], а перевыпас, вызванный пастбищным оленеводством, является серьезной проблемой северных регионов уже многие годы [3]. В большинстве случаев пастбищные сукцессии являются ретрогрессиями, когда изменение растительности приводит к падению запасов биомассы, продуктивности, уменьшению содержания элементов минерального питания в живой части экосистемы [4], снижению альфа- и бета-разнообразия, а также изменению физических условий среды [5–7]. Напочвенные кустистые лишайники, формирующие плотный покров в отсутствии нарушений, являются наиболее чувствительным компонентом северных экосистем [2, 8–10]. В настоящее время в тундровой зоне полуострова Ямал ценные кормовые лишайники сохранили высокую встречаемость (за исключением C. stellaris (Opiz) Pouzar & Vězda), но имеют низкое проективное покрытие и существуют в крайне угнетенном состоянии (незначительная высота подециев, отсутствие отмершей части, механические повреждения подециев). На доминирующие позиции вышли виды, которые устойчивы к выпасу, но не имеют кормового значения [2]. Деградация растительности на пастбищах выражена не только в изменении видового состава, но и в снижении высоты и трансформации структуры напочвенного покрова. Пастбищное воздействие на рост лишайников происходит непосредственно через разрушение лишайниковых талломов и опосредованно – через нарушение целостности мохово-лишайникового яруса. Ежегодный прирост лишайников варьирует в зависимости от интенсивности нагрузки и степени угнетения покрова. Наибольшая нагрузка характерна для летних пастбищ, и той части территории, которую местное население использует в качестве прогонных путей, оказывая мощное воздействие на растительность несколько раз в течение года [11].
Цель данной работы – оценка скорости роста талломов ценных кормовых видов лишайников рода Cladonia P. Browne на летних пастбищах полуостровов Ямал и Гыдан с разным уровнем деградации напочвенного покрова. Для достижения поставленной цели было необходимо: 1) оценить величину прироста кустисто-разветвленных лишайников на широтном и фитоценотическом градиентах при разной степени трансформации напочвенного покрова; 2) определить значимые биотические факторы среды, положительно влияющие на скорость роста лишайников в сообществах, подверженных пастбищному использованию; 3) оценить восстановительный потенциал разных видов кустисто-разветвленных лишайников рода Cladonia.
МАТЕРИАЛЫ И МЕТОДЫ
Тундровая зона имеет значительную протяженность с юга на север около 750 (полуостров Ямал) и 400 км (полуостров Гыдан) в условиях морского субарктического климата и распространения вечной мерзлоты. На открытой для масс арктического воздуха (как морского, так и континентального) территории лето короткое и прохладное, зима продолжительная и морозная. Среднегодовая температура на юге полуострова равна –6.6 °С, на севере –10.2 °С. Температура самого теплого месяца (июль) на юге 14.5 °С, на севере (конец августа) – 7.8 °С. Температура самого холодного (январь) месяца на юге –24.9 °С, на севере –59 °С. Летом температура воздуха может достигать 30 °С на всей территории. Годовое количество осадков от 400 мм на юге до 250 мм на севере, осадки выпадают в летний период в виде моросящих дождей, испарение незначительное – 20–25% от поступающей влаги. Снеговой покров незначителен. Максимальная продолжительность вегетационного периода – 2 месяца (июль, август). Для всей территории характерны высокая влажность воздуха, туманы и облачность, сильные ветры [2, 12–15].
Исследования проводились в подзонах субарктических и арктических тундр, первая из которых делится на две полосы – южных (кустарниковых) и северных (типичных) субарктических тундр. Различия между двумя полосами субарктических тундр выражены в разном участии кустарников в растительном покрове, а также в сочетании растительных сообществ на вершинах и склонах водоразделов. Проективное покрытие (ПП) кустисто-разветвленных лишайников в субарктических тундрах до 40%, в арктических тундрах может достигать 60% [2, 12].
В полосе южных субарктических тундр описан экологический ряд сообществ: кустарниковые тундры – открытые тундровые сообщества – плоскобугристые болота.
Кустарниковые фитоценозы представлены ерниковыми лишайниково-моховыми, кустарниковыми кустарничково-мохово-лишайниковыми, бугорковатыми кустарниковыми кустарничково-мохово-лишайниковыми, заболоченными кустарниковыми кустарничково-травяно-лишайниково-моховыми и бугорковатыми кустарниковыми травяно-моховыми тундрами. Сомкнутость кустарникового яруса (Betula nana L. и Salix lanata L.) в сообществах 0.2–0.6. Кустарнички представлены Empetrum nigrum L., Ledum decumbens (Aiton) Lodd. ex Steud., Arctous alpina (L.) Nied., Vaccinium uliginosum L. и V. vitis-idaea L. (ПП 5–70%), травы – Carex bigelowii Torr. ex Schwein., Equisetum arvense L, Calamagrostis lapponica (Wahlb.) Hartm. (ПП 20–45%). Из мхов наиболее обильны Aulacomnium turgidum (Wahlenb.) Schwägr., Pleurozium schreberi (Brid.) Mitt, Dicranum elongatum Schleich. ex Schwägr. (ПП 20–95%), лишайники представлены Cladonia rangiferina (L.) F. H. Wigg, C. stygia (Fr.) Ruoss., С. arbuscula (Wallr.) Flot, Cetraria islandica (L.) Ach., Flavocetraria cucullata (Bellardi) Kärnefelt et A. Thell, F. nivalis (L.) Kärnefelt et A. Thell, Cladonia cornuta (L.) Hoffm., С. crispata (Ach.) Flot., С. gracilis (L.) Willd., С. pleurota (Flörke) Schaer., Peltigera aphthosa (L.) Willd. (ПП 15–80%), толщина мохово-лишайникового покрова 1–4 см.
К открытым тундровым фитоценозам отнесены бугорковатые кустарничковые, кустарничково-лишайниковые, пятнистые кустарничково-лишайниковые, травяно-кустарничковые тундры. Кустарниковый ярус в сообществах не сформирован, единично встречаются Betula nana, Salix reptans Rupr. Травяно-кустарничковый ярус формируют Salix polaris Wahlenb., Ledum decumbens, Empetrum nigrum, Vaccinium vitis-idaea (ПП кустарничков 5–60%), Carex bigelowii, Rubus chamaemorus L., Poa alpina L., Luzula confusa Lindeb. (ПП трав 5–45%). В напочвенном покрове доминируют лишайники Gowardia nigricans (Ach.) P. Halonen, L. Myllys, S. Velmala & H. Hyvärinen, Sphaerophorus globosus (Huds.) Vain., Flavocetraria nivalis, Ochrolechia frigida (Sw.) Lynge, Bryocaulon divergens (Ach.) Kärnefelt, Cladonia arbuscula, С. rangiferina, C. stygia, С. uncialis (L.) Weber ex F. H. Wigg. (ПП 5–90%), мхи представлены Polytrichum hyperboreum R. Br., Dicranum elongatum, D. fuscescens Turner, Aulacomnium turgidum (ПП 1–30%), толщина покрова 1.5–4.5 см.
В сообществах плоскобугристых болот сомкнутость кустарников (Betula nana) незначительна (до 0.1). В травяно-кустарничковом ярусе доминируют травы Carex rariflora (Wahlenb.) Sm., Eriophorum scheuchzeri Hoppe, Hierochloe alpina (Sw.) Roem. & Schult., Equisetum arvense (ПП 35–45%); кустарнички представлены Arctous alpina, Salix polaris, Ledum decumbens, Empetrum nigrum, (ПП около 15%). Напочвенный покров сформирован мхами Aulacomnium turgidum, Polytrichum strictum Brid., Р. juniperinum Hedw., Dicranum angustum Lindb., Hylocomium splendens (Hedw.) Bruch et al., Sanionia uncinata (Hedw.) Loeske, Sphagnum russowii Warnst., S. balticum (Russow) C.E.O.Jensen (ПП 50–70%) с участием лишайников Sphaerophorus globosus, Flavocetraria cucullata, F. nivalis, Bryocaulon divergens, Cladonia arbuscula, C. stygia, Alectoria ochroleuca (Hoffm.) A. Massal. (ПП около 30%), толщина покрова 1–5 см.
В полосе северных субарктических тундр экологический ряд изученных сообществ включает кустарниковые тундры, открытые тундровые сообщества и бугристые болота.
Кустарниковые тундры представлены заболоченными кустарниковыми кустарничково-травяно-лишайниково-моховыми, кустарниковыми травяно-моховыми, фрагментарными кустарниковыми травяно-моховыми, бугорковатыми травяно-моховыми, кустарниковыми травяно-лишайниково-моховыми тундрами. Кустарниковый ярус в сообществах сформирован низкорослыми кустами Betula nana и Salix lanata (высота до 20 см) сомкнутостью 0.1–0.9. В травяно-кустарничковом ярусе представлены кустарнички Vaccinium vitis-idaea, Ledum decumbens, Salix nummularia Andersson (ПП 20–80%), участие трав Carex bigelowii, Carex concolor R. Br., Rubus chamoemorus, Equisetum arvense, Eriophorum vaginatum L., Festuca ovina L., Calamagrostis stricta (Timm) Koeler, незначительно (ПП до 2%). В напочвенном покрове доминируют мхи Pleurozium schreberi, Polytrichum juniperinum, Р. hyperboreum R. Br., Aulacomnium turgidum, Sphagnum magellanicum Brid. (ПП 40–95%), лишайники представлены Thamnolia vermicularis (Sw.) Schaer., Flavocetraria cucullata, Cladonia amaurocraea (Flörke) Schaer., C. arbuscula, C. rangiferina, C. stygia, C. cornuta, Sphaerophorus globosus, Dactylina arctica (Hook. f.) Nyl., Cetraria islandica, Flavocetraria cucullata, F. nivalis, Peltigera aphthosa (ПП до 30%), толщина покрова 1–10 см.
К открытым тундровым сообществам отнесены пятнистые травяно-мохово-лишайниковые, кустарничково-лишайниковые, моховые, кустарничковые лишайниково-моховые, бугорковатые травяно-лишайниково-моховые тундры. Для этих фитоценозов характерна значительная доля кустарничков Salix nummularia, Vaccinium vitis-idaea, Ledum decumbens (ПП 15–75%), травы представлены Carex bigelowii, Роа arctica R. Br., Calamagrostis holmii Lange, Calamagrostis stricta, Hierochloe alpina, Luzula spicata (L.) DC. (ПП 3–50%). Мохово-лишайниковый ярус образован Aulacomnium turgidum, Racomitrium lanuginosum (Hedw.) Brid. (ПП мхов 15–80%), Alectoria ochroleuca, Gowardia nigricans, Bryocaulon divergens, Cetraria islandica, Cladonia arbuscula, C. rangiferina, C. stygia, C. uncialis, Flavocetraria cucullata, Ochrolechia frigida, Sphaerophorus globosus (ПП лишайников 5–85%), толщина покрова 1–5 см.
В сообществах бугристых болот сомкнутость кустарников (Betula nana) низкая (до 0.2), доминируют травы Rubus chamaemorus, Eriophorum medium Andersson, Роа arctica, Calamagrostis stricta, Carex rariflora (ПП до 70%), ПП кустарничков Vaccinium vitis-idaea, Ledum decumbens незначительно (5–20%). В напочвенном покрове доминируют мхи (ПП 70–95%), доля лишайников Alectoria ochroleuca, Gowardia nigricans, Bryocaulon divergens, Bryoria nitidula (Th. Fr.) Brodo et D. Hawksw., Cetraria islandica, С. laevigata Rass., Flavocetraria cucullata, Cladonia arbuscula, С. rangiferina, C. stygia, C. amaurocraea, С. macroceras (Delise) Hav., С. uncialis, Dactylina arctica, Thamnolia vermicularis невелика (ПП 5–30%), толщина покрова 2–8 см.
В подзоне арктических тундр изученный экологический ряд сообществ включает кустарничково-лишайниковые тундры, пятнистые травяно-мохово-лишайниковые тундры и бугристые болота.
В кустарничково-лишайниковых тундрах преобладают кустарнички Salix polaris, Cassiope tetragona (L.) D. Don, Dryas octopetala L. (ПП 15–40%); травы представлены Carex bigelowii, Arctagrostis latifolia (R. Br.) Griseb., Luzula confusa, Hierochloe alpina (ПП 5–30%). В напочвенном покрове максимальная доля участия лишайников Alectoria ochroleuca, Bryocaulon divergens, Cetrariella delisei (Bory ex Schaer.) Kärnefelt et A. Thell, Flavocetraria cucullata, F. nivalis, Cetraria islandica, Cladonia arbuscula, С. rangiferina, C. stygia, C. uncialis (ПП до 90%), доля мхов (Racomitrium lanuginosum, Aulacomnium turgidum, Dicranum fuscescens, Ptilidium ciliare (L.) Hampe) незначительна (ПП до 10%), толщина покрова 2–3.5 см.
Пятнистые травяно-мохово-лишайниковые тундры характеризуются различной долей участия кустарничка Salix polaris (ПП 5–30%), среди видов трав Arctophila fulva, Saxifraga rivularis, Calamagrostis stricta, Luzula confusa, Juncus triglumis (ПП 1–15%). В структуре напочвенного покрова доминируют мхи Aulacomnium turgidum, Racomitrium lanuginosum, Dicranum elongatum (ПП 50–70%), лишайники представлены Alectoria ochroleuca, Gowardia nigricans, Flavocetraria nivalis, Cetrariella delisei, Cetraria nigricans Nyl., Cladonia uncialis, C. arbuscula, С. rangiferina, C. stygia, Thamnolia vermicularis (ПП 30–50%), толщина покрова 2.5–4.5 см.
В сообществах бугристых болот произрастают Salix polaris, Vaccinium vitis-idaea (ПП кустарничков 20%); из трав представлены Роа arctica, Carex brunnescens (Pers.) Poir., Rubus chamaemorus, Stellaria peduncularis Bunge, Draba nivalis Lilj., Luzula wahlenbergii Rupr., Senecio atropurpureus (Ledeb.) B. Fedtsch. (ПП 25%). В напочвенном покрове доминируют мхи Polytrichum strictum, Hylocomium splendens, Aulacomnium turgidum, А. palustre (Hedw.) Schwägr., Dicranum elongatum (ПП до 70%), доля лишайников Alectoria ochroleuca, Gowardia nigricans, Cladonia rangiferina, С. arbuscula, C. stygia, C. amaurocraea, Flavocetraria cucullata, Cetraria islandica, Cetrariella delisei, C. fastigiata (Delise ex Nyl.) Kärnefelt et A. Thell, Sphaerophorus globosus, невелика (ПП до 25%), толщина покрова 4 см.
На обследованных территориях высоко разнообразие сообществ с доминированием лишайников, чаще всего это открытые местообитания, с куртинным кустарниковым ярусом и разной степенью заболоченности. Покрытие модельных видов в структуре напочвенного покрова обследованных территорий не превышало 25%.
Объектом исследований послужили ценные кормовые виды кустистых лишайников – Cladonia arbuscula (Wallr.) Flot., C. rangiferina (L.) F. H. Wigg., C. stellaris (Opiz) Pouzar & Vӗzda, C. stygia (Fr.) Ruoss. Образцы лишайников модельных видов отбирали в растительных сообществах тундровой зоны полуостровов Ямал и Гыдан (рис. 1).
Рис. 1.
Карта-схема ботанико-географического районирования территории и точек отбора образцов: I – южные субарктические тундры, II – северные субарктические тундры, III – арктические тундры. Fig. 1. The scheme of tundra subzones and sampling points: I – shrub tundra, II –typical tundra, III – arctic tundra.

При проведении исследований в течение вегетационных сезонов 2010–2014 гг. в 11 точках на территории полуострова Ямал и западного побережья полуострова Гыдан сделано 50 геоботанических описаний тундровых сообществ на участках площадью 100 м2, а также 200 описаний мохово-лишайникового яруса (на площадках 25 × 25 см). В каждой подзоне были описаны фитоценозы с различной вертикальной и горизонтальной структурой.
Основные параметры структуры сообществ были включены в анализ для определения биотических факторов среды, оказывающих благоприятное влияние на рост лишайников в условиях интенсивных пастбищных нагрузок.
В ходе камеральных работ было промерено около 1500 лишайниковых подециев модельных видов. Подеции измеряли в предельно увлажненном состоянии. С помощью электронного штангенциркуля (Garant 41 2618_150, точность измерений 0.01 мм) определяли высоту отдельных колен в пределах таллома [16]. Длину подеция или его живой части определяли, как сумму длин колен, входящих в его состав, а регистрируемый возраст как количество этих колен.
В работе использованы два метода расчета прироста кустисто-разветвленных видов лишайников. По методу К.Н. Игошиной [17] прирост определяли как результат деления длины всего лишайникового подеция на количество ветвлений. В результате получены усредненные данные о скорости роста лишайников за длительные промежутки времени от 5 до 25 лет, что наиболее полно характеризует потенциал прироста лишайников на территории исследования. С помощью метода В.Н. Андреева [18] определяли прирост живой части (отличается от отмершей части по цвету и плотности таллома), который может быть использован для оценки продукционного потенциала и охватывает наиболее близкие к моменту исследования сроки (6–13 лет). Этот способ наиболее признан в мировом сообществе [19, 20]; его объективность подтверждена современными изотопными методами [21].
Выпас оленей меняет структуру напочвенного покрова, а, следовательно, влияет на прирост и высотно-возрастные параметры подециев [7, 22]. Для оценки состояния лишайников на территориях, используемых для выпаса, а также опосредованной оценки пастбищной нагрузки и интенсивности стравливания и вытаптывания оленями лишайникового покрова был проведен расчет значений следующих параметров: общая длина и регистрируемый возраст подеция и его живой части.
Общая длина подеция имеет существенное значение при рассмотрении изменений в лишайниковом покрове, т. к. очень чутко реагирует на любые воздействия [18, 23].
Высота живой части – к росту способны только живые колена лишайников, чем больше длина живой части, тем выше скорость и потенциал роста [18] и именно этот показатель в сочетании с наличием/отсутствием мертвой части подециев служит индикатором состояния лишайников на пастбищах [22].
Регистрируемый возраст подеция позволяет определить средний возраст талломов в условиях ежегодного и ежесезонного пастбищного использования.
Регистрируемый возраст живой части позволяет определить, сколько лет в условиях тундровой зоны лишайниковые талломы способны к активному росту, фотосинтезу и продукции биомассы.
Определение толщины мохово-лишайникового покрова позволяет опосредованно оценить интенсивность пастбищного воздействия на растительный покров территории. В сильно нарушенных сообществах тундровой зоны высота напочвенного покрова незначительна (до 1 см), лишайниковые подеции сильно повреждены, отмершая часть практически отсутствует. В сообществах с меньшей нагрузкой толщина покрова более 2 см, доля отмершей части незначительна.
Статистическую оценку значимости различий прироста и высотно-возрастных параметров модельных видов лишайников в разных подзонах и типах тундровых сообществ проводили с помощью одно- и двухфакторного дисперсионного анализа. Вклад биотических факторов среды в изменчивость скорости роста рассчитывали в ходе построения моделей методами множественной (с использованием forward-сортировки) или одномерной регрессии. При проверке статистических гипотез использовали 5%-ный уровень значимости.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
На территории исследования скорость роста кустисто-разветвленных лишайников варьирует в широких пределах от 1.2 до 6.3 мм/год (табл. 1).
Таблица 1.
Высотно-возрастные параметры подециев кустисто-разветвленных лишайников рода Cladonia на пастбищах северного оленя в условиях разных подзон (п-ова Ямал и Гыдан) Table 1. Growth rate, height and age variability of shrubby fruticose lichen podetium (Cladonia ssp.) on domestic reindeer pastures in different subzones (Yamal and Gydan peninsulas)
| Природная зона Tundra subzones |
Cladonia arbuscula | Cladonia rangiferina | Cladonia stygia | Cladonia stellaris | Все виды All species |
|||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean ± SD (N) | min–max | Mean ± SD (N) |
min–max | Mean ± SD (N) |
min–max | Mean ± SD (N) |
min–max | Mean ± SD (N) |
min–max | |
| Прирост подеция, мм/год Podetium growth rate, mm/year |
||||||||||
| Южные субарктические тундры | 2.1 ± 0.3a | 1.7–3.0 | 2.1 ± 0.4a | 1.2–3.1 | 2.8 ± 1.0b | 1.3–6.3 | 2.0 ± 0.3a | 1.8–2.4 | 2.3 ± 0.6a | 1.2–6.3 |
| Southern (shrub) tundra | (274) | (196) | (136) | (92) | (698) | |||||
| Северные субарктические тундры (покров ~2 см) | 2.3 ± 0.5ab | 1.7–3.4 | 2.5 ± 0.4b | 1.9–3.4 | 3.6 ± 0.6c | 2.6–4.3 | N/A | N/A | 2.7 ± 0.7b | 1.7–4.3 |
| Northern (typical) tundra (lichen mat ~ 2 cm) | (141) | (80) | (33) | (254) | ||||||
| Северные субарктические тундры (покров ~1 см) | 2.2 ± 0.4a | 1.6–3.0 | 2.3 ± 0.3a | 1.7–2.6 | 2.8 ± 0.6b | 2.1–4.3 | N/A | N/A | 2.5 ± 0.5ab | 1.6–4.3 |
| Northern (typical) tundra (lichen mat ~ 1cm) | (203) | (117) | (153) | (473) | ||||||
| Арктические тундры | 2.2 ± 0.4a | 1.8–2.6 | 2.2 ± 0.7a | 1.6–3.2 | 3.1 ± 0.8cb | 2.4–3.9 | N/A | N/A | 2.5 ± 0.7ab | 1.6–3.9 |
| Arctic tundra | (56) | (42) | (44) | (142) | ||||||
| Прирост живой части подеция, мм/год Growth rate of the living part of podetium, mm/year |
||||||||||
| Южные субарктические тундры Southern (shrub) tundra |
1.8 ± 0.2a | 1.4–2.5 | 1.9 ± 0.3a | 0.9–2.6 | 1.9 ± 0.5ab | 1.2–2.9 | 1.8 ± 0.2a | 1.6–2.0 | 1.9 ± 0.3a | 0.9–2.9 |
| Северные субарктические тундры (покров ~ 2 см) Northern (typical) tundra (lichen mat ~ 2 cm) |
1.8 ± 0.4a | 1.4–2.8 | 2.0 ± 0.3a | 1.5–2.4 | 2.5 ± 0.5c | 1.7–3.3 | N/A | N/A | 2.0 ± 0.5a | 1.4–3.3 |
| Северные субарктические тундры (покров ~1 см) Northern (typical) tundra (lichen mat ~ 1 cm) |
1.7 ± 0.2a | 1.2–1.9 | 1.9 ± 0.2a | 1.5–2.2 | 2.1 ± 0.3bc | 1.6–2.7 | N/A | N/A | 1.9 ± 0.3a | 1.2–2.6 |
| Арктические тундры Arctic tundra |
1.7 ± 0.1a | 1.5–1.8 | 1.8 ± 0.3a | 1.4–2.1 | 2.3 ± 0.6abc | 1.8–2.9 | N/A | N/A | 1.9 ± 0.4a | 1.4–2.9 |
| Общая длина подеция, мм Height of podetium, mm |
||||||||||
| Южные субарктические тундры Southern (shrub) tundra |
28 ± 8a | 16–48 | 26 ± 8a | 9–51 | 35 ± 12 b | 9–57 | 30 ± 9ab | 23–43 | 29 ± 1a | 9–57 |
| Северные субарктические тундры (покров ~ 2 см) Northern (typical) tundra (lichen mat ~ 2 cm) |
29 ± 9a | 16–51 | 28 ± 7a | 19–44 | 49 ± 16c | 30–84 | N/A | N/A | 34 ± 14b | 16–84 |
| Северные субарктические тундры (покров ~1 см) Northern (typical) tundra (lichen mat ~ 1 cm) |
30 ± 5a | 23–39 | 33 ± 7b | 22–42 | 40 ± 16bc | 20–81 | N/A | N/A | 35 ± 11b | 20–81 |
| Арктические тундры Arctic tundra |
33 ± 6a | 24–37 | 38 ± 20b | 21–67 | 39 ± 8bc | 30–44 | N/A | N/A | 37 ± 12b | 21–67 |
| Длина живой части подеция, мм Height of the living part of podetium, mm |
||||||||||
| Южные субарктические тундры Southern (shrub) tundra |
18 ± 4a1 | 11–28 | 18 ± 5a | 7–29 | 16 ± 5a | 7–28 | 21 ± 5a | 14–26 | 18 ± 5a | 7–29 |
| Северные субарктические тундры (покров ~2 см) Northern (typical) tundra (lichen mat ~2 cm) |
14 ± 4b | 8–24 | 15 ± 3ab | 11–19 | 18 ± 5c | 12–31 | N/A | N/A | 16 ± 3b | 13–22 |
| Северные субарктические тундры (покров ~1 см) Northern (typical) tundra (lichen mat ~1 cm) |
14 ± 2b | 10–17 | 17 ± 3a | 11–24 | 17 ± 4ab | 13–24 | N/A | N/A | 16 ± 3b | 10–24 |
| Арктические тундры Arctic tundra |
17 ± 1a | 16–19 | 17 ± 4a | 13–22 | 16 ± 1ab | 11–17 | N/A | N/A | 17 ± 5b | 8–31 |
| Регистрируемый возраст подеция, лет Relative podetium age, years |
||||||||||
| Южные субарктические тундры Southern (shrub) tundra |
13 ± 2a | 9–17 | 12 ± 2a | 8–17 | 13 ± 4a | 6–18 | 14 ± 2a | 13–18 | 13 ± 3a | 6–18 |
| Северные субарктические тундры (покров ~2 см) Northern (typical) tundra (lichen mat ~2 cm) |
12 ± 2a | 10–15 | 11 ± 2a | 8–15 | 14 ± 4a | 11–21 | N/A | N/A | 12 ±3b | 8–21 |
| Северные субарктические тундры (покров ~ 1 см) Northern (typical) tundra (lichen mat ~1 cm) |
13 ± 1ab | 11–15 | 14 ± 3b | 10–21 | 14 ± 3ab | 9–19 | N/A | N/A | 14 ± 2b | 9–21 |
| Арктические тундры Arctic tundra |
15 ± 2b | 14–16 | 16 ± 3b | 14–21 | 12 ± 2ab | 11–14 | N/A | N/A | 15 ± 3b | 11–21 |
| Регистрируемый возраст живой части подеция, лет Relative age of the living part of podetium, years |
||||||||||
| Южные субарктические тундры Southern (shrub) tundra |
10 ± 1a | 8–13 | 9 ± 1a | 7–12 | 8 ± 1b | 6–10 | 12 ± 2c | 9–14 | 7 ± 1a | 5–10 |
| Северные субарктические тундры (покров ~2 см) Northern (typical) tundra (lichen mat ~2 cm) |
8 ± 1b | 6–10 | 8 ± 1ab | 6–10 | 8 ± 1b | 6–10 | N/A | N/A | 9 ± 2b | 6–14 |
| Северные субарктические тундры (покрова ~1 см) Northern (typical) tundra (lichen mat ~1cm) |
8 ± 1b | 7–10 | 9 ± 1ab | 7–12 | 8 ± 1b | 7–9 | N/A | N/A | 8 ± 1bc | 6–10 |
| Арктические тундры Arctic tundra |
10 ± 1a | 9–10 | 10 ± 1a | 8–11 | 7 ± 1b | 6–8 | N/A | N/A | 8 ± 1ac | 7–12 |
Примечание. (N) – количество измеренных подециев; 1 – значимо различающиеся значения параметров отмечены индексами, не содержащими одинаковых букв; N/A – нет данных. Note. (N) – number of measured podetia; 1 – significantly different parameter values are marked with different letter indices; N/A – data not available.
В таежной зоне Западно-Сибирской равнины, в сообществах близких к коренным, прирост лишайников значительно уменьшается с юга на север [24]. Сходную картину мы предполагали увидеть в тундровых сообществах полуостровов Ямал и Гыдан, учитывая существенную протяженность и выраженную дифференциацию природных и климатических условий на территории исследования. Однако закономерных изменений средних значений скорости роста модельных видов в зональном градиенте не выявлено (рис. 2). Более того, в сообществах южных субарктических тундр отмечены наименьшие значения скорости роста кустисто-разветвленных лишайников (p < 0.01), вопреки ожидаемым наибольшим (табл. 1). Прирост живой части подеция (табл. 1, рис. 2) на территории исследований варьирует в значительно меньших пределах: от 0.9 до 3.3 мм/год, подзональные различия средних значений данного параметра не выявлены (p = 0.14). Различия значений прироста подеция и его живой части (на 25%) свидетельствуют, что потенциально скорость роста лишайниковых талломов выше, чем характерная для последнего десятилетия.
Рис. 2.
Общий прирост (a) и прирост живой части (b) подециев лишайников рода Cladonia в тундровой зоне полуостровов Ямал и Гыдан: 1 – южные субарктические тундры, 2 – северные субарктические тундры, 3 – арктические тундры. По вертикали справа – скорость роста, мм/год. По вертикали слева – климатические параметры: c – сумма активных температур, °С; d – количество осадков, мм. Fig. 2. Growth rate (a) and growth rate of the living part (b) of lichen podetium (Cladonia ssp.) in tundra zone of Yamal and Gydan peninsula: 1 – shrub tundra, 2 –typical tundra, 3 – arctic tundra. Main Y-axis – growth rate, mm/year. The secondary Y‑a-xis – clamate parameters: c – sum of active temperatures, °С; d – precipitation, mm.

Длина подециев модельных видов в южных субарктических тундрах в среднем на 20% ниже, чем в северных субарктических и арктических (р = 0.01). Закономерных изменений длины живой части подециев в градиенте исследованных подзональных единиц не выявлено. Она является более высокой (р < 0.05) в южных субарктических и арктических тундрах (табл. 1).
Средние значения регистрируемого возраста подеция и его живой части незначительно варьируют в градиенте “юг–север” (табл. 1). Наибольший разброс значений высотно-возрастных параметров лишайниковых подециев выявлен в сообществах северных субарктических тундр, где исследованы участки с различной высотой мохово-лишайникового яруса и интенсивностью нагрузки.
Из числа изученных модельных видов у C. stellaris выявлены максимальные значения возраста живой части подеция, среди других изученных видов (табл. 1). Для C. stygia характерны максимальные значения скорости роста подеция, а также длины подециев.
На территории лесотундры Западной Сибири структура растительных сообществ оказывает существенное влияние на ростовые процессы у напочвенных лишайников [16]. Наиболее значимыми биотическими факторами, влияющими на прирост лишайниковых талломов в условиях тундровой зоны и подзоны лесотундры, являются высота и сомкнутость кустарникового яруса, а также структура и толщина напочвенного покрова [18, 25, 26]. Результатом многих десятилетий выпаса домашнего северного оленя стала трансформация видового состава и структуры сообществ, а также толщины лишайникового покрова [2]. Этот фактор может оказывать значительное влияние на скорость роста лишайников и варьирование данного параметра в изученных градиентах.
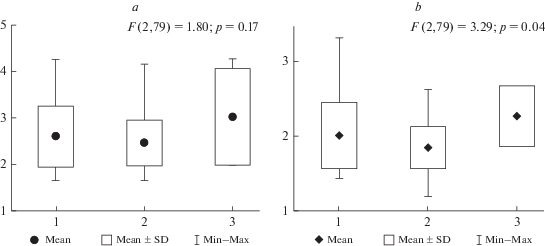
В полосе южных субарктических тундр прирост кустисто-разветвленных лишайников варьирует в пределах от 1.2 до 6.3 мм/год, скорость роста живой части подеция составляет 0.9–2.9 мм/год (табл. 2). Во всем многообразии изученных фитоценозов значимых различий среднего прироста лишайников не выявлено (p = 0.19). Однако при анализе рассматриваемого экологического ряда сообществ показано (табл. 2, рис. 3), что в кустарниковых тундрах модельные виды лишайников растут значимо быстрее (p < 0.05). Различия прироста живой части и подециев в целом в болотных фитоценозах и открытых тундровых сообществах (3–4%) статистически незначимы. Единственным биотическим фактором, оказывающим положительное влияние на скорость роста модельных видов лишайников в сообществах южных тундр Ямала и Гыдана, является сомкнутость кустарников, объясняющая около 20% дисперсии прироста (p < 0.01). Регистрируемый возраст подеция и его живой части в рассматриваемом экологическом ряду не меняются (табл. 2).
Таблица 2.
Высотно-возрастные параметры подециев кустисто-разветвленных лишайников рода Cladonia в разных типах сообществ южных субарктических тундр Table 2. Growth rate, height and age variability of shrubby fruticose lichen podetium (Cladonia ssp.) in southern (shrub) tundra communities
| Вид/Экологический ряд сообществ Species /Types of plant communities |
Cladonia arbuscula | Cladonia rangiferina | Cladonia stygia | Cladonia stellaris | Все виды All species |
|||||
|---|---|---|---|---|---|---|---|---|---|---|
| Mean ± SD (N) | min–max | Mean ± SD (N) | min–max | Mean ± SD (N) | min–max | Mean ± SD (N) | min–max | Mean ± SD (N) | min–max | |
| Прирост подеция, мм/год Podetium growth rate, mm/year |
||||||||||
| Кустарниковые тундры | 2.2 ± 0.4a | 1.8–3.0 | 2.3 ± 0.3a | 1.7–3.1 | 2.9 ± 1.1c | 1.3–6.3 | 2.1 ± 0.3a | 1.8–2.4 | 2.4 ± 0.7a | 1.3–6.3 |
| Shrub tundra | (113) | (98) | (56) | (48) | (315) | |||||
| Открытые тундровые сообщества | 2.0 ± 0.4b | 1.2–2.4 | 1.9 ± 0.2b | 2.3–2.7 | 2.5 ± 0.3bc | 1.7–2.3 | 1.8 ± 0.0ab | 1.8–1.9 | 2.0 ± 0.4b | 1.2–2.7 |
| Exposed tundra | (98) | (47) | (38) | (44) | (227) | |||||
| Бугристые болота | 2.0 ± 0.1b | 1.8–2.1 | 2.1 ± 0.2ab | 1.9–2.3 | 2.1 ± 0.3b | 1.8–2.3 | N/A | N/A | 2.1 ± 0.2b | 1.8–2.3 |
| Hummock bogs | (63) | (51) | (42) | (156) | ||||||
| Прирост живой части подеция, мм/год Growth rate of the living part of podetium, mm/year |
||||||||||
| Кустарниковые тундры Shrub tundra |
1.9 ± 0.3a | 1.4–2.5 | 1.9 ± 0.3a | 1.6–2.6 | 2.1 ± 0.5a | 1.2–2.9 | 1.8 ± 0.2a | 1.6–2.0 | 2.0 ± 0.4a | 1.2–2.9 |
| Открытые тундровые сообщества Exposed tundra |
1.8 ± 0.5a | 0.9–2.3 | 1.7 ± 0.2b | 1.9–2.1 | 2.0 ± 0.1a | 1.6–2.1 | 1.7 ± 0.0a | 1.7–1.8 | 1.8 ± 0.3b | 0.9–2.3 |
| Бугристые болота Hummock bogs |
1.7 ± 0.1a | 1.5–1.8 | 1.8 ± 0.1ab | 1.7–2.0 | 1.4 ± 0.2a | 1.2–1.6 | N/A | N/A | 1.7 ± 0.2b | 1.2–2.0 |
| Общая длина подеция, мм Height of podetium, mm |
||||||||||
| Кустарниковые тундры Shrub tundra |
28 ± 8a | 20–48 | 32 ± 8a | 18–51 | 35 ± 13a | 9–57 | 32 ± 10a | 25–43 | 32 ± 10a | 9–57 |
| Открытые тундровые сообщества Exposed tundra |
21 ± 8b | 9–33 | 23 ± 7b | 32–46 | 39 ± 9ab | 16–39 | 23 ± 0b | 22–23 | 24 ± 9b | 9–46 |
| Бугристые болота Hummock bogs |
26 ± 4ab | 21–31 | 26 ± 5ab | 21–35 | 27 ± 8a | 19–32 | N/A | N/A | 26 ± 5ab | 19–35 |
| Длина живой части подеция, мм Height of the living part of podetium, mm |
||||||||||
| Кустарниковые тундры Shrub tundra |
19 ± 4a | 11–28 | 20 ± 5a | 13–29 | 17 ± 6a | 7–28 | 22 ± 6a | 14–26 | 19 ± 5a | 7–29 |
| Открытые тундровые сообщества Exposed tundra |
15 ± 6b | 7–22 | 17 ± 3a | 15–19 | 16 ± 3ab | 14–22 | 21 ± 0a | 20–21 | 16 ± 4a | 7–22 |
| Бугристые болота Hummock bogs |
17 ± 2ab | 14–21 | 19 ± 4a | 15–24 | 11 ± 2b | 9–14 | N/A | N/A | 17 ± 4a | 9–24 |
| Регистрируемый возраст подеция, лет Relative podetium age, years |
||||||||||
| Кустарниковые тундры Shrub tundra |
13 ± 2a | 11–17 | 12 ± 4a | 10–17 | 14 ± 2a | 6–18 | 5 ± 2a | 13–18 | 13 ± 3a | 6–18 |
| Открытые тундровые сообщества Exposed tundra |
10 ± 3b | 8–15 | 12 ± 2ab | 14–16 | 15 ± 2ab | 9–17 | 13 ± 0ab | 13–14 | 12 ± 3a | 8–17 |
| Бугристые болота Hummock bogs |
13 ± 1a | 12–15 | 12 ± 2a | 11–15 | 13 ± 2a | 11–14 | N/A | N/A | 13 ± 1a | 11–15 |
| Регистрируемый возраст живой части подеция, лет Relative age of the living part of podetium, years |
||||||||||
| Кустарниковые тундры Shrub tundra |
9 ± 1a | 8–13 | 10 ± 2a | 7–12 | 8 ± 1a | 6–10 | 12 ± 3b | 9–14 | 9 ± 2a | 6–14 |
| Открытые тундровые сообщества Exposed tundra |
8 ± 1a | 7–10 | 9 ± 2a | 7–10 | 9 ± 1a | 8–12 | 12 ± 0b | 12–13 | 9 ± 1a | 7–12 |
| Бугристые болота Hummock bogs |
10 ± 1a | 9–11 | 10 ± 1a | 9–12 | 8 ± 1a | 7–8 | N/A | N/A | 10 ± 1a | 7–12 |
Примечание. (N) – количество измеренных подециев; 1 – значимо различающиеся значения параметров отмечены индексами, не содержащими одинаковых букв; N/A – нет данных. Note. (N) – number of measured podetia; 1 – significantly different parameter values are marked with different letter indices; N/A – data not available.
Рис. 3.
Прирост кустисто-разветвленных лишайников (a – всего подеция, b – прирост живой части подеция) в экологическом ряду сообществ южных субарктических тундр: 1 – кустарниковые тундры, 2 – открытые тундры, 3 – бугристые болота. По вертикали – скорость роста, мм/год. Fig. 3. Growth rate of shrubby fruticose lichens (a – podetium growth rate, b – growth rate of the living part of podetium) in different types of shrub tundra subzone plant communities: 1 – shrub tundra, 2 – exposed tundra, 3 – hummock bogs. Y-axis – growth rate, mm/year.
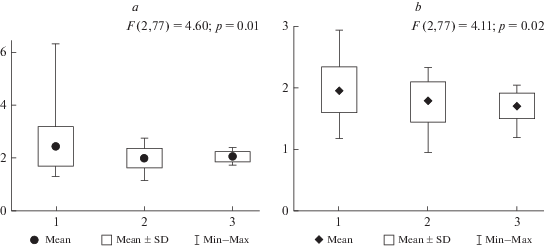
В условиях пастбищной деградации растительных сообществ полосы южных субарктических тундр практически во всех единицах экологического ряда для С. stygia характерны более высокие значения прироста и общей высоты подециев. Однако значимыми являются только различия прироста в кустарниковых тундрах (р = 0.03). Для C. stellaris характерны значимо более высокие показатели регистрируемого возраста живой части подеция (р < 0.05). Различия высотно-возрастных параметров C. arbuscula и C. rangiferina в пределах экологического ряда незначительны.
В полосе северных субарктических тундр прирост кустисто-разветвленных лишайников варьирует в пределах от 1.6 до 4.3 мм/год, прирост живой части подеция – от 1.2 до 3.3 мм/год (табл. 3). Значимых различий в скорости роста подециев во всем разнообразии изученных фитоценозов и в рассматриваемом экологическом ряду (рис. 4) не выявлено. Несколько более высокие значения (р < 0.05) прироста живой части подециев отмечены в сообществах бугристых болот по сравнению с кустарниковыми и открытыми тундрами.
Таблица 3.
Высотно-возрастные параметры подециев кустисто-разветвленных лишайников рода Cladonia в разных типах сообществ северных субарктических тундр Table 3. Growth rate, height and age variability of shrubby fruticose lichen podetium (Cladonia ssp.) in northern (typical) tundra communities
| Вид/Экологический ряд сообществ Species/Types of plant communities |
Cladonia arbuscula | Cladonia rangiferina | Cladonia stygia | Все виды All species |
||||
|---|---|---|---|---|---|---|---|---|
| Mean ± SD (N) |
min–max | Mean ± SD (N) |
min–max | Mean ± SD (N) |
min–max | Mean ± SD (N) |
min–max | |
| Прирост подеция, мм/год Podetium growth rate, mm/year |
||||||||
| Кустарниковые тундры | 2.2 ± 0.4a | 1.7–3.4 | 2.4 ± 0.4a | 1.7–3.4 | 3.4 ± 0.6b | 2.6–4.3 | 2.6 ± 0.7a | 1.7–4.3 |
| Shrub tundra | (129) | (71) | (52) | (252) | ||||
| Открытые тундровые сообщества | 2.3 ± 0.4a | 1.6–3.0 | 2.4 ± 0.3ab | 2.0–3.0 | 2.8 ± 0.6b | 2.2–4.1 | 2.5 ± 0.5a | 1.6–4.1 |
| Exposed tundra | (215) | (95) | (96) | (406) | ||||
| Бугристые болота | N/A | N/A | 2.3 ± 0.1ab | 2.2–2.4 | 3.3 ± 1.1b | 2.1–4.3 | 3.0 ± 1.0a | 2.1–4.3 |
| Hummock bogs | (31) | (38) | (69) | |||||
| Прирост живой части подеция, мм/год Growth rate of the living part of podetium, mm/year |
||||||||
| Кустарниковые тундры Shrub tundra |
1.8 ± 0.4a | 1.4–2.8 | 1.9 ± 0.2ab | 1.5–2.4 | 2.4 ± 0.5b | 1.7–3.3 | 2.0 ± 0.4b | 1.4–3.3 |
| Открытые тундровые сообщества Exposed tundra |
1.7 ± 0.2a | 1.2–2.0 | 1.9 ± 0.3b | 1.5–2.4 | 2.0 ± 0.2b | 1.6–2.3 | 1.8 ± 0.3a | 1.2–2.4 |
| Бугристые болота Hummock bogs |
N/A | N/A | 2.0 ± 0.2b | 1.9–2.1 | 2.4 ± 0.4b | 1.9–2.6 | 2.3 ± 0.4b | 1.9–2.6 |
| Длина подеция, мм Height of podetium, mm |
||||||||
| Кустарниковые тундры Shrub tundra |
28 ± 9a | 16–51 | 30 ± 8a | 19–44 | 48 ± 16b | 30–84 | 34 ± 14a | 16–84 |
| Открытые тундровые сообщества Exposed tundra |
31 ± 6a | 22–40 | 33 ± 8a | 19–42 | 36 ± 9ab | 20–45 | 32 ± 7a | 19–45 |
| Бугристые болота Hummock bogs |
N/A | N/A | 23 ± 7a | 18–28 | 54 ± 27ab | 26–81 | 46 ± 27a | 23–81 |
| Длина живой части подеция, мм Height of the living part of podetium, mm |
||||||||
| Кустарниковые тундры Shrub tundra |
14 ± 4a | 10–24 | 16 ± 3ab | 11–21 | 19 ± 5c | 12–31 | 16 ± 4a | 10–31 |
| Открытые тундровые сообщества Exposed tundra |
14 ± 3a | 8–17 | 17 ± 4b | 11–24 | 15 ± 2ac | 13–17 | 15 ± 3a | 8–24 |
| Бугристые болота Hummock bogs |
N/A | N/A | 15 ± 0b | 14–15 | 19 ± 3bc | 17–22 | 18 ± 3a | 15–22 |
| Регистрируемый возраст подеция, лет Relative podetium age, years |
||||||||
| Кустарниковые тундры Shrub tundra |
12 ± 2a | 10–15 | 12 ± 2a | 8–15 | 14 ± 3a | 11–21 | 13 ± 3a | 8–21 |
| Открытые тундровые сообщества Exposed tundra |
13 ± 2a | 10–15 | 14 ± 4a | 8–21 | 13 ± 3a | 9–16 | 13 ± 2a | 8–21 |
| Бугристые болота Hummock bogs |
N/A | N/A | 10 ± 3a | 8–12 | 16 ± 3a | 13–19 | 14 ± 4a | 10–19 |
| Регистрируемый возраст живой части подеция, лет Relative age of the living part of podetium, years |
||||||||
| Кустарниковые тундры Shrub tundra |
8 ± 1a | 6–10 | 8 ± 1a | 6–10 | 8 ± 1a | 6–10 | 8 ± 1a | 7–9 |
| Открытые тундровые сообщества Exposed tundra |
8 ± 1a | 6–10 | 9 ± 2a | 7–12 | 7 ± 1a | 6–8 | 8 ± 1a | 6–10 |
| Бугристые болота Hummock bogs |
N/A | N/A | 8 ± 1a | 7–8 | 8 ± 1a | 7–9 | 8 ± 1a | 6–12 |
Примечание. (N) – количество измеренных подециев; 1 – значимо различающиеся значения параметров отмечены индексами, не содержащими одинаковых букв; N/A – нет данных. Note. (N) – number of measured podetia; 1 – significantly different parameter values are marked with different letter indices;N/A – data not available.
Рис. 4.
Прирост кустисто-разветвленных лишайников (a – всего подеция, b – прирост живой части подеция) в экологическом ряду сообществ северных субарктических тундр: 1 – кустарниковые тундры, 2 – открытые тундры, 3 – бугристые болота. По вертикали – скорость роста, мм/год. Fig. 4. Growth rate of shrubby fruticose lichens (a – podetium growth rate, b – growth rate of the living part of podetium) in different types of typical tundra subzone plant communities in: 1 – shrub tundra, 2 – exposed tundra, 3 – hummock bogs. Y-axis – growth rate, mm/year.
Скорость роста живой части подециев модельных видов различается в ряду: болотные комплексы – кустарниковые тундры – открытые тундровые сообщества. В последних она на 10–20% ниже (р < 0.05). Остальные высотно-возрастные параметры в исследованном экологическом ряду значимо не различаются (табл. 3).
В условиях пастбищной эксплуатации в сообществах северных субарктических тундр выявлен единственный значимый фактор биотической среды, оказывающий влияние на прирост лишайников – толщина мохово-лишайникового яруса. При этом с увеличением пастбищных нагрузок вклад этого фактора в варьирование прироста возрастает с 6 до 28% (p < 0.01).
Из числа модельных видов наибольшие значения изученных параметров (за исключением возрастных) выявлены у C. stygia в кустарниковых тундрах (p < 0.01), где она растет в среднем на 30–50% быстрее, чем другие виды (табл. 3). Общая скорость роста C. arbuscula и C. rangiferina в изученных экологических группах сообществ значимо не различается. Скорость роста живой части подециев этих видов имеет значимое различие в открытых тундровых сообществах, но оно не превышает 12%.
В подзоне арктических тундр прирост кустисто-разветвленных лишайников варьирует в диапазоне 1.6–4.1 мм/год, живой части 1.4–2.9 мм/год (табл. 4). Более высокие значения общего прироста и прироста живой части подециев (на 45–85%) отмечены в сообществах бугристых болот (p < 0.01). В лишайниковых и пятнистых тундрах значимых различий скорости роста лишайников не выявлено (рис. 5).
Таблица 4.
Высотно-возрастные параметры подециев кустисто-разветвленных лишайников рода Cladonia в разных типах сообществ арктических тундр Table 4. Growth rate, height and age variability of shrubby fruticose lichen podetium (Cladonia ssp.) in arctic tundra communities
| Вид/Экологический ряд сообществ Species/Types of plant communities |
Cladonia arbuscula | Cladonia rangiferina | Cladonia stygia | Все виды All species |
||||
|---|---|---|---|---|---|---|---|---|
| Mean ± SD (N) |
min–max | Mean ± SD (N) |
min–max | Mean ± SD (N) |
min–max | Mean ± SD (N) |
min–max | |
| Прирост подеция, мм/год Podetium growth rate, mm/year |
||||||||
| Кустарничково-лишайниковые тундры | 2.2 ± 0.1a | 2.2–2.2 | 1.9 ± 0.3a | 1.6–2.2 | 2.4 ± 0.6a | 1.9–3.4 | 2.1 ± 0.3a | 1.6–2.4 |
| Dwarf shrub-lichen tundra | (27) | (14) | (15) | (56) | ||||
| Пятнистые травяно-мохово-лишайниковые тундрыSpotty grass-moss-lichen tundra | 2.2 ± 0.6a(29) | 1.8–2.6 | 3.2 ± 0.0a(28) | 3.1–3.2 | 3.1 ± 0.5a(17) | 2.6–4.1 | 2.7 ± 0.7a(74) | 1.8–3.2 |
| Бугристые болота | N/A | N/A | N/A | N/A | 3.9 ± 0.3a | 3.7–4.1 | 3.9 ± 0.3b | 3.7–4.1 |
| Hummock bogs | (12) | (12) | ||||||
| Прирост живой части подеция, мм/год Growth rate of the living part of podetium, mm/year |
||||||||
| Кустарничково-лишайниковые тундры Dwarf shrub-lichen tundra |
1.8 ± 0.1a | 1.7–1.8 | 1.6 ± 0.3a | 1.4–1.9 | 1.8 ± 0.4a | 1.3–2.4 | 1.7 ± 0.2a | 1.4–1.9 |
| Пятнистые травяно-мохово-лишайниковые тундры Spotty grass-moss-lichen tundra |
1.6 ± 0.2a | 1.5–1.7 | 2.1 ± 0.0a | 2.0–2.1 | 2.1 ± 0.5a | 1.4–3.2 | 1.9 ± 0.3a | 1.5–2.1 |
| Бугристые болота Hummock bogs |
N/A | N/A | N/A | N/A | 2.9 ± 0.5a | 2.5–3.2 | 2.9 ± 0.5b | 2.5–3.2 |
| Общая длина подеция, мм Height of podetium, mm |
||||||||
| Кустарничково-лишайниковые тундры Dwarf shrub-lichen tundra |
36 ± 1a | 35–37 | 29 ± 8a | 21–37 | 30 ± 15a | 15–58 | 31 ± 6a | 21–37 |
| Пятнистые травяно-мохово-лишайниковые тундры Spotty grass-moss-lichen tundra |
30 ± 8a | 24–36 | 67 ± 0a | 66–67 | 44 ± 12a | 24–69 | 43 ± 18a | 24–67 |
| Бугристые болота Hummock bogs |
N/A | N/A | N/A | N/A | 43 ± 14a | 33–53 | 43 ± 14a | 33–53 |
| Длина живой части подеция, мм Height of the living part of podetium, mm |
||||||||
| Кустарничково-лишайниковые тундры Dwarf shrub-lichen tundra |
18 ± 1a | 17–19 | 16 ± 5a | 13–22 | 14 ± 6a | 7–24 | 17 ± 3a | 13–22 |
| Пятнистые травяно-мохово-лишайниковые тундры Spotty grass-moss-lichen tundra |
15 ± 1a | 15–16 | 19 ± 0a | 18–19 | 16 ± 5a | 7–23 | 16 ± 2a | 15–19 |
| Бугристые болота Hummock bogs |
N/A | N/A | N/A | N/A | 17 ± 1a | 16–18 | 17 ± 1a | 16–18 |
| Регистрируемый возраст подеция, лет Relative lichen podetium age, year |
||||||||
| Кустарничково-лишайниковые тундры Dwarf shrub-lichen tundra |
16 ± 0a | 16–17 | 15 ± 2a | 14–17 | 12 ± 3ab | 7–17 | 15 ± 2a | 12–17 |
| Пятнистые травяно-мохово-лишайниковые тундры Spotty grass-moss-lichen tundra |
14 ± 0b | 14–15 | 21 ± 0a | 21–22 | 14 ± 3b | 9–19 | 16 ± 4a | 14–21 |
| Бугристые болота Hummock bogs |
N/A | N/A | N/A | N/A | 11 ± 3b | 9–13 | 11 ± 3a | 9–13 |
| Регистрируемый возраст живой части подеция, лет Relative age of the living part of podetium, year |
||||||||
| Кустарничково-лишайниковые тундры Dwarf shrub-lichen tundra |
10 ± 0a | 10–11 | 10 ± 2a | 8–11 | 8 ± 2a | 5–10 | 10 ± 1a | 8–11 |
| Пятнистые травяно-мохово-лишайниковые тундры Spotty grass-moss-lichen tundra |
9 ± 1a | 9–10 | 9 ± 0a | 9–10 | 7 ± 2a | 5–9 | 9 ± 1a | 7–10 |
| Бугристые болота Hummock bogs |
N/A | N/A | N/A | N/A | 6 ± 1a | 5–7 | 6 ± 1b | 5–7 |
Примечание. (N) – количество измеренных подециев; 1 – значимо различающиеся значения параметров отмечены индексами, не содержащими одинаковых букв; N/A – нет данных. Note. (N) – number of measured podetia; 1 – significantly different parameter values are marked with different letter indices;N/A – data not available.
Рис. 5.
Прирост кустисто-разветвленных лишайников (a – всего подеция, b – прирост живой части подеция) в экологическом ряду сообществ арктических тундр: 1 – кустарничково-лишайниковые тундры, 2 – пятнистые травяно-мохво-лишайниковые тундры, 3 – бугристые болота. По вертикали – скорость роста, мм/год. Fig. 5. Growth rate of shrubby fruticose lichens (a – podetium growth rate, b – growth rate of the living part of podetium) in different types of arctic tundra subzone plant communities: 1 – dwarf shrub-lichen tundra, 2 – spotty grass-moss-lichen tundra, 3 – hummock bogs. Y-axis – growth rate, mm/year.

Толщина и структура напочвенного покрова значимо влияет на прирост кустисто-разветвленных лишайников. Толщиной лишайникового покрова объясняется 50% дисперсии прироста (p = 0.01), долей мхов в структуре напочвенного покрова – 45% (p = 0.01). Однако незначительная доля отмерших частей подециев свидетельствует о высокой уязвимости лишайников в условиях арктических тундр Ямала и Гыдана.
Значения высотно-возрастных параметров лишайниковых подециев у разных модельных видов не имеют значимых различий в зависимости от условий местообитания (табл. 4).
Таким образом, в современных условиях на пастбищных территориях полуостровов Ямал и Гыдан не выявлено существенного снижения прироста кустисто-разветвленных видов лишайников рода Cladonia с юга на север. Эту закономерность можно объяснить двумя основными причинами: 1) гидротермические условия произрастания лишайников во всей тундровой зоне предельно суровые и их изменение с юга на север не оказывает значимого влияния; 2) мощное антропогенное воздействие перекрывает влияние климатических факторов и нивелирует подзональные различия прироста лишайников. Значимое снижение прироста лишайников в полосе южных субарктических тундр, относительно северных субарктических и арктических тундр, можно объяснить интенсивным использованием территории, не только как весенних, летних и осенних пастбищ, но и как прогонных путей к северным районам. Это может свидетельствовать о высокой значимости не только стравливания, но и вытаптывания, как факторов, влияющих на скорость роста лишайников кормовых видов.
Вывод о высокой антропогенной нагрузке на сообщества южных субарктических тундр подтверждает также результат анализа изменения длины подециев модельных видов, показывающий значимое снижение величины данного параметра в полосе кустарниковых тундр. В тоже время в арктических тундрах, для которых характерны наиболее суровые природно-климатические условия, отмечена довольно значительная длина подециев и их живой части. Этот факт может свидетельствовать о меньшей пастбищной эксплуатации данных территорий по сравнению с субарктическими тундрами. Кроме того, наибольшие средние значения высоты живой части подеция в сообществах южных субарктических тундр свидетельствуют о более высоком потенциале роста и продуктивности лишайников на этой территории, который не может быть реализован в условиях антропогенного воздействия.
Для сообществ северных субарктических тундр характерна различная интенсивность пастбищной нагрузки и высокая вариабельность показателей толщины мохово-лишайникового покрова, что является причиной наибольшего разброса значений высотно-возрастных параметров лишайниковых подециев. Кроме того, лишайники этих фитоценозов находятся на стадии наибольшей продуктивности [18], что подтверждают наименьшие значения длины живой части подеция и наибольшие значения общего прироста.
Наиболее стабильный показатель состояния лишайниковых подециев – это регистрируемый возраст живой части подеция, не зависящий ни от природно-климатических условий, ни от степени антропогенного воздействия, ни от видовой принадлежности. Продолжительность жизни лишайниковых талломов обусловлена внутренними физиологическими процессами взаимодействия фото- и микобионта [27, 28].
В фитоценотическом градиенте наибольшая скорость роста подеция или живой части модельных видов лишайников выявлена в сообществах кустарниковых тундр (южные субарктические тундры) и бугристых болот (северные субарктические и арктические тундры). Выраженный кустарниковый ярус в полосе южных субарктических тундр выполняет защитную функцию, снижая иссушающее влияние ветров на напочвенный покров и лишайники, что оказывает положительное влияние на прирост подециев. Кроме того зеленые корма и листва кустарников в летнее время более предпочтительны для оленей, что снижает отрицательное влияние стравливания лишайникового покрова. Однако в сообществах северных субарктических тундр низкорослые кусты [2] образуют фрагментарный ярус, который не оказывает положительного влияния на состояние напочвенного покрова и ростовые процессы лишайников. Болотные фитоценозы подвержены меньшей пастбищной эксплуатации, а сомкнутый, имеющий большую толщину моховой покров поддерживает значительную степень увлажненности талломов лишайников. В результате лишайниковые подеции лучше сохраняются при механических нагрузках и достигают довольно высоких значений общей скорости роста или скорости роста живой части особенно в подзоне арктических тундр.
В сообществах северных субарктических и арктических тундр важным биотическим фактором среды, положительно влияющим на ростовые процессы лишайников, является хорошо сформированный мохово-лишайниковый покров с мощным отмершим слоем, который поддерживает подходящие для роста лишайников микроклиматические условия.
У C. stygia выявлены максимальные значения скорости роста подеция и его живой части, а также длины подециев, что характеризует данный вид как наиболее устойчивый к пастбищной нагрузке. Наименьший регистрируемый возраст живой части подеция подтверждает наше предположение о наибольшем восстановительном потенциале этого вида, т.к. он достигает наибольших значений прироста за более короткий период жизни подеция. В болотных фитоценозах и кустарниковых тундрах данный вид растет быстрее, т.к. наиболее требователен к условиям увлажнения [29].
C. stellaris в 30-е годы XX-го столетия считали одним из доминирующих видов на полуострове в составе лишайниковых тундр [30], однако в настоящее время он практически исчез в результате антропогенных нагрузок [9, 10, 30–33]. Прирост у C. stellaris в современных условиях не отличается от прироста C. arbuscula и С. rangiferina и меньше чем у C. stygia. Однако у C. stellaris выявлены достаточно большие значения высоты и возраста живой части подеция, что может свидетельствовать о высоком потенциале роста и продуктивности, а значит C. stellaris может занимать доминирующие позиции в структуре напочвенного покрова при отсутствии механического воздействия.
ВЫВОДЫ
1. В современных условиях на территории полуостровов Ямал и Гыдан высотно-возрастные параметры кустисто-разветвленных лишайников рода Cladonia варьируют в широких пределах.
2. Не выявлено значимого снижения прироста лишайников при переходе от южных субарктических тундр к арктическим, несмотря на изменения гидротермических условий с юга на север. Минимальные значения прироста выявлены в южных субарктических тундрах.
3. Наибольшие величины прироста лишайниковых подециев выявлены в кустарниковых тундрах (южные субарктические тундры) и сообществах болот (северные субарктические тундры и арктические тундры).
4. Значимыми биотическими факторами среды, влияющими на прирост лишайников, являются сомкнутость кустарникового яруса (южные субарктические тундры), толщина мохово-лишайникового покрова (северные субарктические и арктические тундры) и доля мхов в структуре напочвенного покрова (арктические тундры).
5. Максимальный прирост подеция и его живой части, а также длина подеция в целом практически на всей территории исследования отмечены у C. stygia. Можно констатировать, что данный вид характеризуется наибольшей устойчивостью к пастбищным нагрузкам и обладает наибольшим восстановительным потенциалом.
6. Достаточно большие значения высоты живой части, регистрируемого возраста подеция и его живой части у C. stellaris, свидетельствуют о высоком потенциале роста и продукции вида, который в условиях современной пастбищной нагрузки не реализован.
Список литературы
Olofsson J., Moen J., Oksanen L. 2002. Effects of herbivory on competition intensity in two arctic-alpine tundra communities with different productivity – Oikos. 96(2): 265–272. https://doi.org/10.1034/j.1600-0706.2002.960208.x
Полуостров Ямал: растительный покров. 2006. Тюмень. 360 с. https://ipae.uran.ru/sites/default/files/publications/ipae/1054_2006_PoluostrovYamal_RastPokrov.pdf
Ahti T., Oksanen J. 1990. Epigeic lichen communities of taiga and tundra regions – Vegetatio. 86(1): 39–70. https://doi.org/10.1007/BF00045134
Olofsson J., Oksanen L. 2002. Role of litter decomposition for the increased primary production in areas heavily grazed by reindeer: a litterbag experiment – Oikos. 96(3): 507–515. https://doi.org/10.1034/j.1600-0706.2002.960312.x
van der Wal R., Brooker R., Cooper E., Langvatn R. 2001. Differential effects of reindeer on high Arctic lichens – J. Veg. Sci. 12(11): 705–710. https://doi.org/10.2307/3236911
Marozas V., Racinskas J., Bartkevicius E. 2007. Dynamics of ground vegetation after surface fires in hemiboreal Pinus sylvestris forests – Forest. Ecol. Manag. 250(1–2): 47–55. https://doi.org/10.1016/j.foreco.2007.03.008
Olofsson A., Danell O., Forslund P., Åhman B. 2011. Monitoring changes in lichen resources for range management purposes in reindeer husbandry – Ecol. Indic. 11(5): 1149–1159. https://doi.org/10.1016/j.ecolind.2010.12.015
Sveinbjörnsson B. 1987. Reindeer lichen productivity as a function of mat thickness. – Arctic Alpine Res. 19(4): 437–441. https://doi.org/10.2307/1551409
Herder M., Kytöviita M.-M., Niemela P. 2003. Growth of reindeer lichen and effects of reindeer grazing on ground cover vegetation in Scots pine forest and a subarctic heathland in Finnish Lapland – Ecography. 26(1): 3–12. https://doi.org/10.1034/j.1600-0587.2003.03211.x
Растительный покров и растительные ресурсы Полярного Урала. 2006. Екатеринбург. 796 с. https://ipae.uran.ru/sites/default/files/publications/ipae/0154_2006_MorozovaEtAl1.pdf https://ipae.uran.ru/sites/default/files/publications/ipae/0154_2006_MorozovaEtAl2_0.pdf
Golovatin M.G., Morozova L.M., Ektova S.N. 2012. Effect of reindeer overgrazing on vegetation and animals of tundra ecosystems of the Yamal peninsula – Czech Polar Report. 2(2): 80–91. https://doi.org/10.5817/CPR2012-2-8
Природа Ямала. 1995. Екатеринбург. 435 с. https://ipae.uran.ru/sites/default/files/publications/ipae/0305_1995_Природа_ЯмалаМал.pdf
Атлас Ямало-Ненецкого автономного округа (Карты). 2004. Омск. 303 с.
World Weather Information Service. 2020. http://worldweather.wmo.int/en/home.html.
Флора полуострова Ямал. Современное состояние и история формирования. 2013. СПб. 312 с.
Абдульманова С.Ю., Эктова С.Н. 2015. Ростовые процессы некоторых видов кустисто-разветвленных лишайников рода Cladonia (Сladoniaceae) в тундровых сообществах – Растительные ресурсы. 51(3): 344–356. https://www.binran.ru/files/journals/PlantRes/2015_51_3/PlantRes_2015_51_3_Abdulmanova_abstract_r.pdf
Игошина К.Н. 1939. Рост кормовых лишайников на приуральском Севере – Тр. НИИ поляр. земледелия, животноводства и промысл. хоз-ва. Сер. Оленеводство. 4: 7–28.
Андреев В.Н. 1954. Прирост кормовых лишайников и приемы его регулирования – Тр. Ботан. Ин-та. Сер. 3. Геоботаника. 9: 11–74.
Kärenlampi L. 1971. Studies on the relative growth rate of some fruticose lichens – Rep. Kevo Subarctic. 7: 33–39.
Sveinbjörnsson B. 1990. Reindeer lichen productivity: problems and possibilities – Rangifer. 10(3): 91–98. https://doi.org/10.7557/2.10.3.836
Holt E.A., Bench G. 2008. 14C/C measurements support Andreev’s internode method to determine lichen growth rates in Cladonia stygia (Fr.) Ruoss – The Lichenologist. 40(6): 559–565. https://doi.org/10.1017/S0024282908008062
Эктова С.Н. 2003. Изменение лишайникового покрова Заполярного Урала под воздействием выпаса оленей. – Научный вестник Ямало-Ненецкого автономного округа; Вып. 3. Биологические ресурсы Полярного Урала. Часть 2: 88–94. https://ipae.uran.ru/sites/default/files/publications/ipae/1038_2003_3%282%29sm.pdf
Boudreau S., Payette S. 2004. Caribou-induced changes in species dominance of lichen woodland: an analysis of plant remains – AM. J. BOT. 91(3): 422–429. https://doi.org/10.3732/ajb.91.3.422
Абдульманова С.Ю. 2013. Зависимость размерно-возрастных параметров покровообразующих лишайников от условий местообитания – В сб.: Экология: теория и практика: материалы конф. молодых ученых. Екатеринбург. С. 5–15. https://ipae.uran.ru/sites/default/files/publications/ipae/0227_2013_sbornikMKмал.pdf
Пристяжнюк С.А. 1998. Лишайники южных субарктических тундр Ямала и перевыпас – Сиб. экол. журн. 2: 197–200.
Абдульманова С.Ю. 2013. Особенности формирования годичного прироста кустистых лишайников на севере Западной Сибири – В сб.: Биоразнообразие экосистем Крайнего Севера: инвентаризация, мониторинг, охрана: материалы Всероссийской конф. Сыктывкар. С. 172–175. http://byrranga.ru/docs/242_244.pdf
Crittenden P.D. 1991. Ecological significance of necromass production in mat-forming lichens – The Lichenologist. 23(3): 323–331. https://doi.org/10.1017/S0024282991000464
Hammer S. 2000. Meristem growth dynamics and branching patterns in the Cladoniaceae – Am. J. Bot. 87(1): P. 33–47. https://doi.org/10.2307/2656683
Ahti T., Hyvönen S. 1985. Cladina stygia, a common, overlooked species of reindeer lichen – Ann. Bot. Fenn. 22(3): 223–229. https://www.jstor.org/stable/23725708?seq=1
Андреев В.Н. Игошина К.Н., Лесков А.И. 1935. Оленьи пастбища и растительный покров Полярного Приуралья – Сов. оленеводство. 5: 171–406.
Kershaw K.A. 1978. The Role of Lichens in Boreal Tundra Transition Areas – The Bryologist. 81(2): 294–306. https://doi.org/10.2307/3242190
Johnson E.A. 1981.Vegetation organization and dynamics of lichen woodland communities in the Northwest Territories, Canada – Ecology. 62(1): 200–215. https://doi.org/10.2307/1936682
Suominen O., Olofsson J. 2000. Impacts of semi-domesticated reindeer on structure of tundra and forest communities in Fennoscandia: a review – Ann. Zool. Fenn. 37: 233–249. http://www.sekj.org/PDF/anzf37/anzf37-233p.pdf
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы