

Растительные ресурсы, 2020, T. 56, № 4, стр. 335-350
Пространственное распределение запасов напочвенного покрова и лесной подстилки в фоновых и дефолиирующих сосновых лесах Кольского полуострова
И. В. Лянгузова 1, *, П. А. Примак 2, Е. Н. Волкова 1, Ф. С. Салихова 1
1 Ботанический институт им. В.Л. Комарова РАН
г. Санкт-Петербург, Россия
2 Санкт-Петербургский государственный университет
г. Санкт-Петербург, Россия
* E-mail: ILyanguzova@binran.ru
Поступила в редакцию 23.02.2020
После доработки 01.07.2020
Принята к публикации 27.08.2020
Аннотация
Приведены результаты исследования внутриценотической неоднородности почвенно-растительного покрова в средневозрастных сосновых лесах лишайниково-зеленомошной группы типов леса на фоновой территории Кольского полуострова и в пределах буферной зоны комбината “Североникель” (г. Мончегорск). Выявлены особенности в накоплении биомассы различных компонентов напочвенного покрова (лишайников, мхов, кустарничков), а также запасов растительного опада и лесной подстилки в зависимости от положения в тессере, под которой понимается совокупность приствольного, подкронового и межкронового пространств деревьев Pinus sylvestris L., различающихся локальными экологическими условиями. Показано, что существенное внутриэкосистемное варьирование этих параметров наблюдается как в фоновых условиях, так и в условиях аэротехногенного загрязнения. Вследствие адсорбции древесным пологом полиметаллической пыли, выпадающей из загрязненной атмосферы, в приствольных и подкроновых зонах деревьев Pinus sylvestris регистрируется повышенный уровень загрязнения почвы тяжелыми металлами, что отражается в реакции напочвенного покрова. При индексе техногенной нагрузки <10 отн. ед. не нарушаются закономерности накопления органического вещества компонентами почвенно-растительного покрова в среднем в фитоценозе и в зависимости от положения в тессере. При увеличении индекса техногенной нагрузки в среднем до 15 отн. ед. изменяется видовой состав и структура мохово-лишайникового яруса, сокращаются запасы его биомассы, надземной биомассы кустарничков и общего запаса биомассы напочвенного покрова, а также уменьшается толщина лесной подстилки.
Сообщества северотаежных лесов характеризуются ярко выраженной неоднородностью ценотической среды и мозаичностью напочвенного покрова, которая преимущественно обусловлена влиянием древесного яруса. Деревья – эдификаторы лесного фитоценоза вызывают пространственную неоднородность и временнýю изменчивость освещенности, микроклимата, водного, температурного и питательного режимов почвы. Микромозаика растительности в лесных экосистемах определяет пространственную гетерогенность лесных почв, что наиболее четко отражается на составе их органогенных горизонтов, формирующихся из растительного опада. Изучение микромозаики напочвенного покрова и верхних горизонтов почв в пределах лесных биогеоценозов преимущественно проводилось в северотаежных еловых лесах или в темнохвойных лесах средней тайги, где внутриценотическая мозаичность выражена наиболее контрастно [1–8]. В подзоне елово-широколиственных лесов выявлены четкие закономерности изменения в видовом составе живого напочвенного покрова ельников в зависимости от положения в тессере, под которой понимается совокупность приствольного, подкронового и межкронового пространства, различающихся локальными экологическими условиями. От приствольных пространств к межкроновым в составе напочвенного покрова увеличивается разнообразие экологических свит, общее число видов, долевое участие неморальных, светолюбивых и требовательных к общему содержанию элементов питания видов. Толщина и запас лесных подстилок уменьшаются от приствольных пространств к окнам. Положение в тессере определяет фракционный состав подстилок, а также соотношение величин запасов горизонтов внутри подстилки [9]. Значительно меньше работ посвящено изучению пространственной гетерогенности напочвенного покрова и верхних горизонтов почв в сосновых биогеоценозах, что, по-видимому, связано с тем, что фитоценозы сосновых лесов отличаются более равномерной освещенностью [7, 10–13]. Анализ базы данных по биомассе напочвенного покрова бореальных и гемибореальных лесов России показал, что по биомассе видов сосудистых растений в наибольшей степени различаются между собой пространственные микрогруппировки, выделяемые в лесном напочвенном покрове по доминирующей эколого-ценотической группе [14]. Авторами сделан вывод о перспективности использования пространственных микрогруппировок для моделирования динамики напочвенного покрова в круговоротных моделях лесных экосистем.
Аэротехногенное загрязнение окружающей среды оказывает существенное воздействие на структуру и продуктивность фитоценозов, а также вносит дополнительный вклад в формирование пространственной неоднородности накопления и разложения органического вещества, способствует изменению фракционного состава, запасов и скорости разложения растительного опада и приводит к трансформации органических веществ лесных подстилок [15–18].
Для исследования элементов внутриценотической неоднородности растительности и почвенного покрова используют разные термины, в наших исследованиях мы придерживаемся определения “микроместообитания” [13]. В соответствии с устоявшейся терминологией [16, 18] мы называем леса на территории буферной зоны дефолиирующими.
Цель данного исследования – оценка биомассы компонентов напочвенного покрова (лишайники, мхи, кустарнички), а также запасов растительного опада и лесной подстилки в разных микроместообитаниях фоновых и дефолиирующих (буферная зона медно-никелевого комбината) средневозрастных северотаежных сосновых лесов.
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Исследования проводили в средневозрастных сосновых лесах, расположенных в фоновом районе Кольского полуострова в среднем течении р. Ливы, и на территории буферной зоны комбината “Североникель” (Мурманская обл.). Давность последнего пожара в исследованных сообществах составляет в среднем 90 лет. Эдификатором древесного яруса является Pinus sylvestris L., в составе древостоев присутствует Betula pubescens Ehrh. В травяно-кустарничковом ярусе фоновых сосновых лесов доминирующими видами являются кустарнички Vaccinium vitis-idaea L., V. myrtillus L., Empetrum hermaphroditum Hagerup, в мохово-лишайниковом – зеленые мхи Pleurozium schreberi (Brid.) Mitt., Dicranum sp. и лишайники Cladonia rangiferina (L.) Weber ex F.H. Wigg., Cl. stellaris (Opiz.) Pouzar & Vězda, Cl. arbuscula (Wallr.) Flot. На территории буферной зоны в травяно-кустарничковом ярусе доминируют те же виды кустарничков, а в мохово-лишайниковом ярусе преобладают лишайники р. Cladonia с шиловидными и сцифовидными подециями. Согласно современной классификации почв России [19], исследуемые почвы относятся к Al-Fe-гумусовым подзолам или к Albic Rustic Podzols, согласно классификации WRB [20].
Комбинат “Североникель” вступил в строй в 1938 г. и до 1968 г. использовал руду местного происхождения, а затем перешел на руду Норильского месторождения, что привело к резкому увеличению объемов атмосферных выбросов, особенно диоксида серы. Согласно опубликованным данным, в период 1981–1990 гг. ежегодный объем атмосферных выбросов SO2 ОАО “Североникель” превышал в среднем 220 тыс. т, твердых веществ – 16 тыс. т, затем происходило постепенное снижение объемов выбросов, и к концу XX в., согласно официальным данным (http://www/kolagmk.ru), они составляли соответственно 45.8 и 6.0 тыс. т в год. В настоящее время ежегодный объем выбросов SO2 и твердых веществ на комбинате “Североникель” сократился соответственно в 8 и 5 раз по сравнению с их максимальными величинами.
Исследование проводили на постоянных пробных площадях (ППП) размером 20 × 20 м, заложенных в лишайниково-зеленомошных сосновых лесах в фоновом районе (ППП 1) на расстоянии 70 км от комбината “Североникель” и в буферной зоне – ППП 2 и ППП 3, удаленных от комбината на расстояние 35 км в юго-западном и 27 км в северо-восточном направлении соответственно. Средний возраст деревьев Pinus sylvestris составляет 70–80 лет, средняя высота – 8–11 м, средний диаметр ствола на высоте 1.3 м – 7.7–12 см. Общее проективное покрытие травяно-кустарничкового яруса составляет в среднем фоновом районе 22%, в буферной зоне 17–23%; мохово-лишайникового яруса – соответственно 75 и 47–65%, при этом на ППП 3 в лишайниковом покрове присутствуют довольно обильно кустистые лишайники р. Cladonia.
На каждой ППП были заложены трансекты, на которых с интервалом 1 м размещали учетные площадки размером 10 × 10 см. Площадки располагались в следующих типах микроместообитаний: А – в приствольной зоне в 20 см от стволов деревьев сосны с 4-х сторон света; B – в подкроновых пространствах деревьев; C – на межкроновых участках в “окнах” полога древостоя. Всего заложено 170 учетных площадок.
В пределах каждой учетной площадки был срезан напочвенный покров на уровне лесной подстилки и разобран на следующие компоненты: живые части кустарничков, лишайников, мхов; была измерена толщина лесной подстилки, отобран растительный опад и органогенный горизонт (О), из которого были извлечены все подземные части растений. Образцы растительного материала и лесной подстилки высушены до воздушно-сухого состояния и взвешены. Запас надземной биомассы включает надземные части кустарничков, мхов и лишайников, а в общий запас живого напочвенного покрова включена биомасса подземных частей растений – подземные побеги и корни кустарничков, корни травянистых растений. Запас биомассы разных компонентов напочвенного покрова и запасы растительного опада и лесной подстилки рассчитаны в г/м2.
Для оценки уровня загрязнения лесной подстилки тяжелыми металлами рассчитывали индекс техногенной нагрузки, который представляет собой превышение суммарного содержания кислоторастворимых форм преобладающих металлов (Ni, Cu и Co) в подстилке над их фоновым содержанием [15].
Проверка выборок исследуемых параметров на соответствие закону нормального распределения показала, что распределения большинства параметров значимо отличаются от нормального распределения. В связи с этим при оценке значимости различий использовали непараметрические критерии Краскела–Уоллиса (H) и Манна–Уитни (z), при уровне значимости р < 0.05 различия считали достоверными. На рисунках и в таблицах представлены средние значения и их стандартная ошибка.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
В фоновом сосновом лесу на ППП 1 среднее значение суммарной концентрации кислоторастворимых форм Ni, Cu и Со в органогенном горизонте Al–Fe-гумусовых подзолов составляет 20.0 ± ± 0.5 мг/кг, при этом не выявлено достоверных различий в их содержании в разных типах микроместообитаний, поэтому при расчете индекса техногенной нагрузки суммарное содержание этих металлов было принято за 1.
Запас надземной биомассы лишайников, мхов и кустарничков в фоновом сосняке лишайниково-зеленомошном (ППП 1) в разных типах микроместообитаний достоверно не различается (табл. 1), что свидетельствует о достаточно равномерном распределении запасов биомассы компонентов напочвенного покрова по площади фитоценоза. Однако использование объединенной выборки данных по приствольным и подкроновым микроместообитаниям показало, что запас надземной и подземной биомассы кустарничков на этих участках достоверно больше по сравнению с межкроновыми микроместообитаниями (z = 2.39–2.44, p = 0.045–0.047), а для запаса биомассы мхов регистрируется противоположная закономерность (z = 2.56, p = 0.043). Запас надземной биомассы травяно-кустарничкового яруса фактически совпадает с таковым кустарничков, т.к. травянистые растения практически отсутствуют в исследуемом типе северотаежных сосновых лесов.
Таблица 1.
Запас (г/м2) биомассы компонентов напочвенного покрова в разных микроместообитаниях исследуемых сосновых сообществ Table 1. Stock (g/m2) of groundcover biomass components in different microhabitats of pine communities
| Компонент Component |
Микроместообитания Microhabitats |
Критерий
Краскела–Уоллиса (H) Kruskal–Wallis test (H) |
Уровень значимости (p) Significance level (p) |
||
|---|---|---|---|---|---|
| приствольные at tree base |
подкроновые under crowns |
межкроновые on canopy gaps |
|||
| ППП 1 (PSP 1) | |||||
| Лишайники Lichens |
110 ± 31 | 101 ± 34 | 130 ± 39 | 0.04 | 0.98 |
| Мхи Mosses |
67 ± 35 | 34 ± 6 | 172 ± 65 | 1.87 | 0.39 |
| Кустарнички: Dwarf shrubs |
|||||
| надземные части above-ground parts |
106 ± 25 | 91 ± 18 | 68 ± 10 | 2.31 | 0.32 |
| подземные части underground parts |
289 ± 32 | 362 ± 40 | 213 ± 25 | 8.45 | 0.015 |
| ППП 2 (PSP 2) | |||||
| Лишайники Lichens |
155 ± 43 | 139 ± 36 | 175 ± 35 | 2.77 | 0.25 |
| Мхи Mosses |
– | – | – | – | – |
| Кустарнички: Dwarf shrubs |
|||||
| надземные части above-ground parts |
35 ± 11 | 21 ± 5 | 32 ± 17 | 0.31 | 0.85 |
| подземные части underground parts |
136 ± 30 | 161 ± 50 | 266 ± 118 | 0.26 | 0.88 |
| ППП 3 (PSP 3) | |||||
| Лишайники Lichens |
196 ± 38 | 173 ± 29 | 210 ± 31 | 0.94 | 0.62 |
| Мхи Mosses |
42 ± 13 | 12 ± 7 | 45 ± 15 | 7.16 | 0.028 |
| Кустарнички: Dwarf shrubs |
|||||
| надземные части above-ground parts |
116 ± 19 | 111 ± 17 | 56 ± 14 | 8.54 | 0.014 |
| подземные части underground parts |
414 ± 34 | 436 ± 36 | 323 ± 30 | 5.08 | 0.079 |
Примечание. * – Здесь и в табл. 2: жирным шрифтом выделены величины уровня значимости, указывающие на достоверные различия параметра в разных микроместообитаниях. Note. * – Here and in tab. 2 in bold, are the values of significance level indicating significant differences of the parameter in different microhabitats.
Запас биомассы мохово-лишайникового яруса в зависимости от положения в тессере составляет соответственно 177, 135, 302 г/м2 (табл. 1), но вследствие высокой степени варьирования этого показателя достоверные различия в разных типах микроместообитаний отсутствуют. Запас надземной биомассы напочвенного покрова и общая величина его биомассы с учетом подземных частей растений в рассматриваемом сообществе достоверно не различаются в разных частях тессеры и составляют в среднем 290 и 580 г/м2 (рис. 1, 2).
Рис. 1.
Запас надземной биомассы напочвенного покрова в приствольных (А), подкроновых (B) и межкроновых (C) микроместообитаниях исследованных сосновых сообществ. По вертикали: запас надземной биомассы, г/м2; по горизонтали: номера пробных площадей. Fig. 1. Stock of the above-ground biomass in different microhabitats of the studied Scots pine communities: at tree base (А), under crowns (B), on canopy gaps (C) X-axis: total stock of the above-ground biomass, g/m2; y-axis: sample plots.
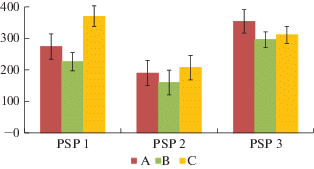
Рис. 2.
Общий запас биомассы живого напочвенного покрова (с учетом подземных частей) в различных микроместообитаниях исследованных сосновых сообществ. Обозначения типов микроместообитаний как на рис. 1. По вертикали: общий запас биомассы живого напочвенного покрова, г/м2; по горизонтали: номера пробных площадей. Fig. 2. Total stock of the live ground cover biomass (including underground parts) in different microhabitats of the studied Scots pine communities. Designation of microhabitat types as in Fig. 1. X-axis: total stock of the live groundcover biomass, g/m2; y-axis: sample plots.

Высокая степень варьирования запаса надземной биомассы напочвенного покрова в пределах тессер в разных типах еловых лесов выявлена в работах В.М. Телесниной с соавторами [21] и О.В. Семенюк с соавторами [9]. Авторы установили, что внутрибиогеоценозное варьирование надземной биомассы напочвенного покрова чрезвычайно высоко (2.5–1800 г/м2) и превышает межбиогеоценозное варьирование, при этом фактором, определяющим увеличение биомассы напочвенного покрова от приствольных к межкроновым пространствам, является освещенность. В редкостойных северотаежных лесах фактор освещенности не играет столь существенной роли, как в еловых лесах южной тайги, однако как в еловых, так и в сосновых фоновых лесах Кольского полуострова влияние деревьев на формирование биомассы напочвенного покрова остается по-прежнему значительным. В.В. Никонов и др [11] указывают, что в сосняках кустарничково-лишайниковых наименьший запас надземной биомассы напочвенного покрова регистрируется в приствольных зонах (758 г/м2), а максимальный – в подкроновых пространствах (1414–1482 г/м2) деревьев сосны, при этом, как видно из приводимой авторами таблицы, запас биомассы в межкроновых (730 г/м2) и приствольных зонах достоверно не различается. Как показали наши предшествующие исследования [13], максимальные величины запаса надземной биомассы напочвенного покрова отмечаются в наиболее сухом лишайниковом сосновом редколесье (430 г/м2), а минимальные – в более влажном сосняке зеленомошном (125 г/м2), при этом наиболее контрастно запасы биомассы нижних ярусов распределены по площади фитоценоза в сосновом редколесье, а наиболее выровнены – в сосняке зеленомошном. Рассматриваемые нами сообщества сосновых лесов находятся лишь на промежуточной стадии постпирогенного восстановления (давность последнего пожара составляет 90 лет), когда общее проективное покрытие и высота травяно-кустарничкового и мохово-лишайникового ярусов еще не достигли стабилизации, регистрируемой при давности пожара свыше 100 лет [22].
В рассматриваемом фоновом сосновом сообществе (ППП 1) масса растительного опада в среднем составляет 1280 г/м2, причем в приствольной зоне и в подкроновом пространстве деревьев P. sylvestris его запас достоверно выше, чем на межкроновых участках (табл. 2), что вполне логично, т.к. именно на приствольные и подкроновые участки попадает основная масса древесного опада (хвоя, кора, сухие ветки, шишки). Согласно данным Н.А. Артемкиной с соавторами [6], в ельниках кустарничково-зеленомошных вклад хвои Picea abies × obovata в общий запас опада еловых элементарных биогеоареалов (ЭБГА) варьирует в пределах 56–93%, в то время как в кустарничково-зеленомошном ЭБГА, формирующемся в межкроновых пространствах, опад на 100% представлен листьями кустарничков, травами и мхами. Сходный характер изменения запаса древесного опада в тессере отмечается и в фоновых сосновых лесах [16].
Таблица 2.
Запас (г/м2) массы опада и подстилки в разных микроместообитаниях исследуемых сосновых лесов Table 2. Stock (g/m2) of plant waste and forest litter in different microhabitats of pine forests
| Компонент Component |
Микроместообитания Microhabitats |
Критерий Краскела–Уоллиса (H) Kruskal–Wallis test | Уровень значимости (p) Significance level (p) |
||
|---|---|---|---|---|---|
| приствольные at tree base |
подкроновые under crowns |
межкроновые on canopy gaps |
|||
| ППП 1, PSP 1 | |||||
| Масса опада Weight of plant waste |
1653 ± 176 | 1446 ± 168 | 734 ± 65 | 20.6 | 0.000 |
| Масса подстилки Weight of forest litter |
3827 ± 640 | 4814 ± 472 | 3890 ± 316 | 3.90 | 0.14 |
| Толщина подстилки, см Thickness of forest litter, cm |
5.0 ± 0.6 | 4.4 ± 0.3 | 3.9 ± 0.5 | 3.78 | 0.15 |
| ППП 2, PSP 2 | |||||
| Масса опада Weight of plant waste |
2660 ± 166 | 1915 ± 193 | 1040 ± 107 | 23.4 | 0.000 |
| Масса подстилки Weight of forest litter |
4700 ± 437 | 3750 ± 290 | 5190 ± 993 | 1.5 | 0.48 |
| Толщина подстилки, см Thickness of forest litter, cm |
3.8 ± 0.5 | 2.7 ± 0.5 | 2.3 ± 0.4 | 6.9 | 0.031 |
| Индекс техногенной нагрузки, отн. ед. Index of man-made load, arb. unit |
17.0 ± 1.3 | 16.0 ± 1.3 | 13.2 ± 1.6 | 3.4 | 0.19 |
| ППП 3, PSP 3 | |||||
| Масса опада Weight of plant waste |
1655 ± 164 | 1480 ± 76 | 1465 ± 159 | 1.89 | 0.39 |
| Масса подстилки Weight of forest litter |
5050 ± 654 | 4650 ± 413 | 4230 ± 441 | 0.69 | 0.71 |
| Толщина подстилки, см Thickness of forest litter, cm |
5.4 ± 0.4 | 3.9 ± 0.3 | 3.1 ± 0.3 | 17.7 | 0.000 |
| Индекс техногенной нагрузки, отн. ед. Index of man-made load, arb. unit |
9.6 ± 0.3 | 9.2 ± 0.4 | 7.2 ± 0.3 | 26.9 | 0.000 |
Толщина лесной подстилки на ППП 1 по типам микроместообитаний достоверно не различается и в среднем составляет 4.4 см, что близко к средней величине, наблюдаемой при полном восстановлении толщины подстилки в сосняках лишайниково-зеленомошных [22]. Диапазон варьирования толщины подстилки очень широк: от 1.5 до 9.0 см. Величина запаса лесной подстилки в среднем составляет 4180 г/м2, интервал варьирования этого показателя составляет от 2020 до 10 200 г/м2, т.е. различия достигают 5 крат. При столь значительном варьировании различия этого показателя по типам микроместообитаний, также как толщины подстилки, недостоверны, т.е. эти параметры не связаны с положением в тессере (табл. 2). Высокая степень пространственной изменчивости толщины и запаса подстилки внутри фитоценоза выявлена в работах В.М. Телесниной с соавторами [21] и О.В. Семенюк с соавторами [9]: величина запасов подстилки в ельниках изменяется в 4 раза (от 1500 до 6000 г/м2), при этом толщина подстилки существенно уменьшается в ряду ствол–крона–окно. Значительные запасы и доля мелких фракций в пределах профиля в гор. F и H, а также в верхних деструктивных горизонтах свидетельствует о низкой скорости биологического круговорота и депонировании органического вещества [21]. На различия в скорости разложения растительных остатков в фоновых хвойных лесах Кольского полуострова указывают данные Е.А. Ивановой с соавторами [18]. Авторами показано, что в течение двух лет опад вечнозеленых растений в еловых лесах разлагался значительно быстрее, чем в сосновых.
Таким образом, следует подчеркнуть высокую внутриценотическую вариабельность всех исследуемых показателей в фоновом сосняке лишайниково-зеленомошном, что в большинстве случаев не позволяет выявить связь запасов компонентов лесных экосистем с положением в тессере.
Как уже отмечалось выше, на территории буферной зоны обе исследованные пробные площади (ППП 2 и 3) находятся в противоположных направлениях от комбината “Североникель”. Согласно многолетним данным, в районе исследования преобладают ветры, дующие в юго-западном направлении от г. Мончегорска, что является причиной более сильной степени загрязнения почв тяжелыми металлами на ППП 2 по сравнению с ППП 3 и нашло отражение в величине индекса техногенной нагрузки, который в среднем составляет соответственно 15.4 и 8.7 отн. ед., то есть во втором случае его среднее значение почти в 2 раза меньше. Следует подчеркнуть, что принадлежность обеих исследуемых площадей к буферной зоне не вызывает сомнений. Мониторинг индекса техногенной нагрузки на территории буферной зоны за период 2005–2019 гг. показал, что на ППП 2 и 3 интервалы его варьирования по годам в значительной степени перекрываются, составляя соответственно 9.4–28.5 и 4.6–26.1 отн. ед. Ранее мы также отмечали высокую степень погодичного варьирования содержания кислоторастворимых форм тяжелых металлов в органогенном горизонте подзолов, как на территории буферной, так и импактной зоны [23]. При анализе многолетней динамики уровня загрязнения почв в окрестностях комбината “Североникель” выявлено 1.5–7-кратное варьирование содержания тяжелых металлов в органогенном горизонте почв [24, 25]. По мнению автора, наиболее важными факторами варьирования концентрации Ni, Cu и Co являются содержание органического вещества в отобранном образце, а также количество атмосферных осадков, выпавших за предшествующий отбору образца год. При этом отмечается, что состояние экосистем в локальной зоне варьирует в широком диапазоне и не всегда согласуется с концентрациями основных металлов-загрязнителей в почве [24].
На ППП 3 величина индекса техногенной нагрузки достоверно меньше в межкроновых микроместообитаниях по отношению к приствольным и подкроновым (табл. 2). На ППП 2 различия в величине индекса техногенной нагрузки в приствольных и подкроновых зонах по сравнению с межкроновыми также достоверны (z = 2.69, p = 0.043). Логично предположить, что кроны деревьев адсорбируют частицы полиметаллической пыли из загрязненного воздуха, а при опадании хвои, коры и сухих ветвей дополнительная часть тяжелых металлов переходит в подстилку из разлагающегося опада. В дефолиирующих лесах коэффициент обогащения Ni и Cu растительных остатков вечнозеленых растений составлял в еловых лесах 2.85 и 7.27, в сосновых – 1.72 и 2.43 соответственно [18]. Ранее было показано, что в пределах импактной зоны относительное содержание тяжелых металлов в разных фракциях фитомассы деревьев сосны находится в убывающем ряду: сухие ветви > неохвоенные живые ветви > > хвоя > корни > древесина [26].
Различия в уровне загрязнения лесной подстилки тяжелыми металлами обусловили различия в накоплении биомассы отдельными компонентами напочвенного покрова. При существенно большем среднем индексе техногенной нагрузки на ППП 2 зеленые мхи практически полностью выпали из состава мохово-лишайникового яруса, как наиболее чувствительные виды к аэротехногенному загрязнению (табл. 1). На ППП 3 величина запаса биомассы мхов в разных типах микроместообитаний в 1.5–3.8 раза меньше соответствующих значений на ППП 1, при этом в подкроновых зонах она достоверно меньше по отношению к приствольным и межкроновым зонам.
В буферной зоне на обеих исследуемых пробных площадях в разных типах микроместообитаний величины запаса биомассы лишайников достоверно не различаются, при этом они в 1.3–1.8 раз больше по отношению к этой величине в фоновом районе. Следует отметить, что, как в фоновом сосняке, так и в буферной зоне, запас биомассы мохово-лишайникового яруса достаточно равномерно распределен по площади фитоценоза и не связан с положением в тессере. Полученные результаты вполне согласуются с более ранними нашими данными [27], где на ППП 2 запас биомассы лишайников, равно как и запас биомассы мохово-лишайникового яруса, составлял в среднем 179 г/м2. При этом мы констатировали процесс восстановления лишайникового покрова, поскольку запас биомассы лишайников вырос в 7.5 раз по сравнению с его запасом в период высокой аэротехногенной нагрузки, когда он составлял лишь 24 г/м2 [28]. Кроме того, в условиях полевого эксперимента по искусственному загрязнению полиметаллической пылью было установлено, что в сосняках лишайниковых и лишайниково-зеленомошных при индексе техногенной нагрузки ≤10 отн. ед. состояние мохово-лишайникового яруса оценивается как ненарушенное, и это значение индекса техногенной нагрузки является пороговым для снижения запаса биомассы мохово-лишайникового яруса [29].
На сравниваемых пробных площадях в буферной зоне запас надземной биомассы кустарничков различается более чем в 3 раза. На ППП 2 его средняя величина составляет лишь 29 г/м2, что очень близко к зарегистрированному значению на этой пробной площади в 2014 г., когда оно было равно 35 г/м2 [27]. На ППП 3 запас надземной биомассы кустарничков составляет 94 г/м2, что практически равно запасу надземной биомассы кустарничков в фоновом сосняке (88 г/м2). На ППП 2 запас как надземной, так и подземной биомассы кустарничков распределен достаточно равномерно по площади фитоценоза, достоверные различия в этих показателях в разных типах микроместообитаний отсутствуют, при этом регистрируются наименьшие значения обоих параметров среди исследуемых сосновых лесов (табл. 1). На ППП 3 запас надземной биомассы кустарничков, равно как и травяно-кустарничкового яруса в целом, достоверно меньше в межкроновых микроместообитаниях по отношению к приствольным и подкроновым участкам. Значения запаса надземной биомассы и характер его изменения в зависимости от положения в тессере сходны с наблюдаемыми в фоновом фитоценозе. Запас подземной биомассы кустарничков на ППП 3 2-кратно превышает его величину на ППП 2, средние значения составляют соответственно 390 и 188 г/м2, в то время как в фоновом сосняке эта величина равна 288 г/м2.
Анализ соотношения запасов биомассы надземных и подземных частей кустарничков в межкроновых пространствах, под кронами и в непосредственной близости от стволов Pinus sylvestris выявил сходный характер его изменения, как в фоновом сосняке, так и в буферной зоне. Во всех случаях запас надземной биомассы кустарничков существенно меньше, чем запас биомассы подземных частей (рис. 3). В фоновом сосняке (ППП 1) запас надземной биомассы в разных микроместообитаниях составляет 20–27% от их общего запаса, на территории буферной зоны – 11.5–22%. На всех пробных площадях более высокая доля надземных частей в общем запасе биомассы кустарничков регистрируется в приствольной зоне, а более низкая – в межкроновых пространствах. В ходе анализа базы данных по надземной и подземной биомассе напочвенного покрова бореальных и гемибореальных лесов России установлено, что подземная биомасса видов разных эколого-ценотических групп различается между собой в большей степени, чем надземная, причем для бореальной группы кустарничков характерны более высокие значения подземной биомассы по сравнению с надземной [14].
Рис. 3.
Соотношение запасов биомассы подземных (1) и надземных (2) частей кустарничков в приствольном (А) и подкроновом (B) пространстве деревьев Pinus sylvestris и в межкроновых микроместообитаниях (C) исследованных сосновых сообществ. По вертикали: относительный запас биомассы, %; по горизонтали: тип микроместообитания. Fig. 3. Proportion of the stock of the above- (1) and underground (2) shrub biomass in microhabitats at tree base (A) and under crown (B) of Pinus sylvestris and on canopy gaps (C) of the studied Scots pine communities. X-axis: biomass relative stock, %; y-axis: microhabitat type.

Запас надземной биомассы и общий запас живого напочвенного покрова с учетом подземных частей в буферной зоне достоверно различается на ППП 2 и ППП 3 (z = 2.31–5.90, p = 0.000–0.021): на ППП 3 эти величины (соответственно 320 и 710 г/м2) почти в 2 раза больше, чем на ППП 2 (185 и 375 г/м2) и сопоставимы со значениями, регистрируемыми в фоновом сосняке (290 и 580 г/м2). В условиях экспериментального загрязнения почвы полиметаллической пылью при среднем значении индекса техногенной нагрузки 15 отн. ед. (таком же, как на ППП 2) общий запас надземной биомассы напочвенного покрова варьировал от 250 (в лишайниково-зеленомошном сосняке) до 350 г/м2 (в старовозрастном лишайниковом сосняке) и достоверно не различался в разных сообществах [29].
Запас надземной биомассы напочвенного покрова достоверно не различается в разных типах микроместообитаний как на ППП 2, так и на ППП 3. В фоновом сосняке в межкроновых зонах величина этого показателя примерно в 1.5 раза выше его среднего значения на приствольных и подкроновых участках (рис. 1). Характер изменения запаса живого напочвенного покрова с учетом подземных частей от приствольных к межкроновым участкам несколько различается в исследуемых сообществах (рис. 2). В фоновом сосняке за счет увеличения массы подземных частей кустарничков пространственные различия в общем запасе нивелируются. На территории буферной зоны прослеживаются противоположные тенденции изменения этого показателя в зависимости от положения в тессере. На ППП 2 запас биомассы с учетом подземных частей на приствольных и подкроновых участках достоверно меньше (z = 2.35, p = 0.023), а на ППП 3, напротив, достоверно больше (z = 3.15, p = 0.011) по отношению к его величине в межкроновых микроместообитаниях, что связано с разнонаправленными тенденциями изменения запасов подземных частей растений в тессере на сравниваемых пробных площадях.
Соотношение запасов надземной биомассы травяно-кустарничкового и мохово-лишайникового ярусов в межкроновых пространствах, под кронами и в непосредственной близости от стволов Pinus sylvestris оказалось различным в фоновом сосняке и на наиболее загрязненной ППП 2 в буферной зоне (рис. 4). Основной вклад в общий запас надземной биомассы напочвенного покрова во всех исследуемых сосновых лесах вносит мохово-лишайниковый ярус, и аэротехногенное загрязнение не оказывает влияния на это соотношение, что согласуется с данными других исследователей [11]. Сходную закономерность преобладания вклада мохово-лишайникового яруса в общий запас надземной биомассы отмечали и другие исследователи при изучении микромозаики напочвенного покрова в сосновых лесах [10, 11, 27, 28]. Напротив, в темнохвойных лесах северной и средней тайги вклад сосудистых растений в общий запас надземной биомассы может достигать 57–90%, что значительно больше, чем вклад мхов [3, 4, 6, 10, 11].
Рис. 4.
Соотношение запасов биомассы мохово-лишайникового (1) и травяно-кустарничкового (2) ярусов в приствольном (А) и подкроновом (B) пространстве деревьев Pinus sylvestris и межкроновых микроместообитаниях (C) исследованных сосновых сообществ. По вертикали: относительный запас биомассы, %; по горизонтали: тип микроместообитания. Fig. 4. Proportion of the stock of lichen-moss (1) and grass-dwarf shrub (2) layers biomass in microhabitats at tree base (A) and under crown (B) of Pinus sylvestris and on canopy gaps (C) of the studied Scots pine communities. X-axis: biomass relative stock, %; y-axis: microhabitat type.
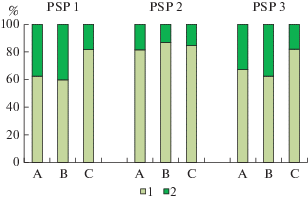
Доля травяно-кустарничкового яруса на ППП 1 и на ППП 3 является более высокой (33–40%) в приствольных и подкроновых микроместообитаниях, а на межкроновых участках его вклад одинаков на обеих ППП и не превышает 18.5%. Наиболее низкая доля биомассы растений травяно-кустарничкового яруса в общем запасе надземной биомассы (13–18%), которая остается постоянной в разных микроместообитаниях, наблюдается на ППП 2, что, возможно, обусловлено большей долей участия шиловидных и бокальчатых видов лишайников р. Cladonia в проективном покрытии мохово-лишайникового яруса. Аналогичная закономерность снижения доли участия травяно-кустарничкового яруса в общем запасе надземной биомассы от приствольных к межкроновым зонам выявлена в фоновых сосновых (от 38 до 20%) и еловых лесах (от 90 до 57%) [11].
Несмотря на почти 2-кратные различия в уровне загрязнения органогенного горизонта почв тяжелыми металлами в буферной зоне, масса опада достоверно не различается на ППП 2 и ППП 3, средние величины составляют соответственно 1870 и 1535 г/м2, что почти в 1.5 раза больше, чем в фоновом сосняке. Характерно, что в исследуемых сообществах в приствольных микроместообитаниях накапливается больше опада по сравнению с межкроновыми, за исключением ППП 3, где масса опада на связана с положением в тессере (табл. 2). Полученные результаты хорошо согласуются с данными других авторов, согласно которым в дефолиирующих сосновых лесах суммарная масса опада в среднем в 1.2 раза превышает ее значение в фоновых условиях, при этом в подкроновых пространствах по сравнению с межкроновыми регистрируется 3.5-кратное возрастание запаса опада [16]. Увеличение мортмассы опада в условиях аэротехногенного загрязнения обусловлено более медленным его разложением, при этом особенно ярко эта закономерность проявляется в техногенном редколесье [18]. Кроме того, аэротехногенное загрязнение тормозит процессы разложения крупных древесных остатков (КДО). Так, вблизи медеплавильного завода в елово-пихтовых лесах в 3–4 раза увеличивается доля фрагментов КДО на начальных этапах разложения, что свидетельствует о сильном торможении их деструкции [30].
Средние значения запасов подстилки (4180, 4545 и 4770 г/м2) на всех исследуемых пробных площадях достоверно не различаются, так же, как и в разных типах микроместообитаний. В то время толщина подстилки имеет достоверные различия в буферной зоне. Самая тонкая подстилка наблюдается на ППП 2 – ее толщина в среднем не превышает 3 см; на ППП 3 средняя толщина подстилки (4.1 см) достоверно не отличается от ее величины в фоновом сосняке (4.4 см), что может свидетельствовать о более благоприятных условиях увлажнения на ППП 3 по сравнению с ППП 2. Корреляционный анализ данных выявил значимые связи между толщиной и запасом лесной подстилки только для буферной зоны (r = 0.50–0.64, p < 0.05), в фоновом сосняке такая связь отсутствует. Кроме того, следует подчеркнуть, что на всех исследуемых пробных площадях отсутствуют закономерности в изменении запаса лесной подстилки в зависимости от положения в тессере (табл. 2). Возможно, достоверные различия в запасе лесной подстилки в разных типах микроместообитаний сложно выявить из-за высокой степени вариабельности данного показателя, что отмечается и другими исследователями, как в еловых, так и сосновых лесах [9, 21, 31].
Таким образом, аэротехногенное загрязнение оказывает как прямое негативное воздействие на компоненты напочвенного покрова и верхнего горизонта почв, так и опосредованное через изменение биогеоценотических условий. К прямому негативному воздействию можно отнести увеличение содержания тяжелых металлов в разных компонентах наземных экосистем, что отмечалось многими исследователями, при этом необходимо подчеркнуть, что уровень загрязнения почв непосредственно связан не только с расстоянием от источника атмосферных выбросов, но и с розой ветров. Преобладание в Мончегорском районе ветров, дующих в юго-западном направлении, привело к 2-кратному повышению индекса техногенной нагрузки на ППП 2 по отношению к его величине на ППП 3, расположенной в северо-восточном направлении от комбината “Североникель”, хотя обе пробные площади находятся на территории буферной зоны.
Все исследуемые растительные сообщества относятся группе типов лишайниково-зеленомошных сосновых лесов, для которых, начиная с 80–100 лет после пожара, характерно восстановление соотношения лишайников и мхов в напочвенном покрове [22]. При этом доля мхов в общем покрытии мохово-лишайникового яруса составляет в среднем от 40 до 60%. Однако воздействие аэротехногенного загрязнения диоксидом серы совместно с полиметаллической пылью привело к изменению видового состава и структуры яруса, вплоть до полного выпадения доминанта мохового покрова Pleurozium schreberi на ППП 2. В свою очередь, эти нарушения обусловили изменение гидротермического режима верхних горизонтов почв. Известно, что в сосняках лишайниково-зеленомошных плотный сомкнутый мохово-лишайниковый ярус и довольно значительный по толщине, обладающий высокой порозностью и влагоемкостью слой подстилки и опада, характеризуются более высокими водоудерживающими и теплоизолирующими свойствами по сравнению с лишайниковым покровом и более тонким слоем подстилки и опада в лишайниковых сообществах [31–33]. В результате нарушения сложения мохово-лишайникового яруса под воздействием аэротехногенного загрязнения на территории буферной зоны создаются более контрастные гидротермические условия. Сочетанное воздействие этих факторов привело к 1.5–2-кратным различиям в запасе биомассы этого яруса, а также общем запасе напочвенного покрова на ППП 2 и ППП 3 и способствовало замедлению скорости разложения растительных остатков и накоплению мортмассы опада, которая в 1.2–1.5 раза превышает соответствующую величину в фоновом сосняке.
Логично предположить, что распределение по площади фитоценоза толщины и запаса лесной подстилки должно быть связано с запасом растительного опада, однако, как оказалось, согласованное изменение перечисленных параметров регистрируется не всегда. Если толщина подстилки и масса опада в большинстве случаев закономерно снижаются от приствольных к межкроновым зонам, то масса подстилки никак не связана ни с положением в тессере, ни с уровнем загрязнения местообитания. Возможно, это обусловлено высокой степенью вариабельности параметров лесных подстилок, что отмечается в ряде работ [9, 21, 31].
Сравнительный анализ различий в накоплении органического вещества компонентами сообществ лишайниково-зеленомошных сосновых лесов в разных типах микроместообитаний позволяет сделать ряд заключений, касающихся изменения средообразующей роли древесного яруса в условиях аэротехногенного загрязнения. Если принять за 1 величины исследованных параметров на межкроновых участках, то оказывается, что запас биомассы лишайников достаточно равномерно распределен по площади фитоценоза как в фоновой, так и в буферной зоне (табл. 3). В то же время хорошо выраженная в фоновой зоне контрастность величин запаса биомассы мхов в подкроновом и межкроновом пространстве сглаживается в буферной зоне за счет исчезновения различий между приствольными и межкроновыми участками. Различия в запасе надземной биомассы кустарничков в разных частях тессеры более четко выражены в фоновом сосняке и на ППП 3, где уровень загрязнения почвы еще не превышает порогового значения, по сравнению с ППП 2. Соотношение биомассы подземных частей растений в разных частях тессеры свидетельствует, что в пределах буферной зоны на ППП 2 подземные органы кустарничков слабее развиты на наиболее загрязненных подкроновых участках по сравнению с межкроновыми, тогда как в фоновом сообществе наблюдается обратное соотношение. Величина общего запаса биомассы напочвенного покрова наиболее ясно отражает тенденцию сглаживания различий между разными типами микроместообитаний в буферной зоне по сравнению с фоновыми условиями (табл. 3), что свидетельствует о снижении средообразующей роди древесного яруса в условиях умеренного загрязнения.
Таблица 3.
Соотношение запасов биомассы разных компонентов, толщины подстилки и индекса техногенной нагрузки в разных типах микроместообитаний исследуемых сосновых лесов Table 3. Ratio of the biomass stock of different components, thickness of forest litter and index of man-made load in different microhabitats of pine forests
| Компонент Component |
ППП 1 PSP 1 |
ППП 2 PSP 2 |
ППП 3 PSP 3 |
|---|---|---|---|
| Лишайники Lichens |
0.8 : 0.8 : 1 | 0.9 : 0.8 : 1 | 0.9 : 0.8 : 1 |
| Мхи Mosses |
0.4 : 0.2 : 1 | – | 0.9 : 0.3 : 1 |
| Кустарнички: Dwarf-shrubs |
|||
| надземные части above-ground parts |
1.6 : 1.3 : 1 | 1.1 : 0.7 : 1 | 2.1 : 2 : 1 |
| подземные части underground parts |
1.4 : 1.7 : 1 | 0.5 : 0.6 : 1 | 1.3 : 1.3 : 1 |
| Напочвенный покров в целом Groundcover |
0.6 : 0.4 : 1 | 0.9 : 0.8 : 1 | 0.9 : 0.7 : 1 |
| Масса опада Weight of plant waste |
2.3 : 2 : 1 | 2.6 : 1.8 : 1 | 1.1 : 1 : 1 |
| Масса подстилки Weight of forest litter |
1 : 1.2 : 1 | 0.9 : 0.7 : 1 | 1.2 : 1.1 : 1 |
| Толщина подстилки, см Thickness of forest litter, cm |
1.3 : 1.1 : 1 | 1.7 : 1.2 : 1 | 1.7 : 1.3 : 1 |
| Индекс техногенной нагрузки, отн. ед. Index of man-made load,arb. unit |
1 : 1 : 1 | 1.3 : 1.2 : 1 | 1.3 : 1.3 : 1 |
Контрастность в распределении запаса растительного опада и подстилки по типам микроместообитаний в сообществах буферной зоны по сравнению в фоновым сообществом существенно не изменяется. В то же время различия по толщине подстилки между приствольными и межкроновыми участками заметно усиливаются в дефолиирующих лесах по сравнению с фоновым сообществом (табл. 3).
В буферной зоне древесный ярус способствует неравномерному пространственному распределению загрязнителей по территории фитоценоза: в приствольных и подкроновых зонах уровень загрязнения органогенного горизонта почвы достоверно больше по сравнению с межкроновым пространством. Определенные трудности в оценке влияния древесного яруса на формирование микромозаики напочвенного покрова в северотаежных сосновых лесах в значительной мере обусловлены особенностями структуры крон сосны обыкновенной, их более низкой плотностью, ажурностью по сравнению с кронами ели. Тем не менее, полученные результаты показывают, что в дефолиирующих сосновых лесах средообразующая роль древесного яруса, формирующая пространственную неоднородность нижних ярусов сообществ, снижается. Это особенно четко проявляется в изменении соотношения величин общей биомассы напочвенного покрова по типам микроместообитаний.
ЗАКЛЮЧЕНИЕ
Исследование, проведенное в средневозрастных сосновых лесах Кольского полуострова позволило оценить общие закономерности и отличительные черты внутриценотической неоднородности распределения запасов биомассы компонентов напочвенного покрова, мортмассы опада и лесной подстилки в фоновых условиях и при умеренном уровне аэротехногенного загрязнения в буферной зоне медно-никелевого комбината.
В фоновом сосняке лишайниково-зеленомошном выявлена высокая вариабельность запасов биомассы лишайников, мхов и кустарничков, растительного опада и лесной подстилки, что в определенной степени затрудняет оценку связи запасов компонентов лесных экосистем с положением в тессере.
Подтверждено предположение, что роза ветров является одним из факторов, определяющим уровень загрязнения органогенного горизонта Al-Fe-гумусовых подзолов, что проявилось в почти 2-кратном различии индекса техногенной нагрузки в сообществах сосновых лесов на территории буферной зоны комбината “Североникель”, расположенных в разных направлениях от источника выбросов. Указанные различия повлекли за собой неодинаковую реакцию напочвенного покрова на воздействие аэротехногенного загрязнения. При индексе техногенной нагрузки <10 отн. ед. не нарушаются закономерности накопления органического вещества компонентами почвенно-растительного покрова в среднем в фитоценозе и в разных частях тессеры. При увеличении индекса техногенной нагрузки в среднем до 15 отн. ед. изменяется по сравнению с фоновыми условиями видовой состав и структура мохово-лишайникового яруса, сокращаются запасы его биомассы, надземной биомассы кустарничков и общего запаса биомассы напочвенного покрова, а также уменьшается толщина лесной подстилки. Полог древостоя адсорбирует выпадающие из атмосферы частицы полиметаллической пыли, которые впоследствии накапливаются в растительном опаде, что приводит к повышенному уровню загрязнения органогенного горизонта подзолов тяжелыми металлами в приствольных и подкроновых зонах деревьев Pinus sylvestris по отношению к межкроновому пространству. В условиях аэротехногенного загрязнения, регистрируемого на территории буферной зоны, средообразующая роль древесного яруса заметно ослабляется. Запас биомассы мохово-лишайникового и травяно-кустарничкового ярусов более равномерно распределен по площади фитоценоза по сравнению с его распределением в фоновом сосняке. В то же время в буферной зоне толщина лесной подстилки более контрастно различается в приствольных к межкроновых микроместообитаниях, однако ее запас не связан с положением в тессере. Для более полного выявления влияния древесного яруса на микромозаику напочвенного покрова необходимы дальнейшие исследования.
Список литературы
Орлова М.А., Лукина Н.В., Камаев И.О., Смирнов В.Э., Кравченко Т.В. 2011. Мозаичность лесных биогеоценозов и продуктивность почв. – Лесоведение. 6: 39–48. https://elibrary.ru/item.asp?id=17097643&
Орлова М.А., Лукина Н.В., Смирнов В.Э., Артемкина Н.А. 2016. Влияние ели на кислотность и содержании элементов питания в почвах северотаежных ельников кустарничково-зеленомошных. – Почвоведение. 11: 1355–1367. https://doi.org/10.7868/S0032180X16110071
Смирнова О.В., Алейников А.А., Семиколенных А.А., Бовкунов А.Д., Запрудина М.В., Смирнов Н.С. 2011. Пространственная неоднородность почвенно-растительного покрова темнохвойных лесов в Печоро-Илычском заповеднике. – Лесоведение. 6:67–78. https://elibrary.ru/item.asp?id=17097645
Луговая Д.Л., Смирнова О.В., Запрудина М.В., Алейников А.А., Смирнов В.Э. 2013. Микромозаичная организация и фитомасса напочвенного покрова в основных типах темнохвойных лесов Печоро-Илычского заповедника. – Экология. 1: 3–10. https://elibrary.ru/item.asp?doi=10.7868/S0367059713010083
Лебедева В.Х., Ипатов В.С., Тиходеева М.Ю. 2015. Неоднородность пространственной структуры живого напочвенного покрова в лесных сообществах. – Вестник СПбГУ. Сер. 3. Биология. 2: 32–46. https://biocomm.spbu.ru/article/view/859/750
Артемкина Н.А., Орлова М.А., Лукина Н.В. 2018. Микромозаика растительности и вариабельность химического состава L-горизонтов подстилки северотаежных ельников кустарничково-зеленомошных. – Лесоведение. 2: 97–106. https://doi.org/10.7868/S002411481802002X
Лукина Н.В., Ершов В.В., Горбачева Т.В., Орлова М.А., Исаева Л.Г., Тебенькова Д.Н. 2018. Оценка состава почвенных вод северотаежных хвойных лесов фоновых территорий индустриально развитого региона. – Почвоведение. 3:284–296. https://doi.org/10.7868/S0032180X18030036
Данилова М.А., Лукина Н.В., Кузнецова А.И., Смирнов В.Э. 2019. Влияние древесных растений на плодородие почв таежных лесов. – В сб.: Экологические проблемы северных регионов и пути их решения: Тез. докл. VII Всеросс. научн. конф. с международ. участием. Апатиты. С. 18–19. http://inep.ksc.ru/documents/11_ecol_prob_19.pdf
Семенюк О.В., Телеснина В.М., Богатырев Л.Г., Бенедиктова А.И., Кузнецова Я.Д. 2020. Оценка внутрибиогеоценозной изменчивости лесных подстилок и травяно-кустарничковой растительности в еловых насаждениях. – Почвоведение. 1: 31–43. https://doi.org/10.31857/S0032180X2001013X
Никонов В.В., Лукина Н.В. 1994. Биогеохимические функции лесов на северном пределе распространения. Апатиты. 315 с.
Никонов В.В., Лукина Н.В., Смирнова Е.В., Исаева Л.Г. 2002. Влияние Picea obovata и Pinus sylvestris на первичную продуктивность нижних ярусов хвойных лесов Кольского полуострова. – Ботан. журн. 87(8):107–119.
Иванова Е.А., Артемкина Н.А., Лукина Н.В. 2019. Разложение опада в сосновых лесах на северном пределе распространения на Кольском полуострове. – В сб.: Экологические проблемы северных регионов и пути их решения: Тез. докл. VII Всеросс. научн. конф. с международ. участием. Апатиты. С. 30–32. http://inep.ksc.ru/documents/11_ecol_prob_19.pdf
Лянгузова И.В., Примак П.А. 2019. Пространственное распределение запасов напочвенного покрова и лесной подстилки в средневозрастных сосновых лесах Кольского полуострова. – Растительные ресурсы. 4: 473–489. https://doi.org/10.1134/S003399461904006X
Ханина Л.Г., Грозовская И.С., Смирнов В.Э., Романов М.С., Бобровский М.В. 2013. Анализ базы данных по биомассе лесного напочвенного покрова для моделирования его динамики в круговоротных моделях лесных экосистем. – Хвойные бореальной зоны. XXXI (1–2): 22–29. https://www.elibrary.ru/item.asp?id=20285336
Динамика лесных сообществ Северо-Запада России. 2009. СПб. 276 с.
Иванова Е.А., Лукина Н.В. 2017. Варьирование массы и фракционного состава древесного опада в сосняках кустарничково-лишайниковых при аэротехногенном загрязнении. – Лесоведение. 5: 47–58. http://lesovedenie.ru/index.php/forestry/article/view/1049
Одинцов П.Е., Караванова Е.И., Степанова А.А. 2018. Трансформация водорастворимых органических веществ подстилок подзолов фоновых и техногенных территорий Кольского полуострова. – Почвоведение. 8: 1022–1032. https://doi.org/10.1134/S0032180X18080099
Иванова Е.А., Лукина Н.В., Данилова М.А., Артемкина Н.А., Смирнов В.Э., Ершов В.В., Исаева Л.Г. 2019. Влияние аэротехногенного загрязнения на скорость разложения растительных остатков в сосновых лесах на северном пределе распространения. – Лесоведение. 6: 533–546. http://lesovedenie.ru/index.php/forestry/article/view/1214
Шишов Л.Л., Тонконогов В.Д., Лебедева И.И., Герасимова М.И. 2004. Классификация и диагностика почв России. Смоленск. 342 с. https://www.rfbr.ru/rffi/ru/books/o_28031#7
Мировая реферативная база почвенных ресурсов 2014, исправленная и дополненная версия 2015. Международная система почвенной классификации для диагностики почв и создания легенд почвенных карт. Доклады о мировых почвенных ресурсах №106. ФАО, Рим. 203 с. http://www.fao.org/3/i3794ru/I3794RU.pdf
Телеснина В.М., Семенюк О.В., Богатырев Л.Г. 2017. Свойства лесных подстилок во взаимосвязи с напочвенным покровом в лесных экосистемах Подмосковья (на примере УОПЭЦ “Чашниково”). – Вестн. МГУ. Сер. 17. Почвоведение. 4: 11–20. https://www.elibrary.ru/item.asp?id=30267046
Горшков В.В., Ставрова Н.И., Баккал И.Ю. 2009. Основные этапы восстановительной динамики северотаежных лесов. – В кн.: Динамика лесных сообществ Северо-Запада России. СПб. С. 228–236.
Лянгузова И.В. 2017. Динамические тренды содержания тяжелых металлов в растениях и почвах при разном режиме аэротехногенной нагрузки. – Экология. 4: 250–260. https://doi.org/10.7868/S0367059717040114
Кашулина Г.М. 2017. Экстремальное загрязнение почв выбросами медно-никелевого предприятия на Кольском полуострове. – Почвоведение. 7: 860–873. https://doi.org/10.7868/S0032180X17070036
Кашулина Г.М. 2018. Мониторинг загрязнения почв тяжелыми металлами в окрестностях медно-никелевого предприятия на Кольском полуострове. – Почвоведение. 4: 493–505. https://doi.org/10.7868/S0032180X1804010X
Лянгузова И.В. 2016. Тяжелые металлы в северотаежных экосистемах России. Saarbrucken (Германия). 260 с.
Лянгузова И.В., Ярмишко В.Т., Евдокимов А.С., Беляева А.И. 2018. Состояние сосновых лесов Кольского полуострова на фоне снижения объемов атмосферных выбросов предприятием цветной металлургии. – Растительные ресурсы. 54(4): 516–531. https://doi.org/10.1134/S0033994618040039
Ярмишко В.Т., Ярмишко М.А. 2002. Влияние пожаров и атмосферного загрязнения на структуру и продуктивность напочвенного покрова и травяно-кустарничкового яруса сосновых лесов на европейском севере России. – Растительные ресурсы. 38(2): 40–54. https://www.elibrary.ru/item.asp?id=29012198
Бондаренко М.С., Лянгузова И.В., Горшков В.В., Баккал И.Ю. 2018. Изменение фитомассы нижних ярусов северотаежных сосновых лесов при экспериментальном загрязнении почв тяжелыми металлами. – Растительные ресурсы. 54(1): 59–74. https://www.elibrary.ru/item.asp?id=32328491
Бергман И.Е., Воробейчик Е.Л. 2017. Влияние выбросов медеплавильного завода на формирование запаса и разложение крупных древесных остатков в елово-пихтовых лесах. – Лесоведение. 1: 24–38.
Демаков Ю.П., Исаев А.В., Шарафутдинов Р.Н. 2017. Роль лесной подстилки в борах Марийского Заволжья и вариабельность ее параметров. – В сб.: Научные Труды Государственного природного заповедника “Большая Кокшага”. Вып. 8. С. 15–43. https://www.elibrary.ru/item.asp?id=30729474
Семко А.П. 1982. Гидротермический режим почв лесной зоны Кольского полуострова. Апатиты. 142 с.
Ипатов B.C., Трофимец В.И. 1988. Влияние лишайниковых и зеленомошных ковров на водный режим верхнего корнеобитаемого слоя почвы в сухих сосняках – Экология. 1:19–23.
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы