

Растительные ресурсы, 2020, T. 56, № 4, стр. 314-325
Скорость роста и структура фитомассы Pinus sylvestris (Pinaceae) в средневозрастных сосняках Мурманской области
В. Т. Ярмишко 1, *, О. В. Игнатьева 2
1 Ботанический институт им В.Л. Комарова РАН
г. Санкт-Петербург, Россия
2 Санкт-Петербургский государственный лесотехнический университет им. С.М. Кирова
г. Санкт-Петербург, Россия
* E-mail: vasiliyarmishko@yandex.ru
Поступила в редакцию 19.05.2020
После доработки 24.07.2020
Принята к публикации 16.09.2020
Аннотация
Исследованы особенности динамики роста и формирования фитомассы сосны обыкновенной Pinus sylvestris L. в 60–80-летних средневозрастных лишайниково-зеленомошных лесах, формирующихся на вырубках и гарях в Мурманской обл. В районах исследований была выявлена тенденция увеличения ежегодного линейного прироста сосны в высоту до 55–65-летнего возраста, а в последние годы – его стабилизация. Интенсивность прироста по диаметру у сосны обыкновенной снижается во всем исследованном возрастном интервале. В составе надземной фитомассы сосновых древостоев древесина стволов составляет 58–67%, живые неохвоенные ветви – 15.4–17.3%, охвоенные побеги вместе с хвоей – 21–24%. Масса сухих ветвей варьирует в пределах 1.4–3.6%. Установлена тесная линейная связь таксационных показателей деревьев с их фитомассой. Максимальная теснота связи выявлена для фракции ствола, минимальная – для сухих ветвей.
Интенсивное освоение природных ресурсов Европейского Севера создает постоянную и непрерывно возрастающую нагрузку на северотаежные леса, которые выполняют важнейшие природоохранные и средостабилизирующие функции. Особенно это актуально для сосновых лесов, которые преобладают на территории Европейского Севера и повсеместно вовлекаются в использование народным хозяйством. В довольно широком спектре экологических условий сосняки являются устойчивыми, саморегулирующимися экосистемами, в которых идет постоянная смена поколений, хотя и значительно более растянутая во времени, чем в южных районах. Однако рубки и пожары приводят к нарушению этого природного равновесия.
На бо́льшей части территории Мурманской обл. лесная растительность не образует больших сплошных массивов. Леса чередуются с озерами, болотами, участками тундровой растительности в верхнем поясе низких глыбовых гор. В биологии сосны обыкновенной, обитающей в Мурманской обл., имеются некоторые особенности по сравнению с сосной, произрастающей в более южных частях таежной зоны. Она отличается более медленным ростом в высоту и по диаметру, сжатостью фенофаз, способностью начинать рост при относительно низких положительных температурах, более продолжительным функционированием хвои на деревьях, слабым и редким плодоношением [1–3].
Исследования сосновых лесов на Европейском Севере имеют немалую историю. Однако, как и во многих других районах Севера, изученность сосняков Мурманской обл. крайне неоднородна. Как справедливо отмечалось исследователями [4, 5], наряду с обстоятельным рассмотрением вопросов флористического и геоботанического плана, классификации сообществ и вырубок, достаточно глубоким анализом закономерностей естественного возобновления сосновых лесов, много описательных публикаций, иногда весьма поверхностных, посвященных частным аспектам, мало относящихся к насущным проблемам лесоведения.
На бо́льшей части площадей эксплуатационного назначения в Мурманской обл. сосняки лишайниковые и лишайниково-зеленомошные сведены рубками. На их месте сформировались относительно густые молодняки. Количество деревьев в молодом возрасте может достигать 30–40 тыс. шт./га и более [3]. В настоящее время первостепенное значение, наряду с нерешенными вопросами охраны лесов от пожаров и совершенствования процессов лесопользования, на наш взгляд, приобретают научные разработки по лесовосстановлению и формированию насаждений на нарушенных территориях (вырубки, гари), изучению строения и роста сосновых древостоев, их продуктивности.
Цель настоящей работы состояла в изучении особенностей роста и формирования надземной фитомассы сосны обыкновенной Pinus sylvestris L. в сосняках лишайниковых и лишайниково-зеленомошных, естественно восстанавливающихся на вырубках и гарях в Мурманской обл.
МАТЕРИАЛ И МЕТОДЫ
Исследованные сосновые леса расположены в трех районах (Уполокшском, Ено-Ковдорском, Ливском) на территории Мурманской обл. (Кольский п-ов). Древостои являются одновозрастными или условно одновозрастными, одноярусными, реже с глубоким высотным профилем без выраженности ярусов, преимущественно чистые или почти чистые. Примесь березы пушистой Betula pubescens Ehrh. составляет не более 10%. Встречаются также разновозрастные древостои, формирование которых идет при часто повторяющихся пожарах. В табл. 1 приведена краткая характеристика исследованных древостоев.
Таблица 1.
Краткая характеристика древостоев исследованных лишайниково-зеленомошных сосновых лесов в Мурманской обл. Table 1. Stand description in studied lichen-green moss Scots pine forests in the Murmansk region
| №№ ППП №№ PSP |
Районы исследований Study area |
Состав древостоя Stand composition |
Таксационные характеристики Stand characteristics |
||||
|---|---|---|---|---|---|---|---|
| высота, м height, m | диаметр, см diameter, cm | число стволов, экз./га density, ind./hа |
возраст, лет age, years |
класс бонитета capacity class |
|||
| 1 | Уполокшский Upolokshsky |
10 С + Б 10 P + B |
3.9 | 4.9 | 1512 | 50 | V |
| 2 | 10 С 10 P |
8.5 | 10.1 | 2804 | 60 | IV–V | |
| 3 | 10 С 10 P |
8.0 | 8.3 | 5273 | 60 | IV–V | |
| 4 | Ено-Ковдорский Eno-Kovdorsky |
10 С 10 P |
8.7 | 8.3 | 2290 | 70 | V |
| 5 | 10 С + Б 10 P + B |
13.5 | 12.6 | 2300 | 70 | V | |
| 6 | 10 С 10 P |
10.4 | 16.1 | 575 | 80 | Va | |
| 7 | Ливский Livsky |
10 С 10 P |
9.1 | 10.6 | 1336 | 70 | IV–V |
| 8 | 10 С + Б 10 P + B |
8.8 | 9.0 | 1386 | 70 | IV–V | |
В средневозрастных лишайниково-зеленомошных сосновых лесах доминирующими видами в травяно-кустарничковом ярусе являются: Vaccinium myrtillus L., V. vitis-idaea L., Empetrum hermaphroditum Hagerup. Мохово-лишайниковый ярус формируют лишайники рода Cladonia: C. stellaris (Opiz.) Brodo, C. rangiferina (L.) F.H. Wigg, C. mitis Sandst. Среди мхов чаще всего встречаются Pleurozium schreberi (Brid.) Mitt., Polytrichum juniperinum Hedw., виды рода Dicranum Hedw.
В районах исследований преобладают хорошо водопроницаемые, сильно завалуненные, песчаные моренные и водно-ледниковые отложения, на которых развиваются подзолы иллювиально-железистые ненасыщенные, мелкоподзолистые, иллювиально-малогумусовые, средне скелетные [6, 7]. Отличительной чертой лесных почв районов исследований является отсутствие многолетней мерзлоты. По особенностям термического режима эти почвы относятся к типу длительно сезонно промерзающих.
В настоящей работе рассмотрены данные, полученные на 8-ми постоянных пробных площадях (ППП), размером 0.15–0.20 га, на каждой из которых проводился сплошной перечет деревьев по элементам леса и ступеням толщины; измерялись параметры кроны деревьев и их состояние; определялся возраст хвои [8]. Сухостойные деревья учитывались отдельно. При среднем диаметре древостоя до 10 см ступени толщины принимались равными 1 см, от 10 до 20 см – 2 см. Высоты измеряли при помощи высотомера SUUNTO у 20–30 живых деревьев, отобранных случайным образом. По этим данным строили графики зависимости высот деревьев от их диаметров, которые использовали для определения средней высоты древостоя. По выполненным измерениям рассчитывали таксационные показатели исследованных древостоев.
Для определения возраста и величины радиального прироста древесины у модельных деревьев (10–12 экз. на каждой ППП) отбирали образцы древесины (керны) буравом Пресслера или спилы ствола. Ширину годичных колец определяли на приборе LINTAB-6. Анализ данных по динамике радиального прироста сосны обыкновенной основывался на современных принципах и методах, используемых отечественными и зарубежными авторами [9–11].
Для определения фитомассы деревьев и древостоев подбирались модельные деревья. Их выборка формировалась в соответствии с рядами распределения деревьев по диаметру. На каждой ППП отбирались по 10–12 модельных деревьев, имеющих близкие к средним значения диаметра, высоты и размеров кроны для каждой ступени толщины в пределах всего диапазона варьирования диаметров. После предварительного описания и измерений модельные деревья спиливали на уровне корневой шейки и аккуратно укладывали на брезент. Затем модели размечали и измеряли линейные приросты центрального и боковых побегов, расстояние до начала кроны, общую длину ствола, диаметр ствола через 0.5 или 1 м.
Оценка вертикально-фракционной структуры надземной фитомассы древостоев сосны обыкновенной на каждой пробной площади выполнялась с выделением следующих фракций: ствол, живые неохвоенные ветви, охвоенные ветви, сухие ветви. Фитомасса структурных частей кроны определялась после ее деления на 3 или 4 одинаковые по длине секции. Из общей массы охвоенных ветвей отбирались навески для установления соотношения массы хвои и побегов. Самым распространенным в лесоэкологических исследованиях методом выравнивания эмпирических данных массы модельных деревьев в зависимости от их таксационных показателей является степенная функция преобразованная логарифмированием к линейному виду, аргументом которой служит произведение квадрата диаметра ствола на высоте 1.3 м от корневой шейки на высоту дерева [12].
При обработке количественных данных использованы методы описательной статистики, корреляционный и регрессионный анализы.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
В исследованных средневозрастных сосновых лесах густота древостоя варьирует в пределах от 575 до 5273 экз./га (табл. 1). Строение исследованных сосняков может быть охарактеризовано на основе анализа распределения особей сосны обыкновенной по ступеням толщины (табл. 2).
Таблица 2.
Распределение деревьев сосны обыкновенной по 2-х сантиметровым ступеням толщины в средневозрастных лишайниково-зеленомошных сосновых лесах Мурманской обл. Table 2. Distribution of Scots pine trees by 2 cm diameter intervals in medium-aged lichen-green-moss pine forests in the Murmansk region
| №№ ППП №№ PSP |
Ступени толщины деревьев, см Diameter classes, cm |
|||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 4 | 6 | 8 | 10 | 12 | 14 | 16 | 18 | 20 | 22 | |
| Количество деревьев, экз./га Number of trees, ind./ha |
||||||||||
| 1 | $\frac{{575}}{{58.1}}$ | $\frac{{238}}{{24.1}}$ | $\frac{{150}}{{15.2}}$ | $\frac{{13}}{{1.3}}$ | $\frac{{13}}{{1.3}}$ | – | – | – | – | – |
| 6 | – | $\frac{{15}}{{2.8}}$ | $\frac{{20}}{{3.7}}$ | $\frac{{65}}{{12.1}}$ | $\frac{{95}}{{17.8}}$ | $\frac{{115}}{{21.5}}$ | $\frac{{75}}{{14.0}}$ | $\frac{{60}}{{11.2}}$ | $\frac{{65}}{{12.1}}$ | $\frac{{25}}{{4.8}}$ |
| 4 | $\frac{{484}}{{21.1}}$ | $\frac{{384}}{{16.8}}$ | $\frac{{468}}{{20.4}}$ | $\frac{{217}}{{9.5}}$ | $\frac{{351}}{{15.3}}$ | $\frac{{184}}{{8.0}}$ | $\frac{{84}}{{}}3.7$ | $\frac{{67}}{{2.9}}$ | $\frac{{34}}{{1.5}}$ | $\frac{{17}}{{0.8}}$ |
| 5 | – | $\frac{{50}}{{2.2}}$ | $\frac{{400}}{{17.4}}$ | $\frac{{575}}{{25.0}}$ | $\frac{{400}}{{17.4}}$ | $\frac{{425}}{{18.5}}$ | $\frac{{300}}{{13.0}}$ | $\frac{{50}}{{2.2}}$ | $\frac{{50}}{{2.2}}$ | $\frac{{50}}{{2.2}}$ |
| 7 | $\frac{{217}}{{16.2}}$ | $\frac{{184}}{{13.8}}$ | $\frac{{267}}{{20.0}}$ | $\frac{{267}}{{20.0}}$ | $\frac{{167}}{{12.5}}$ | $\frac{{117}}{{8.8}}$ | $\frac{{50}}{{3.7}}$ | $\frac{{33}}{{2.5}}$ | $\frac{{17}}{{1.3}}$ | $\frac{{17}}{{1.3}}$ |
| 8 | $\frac{{100}}{{7.2}}$ | $\frac{{234}}{{16.9}}$ | $\frac{{534}}{{38.5}}$ | $\frac{{234}}{{16.9}}$ | $\frac{{134}}{{9.7}}$ | $\frac{{134}}{{9.6}}$ | – | $\frac{{17}}{{1.2}}$ | – | – |
Показатели асимметрии таких рядов распределения на всех исследованных участках характеризуются положительными значениями, что согласуется с мнением исследователей о строении модальных древостоев во многих других районах лесной зоны [4, 13–16]. При общих положительных значениях этого показателя выровненные кривые рядов распределения в исследованных сосновых древостоях Мурманской обл. существенно различаются между собой. Ближе всего к нормальному ряду распределения кривые в древостоях III–IV классов возраста. Самые большие отклонения от нормального ряда отмечены в молодняках сосны I–II классов возраста, где наблюдаются интенсивные процессы дифференциации особей в древесном ярусе. В большинстве исследованных сообществ распределение деревьев по ступеням толщины соответствует или приближается к нормальному, что свидетельствует об одновозрастности исследованных сосновых древостоев.
Процессы возобновления сосны, оцениваемые по числу подроста на лишайниковых вырубках и гарях, обычно признаются удовлетворительными, однако разовые оценки, какими бы детальными не были обследования, и какую бы численность подроста они не фиксировали, еще не являются окончательными. Они отражают лишь текущее состояние лесовозобновления. Многолетние наблюдения на вырубках и гарях свидетельствуют о затяжном процессе естественного лесовозобновления [17]. Эти процессы в борах-беломошниках Северо-Востока, например, могут затягиваться на 20 и более лет [18]. Среди факторов, затрудняющих возобновление лесов на северном пределе распространения, необходимо отметить редкую периодичность плодоношения деревьев, низкую всхожесть семян, повышенную гибель молодых растений вследствие неблагоприятных климатических и почвенно-гидрологических условий [3, 19]. К этому можно добавить развитый напочвенный покров на вырубках, препятствующий проникновению семян к поверхности почвы и их прорастанию. Лесовозобновительный процесс можно считать успешным при формировании на вырубках и гарях относительно устойчивого поколения сосны. Это характерно практически для всех обследованных нами средневозрастных древостоев сосны обыкновенной.
На вырубках и гарях лишайниковых сосняков формируются как густые, так и разреженные древостои. Последние характеризуются сравнительно медленным ростом в высоту, умеренно густые сосняки – более быстрым. Необходимо отметить, что изменчивость диаметров, высот, относительных высот, объемов крон, а также редукционных чисел по высоте и диаметру в перегущенных древостоях меньше, чем в редкостойных сосняках. Древостои, формирующиеся из подроста предварительных генераций, в возрасте 25–40 лет отличаются наиболее медленным ростом в высоту. Это связано с тем, что предварительный подрост определенное время испытывал жесткую конкуренцию со стороны материнского древостоя в надземной части за свет, а в зоне корневых систем за элементы минерального питания. Под пологом особи подроста постепенно превращались в сильно угнетенные, отстающие в росте деревца, которые долгое время не могут восстановить свою нормальную форму даже после удаления материнского древостоя. Сходные результаты были получены при исследовании сосновых молодняков в Карелии [20].
Под пологом исследованных сосняков лишайниковых и лишайниково-зеленомошных количество подроста сосны в возрасте от 15 до 45 лет, высотой от 19 до 152 см (ср. высота 60.6 см) составляет в среднем около 3 тыс. экз./га. В древостоях с полнотой 0.4–0.6 и выше, которые в течение последних 15–20 лет не подвергались хозяйственным воздействиям или низовым пожарам, создается высокий уровень корневой конкуренции и появившийся здесь подрост в течение многих лет находится в угнетенном состоянии, а затем постепенно отмирает.
Проведенное нами изучение корневых систем [21, 22] показало, что при более высокой густоте древостоя, благодаря наличию якорных корней, сосна полнее использует глубокие горизонты почвы, а при редком расположении деревьев – верхний почвенный слой (подстилку), в котором создается значительное корневое насыщение, оказывающее затем отрицательное влияние на рост и формирование нового поколения. Последнее мы наблюдали на многих участках, особенно на старых, плохо возобновившихся гарях с наличием отдельных старовозрастных деревьев и хорошо развитым напочвенным покровом [23].
При оценке жизненного состояния деревьев и древостоев сосны важнейшими показателями являются сведения о хвое, охвоенности побегов, степени развития крон. Продолжительность жизни хвои Р. sylvestris на отдельных участках находится в пределах от 5.7 до 6.7 лет, доля здоровой хвои составляет 95–100%. Комплексная оценка состояния средневозрастных древостоев в сосняках лишайниковых и лишайниково-зеленомошных в Мурманской обл. позволила установить, что здоровые особи сосны обыкновенной в древостоях составляют 65–70%, на долю ослабленных и сильно ослабленных приходится в среднем по 15%, на долю сухих особей – менее 5%. Такой характер распределения по категориям жизненного состояния обусловлен конкурентным взаимодействием особей сосны, особенно в зоне корневых систем, и усиливающимися с возрастом процессами их дифференциации.
Одним из интегральных показателей состояния и ресурсного потенциала формирующихся сосняков на вырубках и гарях является их рост: линейный прирост центрального и боковых побегов, прирост ствола по диаметру и, наконец, рост и развитие корневых систем. При удалении древесного яруса в результате рубки или пожара резко изменяются экологические условия в надземной сфере: увеличивается освещенность, уровень увлажнения и прогреваемость верхних горизонтов почвы, существенно изменяется состояние напочвенного покрова (он полностью уничтожается при низовых пожарах, частично повреждается при лесозаготовках, при этом отдельные теневыносливые виды снижают свое обилие). В подземной сфере резко снижается конкуренция за влагу и элементы питания. В таких условиях молодые растения сосны активно осваивают свободные экологические ниши, достаточно быстро растут и развиваются, несмотря на большую во многих случаях густоту (несколько десятков тысяч на 1 га), бедность и сухость маломощных, завалуненных почв. Ранее нами было установлено [24], что скелетная основа корневой системы сосны обыкновенной в условиях Кольского п-ова проявляет характерные морфологические особенности уже к 15–20 годам. Корни интенсивно распространяются к этому времени на 6–7, нередко 8–10 м от ствола.
В конце 1980-х гг. в условиях Кольского п-ова нами проводились анализ и оценка погодичной динамики прироста в высоту 35–40-летних древостоев сосны (рис. 1). При этом было установлено, что интенсивность линейного прироста центральных побегов сосны в исследованных сосняках еще не достигала своего максимума [24]. Исследуя сосняки Кольского п-ова в 1950–60-х гг., отмечалось [25], что текущий прирост в высоту деревьев увеличивается в среднем до 35-летнего возраста. В северной Карелии [26] и на юге Мурманской обл. [20] этот период в зависимости от типа леса оценивался в 40–50 лет.
Рис. 1.
Динамика прироста в высоту стволов сосны обыкновенной в 35-летних сосняках лишайниковых на Кольском полуострове [24]. По горизонтали – годы; по вертикали – линейный прирост, см. Fig. 1. Dynamics of Scots pine height growth in 35-year old lichen pine forests in the Kola Peninsula [24]. X-axis – years; y-axis – height growth, cm.
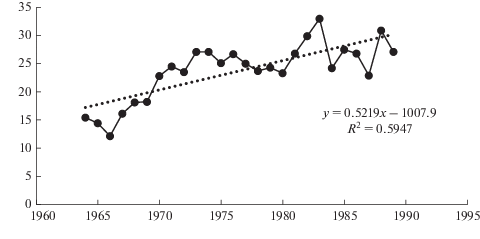
В ходе наших последних исследований в средневозрастных сосняках лишайниково-зеленомошных, имеющих возраст 60–70 лет, была выявлена тенденция последовательного увеличения ежегодного линейного прироста сосны в высоту с некоторым замедлением в последние годы (рис. 2). Для подтверждения предварительных выводов мы разбили кривые прироста на временные отрезки, которые определяет сам их ход. В результате были получены следующие количественные данные: Ено-Ковдорский р-н: 1989–2008 гг. – линейный рост (P < 0.001, R2 = 0.647) от ~16 до 32 см год–1, 2009–2014 г. – плато на среднем уровне 27 ± 1 см год–1. Ливский р-н: 1989–2008 гг. – линейный рост от ~19 до 28 см год–1 (P < 0.001, R2 = 0.517), 2009–2014 – плато на среднем уровне 22.4 ± 0.5 см год–1. В р-не Уполокши в течение всего рассматриваемого периода (1989–2014 гг.) величина прироста оставалась постоянной на среднем уровне 19.4 ± ± 0.6 см год–1. В Ено-Ковдорском и Ливском р-нах в период 1989–2008 гг. возрастной тренд объясняет, как следует из величины коэффициента детерминации, 52–65% варьирования прироста в высоту. Если обобщать данные по этим трем районам, то можно заметить, что с 2009 г. прирост в высоту во всех случаях остается стабильным: в Ливском и Уполокшском р-нах на среднем уровне около 20 см год–1, в Ено-Ковдорском – на несколько более высоком уровне – 27 см год–1. Таким образом, интенсивность прироста центральных побегов сосны обыкновенной стабилизируется к 55–65 годам и связано это, скорее, не с изменением погодных условий, а с закономерностями динамики ростовых процессов у исследуемого вида в изученных сообществах.
Рис. 2.
Динамика прироста в высоту стволов сосны обыкновенной в 60–80-летних сосняках лишайниково-зеленомошных в Ено-Ковдорском (кружки), Уполокшском (квадраты) и Ливском (треугольники) р-нах Мурманской обл. По горизонтали – годы; по вертикали – линейный прирост, см. Fig. 2. Dynamics of Scots pine height growth in 60–80 year old lichen-green moss pine forests in Eno-Kovdor (circles), Upoloksha (squares) and Liva (triangules) districts of the Murmansk region. X-axis – years; y-axis – height growth, cm.
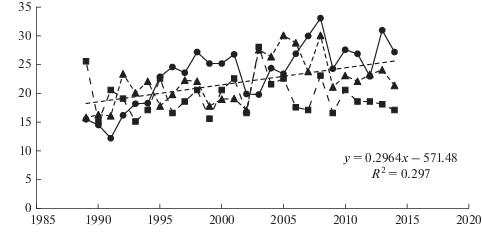
Одной из важнейших интегральных характеристик состояния и ресурсного потенциала древостоев является радиальный прирост стволов. В этой характеристике проявляются результаты конкуренции между деревьями, находят отражение антропогенные воздействия на лесные экосистемы (рубки, пожары, мелиорация), а также другие процессы, протекающие в природных сообществах [24, 27, 28]. Нами были получены серии абсолютных значений ширины годичных слоев сосны обыкновенной, позволяющие оценить динамику радиального прироста сосны в средневозрастных древостоях Уполокшского, Ено-Ковдорского и Ливского р-нов Мурманской обл. (рис. 3). Несмотря на удаленность друг от друга исследуемых сообществ и разную густоту древостоев, прослеживается достаточно высокая синхронность погодичной изменчивости приростов и близкие их значения. Интенсивность прироста по диаметру у сосны обыкновенной продолжает снижаться в исследованном временном интервале, не достигая стабильности, в отличие от прироста в высоту. Общую тенденцию динамики в период от 1980 до 2014 года отражает отрицательная линейная функция (рис. 3). При этом величина прироста снижается в среднем в 2–2.5 раза: от 0.8–1.1 мм год–1 до 0.3–0.5 мм год–1. В трех исследованных районах эта функция объясняет в среднем ~20% варьирования прироста по диаметру. Остальное варьирование происходит за счет других факторов. Анализ кривых абсолютных значений радиального прироста сосны обыкновенной показывает, что у особей во всех исследованных сообществах выражены кратковременные циклы флуктуаций прироста по диаметру. Периоды депрессий и экспрессий прироста охватывают ряд лет с четко выраженными минимумами и максимумами, приходящимися на определенные годы. Они, как правило, имеют синхронный характер в разных районах и заметно различаются по амплитудам. Связано это, скорее всего, на наш взгляд, с флуктуациями климатических составляющих.
Рис. 3.
Динамика радиального прироста сосны обыкновенной в 60–80-летних сосняках лишайниково-зеленомошных в Ено-Ковдорском (кружки), Уполокшском (квадраты) и Ливском (треугольники) р-нах Мурманской обл. По горизонтали – годы; по вертикали – прирост по диаметру, мм. Fig. 3. Dynamics of Scots pine radial increment in 60–80 year old lichen-green moss pine forests in Eno-Kovdor (circles), Upoloksha (squares) and Liva (triangules) districts of the Murmansk region. X-axis – years; y-axis – radial increment, mm.

Еще одним важным интегральным показателем, отражающим состояние лесных экосистем и степень их антропогенной нарушенности, является биологическая продуктивность древостоев, определяемая методами традиционной и весовой таксации [16]. В настоящее время фитомасса лесов рассматривается как их основная характеристика, отражающая ход процессов в лесных экосистемах и используемая в целях экологического мониторинга, изучения структуры и биоразнообразия лесного покрова, моделирования продуктивности лесов, устойчивого ведения лесного хозяйства с учетом глобальных изменений.
Несмотря на важность изучения фитомассы в лесах, количество эмпирических данных остается незначительным из-за трудоемкости работ. В лесах Мурманской обл. до начала наших исследований такие работы практически не проводились. Между тем, реальный прогресс в оценке биологической продуктивности лесов возможен только на основе достаточного фактологического материала.
Ранее мы отмечали [24], что при выравнивании эмпирических данных массы и продукции модельных деревьев в зависимости от их таксационных показателей исследователи пользуются лишь степенной функцией, преобразованной логарифмированием к линейному виду, аргументом которой служит произведение квадрата диаметра на высоте груди на высоту дерева. Такой вид выравнивания является самым распространенным в лесоэкологических исследованиях [12, 29]. В процессе обработки фактических данных была произведена оценка связи таксационных показателей с продуктивностью молодых сосновых древостоев на Кольском Севере с использованием нескольких функций [30]. В результате было установлено, что использование линейного уравнения вида y = ax + b для аппроксимации данных по фракционному составу фитомассы сосны обыкновенной дает вполне удовлетворительные результаты. Анализ связи общей надземной фитомассы и ее фракций с таксационными характеристиками модельных деревьев сосны обыкновенной в средневозрастных древостоях разных районов Мурманской обл. (рис. 4) подтверждает этот вывод. Максимальная теснота связи выявлена для массы ствола, минимальная – для массы сухих ветвей. Использование этой модели позволяет легко и достаточно надежно получать вероятные значения фитомассы древостоев сосны обыкновенной.
Рис. 4.
Зависимость фитомассы ствола (a), живых ветвей (b), сухих ветвей (c) и общей надземной фитомассы (d) сосны обыкновенной от таксационных характеристик деревьев в 60–70-летних древостоях Мурманской обл. По горизонтали – произведение высоты (H) на квадрат диаметра (D2) модельных деревьев; по вертикали – фитомасса, кг АСВ. Fig. 4. Dependence of trunk (a), live branches (b) and dry branches (c) phytomass and total above-ground Scots pine phytomass (d) on the tree mensuration parameters in 60–70-year-old stands of the Murmansk region. X-axis – height (H) multiplied by the square of diameter (D2) of the model trees; y-axis – phytomass, absolute dry weight, kg.
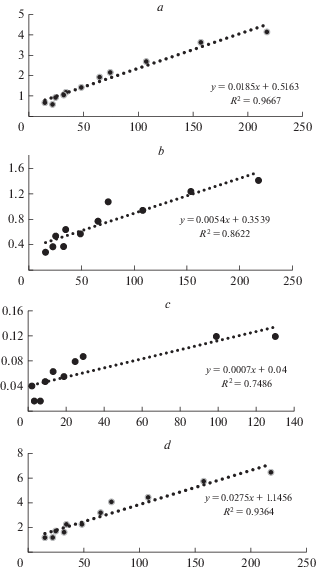
Согласно полученным данным, в исследованных сообществах на древесину ствола приходится 58–67% от общей надземной фитомассы (табл. 3). Фитомасса ветвей – один из важнейших компонентов – занимает второе место после стволовой древесины. Живые неохвоенные ветви в средневозрастных сообществах сосны обыкновенной составляют 15.4–17.3% от общей надземной фитомассы, а доля охвоенных побегов вместе с хвоей достигает 21–24%. Наибольшая доля ветвей отмечена у сосны обыкновенной в древостоях с пониженной полнотой. Фитомасса сухих ветвей в исследованных древостоях варьирует в пределах 1.4–3.6% от фитомассы надземной части.
Таблица 3.
Структура надземной фитомассы сосны обыкновенной в средневозрастных древостоях сосны обыкновенной в Мурманской обл. (в расчете на 1 модельное дерево) Table 3. Structure of aboveground phytomass of Scots pine in the middle-aged stands in the Murmansk region (per 1 model tree)
| Районы исследований Study area |
Доля в общей надземной фитомассе, % Share in total aboveground phytomass, % |
|||
|---|---|---|---|---|
| ствол trunk |
живые ветви live branches |
охвоенные побеги foliated shoots |
сухие ветви dry branches |
|
| Уполокшский Upolokshsky |
66.6 | 17.3 | 13.1 | 3.0 |
| Ено-Ковдорский Eno-Kovdorsky |
59.9 | 15.4 | 21.1 | 3.6 |
| Ливский Livsky |
58.1 | 16.3 | 24.2 | 1.4 |
| В среднем On the average |
61.5 | 16.3 | 19.5 | 2.7 |
ЗАКЛЮЧЕНИЕ
Проведенные обследования сосновых лесов на территории Мурманской обл. показали, что формирующиеся на вырубках и гарях древостои по многим показателям заметно отличаются от древостоев-эталонов. Неодинаковая густота и возрастная структура древостоев обусловливают существенные различия в ходе роста. Самым медленным ростом в высоту в возрасте 25–40 лет отличаются древостои, формирующиеся из подроста предварительных генераций. Сравнительно медленным ростом в высоту характеризуются разреженные сосняки, более быстрым – умеренно густые.
В средневозрастных сосняках лишайниково-зеленомошных выявлена тенденция постепенного увеличения ежегодного линейного прироста в высоту сосны до 55–65-летнего возраста. В дальнейшем прирост в высоту сосны обыкновенной в Мурманской обл. перестает увеличиваться. Радиальный прирост сосны обыкновенной имеет максимальные значения в первые 10–15 лет после рубок и пожаров, затем наблюдается его постепенный спад. Эта тенденция характерна для подавляющего большинства лесных насаждений. Наблюдаются заметные различия в характере динамики радиального прироста и прироста в высоту. Интенсивность прироста по диаметру сосны обыкновенной в средневозрастных лесах последовательно снижается, в то время как интенсивность прироста в высоту до 55–65 лет продолжает увеличиваться.
Для исследованных сообществ характерна тесная линейная связь таксационных показателей деревьев с их фитомассой. Максимальная теснота связи с таксационными показателями выявлена для массы ствола, минимальная – для массы сухих ветвей. Оценка запасов и фракционного распределения общей надземной фитомассы сосны обыкновенной в изученных средневозрастных сосновых лесах показала, что на древесину стволов приходится 58–67%. Живые неохвоенные ветви составляют 15.4–17.3%, доля охвоенных побегов вместе с хвоей достигает 21–24%. Наибольшая масса ветвей отмечена у сосны обыкновенной в древостоях с пониженной полнотой. Фитомасса сухих ветвей варьирует в пределах 1.4–3.6% от фитомассы надземной части.
Список литературы
Максимов В.А. 1967. Строение и ход роста насаждений сосны лапландской в Мурманской области. – Вопросы таежного лесоводства на Европейском Севере. М. С. 79–86. https://www.booksite.ru/fulltext/rusles/voprostae/text.pdf
Правдин Л.Ф. 1964. Сосна обыкновенная. М. 189 с.
Цветков В.Ф., Семенов Б.А. 1985. Сосняки Крайнего Севера. М. 116 с.
Цветков В.Ф. 2002. Сосняки Кольской лесорастительной области и ведение хозяйства в них. Архангельск. 379 с. https://www.booksite.ru/fulltext/rusles/sosn/text.pdf
Цветков В.Ф., Семенов Б.А. 2012. Современные проблемы притундровых лесов. – Материалы Всероссийской конференции с международным участием “Современные проблемы притундровых лесов”, 4–9 сентября 2012 г. Архангельск. С. 5–14. https://narfu.ru/university/library/books/0685.pdf
Никонов В.В., Переверзев В.Н. Почвообразование в Кольской Субарктике. Л. 1989. 168 с.
Шишов Л.Л., Тонконогов В.Д., Лебедева И.И., Герасимова М.И. 2004. Классификация и диагностика почв России. Смоленск. 342 с. https://www.rfbr.ru/rffi/ru/books/o_28031#7
Методы изучения лесных сообществ. 2002. СПб. 240 с.
Fritts H.C. 1976. Tree rings and climate. London, New York, San Francisco. 582 p. https://doi.org/10.1016/B978-0-12-268450-0.X5001-0
Юкнис Р.А. 1990. Рост и продуктивность одновозрастных сосняков в условиях загрязненной природной среды. Автореферат дисс.… д-ра биол. наук. Красноярск. 40 с.
Усольцев В.А. 2010. Фитомасса и первичная продукция лесов Евразии. Екатеринбург. 573 с. https://elar.usfeu.ru/bitstream/123456789/2606/1/Usoltsev_Fitomassa.pdf
Ильина Н.А., Рождественский С.Г., Уткин А.И., Гульбе. Я.И. Каплина Н.Ф., Арутюнян С.Г. 1986. Сравнение разных функций, используемых при аппроксимации эмпирических данных модельных деревьев. – Вертикально-фракционное распределение фитомассы в лесах. М. С. 14–18.
Моисеев В.С. 1971. Таксация молодняков. Л. 343 с.
Луганский Н.А. 1974. Научное обоснование способов возобновления и формирования молодняков на вырубках в сосновых лесах Урала: автореф. дис. … д-ра биол. наук. Алма-Ата. 56 с.
Бузыкин А.И., Пшеничникова Л.С. 1978. Фитомасса и особенности ее продуцирования деревьями разного ценотического положения. – Продуктивность сосновых лесов. М. С. 69–89.
Усольцев В.А., Воробейчик Е.Л., Бергман И.Е. 2012. Биологическая продуктивность лесов Урала в условиях техногенного загрязнения. Исследование системы связей и закономерностей. Екатеринбург. 366 с. https://elar.usfeu.ru/bitstream/123456789/458/5/Usolcev_Vorobeycshik.pdf
Цветков В.Ф. 1989. Формирование сосняков Кольского полуострова в связи со сплошными рубками. – В сб.: Динамическая типология лесов. Сб. научн. трудов. М. С. 119–143.
Листов А.А. 1986. Боры-беломошники. М. 191 с.
Барабин А.И. 2012. Прогнозирование урожаев семян хвойных на Европейском Севере. – Современные проблемы притундровых лесов. Материалы Всероссийской конференции с междунар. Участием, 4–9 сентября 2012 г., Архангельск. Сев (Арк.) федер. ун-т им. М.В. Ломоносова. С. 313–317. https://narfu.ru/university/library/books/0685.pdf
Зябченко С.С. Иванчиков А.А. 1978. Зональные особенности формирования сосняков черничных Карелии и Кольского полуострова и динамика структуры растительной массы в них. – Формирование и продуктивность сосновых насаждений Карельской АССР и Мурманской области. Петрозаводск. С. 30–75.
Ярмишко В.Т., Цветков В.Ф. 1987. Строение, запасы и распределение в почве корневых систем растений в сообществах сосновых молодняков Кольского полуострова. – Бот. журнал. 2(4): 496–505.
Ярмишко В.Т., Лумме И., Ярмишко М.А. 2007. Реакция тонких корней Pinus sylvestris L. на изменение среды их обитания на Карельском перешейке и юго-восточной Финляндии. – В сб: Биоразнообразие, охрана и рациональное использование растительных ресурсов Севера: Материалы XI Перфильевских научных чтений. Архангельск. С. 93–98.
Динамика лесных сообществ Северо-Запада России. 2009. СПб. 276 с.
Ярмишко В.Т. 1997. Сосна обыкновенная и атмосферное загрязнение на Европейском Севере. СПб.: НИИ химии СПбГУ. 210 с.
Максимов В.А. 1963. Составление эскизов таблиц хода роста сосновых насаждений Мурманской области. – Сборник статей по обмену произв.-техн. опытом по лесн. хозяйству и лесоустройству. Л. С. 53–91.
Казимиров Н.И., Кабанов В.В., Преснухин Ю.В. 1978. Ход роста сосновых насаждений северной Карелии. – Формирование и продуктивность сосновых насаждений Карельской АССР и Мурманской области. Петрозаводск. С. 102–107.
Yarmishko V.T. 2015. Radial growth of Scots pine (Pinus sylvestris L.) in varied environment influenced by air pollution in the European North of Russia. – Forestry Ideas. 21(2): 375–383. https://forestry-ideas.info/issues/issues_Download.php?download=260
Ярмишко В.Т., Игнатьева О.В. 2019. Многолетний импактный мониторинг состояния сосновых лесов в центральной части Кольского полуострова. – Известия РАН. Серия биологическая. 6: 658–668. https://doi.org/10.1134/S0002332919060134
Биологическая продуктивность лесов Поволжья. 1982. М. 282 с.
Лянгузова И.В., Ярмишко В.Т. 1990. Использование некоторых корреляционных уравнений для выражения связи таксационных показателей с продуктивностью древостоя в фоновых условиях и при атмосферном загрязнении. – Всесоюзн. cовещание “Проблемы лесоведения и лесной экологии”. Тез. докладов. Минск. С. 76–77.
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы