

Растительные ресурсы, 2021, T. 57, № 1, стр. 58-67
Эколого-биологические особенности Dactylorhiza incarnata (Orchidaceae) в луговых фитоценозах Центрального Кавказа
В. А. Чадаева 1, *, Г. А. Кярова 1
1 Институт экологии горных территорий им. А.К. Темботова РАН
г. Нальчик, Россия
* E-mail: v_chadayeva@mail.ru
Поступила в редакцию 13.05.2020
После доработки 13.08.2020
Принята к публикации 10.12.2020
Аннотация
В 2015–2019 гг. на территории Кабардино-Балкарской Республики изучены изменчивость морфологических признаков особей, возрастная структура и жизненность 14 ценопопуляций Dactylorhiza incarnata (L.) Soó. Исследования проведены в луговых фитоценозах при разном режиме антропогенной нагрузки (рекреация, выпас скота). Для вида характерна высокая пластичность (Ip = 41–87%) и изменчивость ($C{{V}_{{\bar {х}\,{\text{ср}}}}}$ = 21.4–65.1%) морфологических признаков при изменении условий произрастания. В составе ненарушенных сильно увлажненных лугов наблюдается интенсификация ростовых процессов особей с максимальным повышением жизненности ценопопуляций (IVC = 1.18–1.23). Возрастает доля генеративных особей в возрастных спектрах (42.3–57.5%), отмечено эффективное возобновление ценопопуляций. Усиление антропогенного воздействия (рекреация, выпас скота) приводит к снижению жизненности ценопопуляций (IVC = 0.82–1.12) и доли генеративных растений (8.8–29.1%), накоплению в возрастных спектрах виргинильных особей (34.2–42.7%). При этом на щебнистых субстратах с низким проективным покрытием травостоя (70–80%) высока интенсивность возобновления ценопопуляций, плотность особей в которых достигает 45.1–58.2 особ./м2.
Изучение эколого-биологических особенностей растений позволяет определить наиболее оптимальные условия для их произрастания, установить ограничивающие факторы, выявить адаптационные механизмы и стратегии жизни видов в природе. Это важный аспект сохранения биологического разнообразия и рационального использования растительных ресурсов, особенно актуальный для редких и охраняемых видов. В то же время, оценка состояния популяций редких видов растений может служить инструментом биоиндикации степени нарушенности конкретных фитоценозов.
Среди редких видов флоры многих регионов мира – представители семейства Orchidaceae Juss., имеющие большое хозяйственное значение как лекарственные, декоративные растения, подверженные высокой антропогенной нагрузке (сбор на букеты, в качестве лекарственного сырья, нарушение местообитаний и т.д.) [1]. Редкость видов семейства Orchidaceae обусловлена также особенностями их биологии и экологии: редукцией зародышевых структур, наличием мелких семян, высокой специализацией опыления, необходимостью симбиоза с почвенными грибами и т.д. [2].
Наиболее полной сводкой по флоре орхидных Кабардино-Балкарской Республики (Кабардино-Балкарии, КБР) до сих пор остается работа С.Х. Шхагапсоева “Орхидные Кабардино-Балкарии” [3], в которой автор приводит 32 вида из родов Orchis L., Neottia Guett., Listera R.Br., Goodyera R.Вr., Epipactis Zinn, Cephalanthera Rich., Dactylorhiza Neck. ex Nevski, Platanthera Rich., Gymnadenia R.Вr., Anacamptis Rich. и др. При этом данные об эколого-биологических особенностях орхидных, в том числе структуре и функционировании природных популяций, для территории региона практически отсутствуют.
В новое издание Красной книги Кабардино-Балкарии внесены 22 вида семейства Orchidaceae [4], одним из которых является пальчатокоренник мясо-красный Dactylorhiza incarnata (L.) Soó – травянистый многолетник, представитель тубероидных орхидных с пальчатораздельными тубероидами [5]. Общий ареал вида охватывает Европу, Средиземноморье, Иран, Монголию, Китай [6]. В регионе встречается на сильно увлажненных и заболоченных лугах, по берегам рек на высоте 800–2300 м над ур. моря [7]. Местообитания D. incarnata в КБР нередко подвержены высокой антропогенной нагрузке в форме выпаса скота и вытаптывания при рекреации, сбора на букеты. Реже стеблекорневые тубероиды растений заготавливают в качестве лекарственного сырья. Соответственно, общими рекомендациями по охране данного вида на территории республики являются: организация особо охраняемых природных территорий (ООПТ), запрет сбора клубней, интродукция и мониторинг состояния популяций [4]. Отдельные аспекты мониторинга ценопопуляций D. incarnata рассматриваются в рамках настоящей работы.
Цель исследования – изучить изменчивость морфологических признаков особей, возрастную структуру и жизненность ценопопуляций D. incarnata при произрастании в различных по степени антропогенной нагрузки условиях среднегорий и высокогорий Центрального Кавказа (в границах Кабардино-Балкарской Республики).
МАТЕРИАЛ И МЕТОДЫ
В период 2015–2019 гг. на территории Кабардино-Балкарии изучены биометрические параметры растений, численность, плотность, возрастная структура и жизненность 14 ценопопуляций (ЦП) D. incarnata в естественных местообитаниях на высоте 1450–2550 м над ур. моря (табл. 1). Большая часть исследованных ЦП приурочены к луговым фитоценозам на пологих склонах крутизной 5°–20° с умеренным увлажнением (ЦП 1 – уроч. Джилы-Су, ЦП 2 – верховья р. Малка, ЦП 5 – окр. с. Тегенекли, ЦП 6 и 8 – восточный и южный склоны г. Чегет, ЦП 10, 14 – верховья Суканского ущелья, ЦП 12 – окр. с. Верхняя Балкария, ЦП 13 – Зольские пастбища). Ряд ЦП изучены в составе сильно увлажненных лугов, расположенных по берегам ручьев (ЦП 3 и 7 – окр. с. Терскол, ЦП 4 – окр. с. Байдаево, ЦП 9 – окр. оз. Гижгит, ЦП 11 – Тызыльское ущелье).
Таблица 1.
Характеристика фитоценозов с произрастанием Dactylorhiza incarnate Table 1. Characteristics of plant communities with Dactylorhiza incarnata
| ЦП СР |
Координаты: с.ш., в.д. Coordinates: N, E |
Фитоценозы, нарушение Plant communities, disturbance |
Высота над ур. моря, м Elevation a.s.l., m |
Проективное покрытие
травостоя, % Total plant projective cover, % |
Высота травостоя, см Herbage height, cm |
|---|---|---|---|---|---|
| 1 | 43.446930, 42.549786 | Вытаптываемый каменистый луг Trampled skeletal meadow |
2200 | 70 | 10 |
| 2 | 43.595298, 42.577541 | Ненарушенный субальпийский луг Undisturbed subalpine meadow |
2300 | 100 | 30 |
| 3 | 43.261428, 42.515807 | » | 2100 | 95 | 30 |
| 4 | 43.246904, 42.578983 | » | 2100 | 95 | 30 |
| 5 | 43.247048, 42.630219 | » | 1850 | 90 | 20 |
| 6 | 43.241396, 42.518332 | Вытаптываемый субальпийский луг Trampled subalpine meadow |
2200 | 80 | 10 |
| 7 | 43.259985, 42.514750 | » | 2100 | 70 | 5 |
| 8 | 43.233677, 42.501335 | Ненарушенный субальпийский луг Undisturbed subalpine meadow |
2550 | 95 | 20 |
| 9 | 43.457300, 42.994127 | » | 1450 | 95 | 25 |
| 10 | 43.10048, 43.55476 | Выпасаемый субальпийский луг Grazing subalpine meadow |
1900 | 90 | 7 |
| 11 | 43.118032, 43.486726 | Ненарушенный субальпийский луг Undisturbed subalpine meadow |
1700 | 100 | 30 |
| 12 | 43.118032, 43.486726 | » | 1700 | 95 | 30 |
| 13 | 43.75319, 42.74607 | Выпасаемый субальпийский луг Grazing subalpine meadow |
1800 | 90 | 15 |
| 14 | 43.09537, 43.52536 | Ненарушенный субальпийский луг Undisturbed subalpine meadow |
1950 | 95 | 30 |
Фитоценозы в пределах мест произрастания D. incarnata различаются по степени антропогенной нагрузки: вытаптываемые туристами участки луговой растительности по обочинам троп и дорог (ЦП 1, 6, 7), интенсивно используемые пастбища (ЦП 10, 13), ненарушенные субальпийские луга (ЦП 2–5, 8, 9, 11, 12, 14). Так, например, в 2017 г. на момент проведения исследований в уроч. Джилы-Су (ЦП 1) на площади 200 га в летний период ежедневно находилось до 2000 человек, что определяет высокую рекреационную нагрузку на растительный покров (вытаптывание почвы туристами вдоль троп и при установке палаточных лагерей, сбор растений на букеты и т.п.). Склоны г. Чегет (ЦП 6) и Терскольского ущелья (ЦП 7), расположенных на территории национального парка “Приэльбрусье”, также традиционно являются местами массового посещения любителями активного летнего отдыха. Всего территорию национального парка, со слов егерей и сотрудников, ежегодно в летний период посещают десятки тысяч человек. При этом интенсивному антропогенному воздействию в основном подвергаются участки растительности, непосредственно примыкающие к горным тропам и дорогам. ЦП 10 в верховьях Суканского ущелья была исследована в июле 2018 г. на территории, через которую после весенне-летнего стравливания растительности фермеры дважды в день осуществляли прогон 1200 овец на более удаленные пастбищные угодья. Пастбищная нагрузка на данном участке составляла 50–60 голов/га мелкорогатого скота. Высокогорные Зольские пастбища (ЦП 13) также традиционно используются для осуществления отгонного животноводства. Поголовье ежегодно выпасаемого скота здесь составляет 130 тыс. голов (из них 81 тыс. овец и коз) при общей площади пастбищных угодий 129.6 тыс. га [8].
В качестве счетной единицы рассматривали морфологическую особь – побег генеративного или вегетативного происхождения. Возрастные состояния растений выделены на основании комплекса качественных и количественных морфологических признаков надземных органов (исследования проведены без выкапывания особей) согласно методике Т.А. Работнова [9] и А.А. Уранова [10]. Онтогенетический спектр ЦП, численность и плотность растений изучали методом учетных площадок (20 квадратов размером 1 м2 для каждой ЦП), которые в границах ЦП размещали таким образом, чтобы охватить участки как с высокой, так и с низкой численностью особей. Возрастную структуру ЦП анализировали по критерию “∆–ω” Л.А. Животовского [11] и с использованием индекса восстановления ${{I}_{B}}$ (1) [12].
(1)
${{I}_{{\text{в}}}}~\, = \,~\frac{{\sum\limits_{i - 3}^5 {{{n}_{i}}} }}{{\sum\limits_{i - 6}^8 {{{n}_{i}}} }}~,$Оценка жизненности (виталитета) ЦП проведена с вычислением показателя IVC путем взвешивания средних значений каждого из исследуемых морфологических параметров растений в каждой ЦП по среднему значению параметра для всей выборки (2) [13].
(2)
$IVC~\,\, = \,\,~\frac{{\sum\limits_{i - 1}^N {\frac{{{{X}_{i}}}}{{{{{\bar {X}}}_{i}}}}} }}{N}~~,$При определении изменчивости признаков растений и виталитета ЦП анализировали 11 морфологических параметров 30 зрелых генеративных особей в каждой ЦП. В их числе: высота побега и диаметр его основания, см; длина и ширина нижнего и верхнего листьев, см; диаметр цветоноса, см; высота и диаметр соцветия, см; число цветков в соцветии и число листьев на побеге, шт. В качестве показателей изменчивости и пластичности признаков особей использовали фитоценотическую пластичность (Ip) (3) и индивидуальную (внутрипопуляционную) изменчивость CVср, % (4), а также внутривидовую (межпопуляционную) изменчивость $C{{V}_{{\bar {х}\,{\text{ср}}}}},$ %, вычисляемую по среднему значению признака в ряду ценопопуляций [14, 15]. Уровни варьирования параметров приняты по Г.Н. Зайцеву [16]: CV > > 20% – высокий; CV < 10% – низкий; CV = 11–20% – средний.
где Ip – показатель фитоценотической пластичности, А и В – максимальное и минимальное среднее значение признака соответственно. где CVср – индивидуальная (внутрипопуляционная) изменчивость признака, CV – изменчивость признака в пределах конкретной ЦП, n – число ЦП.Первичный материал обработан с использованием пакетов программ Statistica 10 (однофакторный дисперсионный анализ ANOVA), EXCEL.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
D. incarnata – морфологически изменчивый вид, для которого характерна широкая вариабельность окраски цветков и прицветников, формы и пигментации листьев [1, 17]. В условиях Центрального Кавказа вид обладает высокой фитоценотической пластичностью признаков (Ip = = 41–87%, в среднем 68%), отражающей изменение средних значений морфологических параметров в разных условиях произрастания и являющейся важным показателем адаптивности растений (табл. 2). Наиболее пластичными признаками являются высота побега и длина нижнего листа со значениями Ip более 80%, наименее пластичны диаметр цветоноса и число листьев на побеге.
Таблица 2.
Фитоценотическая пластичность морфологических признаков Dactylorhiza incarnate Table 2. Phytocenotic plasticity of Dactylorhiza incarnata morphological characters
| ЦП СР |
Средние значения морфологических признаков, см Average values of morphological characters, cm |
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | |
| 1 | 18.6 ± 2.2 | 0.45 ± 0.03 | 8 ± 0.9 | 1.8 ± 0.4 | 5.9 ± 0.3 | 1.3 ± 0.1 | 4.9 ± 0.7 | 2.7 ± 0.1 | 0.33 ± 0.02 | 4.1 ± 0.6 | 13.2 ± 8.4 |
| 2 | 34 ± 3.9 | 0.50 ± 0.04 | 16.2 ± 1.8 | 2.3 ± 0.4 | 6 ± 0.4 | 1.8 ± 0.4 | 5.7 ± 0.6 | 3.4 ± 0.2 | 0.34 ± 0.03 | 5.5 ± 0.5 | 14 ± 5.4 |
| 3 | 63.4 ± 5.8 | 0.98 ± 0.05 | 33.8 ± 3.3 | 4.3 ± 0.5 | 11.9 ± 0.6 | 3.4 ± 0.5 | 11.8 ± 0.9 | 6.1 ± 0.3 | 0.54 ± 0.03 | 6.2 ± 0.3 | 27.1 ± 9.3 |
| 4 | 56.3 ± 4.2 | 0.81 ± 0.04 | 23.6 ± 3.1 | 3.4 ± 0.4 | 9.8 ± 0.4 | 2.7 ± 0.5 | 9.5 ± 0.7 | 4.5 ± 0.2 | 0.50 ± 0.08 | 5.9 ± 0.8 | 23.4 ± 5.7 |
| 5 | 36.3 ± 3.9 | 0.51 ± 0.06 | 17.7 ± 2.4 | 2.4 ± 0.3 | 6.2 ± 0.3 | 1.8 ± 0.2 | 6.5 ± 0.5 | 3.8 ± 0.1 | 0.37 ± 0.03 | 5.7 ± 0.7 | 17.1 ± 6.8 |
| 6 | 16.2 ± 2.1 | 0.38 ± 0.02 | 5.9 ± 0.5 | 1.6 ± 0.2 | 3.8 ± 0.2 | 1.1 ± 0.1 | 3.7 ± 0.5 | 2.2 ± 0.1 | 0.29 ± 0.02 | 4.2 ± 0.3 | 11.4 ± 5.1 |
| 7 | 14.4 ± 2.4 | 0.35 ± 0.02 | 4.7 ± 0.7 | 1.6 ± 0.2 | 3.6 ± 0.6 | 1 ± 0.1 | 3.5 ± 0.5 | 2 ± 0.2 | 0.27 ± 0.03 | 4.2 ± 0.4 | 9.7 ± 3.8 |
| 8 | 35.2 ± 3.3 | 0.49 ± 0.04 | 15.5 ± 1.9 | 2.3 ± 0.4 | 5.9 ± 0.4 | 1.8 ± 0.1 | 6.3 ± 0.6 | 3.6 ± 0.1 | 0.35 ± 0.05 | 5.7 ± 0.6 | 15 ± 4.6 |
| 9 | 51.5 ± 5.9 | 0.69 ± 0.06 | 19.3 ± 2.4 | 3.1 ± 0.5 | 7.8 ± 0.7 | 2.3 ± 0.2 | 8.6 ± 0.8 | 4.2 ± 0.3 | 0.44 ± 0.04 | 6.1 ± 0.6 | 20.3 ± 6.8 |
| 10 | 12.1 ± 2.6 | 0.32 ± 0.02 | 4.5 ± 0.5 | 1.4 ± 0.1 | 3.5 ± 0.4 | 0.9 ± 0.1 | 3.3 ± 0.4 | 2 ± 0.3 | 0.27 ± 0.02 | 4.1 ± 0.3 | 8.6 ± 3.7 |
| 11 | 63 ± 6.7 | 0.97 ± 0.07 | 33.2 ± 3.7 | 4.1 ± 0.1 | 11.2 ± 0.9 | 3.3 ± 0.5 | 11.1 ± 1.2 | 5.5 ± 0.8 | 0.49 ± 0.04 | 5.8 ± 0.5 | 22.5 ± 6.7 |
| 12 | 33.2 ± 3.5 | 0.48 ± 0.05 | 13.5 ± 1.2 | 2.2 ± 0.2 | 5.6 ± 0.4 | 1.6 ± 0.3 | 6.2 ± 0.7 | 3.5 ± 0.6 | 0.33 ± 0.08 | 5.5 ± 0.6 | 13.1 ± 5.2 |
| 13 | 15 ± 1.4 | 0.43 ± 0.04 | 5.6 ± 0.4 | 1.6 ± 0.3 | 3.8 ± 0.4 | 1 ± 0.1 | 4.2 ± 0.4 | 2.7 ± 0.2 | 0.30 ± 0.03 | 4.3 ± 0.7 | 11.5 ± 3.4 |
| 14 | 33.1 ± 4.2 | 0.47 ± 0.04 | 13 ± 1.1 | 2.1 ± 0.2 | 5.6 ± 0.6 | 1.6 ± 0.1 | 6 ± 0.5 | 3.3 ± 0.2 | 0.33 ± 0.04 | 5.6 ± 0.6 | 13.2 ± 4.8 |
| Ip | 0.81 | 0.67 | 0.87 | 0.68 | 0.7 | 0.72 | 0.72 | 0.68 | 0.50 | 0.41 | 0.68 |
Примечание. ЦП – ценопопуляции (1–14); I–XI – порядковый номер признака: высота побега и диаметр его основания (I и II, см), длина и ширина нижнего (III и IV, см) и верхнего (V и VI, см) листьев, высота и диаметр соцветия (VII и VIII, см), диаметр цветоноса (IX, см), число листьев (X, шт.), число цветков в соцветии (XI, шт.); Ip – показатель фитоценотической пластичности признака. Note. СР – coenopopulations (1–14); I–XI – character’s ordinal number: shoot height and base diameter (I and II, cm), length and width of the lower (III and IV, cm) and the upper (V and VI, cm) leaves, inflorescence height and diameter (VII and VIII, cm), peduncle diameter (IX, cm), number of leaves (X), number of flowers in the inflorescence (XI); Ip – phytocoenotic plasticity index.
Индивидуальная (внутрипопуляционная) изменчивость большинства биометрических признаков D. incarnata, характеризующая морфологическую гетерогенность ЦП, имеет средний уровень варьирования (CVср = 11–20%) (табл. 3). Межпопуляционная изменчивость вида, характеризующая габитуальные различия растений разных ЦП, отличается высокими показателями ($C{{V}_{{\bar {х}\,{\text{ср}}}}}$ = 21.4–65.1%). При этом наименее вариабельными являются диаметр цветоноса и число листьев на побеге.
Таблица 3.
Коэффициенты изменчивости морфологических признаков Dactylorhiza incarnate Table 3. Variation coefficients of Dactylorhiza incarnata morphological characters
| ЦП СР |
Коэффициенты изменчивости морфологических признаков CV, % Variation coefficients of morphological characters CV, % |
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | |
| 1 | 9.42 | 13.9 | 21.8 | 15.4 | 22.2 | 17.5 | 12.9 | 11.7 | 14.6 | 17.4 | 24.5 |
| 2 | 10.1 | 6.4 | 12.4 | 9.7 | 10 | 9.6 | 9.7 | 6.1 | 6.3 | 9.2 | 22.7 |
| 3 | 12.2 | 14.1 | 17.9 | 12.3 | 14.3 | 21.9 | 14.1 | 14.8 | 19.9 | 12.9 | 20.4 |
| 4 | 13.4 | 16.2 | 24.4 | 16.6 | 17.5 | 28.7 | 16.6 | 13.6 | 15.3 | 11.4 | 30.7 |
| 5 | 11.7 | 11.7 | 16.3 | 14 | 13.5 | 11.6 | 18.3 | 11.9 | 11.2 | 14.4 | 25.8 |
| 6 | 19.5 | 16.5 | 28 | 19.7 | 17 | 14.1 | 15 | 24.6 | 9.7 | 17.9 | 27.2 |
| 7 | 22.9 | 18.2 | 36.4 | 24.5 | 16.2 | 16.1 | 16.4 | 27 | 10.4 | 21.6 | 20.7 |
| 8 | 11.7 | 11.7 | 16.5 | 14.4 | 14 | 11.6 | 18.2 | 14.3 | 11.1 | 13.9 | 24 |
| 9 | 16 | 18.2 | 25.6 | 18.5 | 21.6 | 25 | 18.3 | 14.5 | 18.1 | 13.2 | 32.3 |
| 10 | 29.1 | 18.3 | 38.6 | 29.3 | 17.7 | 17.8 | 24.9 | 28.4 | 12.6 | 24 | 21.2 |
| 11 | 11.3 | 12.3 | 16.4 | 12.9 | 16.7 | 22.2 | 12.9 | 18.6 | 20.2 | 12.9 | 24 |
| 12 | 12.2 | 9.9 | 17.7 | 15.7 | 14.9 | 11.4 | 17.1 | 13.3 | 10.1 | 13.1 | 21.4 |
| 13 | 20.4 | 13.5 | 31.8 | 19.5 | 26.7 | 27.1 | 17 | 11.8 | 9.4 | 19.1 | 14.5 |
| 14 | 10.3 | 9.4 | 14.4 | 15.7 | 14.3 | 10.6 | 16.9 | 14 | 8.4 | 15.2 | 14.4 |
| CVср, % | 15 | 13.6 | 22.7 | 17 | 16.9 | 17.5 | 16.3 | 16 | 12.7 | 15.5 | 23.1 |
| $C{{V}_{{\bar {х}\,{\text{ср}}}}}$, % | 53 | 40.6 | 65.1 | 40.5 | 44.7 | 48.3 | 44.2 | 37.6 | 27.6 | 21.4 | 43 |
Примечание. ЦП – ценопопуляции (1–14); I–XI – порядковый номер признака (см. табл. 2); CVср, % – внутрипопуляционная (индивидуальная) изменчивость признака; $C{{V}_{{\bar {х}\,{\text{ср}}}}}$, % – межпопуляционная (внутривидовая) изменчивость признака. Note. СР – cenopopulations (1–14); I–XI – ordinal number of characters (tabl. 2); CVср, % – individual variation of characters; $C{{V}_{{\bar {х}\,{\text{ср}}}}}$, % – interpopulation variation of the characters.
Параметры внутрипопуляционного варьирования всех признаков ниже показателей межпопуляционной изменчивости (CVср < $C{{V}_{{\bar {х}\,{\text{ср}}}}}$), что свидетельствует об их высокой вариабельности в разных условиях произрастания. Учитывая это, морфологические параметры D. incarnate целесообразно рассматривать в качестве индикаторов при оценке соответствия условий среды эколого-биологическим требованиям вида, отражением которого является максимально возможное развитие габитуса растений.
На эколого-ценотическом градиенте, характеризующем степень благоприятствования условий среды росту и развитию растений, формируется следующий ряд ЦП: ЦП3 (IVC = 1.23)–ЦП 11 (1.22)–ЦП 4 (1.20)–ЦП 9 (1.18)–ЦП 5 (1.12)–ЦП 8 (1.10)–ЦП 2 (1.07)–ЦП 12 (1.06)–ЦП 14(1.05)–ЦП 11(0.88)–ЦП 6(0.86)–ЦП 13(0.84)–ЦП 7(0.83)–ЦП 10 (0.82). Наиболее приближенные к оптимальным для роста и развития D. incarnata условия (IVC = 1.18–1.23) складываются в ЦП 3, 4, 9, 11, расположенных в ненарушенных луговых фитоценозах по берегам ручьев, что, вероятно, обусловлено благоприятным водным режимом и отсутствием выраженной антропогенной нагрузки на растительный покров. Достаточно благоприятные для реализации ростовых потенций растений условия (IVC = 1.05–1.12) отмечены также при произрастании на склонах гор в составе субальпийских и остепненных субальпийских лугов с низким уровнем антропогенной нагрузки (ЦП 2, 5, 8, 12, 14). Перевыпас скота, вытаптывание при рекреации, сопровождающиеся переуплотнением почвы и механическими повреждениями растений, приводят к угнетению их роста и развития в ЦП 1, 6, 7, 10, 13 (IVC = 0.82–0.88).
Данный вывод подтвердили результаты однофакторного дисперсионного анализа, показавшие, что средние значения всех анализируемых морфологических параметров растений достоверно отличаются в ЦП трех независимых групп (ЦП 3, 4, 9, 11; ЦП 2, 5, 8, 12, 14; ЦП 1, 6, 7, 10, 13) (табл. 4).
Таблица 4.
Результаты однофакторного дисперсионного анализа морфологических признаков Dactylorhiza incarnata трех независимых групп ценопопуляций Table 4. Results of one-way variation analysis of Dactylorhiza incarnata morphological characters for three independent coenopopulations
| Параметр Morphological characters |
F | p |
|---|---|---|
| Высота побега, см/Height of the shoot, cm | 179.39 | 0.000 |
| Диаметр основания побега, см/Diameter of shoot base, cm | 42.22 | 0.000 |
| Длина нижнего листа, см/Length of the lower leaves, cm | 32.40 | 0.000 |
| Ширина нижнего листа, см/Width of the lower leaves, cm | 48.13 | 0.000 |
| Длина верхнего листа, см/Length of the upper leaves, cm | 32.46 | 0.000 |
| Ширина верхнего листа, см/Width of the upper leaves, cm | 40.83 | 0.000 |
| Высота соцветия, см/Height of the inflorescence, cm | 58.92 | 0.000 |
| Диаметр соцветия, см/Diameter of the inflorescence, cm | 30.72 | 0.000 |
| Диаметр цветоноса, см/Diameter of the peduncle, cm | 59.44 | 0.000 |
| Число листьев, шт. /Number of leaves, pcs | 284.72 | 0.000 |
| Число цветков в соцветии, шт. /Number of flowers, pcs | 40.62 | 0.000 |
Согласно результатам попарного апостериорного сравнения средних значений морфологических признаков между тремя группами ценопопуляций по критерию Фишера (Fisher LSD), все три группы достоверно отличаются друг от друга (р < 0.05) (табл. 5).
Таблица 5.
Результаты расчета критерия Фишера (Fisher LSD) при попарном апостериорном сравнении трех групп ценопопуляций Table 5. Results of Fisher LSD pairwise post hoc comparison of the three groups of coenopopulations
| Группы ценопопуляций Groups of cenopopulations |
1 M = 58.6 |
2 M = 34.4 |
3 M = 15.3 |
|---|---|---|---|
| 1 | – | 0.000000 | 0.000000 |
| 2 | 0.000000 | – | 0.000002 |
| 3 | 0.000000 | 0.000002 | – |
Примечание. М – значения среднего статистического по группам; в таблице представлены полученные при попарных сравнениях уровни статистической значимости; выделенные полужирным значения достоверны при уровне значимости p < 0.05. Note. M – average values for groups; the table shows the levels of statistical significance obtained in pairwise comparisons; values reliable at significance level p < 0.05 are given in bold.
В большом жизненном цикле D. incarnata изучены шесть возрастных состояний: ювенильные (j), имматурные (im), виргинильные (v), молодые, зрелые и старые генеративные (g1, g2, g3) растения. Морфологию и численность проростков с преимущественно подземным образом жизни не изучали, особи постгенеративного онтогенетического периода в исследованных ЦП не выявлены.
Ювенильные растения характеризуются общей простотой строения. Развивают два прикорневых ассимилирующих разноразмерных листа, длина и ширина меньшего из которых составляет 0.3–3.2 и 0.1–0.4 см соответственно, более крупного – 1.5–5.6 и 0.2–0.5 см. Имматурные особи формируют один–два килеватых узколанцетных листа длиной 4.1–11.4 см, шириной 0.5–0.6 см. Виргинильные растения характеризуются значительным накоплением вегетативной массы, образуют два–три крупных жестких килеватых листа, отличающихся значительной длиной – 7.6–19.3 см при ширине 1.3–4.6 см. На данной стадии изредка образуются компактные моноцентрические клоны из двух одновозрастных растений или из материнской виргинильной и дочерней имматурной особей. Молодые генеративные растения – относительно низкорослые немногоцветковые (обычно до 15–20 шт. в соцветии), с 3–6 некрупными широколанцетными листьями и бороздчатым цветоносом. Наиболее мощные зрелые генеративные растения отличаются максимальным развитием вегетативных и генеративных органов. Они имеют 3–8 узколанцетных листьев, нижние из которых в отдельных случаях достигают длины 40 см и ширины 5–5.5 см и до 30–40 шт. цветков в длинном, до 14–15 см, соцветии. Довольно редкие случаи вегетативного размножения на данной стадии выражаются в образовании дочернего растения того же возрастного состояния или омоложенной виргинильной особи. Параметры надземных вегетативных и генеративных органов старых генеративных растений характеризуются несколько меньшими значениями. Это относительно низкорослые, коренастые, немногоцветковые особи, отличительным признаком которых нередко является наличие остатков старых цветоносов.
Таким образом, для D. incarnata на Центральном Кавказе характерен смешанный способ размножения, однако низкая интенсивность вегетативного размножения, свойственная и для других видов тубероидных орхидных [2], обусловливает его незначительный вклад в возобновление ЦП.
Базовый возрастной спектр D. incarnata, позволяющий выделить общие закономерности, повторяющиеся в возрастной структуре отдельных ЦП, и отражающий особенности биологии вида, нормальный со слабым пиком на генеративной группе (39.1%), пониженной долей ювенильных растений (10.4%) и практически равным участием имматурных и виргинильных особей (24.9 и 25.6%). Соответственно, несмотря на различия в условиях произрастания, для D. incarnata в целом характерно эффективное возобновление ЦП.
В то же время, особенности возрастной структуры отдельных ЦП вида в значительной степени определяются степенью и характером антропогенной нагрузки, уровнем межвидовой конкуренции в фитоценозе, косвенным показателем которого является общее проективное покрытие травостоя. Так, при произрастании вида на ненарушенных лугах с высоким общим проективным покрытием (90–100%) в возрастных спектрах ЦП 2–5, 8, 9, 11, 12, 14 отмечено максимальное увеличение доли растений генеративного периода (42.3–57.5%) (рис. 1). Данные ЦП по критерию “Δ–ω” относятся к типу зреющих и зрелых (табл. 6). Это обусловлено, вероятно, угнетением молодых особей D. incarnata более конкурентоспособными растениями сопутствующих видов с сокращением процента их представленности в возрастных спектрах. Так, для ЦП 2–5, 8, 9, 11, 12, 14 отмечены наименьшие значения индекса восстановления Iв (3.7–6.2), характеризующего эффективность возобновления. Плотность особей в ЦП не превышает 10.4–20.1 особ./м2.
Рис. 1.
Возрастные спектры ценопопуляций Dactylorhiza incarnata. j – ювенильные, im – имматурные, v – виргинильные, g – генеративные растения. По горизонтали – номера ценопопуляций (1–14); по вертикали – доля возрастной группы в спектре, %. Fig. 1. Ontogenetic spectra of Dactylorhiza incarnata cenopopulations. j – juvenile, im – immature, v – virginal, g – generative individuals. X-axis – cenopopulations numbers (1–14); y-axis – % of individuals in the spectrum.
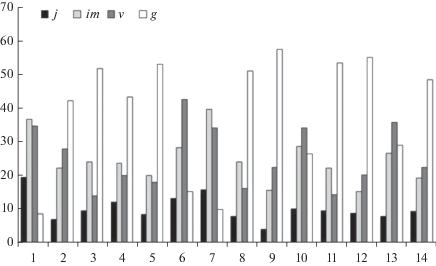
Таблица 6.
Плотность особей и демографические показатели ценопопуляций Dactylorhiza incarnate Table 6. Demographic indicators of Dactylorhiza incarnata cenopopulations
| ЦП CP |
S, м2 S, m2 |
N, шт. N, ind. |
M, особ./м2 M, ind./m2 |
Iв | Доля генеративных особей, % Percentage of generative individuals, % |
Δ | ω | Тип ЦП Type of CP |
||
|---|---|---|---|---|---|---|---|---|---|---|
| g1 | g2 | g3 | ||||||||
| 1 | 460 | 26 772 | 58.20 | 34.16 | 46.4 | 42.7 | 10.9 | 0.18 | 0.44 | Молодая/Young |
| 2 | 900 | 9378 | 10.42 | 6.21 | 32.4 | 59.5 | 8.1 | 0.32 | 0.62 | Зреющая/Maturing |
| 3 | 300 | 5259 | 17.53 | 4.19 | 32 | 53.2 | 14.8 | 0.41 | 0.74 | Зрелая/Mature |
| 4 | 150 | 3018 | 20.12 | 6.12 | 34.8 | 55.3 | 9.9 | 0.34 | 0.64 | Зреющая/Maturing |
| 5 | 400 | 6268 | 15.67 | 4.13 | 25.5 | 62.3 | 12.2 | 0.44 | 0.77 | Зрелая/Mature |
| 6 | 640 | 28 896 | 45.15 | 24.73 | 42.4 | 48.9 | 8.7 | 0.24 | 0.48 | Молодая/Young |
| 7 | 160 | 9030 | 56.44 | 32.81 | 52.1 | 40.7 | 7.2 | 0.21 | 0.45 | Молодая/Young |
| 8 | 640 | 10 380 | 16.22 | 4.22 | 33.1 | 50.7 | 16.2 | 0.40 | 0.74 | Зрелая/Mature |
| 9 | 900 | 11 232 | 12.48 | 3.72 | 29.3 | 57.2 | 13.5 | 0.52 | 0.81 | Зрелая/Mature |
| 10 | 600 | 9618 | 16.03 | 12.05 | 50.1 | 44.1 | 5.8 | 0.27 | 0.50 | Молодая/Young |
| 11 | 460 | 7875 | 17.12 | 4.08 | 35.2 | 46.4 | 18.4 | 0.45 | 0.77 | Зрелая/Mature |
| 12 | 800 | 12 520 | 15.65 | 3.84 | 36.7 | 50.8 | 12.5 | 0.49 | 0.79 | Зрелая/Mature |
| 13 | 900 | 10 935 | 12.15 | 10.72 | 40.2 | 54.2 | 5.6 | 0.29 | 0.52 | Молодая/Young |
| 14 | 1200 | 17 064 | 14.22 | 4.47 | 40.1 | 48.5 | 11.4 | 0.37 | 0.72 | Зрелая/Mature |
Примечание. S, N, M – площадь участка, общая численность особей, плотность особей в составе ценопопуляций; Iв – индекс восстановления; g1 – молодые генеративные, g2 – зрелые генеративные, g3 – старые генеративные особи; Δ и ω – индексы возрастности и эффективности. Note. S, N, M – plot area, total number of individuals, density of individuals in cenopopulations; Iв – renewal index; g1 – young generative, g2 – mature generative, g3 – old generative individuals; Δ and ω – indices of age and efficiency.
При интенсивном выпасе скота (ЦП 10, 13) доля генеративных растений снижается до 26.6–29.1% при выраженном накоплении в возрастных спектрах виргинильных особей (34.3–35.9%). Это может быть обусловлено низкими темпами развития молодых растений на ресурсо- и энергозатратном этапе формирования генеративных органов при произрастании в неблагоприятных условиях. Соответственно, несмотря на возрастание в ЦП 10, 13 индекса восстановления Iв, плотность особей остается относительно низкой.
Максимальная плотность растений отмечена в ЦП 1, 6, 7 (45.1–58.2 особ./м2) в луговых фитоценозах на щебнистых склонах, подверженных высокой рекреационной нагрузке и перевыпасу скота. При сниженном общем проективном покрытии травостоя (70–80%) эффективное, несмотря на антропогенную нагрузку, возобновление вида и высокая приживаемость молодых растений обусловливают максимальное возрастание в онтогенетических спектрах доли ювенильных (13.4–19.6%) и имматурных (28.5–39.7%) растений, что в свою очередь приводит к повышению индекса восстановления (24.73–34.16); ценопопуляции относятся к типу молодых. Доля особей генеративного периода в спектрах, напротив, не превышает 8.8–15.4%, что может быть связано с низкими темпами развития виргинильных растений (34.2–42.7%).
ЗАКЛЮЧЕНИЕ
Согласно результатам изучения Dactylorhiza incarnata на территории Кабардино-Балкарской Республики для этого вида в целом характерна высокая пластичность и изменчивость морфологических признаков при изменении условий произрастания. При этом наиболее благоприятные для реализации потенций роста и развития вида условия складываются в ненарушенных высоко увлажненных луговых фитоценозах. Усиление антропогенного воздействия (рекреация, выпас скота) приводит к снижению жизненности ценопопуляций.
Онтогенетически обусловленным процессом, влияющим на возрастной спектр и плотность особей в ценопопуляциях D. incarnata, является низкая интенсивность вегетативного размножения, а в роли основных внешних факторов выступают степень антропогенной нагрузки, наличие фитоценотических конкурентов, характер субстрата (плотность, состав, влажность). При стрессовом характере воздействия внешних факторов для вида отмечены задержка в развитии особей на виргинильной стадии, снижение доли генеративных растений в возрастных спектрах, но эффективное возобновление ценопопуляций.
Таким образом, при произрастании в условиях Центрального Кавказа D. incarnata проявляет признаки SR-стратегии жизни: интенсификация процессов роста, стабильное возобновление и успешное развитие при высоком уровне межвидовой конкуренции с сохранением подчиненного положения в фитоценозе (фитоценотическая патиентность); подавление ростовых процессов и обеспечение репродукции при высоком антропогенном воздействии (экологическая патиентность); интенсивное возобновление ценопопуляций, способность захватывать незанятые территории в условиях пониженного уровня межвидовой конкуренции (эксплерентность).
Несмотря на реализацию в условиях антропогенной нагрузки эксплерентной компоненты стратегии жизни вида, приводящей к увеличению численности и плотности растений, наиболее оптимальными для произрастания D. incarnata следует считать ненарушенные увлажненные луговые фитоценозы, для которых отмечены максимальные показатели развития габитуса растений, высокая доля генеративных растений в возрастных спектрах и стабильное возобновление ценопопуляций. Соответственно, эффективной мерой по охране данного вида в природе является ограничение антропогенной нагрузки, в частности, создание ООПТ в границах произрастания вида, а также существенное усиление контроля над соблюдением природоохранного режима в границах национального парка “Приэльбрусье”.
Список литературы
Стецук Н.П. 2006. Основные механизмы устойчивости ценопопуляций некоторых видов орхидных Южного Приуралья. – Вестник Оренбургского государственного университета. 4 (Приложение “Биоразнообразие и биоресурсы”): 93–96. http://vestnik.osu.ru/2006_4_1/28.pdf
Перебора Е.А. 2008. Особенности развития некоторых тубероидных орхидных в условиях Северо-Западного Кавказа. – Экологический вестник Северного Кавказа. 4(2): 106–124. http://ecokavkaz.ru/media/docs/2008/2/2008-2-9.pdf
Шхагапсоев С.Х. 1998. Орхидные Кабардино-Балкарии. – В сб.: Охрана и культивирование орхидей. Краснодар. С. 12–14.
Красная книга Кабардино-Балкарской Республики. 2018. Нальчик. 496 с.
Перебора Е.А. 2003. Типы побегов и жизненные формы орхидных Северо-Западного Кавказа. – Бюллетень Ботанического сада им. И.С. Косенко. 21: 118–180.
Конспект флоры Кавказа. 2006. Т. 2. СПб. 467 с. https://www.binran.ru/resursy/informatsionnyye-resursy/tekuschie-proekty/caucasian-flora/
Шхагапсоев С.Х. 2015. Растительный покров Кабардино-Балкарии. Нальчик. 350 с.
Гукежев В.М., Бербекова Н.В., Габаев М.С., Батырова О.А. 2017. Горные пастбища КБР – реальный источник производства органической мясной продукции. – Вестник экспертного совета. 1(8): 37–41. https://ski.ranepa.ru/about/files/journal_vestnik/VESTNIK_No18_2017.pdf
Работнов Т.А. 1950. Жизненный цикл многолетних травянистых растений в луговых ценозах. – Труды БИН АН СССР. Серия Геоботаника. 6: 7–204.
Уранов А.А. 1975. Возрастной спектр фитоценопопуляций как функция времени и энергетических волновых процессов. – Научные доклады высшей школы. Биологические науки. 2: 7–34.
Животовский Л.А. 2001. Онтогенетические состояния, эффективная плотность и классификация популяций. – Экология. 1: 3–7.
Жукова Л.А. 1995. Популяционная жизнь луговых растений. Йошкар-Ола. 224 с.
Ишбирдин А.Р., Ишмуратова М.М. 2004. Адаптивный морфогенез и эколого-ценотические стратегии выживания травянистых растений. – В сб.: Методы популяционной биологии: Мат. VII Всероссийского популяционного семинара. Сыктывкар. С. 113–120.
Злобин Ю.А. 1989. Теория и практика оценки виталитетного состава ценопопуляций растений. – Ботанический журнал. 74(6): 769–781.
Ишбирдин А.Р., Ишмуратова М.М., Жирнова Т.В. 2005. Стратегии жизни ценопопуляции Cephalanthera rubra (L.) Rich. на территории Башкирского государственного заповедника. – В сб.: Популяции в пространстве и времени: Мат. VIII Всероссийского популяционного семинара. Нижний Новгород. С. 85–98.
Зайцев Г.Н. 1990. Математика в экспериментальной биологии. М. 296.
Мамаев С.А., Князев М.С., Куликов П.В., Филиппов Е.Г. 2004. Орхидные Урала: систематика, биология, охрана. Екатеринбург. 124 с.
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы