

Растительные ресурсы, 2021, T. 57, № 1, стр. 68-82
Характеристики ценопопуляций и запасы сырья Trifolium pratense (Fabaceae) в разных экологических условиях на территории Республики Татарстан
Н. Б. Прохоренко 1, *, Л. Р. Кадырова 1, Г. В. Демина 1, О. А. Тимофеева 1
1 Казанский (Приволжский) федеральный университет
г. Казань, Россия
* E-mail: nbprokhorenko@mail.ru
Поступила в редакцию 19.02.2020
После доработки 21.08.2020
Принята к публикации 10.12.2020
Аннотация
Исследованы 8 ценопопуляций Trifolium pratense L. из разных местообитаний на территории Республики Татарстан. В работе приводятся результаты анализа связи морфоструктурных параметров растений, потенциальной и реальной семенной продуктивности и запасов фитомассы с величиной экологических факторов. Установлено, что в пределах Республики Татарстан на свежих со средним содержанием минерального азота почвах (юго-запад подтаежной подзоны и юг широколиственной подзоны) ценопопуляции T. pratense имеют сравнительно низкую плотность и преимущественно состоят из генеративных растений высокого морфологического статуса. В условиях сухих периодически свежих с низким содержанием азота почв (отмеченных в центре широколиственной подзоны и на западе лесостепной подзоны) формируются ценопопуляции T. pratense высокой плотности, состоящие из генеративных и молодых вегетативных растений, причем генеративные растения имеют средний морфологический статус. В ценопопуляциях T. pratense высокой плотности выявлена наибольшая урожайность воздушно-сухой надземной фитомассы (около 413–432 кг/га), что позволяет рекомендовать их для заготовки растительного сырья.
Клевер луговой Trifolium pratеnse L. – многолетний летнезеленый или летне-зимнезеленый травянистый стержнекорневой с многоглавым каудексом симподиально нарастающий поликарпик с полурозеточным прямостоячим или приподнимающимся побегом [1, 2]. T. pratense имеет Евро-Западно-Азиатский ареал, в пределах которого широко распространен в составе луговых и опушечных сообществ, а также по сорным местам и у дорог [1, 3]. Данный вид – ценное кормовое и пищевое, медоносное, красильное и лекарственное растение, используемое в народной медицине. В его надземных органах накапливаются тритерпеноиды, каротиноиды, кумарины, антоцианы, высшие жирные кислоты (пальмитиновая, олеиновая, стеариновая и др.) и их производные, бензол, фенолы, фенолкарбоновые кислоты и их производные, флавоноиды и изофлавоноиды (всего 96 соединений), а также некоторые другие вещества [4–6]. Экспериментально показано, что флавоноиды, накапливающиеся в траве T. pratense, оказывают антиаритмическое и антигипертензивное действие [5], изофлавоны замедляют процесс старения кожи, т.к. стимулируют активность остеобластов, оказывают положительное влияние на уровень холестерина [4]. Кроме того, экстракт T. pratense при клинических испытаниях обнаруживает эстрогенную и антиоксидантную активность [7, 8], что доказывает его эффективность при лечении симптомов менопаузы. Как показывают исследования, благодаря фитохимическому составу водные и этилацетатные экстракты листьев T. pratense эффективно защищают ткани от окислительного стресса, поэтому данный вид можно рассматривать как важный источник биоактивных природных соединений и использовать в качестве пищевой добавки или лечебного средства [9].
В разных местообитаниях виды растений имеют специфическую структуру популяций, семенную продуктивность и интенсивность роста, что отражает определенную степень пластичности вида [10]. При характеристике ресурсов дикорастущих лекарственных растений какого-либо региона важным аспектом исследований выступает анализ структуры их ценопопуляций и урожайности в различных эколого-ценотических условиях [11–13]. Это позволяет выявить растительные сообщества, в составе которых ценопопуляции лекарственных растений можно рекомендовать для сбора лекарственного сырья, а также получения семенной продукции, необходимой для возделывания или реинтродукции.
Республика Татарстан (РТ) охватывает четыре ландшафтные подзоны: южно-таежную и подтаежную подзоны в составе бореальной ландшафтной зоны, широколиственную и лесостепную подзону в составе суббореальной северной семигумидной ландшафтной зоны [14]. При этом в пределах РТ площадь лесостепной подзоны наибольшая и составляет около 35 000 км2, подтаежной и широколиственной – около 14 000 км2 каждая, южнотаежной – 5000 км2. В связи с разнообразием ландшафтных подзон на территории РТ актуальны исследования структурного и экологического разнообразия природных ценопопуляций хозяйственно значимых растений, а также механизмов их адаптаций к среде обитания. T. pratense широко распространен в составе различных ландшафтов в фитоценозах открытого и полуоткрытого типа и имеет перспективы использования в качестве лекарственного растительного сырья.
Задачи настоящего исследования – выявить экологические характеристики местообитаний T. pratense, плотность и структуру его ценопопуляций, семенную продуктивность, а также запасы фитомассы на территории РТ с целью определения наиболее оптимальных условий для формирования высоко продуктивных ценопопуляций.
МАТЕРИАЛ И МЕТОДЫ
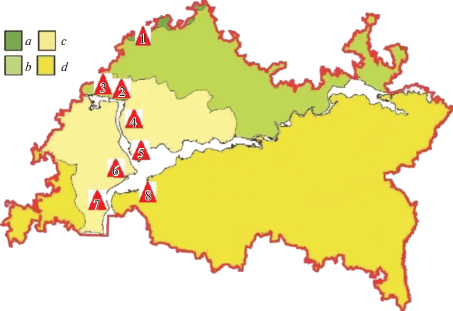
Сбор материала проводился в вегетационные периоды 2018–2019 гг. на территории 5 административных р-нов РТ: Высокогорского, Зеленодольского, Лаишевского, Тетюшского и Спасского. В различных природно-климатических условиях на западе РТ в радиусе около 150 км были исследованы 8 ценопопуляций (ЦП) Trifolium pratеnse (рис. 1). ЦП 1 (Высокогорский р-н) располагается в южно-таежной ландшафтной подзоне, ЦП 2 и 3 (Зеленодольский р-н) – в подтаежной подзоне, ЦП 4 и 5 (Лаишевский р-н), ЦП 6 (Тетюшский р-н) и 7 (Камско-Устьинский р-н) – в широколиственной подзоне. При этом ЦП 6 (Тетюшский р-н) находится на границе с подзоной лесостепи, а ЦП 8 (Спасский р-н) располагается на территории лесостепной подзоны. В каждом из сообществ с участием T. pratense закладывалось 5 площадок размером 1 × 1 м2, на которых выкапывались все растения T. pratense для гербаризации и дальнейшего морфометрического анализа. Геоботанические описания растительных сообществ проводились на площади 200–500 м2 в соответствии с методическими рекомендациями [16].
Рис. 1.
Расположение исследованных ЦП (1–8) в разных ландшафтных подзонах Республики Татарстан. Карта ландшафтного районирования [15]: a – южно-таежная ландшафтная подзона, b – подтаежная ландшафтная подзона, c – широколиственная ландшафтная подзона, d – лесостепная ландшафтная подзона. Fig. 1. The location of the studied coenopopulations (1–8) in different landscape subzones of the Republic of Tatarstan. Map of landscape zoning [15]: a – south taiga landscape subzone, b – coniferous-deciduous landscape subzone, c – broadleaf landscape subzone, d – forest-steppe landscape subzone.
Оценку условиям местообитания T. pratense проводили индикаторными методами по 6 прямодействующим факторам среды с использованием 6 оптимумных экологических шкал Г. Элленберга: освещенность (L), температурный режим (T), континентальность климата (K), влажность почвы (F), кислотность почвы (R), богатство почвы азотом (N) [17, 18]. Значения экологических факторов для местообитания каждой ЦП рассчитывали в программе EcoScaleWin по видовому составу сообществ и обилию видов в них [19, 20].
В процессе морфологического исследования у высушенных генеративных растений определялись следующие параметры: количество генеративных побегов на растении (NG), их высота (H), общее число соцветий (NF), воздушно-сухая фитомасса надземных побегов (W), фитомасса репродуктивных органов (WG), фитомасса листьев (WL), репродуктивное усилие (RE = WG/W) и фотосинтетическое усилие (LWR = WL/W). Коэффициент продуктивности в разных эколого-ценотических условиях рассчитывали как соотношение между количеством выполненных семян и заложившихся семязачатков, исходя из значений потенциальной и реальной семенной продуктивности соцветия (ПСП и РСП) [21].
При оценке онтогенетического состояния растений учитывали разработки Н.П. Крылова и Т.А. Работнова [22]. Плотность ЦП определяли на метровых площадках, встречаемость – на 10 м трансектах в пределах обследованной площади сообщества. Урожайность в разных местообитаниях определяли путем суммирования воздушно-сухой надземной фитомассы генеративных растений, собранных с 5-метровых площадок.
Все морфометрические данные и значения плотности обработаны статистически в программе Excel c определением среднего и его ошибки, стандартного отклонения и коэффициента вариации. Достоверность различий между средними значениями оценивали с помощью критерия Стьюдента при 5% уровне значимости. Объем выборок исследуемых ЦП составил от 12 до 43 растений. Для всех рассматриваемых факторов среды, морфометрических и популяционных параметров рассчитаны коэффициенты корреляции (R) с использованием пакета программ статистического и биометрико-генетического анализа в растениеводстве и селекции AGROS.
Латинские названия растений в работе приводятся в соответствии с международной базой данных The Plant List [23].
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Согласно геоботаническим описаниям, исследуемые ЦП Trifolium pratеnse входят в состав опушечных сообществ (ЦП 1–3), вторичных вейниково-разнотравных (ЦП 4, 5), овсяницево-разнотравных (ЦП 6, 7) и остепненных овсянициево-разнотравных (ЦП 8) лугов (табл. 1). В данных фитоценозах частное проективное покрытие T. pratense варьирует в пределах 15–35%. В соответствии с почвенным районированием территории РТ [14] ЦП 1–3 распространены на дерново-подзолистых почвах, ЦП 4–7 – на темно-серых лесных и серых лесных почвах, ЦП 8 – на светло-серых лесных почвах.
Таблица 1.
Особенности исследованных фитоценозов в разных ландшафтных районах на территории Республики Татарстан Table 1. Features of the studied phytocoenoses from different landscape subzones of the Republic of Tatarstan
| Ландщафтная подзона/ЦП Landscape subzones / Coenopopulation (CP) |
Ассоциация Association |
Проективное покрытие, %/ количество видов Projective cover, %/ Number of species |
Доминирующие виды
(проективное покрытие > 12%) Dominant species (projective cover > 12%) |
|---|---|---|---|
| Южнотаежная/ЦП 1 South taiga/CP 1 56°24′54″ с.ш./N 49°11′05″ в.д /E |
Опушка Querco-Tilietum nemoroherbosum (восточная экспозиция) Querco-Tilietum nemoroherbosum edge (eastern exposure) |
85/36 | Pimpinellasaxifraga L. (50%), Melilotusofficinalis (L.) Pall. (50%); Bromusinermis Leyss., Centaureajacea L., Cichoriumintybus L., Clinopodiumvulgare L., Fragariavesca L., Taraxacum officinale (L.) Webb ex F.H. Wigg., Trifoliumhybridum L., Trifoliumpratense L. (15%), Trifoliumrepens L. |
| Подтаежная/ЦП 2 Coniferous-deciduous/CP 2 55°83′68″ с.ш./N 48°78′53″ в.д./E |
Опушка Querco-Tilietum nemoroherbosum (южная экспозиция) Querco-Tilietum nemoroherbosum edge (southern exposure) |
90/49 | Dactylisglomerata L. (55%), Festucapratensis Huds. (45%) Berteroaincana (L.) DC., Campanulapatula L., Dianthusdeltoides L., Galiummollugo L., Lathyruspratensis L., Plantagomajor L, Stellariagraminea L., Trifoliumhybridum L., Trifoliumpratense L. (15%), T-rifoliumrepens L. |
| Подтаежная/ЦП 3 Coniferous-deciduous /CP 3 55°84′34″ с.ш./N 48°73′90″ в.д./E |
Опушка Querco-Tilietum nemoroherbosum (южная экспозиция) Querco-Tilietum nemoroherbosum edge (southern exposure) |
85/53 | Calamagrostis epigejos (L.) Roth (40%), Trifolium pratense L. (25%), Achillea millefolium L., Cichorium intybus L., Dactylis glomerata L., Festuca pratensis Huds., Leontodon autumnalis L., Leucanthemum vulgare Lam., Medicago sativa L., Plantago major L., Silene viscaria (L.) Borkh., Taraxacum officinale (L.) Webb ex F.H. Wigg., Trifolium medium L., Trifolium repens L., Vicia sepium L. |
| Широколиственная/ЦП 4 Broadleaf/CP 4 55°53′60″ с.ш./N 49°13′77″ в.д./E |
Calamagrostetum varioherbosum | 90/36 | Calamagrostisepigejos (L.) Roth (70%), Trifoliumpratense L. (25%), Centaureascabiosa L., Convolvulusarvensis L., Equisetumarvense L., Eryngiumplanum L., Leontodonhispidus L., Pimpinellasaxifraga L., Poaangustifolia L., Jacobaeavulgaris Gaertn. |
| Широколиственная/ЦП 5 Broadleaf/CP 5 55°28′99″ с.ш./N 49°37′23″ в.д./E |
Calamagrostetum varioherbosum | 65/35 | Calamagrostisepigejos (L.) Roth (35%), Trifoliumpratense L. (25%), Dianthus fischeri Spreng., Festuca valesiaca Schleich. ex Gaudin, Fragariaviridis Weston, Hypericumperforatum L., Leucanthemumvulgare (Vaill.) Lam. (50%), Pimpinellasaxifraga L., Prunellavulgaris L. |
| Широколиственная/ЦП 6 Broadleaf/CP 6 55°16′50″ с.ш./N 48°96′07″ в.д./E |
Festucetum varioherbosum | 70/27 | Festucapratensis Huds. (50%), Trifoliumpratense L. (30%), Astragaluscicer L., Carduuscrispus Guirão ex Nyman, Fragariaviridis Weston, Knautiaarvensis (L.) Coult., Lavaterathuringiaca L., Lotuscorniculatus L., Melilotusofficinalis (L.) Pall., Plantagolanceolata L., Potentillaargentea L., Trifoliumrepens L. |
| Широколиственная/ЦП 7 Broadleaf /CP 7 54°92′15″ с.ш./N 48°81′85″ в.д./E |
Festucetum varioherbosum | 60/20 | Festucapratensis Huds. (45%), Trifoliumpratense L. (35%), Achilleamillefolium L., Medicagosativa L., Melilotusofficinalis (L.) Pall., Phleumpratense L., Rumexconfertus Willd., Tripleurospermuminodorum (L.) Sch. Bip., Trifoliumrepens L. |
| Лесостепная/ЦП 8 Forest-steppe/CP 8 54°97′86″ с.ш./N 49°04′93″ в.д./E |
Valesiaca festucetum varioherbosum | 90/26 | Festucavalesiaca Schleich. ex Gaudin (45%), Trifoliumpratense L. (35%), Achilleamillefolium L., Artemisiacampestris L., Astragaluscicer L., Cichoriumintybus L., Medicagofalcata L., Plantagolanceolata L., Plantagomedia L., Poaangustifolia L., Scabiosaochroleuca L. |
Согласно расчету значений экологических факторов на основе шкал Г. Элленберга для местообитаний исследованных ЦП T. pratense характерны следующие параметры: освещенность 50% и более от полной (L = 7.03–7.74), климат умеренно теплый (Т = 5.65–6.09) от субокеанического до слабо субконтинентального (К = 4.03–4.77), почвы от сухих периодически свежих до свежих (F = = 3.78–4.87), нейтральные или слабокислые (R = = 6.01–7.36), бедные или со средним содержанием минерального азота (N = 3.28–5.45) (табл. 2).
Таблица 2.
Характеристика местообитаний Trifolium pratense в Республике Татарстан по экологическим шкалам Г. Элленберга Table 2. Characteristics of Trifolium pratense habitats in the Republic of Tatarstan according to Ellenberg ecological indicator values
| Ландщафтная подзона/ЦП Landscape subzones/ Coenopopulation (CP) |
Значения экологических факторов (баллы) Values of ecological indicators (points) |
|||||
|---|---|---|---|---|---|---|
| L | T | K | F | R | N | |
| Южнотаежная/ЦП 1 South taiga/CP 1 |
7.46 | 5.92 | 4.51 | 4.41 | 7.36 | 4.75 |
| Подтаежная/ЦП 2 Coniferous-deciduous/CP 2 |
7.03 | 5.65 | 4.03 | 4.87 | 6.01 | 5.22 |
| Подтаежная/ЦП 3 Coniferous -deciduous/CP 3 |
7.2 | 5.72 | 4.28 | 4.76 | 6.31 | 5.06 |
| Широколиственная/ЦП 4 Broadleaf/CP 4 |
7.37 | 5.76 | 4.3 | 3.78 | 7.04 | 3.36 |
| Широколиственная/ЦП 5 Broadleaf/CP 5 |
7.28 | 5.95 | 4.2 | 3.93 | 6.85 | 3.5 |
| Широколиственная/ЦП 6 Broadleaf/CP 6 |
7.42 | 5.98 | 4.21 | 4.3 | 7.1 | 4.46 |
| Широколиственная/ЦП 7 Broadleaf subzone/CP 7 |
7.74 | 5.94 | 4.67 | 4.57 | 7.1 | 5.45 |
| Лесостепная/ЦП 8 Forest-steppe/CP 8 |
7.44 | 6.09 | 4.77 | 3.99 | 7.17 | 3.28 |
Примечание: L – освещенность, T – температурный режим, K – континентальность климата, F – влажность почвы, R – кислотность почвы, N – богатство почвы азотом. Note: L – light availability, T – temperature condition, K – climatic continentality, F – soil moisture, R – soil acidity, N – soil richness with nitrogen.
Наиболее сухие почвы характеризуются сравнительно низким содержанием минерального азота, между этими параметрами отмечена высокая положительная корреляция (R = 0.93). В фитоценозах с участием T. pratense наибольшее варьирование характерно для параметров почвы. Анализ значений экологических факторов показал, что наименьшее увлажнение почв (сухие периодически свежие) и низкое содержание в них азота отмечено в местообитаниях ЦП 4, 5 (Calamagrostetum varioherbosum, широколиственная подзона) и ЦП 8 (Valesiaca festucetum varioherbosum, лесостепная подзона). Свежие почвы со средним содержанием азота и преимущественно слабокислой реакцией выявлены в местообитаниях ЦП 1 (опушка сообщества Querco-Tilietum nemoroherbosum, южнотаежная подзона), ЦП 2, 3 (опушка сообщества Querco-Tilietum nemoroherbosum, подтаежная подзона), а также ЦП 6, 7 (Festucetum varioherbosum, широколиственная подзона).
Согласно полученным данным, у генеративных растений T. pratense в среднем формируется 3–7 генеративных побегов, средняя высота которых в разных районах РТ варьирует от 29 (ЦП 6) до 48 см (ЦП 2) (табл. 3, 4). При этом в разных условиях произрастания на одном растении может формироваться в среднем от 3 (ЦП 8) до 17 (ЦП 3) соцветий. Средние значения общей надземной фитомассы генеративных растений варьирует от 2.4 (ЦП 5) до 8.5 г (ЦП 3). Доля листьев в составе общей фитомассы растения (LWR) составляет в среднем от 0.22 до 0.36, в то время как репродуктивное усилие (RE) находится в пределах 0.18–0.29. С-огласно значениям коэффициента вариации, в наименьшей степени варьирует высота растения (V = 20–40%). Вариабельность остальных параметров выше 40%. Пределы изменчивости числа генеративных побегов (NG) в большинстве случаев составляют 40–70%, числа соцветий (NF) и общей фитомассы (W) – 53–98%, массы листьев (WL) и массы соцветий (WG) – 65–99%.
Таблица 3.
Параметры генеративных растений Trifolium pratense в южно-таежной и подтаежной подзонах Республики Татарстан Table 3. Parameters of Trifolium pratense generative plants in south taiga and mixed coniferous-deciduous subzones of the Republic of Tatarstan
| Параметры Parameters |
ЦП 1 CP 1 |
ЦП 2 CP 2 |
ЦП 3 CP 3 |
|||
|---|---|---|---|---|---|---|
| X mean |
V, % coefficient of variation, % |
X mean |
V, % coefficient of variation, % |
X mean |
V, % coefficient of variation, % |
|
| H, см Н, cm |
34.3 ± 1.2 | 20.3 | 48.1 ± 5.8 | 28.9 | 39.7 ± 3.4 | 19.7 |
| NG, шт. NG, pcs. |
3.8 ± 0.5 | 73.2 | 5.7 ± 0.6* | 38.6 | 8.6 ± 0.3** | 68.7 |
| NF, шт. NF, pcs. |
11.0 ± 1.8 | 86.5 | 11.1 ± 0.5 | 23.8 | 16.7 ± 0.2** | 53.5 |
| W, г W, g |
3.6 ± 0.5 | 81.6 | 6.0 ± 0.7* | 64.9 | 8.5 ± 0.3*** | 33.3 |
| WL, г WL, g |
1.0 ± 0.2 | 86.6 | 1.3 ± 0.03 | 84.3 | 1.9 ± 0.1*** | 75.1 |
| WG, г WG, g |
0.7 ± 0.1 | 94.02 | 0.9 ± 0.02 | 50.3 | 1.9 ± 0.04*** | 63.9 |
| LWR | 0.3 ± 0.01 | 30.7 | 0.2 ± 0.1 | 20.8 | 0.2 ± 0.1 | 32.1 |
| RE | 0.2 ± 0.02 | 52.7 | 0.2 ± 0.1 | 42.1 | 0.2 ± 0.1 | 23.9 |
| n (объем выборки) n (sample size) |
32 | 12 | 14 | |||
Примечание. H – высота; NG – количество генеративных побегов на растении; NF – общее число соцветий, W – общая надземная фитомасса одного растения; WG – фитомасса репродуктивных органов; WL – фитомасса листьев; RE (WG/W) – репродуктивное усилие; LWR (WL/W) – фотосинтетическое усилие. * – величина достоверно отличается от соответствующей величины ЦП 1. ** – величина достоверно отличается от соответствующих величин ЦП 1 и 2. *** – величина достоверно отличается от соответствующих величин ЦП 1, 2 и 4–8 (табл. 4). Note. H – shoot height; NG – number of generative shoots per plant; NF – total number of inflorescences per plant; W – aboveground phytomass of an individual plant; WG – inflorescence mass; WL – leaf mass; RE (WG/W) – reproductive effort; LWR (WL/W) – photosynthetic effort. * – the value significantly differs from the corresponding value of the CP 1. ** – the value significantly differs from the corresponding values of CP 1 and 2. *** – the value significantly differs from the corresponding values of CP 1, 2, and 4–8 (table 4).
Таблица 4.
Параметры генеративных растений Trifolium pratense в широколиственной и лесостепной подзонах Республики Татарстан Table 4. Parameters of Trifolium pratense generative plants in broadleaf and forest-steppe subzones of the Republic of Tatarstan
| Параметры Parameters |
ЦП 4 CP 4 |
ЦП 5 CP 5 |
ЦП 6 CP 6 |
ЦП 7 CP 7 |
ЦП 8 CP 8 |
|||||
|---|---|---|---|---|---|---|---|---|---|---|
| X mean |
V, % coefficient of variation, % |
X mean |
V, % coefficient of variation, % |
X mean |
V, % coefficient of variation, % |
X mean |
V, % coefficient of variation, % |
X mean |
V, % coefficient of variation, % |
|
| H, см Н, cm |
32.5 ± 2.0 | 39.9 | 38.1 ± 1.5 | 25.9 | 28.9 ± 0.7 | 28.4 | 32.3 ± 1.1 | 24.1 | 29.9 ± 0.9 | 34.5 |
| NG, шт. NG, pcs. |
2.5 ± 0.4 | 85.3 | 2.5 ± 0.2 | 57.9 | 7.0 ± 0.9* | 58.8 | 3.5 ± 0.4 | 44.8 | 3.3 ± 0.3 | 63.2 |
| NF, шт. NF, pcs. |
6.8 ± 1.1 | 96.2 | 8.9 ± 1.3 | 94.1 | 14.2 ± 1.9 | 57.6 | 10.1 ± 1.7 | 63.8 | 2.9 ± 0.2** | 74.7 |
| W, г W, g |
2.6 ± 0.4 | 98.1 | 2.4 ± 0.3 | 77.9 | 5.3 ± 0.9 | 72.6 | 3.4 ± 0.5 | 53.2 | 3.2 ± 0.5 | 93.0 |
| WL, г WL, g |
0.4 ± 0.04 | 64.5 | 0.4 ± 0.1 | 76.9 | 1.2 ± 0.3 | 93.7 | 0.7 ± 0.1 | 32.8 | 0.7 ± 0.1 | 97.7 |
| WG, г WG, g |
0.6 ± 0.1 | 94.4 | 0.4 ± 0.1 | 87.2 | 0.8 ± 0.1 | 68.7 | 1.1 ± 0.21 | 75.6 | 0.7 ± 0.1 | 98.7 |
| LWR | 0.4 ± 0.1 | 95.9 | 0.2 ± 0.02 | 54.5 | 0.2 ± 0.01 | 30.2 | 0.3 ± 0.04 | 65.2 | 0.2 ± 0.01 | 42.4 |
| RE | 0.2 ± 0.02 | 50.01 | 0.2 ± 0.02 | 78.9 | 0.2 ± 0.1 | 68.7 | 0.3 ± 0.2 | 75.6 | 0.2 ± 0.01 | 38.6 |
| n (объем выборки) n (sample size) |
37 | 43 | 15 | 18 | 38 | |||||
Примечание. H – высота; NG – количество генеративных побегов на растении; NF – общее число соцветий, W – надземная фитомасса одного растения; WG – фитомасса репродуктивных органов; WL – фитомасса листьев; RE (WG/W) – репродуктивное усилие; LWR (WL/W) – фотосинтетическое усилие. * – величина достоверно отличается от соответствующих величин ЦП 4, 5, 7 и 8. ** – величина достоверно отличается от соответствующих величин ЦП 1–3 (табл. 3) и 4–7. Note. H – shoot height; NG – number of generative shoots per plant; NF – total number of inflorescences per plant; W – aboveground phytomass of an individual plant; WG – inflorescence mass; WL – leaf mass; RE (WG/W) – reproductive effort; LWR (WL/W) – photosynthetic effort. * – the value significantly differs from the corresponding values of CP 4, 5, 7 and 8. ** – the value significantly differs from the corresponding values of CP 1–3 (tab. 3) и 4–7.
Генеративные растения T. pratense на свежих почвах со средним содержанием минерального азота в ЦП 2, 3 (подтаежная подзона) и ЦП 6 (юг широколиственной подзоны) отличаются более высоким морфологическим статусом. Так, в ЦП 2, 3, 6 по сравнению с ЦП 4, 5 достоверно выше количество побегов у растений (в 2.5–3.5 раза) и общая надземная фитомасса (в 1.8–4 раза). В ЦП 3 число соцветий (головок) в 1.7–5.7 раз превышает соответствующее значение в ЦП 4, 5, 8. ЦП 4, 5, 8, распространенные в условиях наименьшего увлажнения и минерального богатства почв оказались наименее продуктивными (табл. 4).
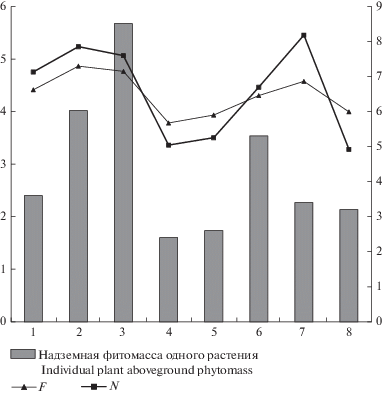
Общая надземная фитомасса растения имеет закономерную положительную корреляцию с числом генеративных побегов (R = 0.97; p < 0.05), массой репродуктивных органов (R = 0.84; p < 0.05) и массой листьев (R = 0.99; p < 0.05). Сравнение изменчивости значений влажности (F) и богатства (N) почвы по шкалам Г. Элленберга с показателем общей надземной фитомассы растений T. pratense в разных ЦП показало, что уменьшение влажности и обеспеченности почвы минеральным азотом приводит к снижению их фитомассы в ЦП 4, 5, 8 (рис. 2). Это подтверждается и выявленной положительной корреляцией между общей фитомассой и балльной оценкой влажности почвы (R = 0.86; p < 0.05). Сходная закономерность выявлена в отношении связи количества генеративных побегов на растении и массы репродуктивных органов с уровнем влажности почвы (соответственно R = 0.78 и 0.86; p < 0.05).
Рис. 2.
Соотношение показателей влажности, богатства почвы и надземной фитомассы одного растения в ЦП Trifolium pratеnse на территории Республики Татарстан. F – влажность почвы, N – богатство почвы минеральным азотом. По горизонтали – ценопопуляции; по вертикали (левая шкала) – балльная оценка факторов среды по шкалам Г. Элленберга; по вертикали (правая шкала) – надземная фитомасса одного растения, г. Fig. 2. Correlation of moisture, soil richness and Trifolium pratense individual plant aboveground phytomass in coenopopulations in the Republic of Tatarstan. F – soil moisture, N – soil richness with mineral nitrogen. X-axis – coenopopulations; y-axis (left scale) – values of environmental factors according to Ellenberg’s ecological indicators; y-axis (right scale) – aboveground phytomass of an individual plant, g.
Встречаемость T. pratense в изученных сообществах составляет 100%, плотность ЦП варьирует от 3 до 17.2 экз./м2 в зависимости от экологических условий (табл. 5). Общая надземная фитомасса растения, масса листьев, число генеративных побегов и масса репродуктивных органов имеют тесную отрицательную корреляцию с плотностью ЦП (R = –|0.85–0.89|; p < 0.05). В ЦП 4, 5 отмечается наибольшая плотность (16.6–17.2 экз./м2), в 2–5 раз превышающая плотность других ЦП. Самая низкая плотность T. pratense (3–3.8 экз./м2) выявлена в ЦП 2, 3 и ЦП 6. Для растений в ЦП 2, 3, 6 характерны наибольшие значения общей надземной фитомассы, массы листьев, числа побегов и наименьшая плотность.
Таблица 5.
Плотность ценопопуляций Trifolium pratense на территории Республики Татарстан Table 5. Density of Trifolium pratense coenopopulations in the Republic of Tatarstan
| Ландшафтная подзона/ЦП Landscape subzone/Coenopopulation (CP) |
Плотность, экз./м2 Density, ind./m2 |
|---|---|
| Южнотаежная/ЦП 1 South taiga/CP 1 |
9.5 ± 4.7 |
| Подтаежная/ЦП 2 Coniferous-deciduous/CP 2 |
3.0 ± 0.4 |
| Подтаежная/ЦП 3 Coniferous-deciduous/CP 3 |
3.5 ± 0.3 |
| Широколиственная/ЦП 4 Broadleaf/CP 4 |
17.2 ± 9.7 |
| Широколиственная/ЦП 5 Broadleaf subzone/CP 5 |
16.8 ± 4.2 |
| Широколиственная/ЦП 6 Broadleaf/CP 6 |
4.5 ± 0.9 |
| Широколиственная/ЦП 7 Broadleaf/CP 7 |
5.0 ± 0.6 |
| Лесостепная/ЦП 8 Forest-steppe/CP 8 |
8.8 ± 1.8 |
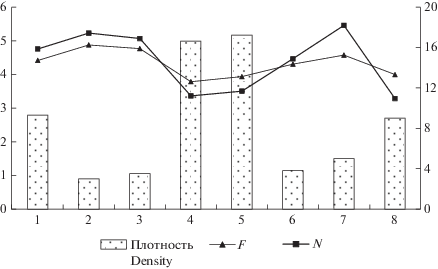
Проведенный анализ соотношения оценок экологических факторов (F и N) и плотности ЦП показал, что с уменьшением влажности и количества минерального азота в почве достоверно увеличивается плотность ЦП T. pratense (рис. 3). Это подтверждают значения коэффициентов корреляции между плотностью ЦП и влажностью почвы (R = –0.87; p < 0.05) и между плотностью и богатством почвы (R = –0.79; p < 0.05). Наиболее высокая плотность характерна для ЦП 4 и 5, распространенных на сухих периодически свежих почвах с низким содержанием минерального азота.
Рис. 3.
Соотношение показателей влажности, богатства почвы и плотности ЦП Trifolium pratеnse на территории Республики Татарстан. F – влажность почвы, N – богатство почвы минеральным азотом. По горизонтали – ценопопуляции, по вертикали слева – балльная оценка факторов среды по шкалам Г. Элленберга, по вертикали справа – плотность ценопопуляций, экз./м2. Fig. 3. Correlation of moisture, soil richness and density of Trifolium pratense coenopopulations in the Republic of Tatarstan. F – soil moisture, N – soil richness with mineral nitrogen. X-axis – coenopopulations; y-axis (left scale) – values of environmental factors according to Ellenberg’s ecological indicators; y-axis (right scale) – density of coenopopulations, inds./m2.
Разная плотность может быть связана с особенностями самоподдержания ЦП семенным путем. По замечанию Л.А. Жуковой [24] для ЦП стержнекорневых олиго- и поликарпиков характерно резкое варьирование количества генеративных и вегетативных растений, при этом доля генеративных растений может колебаться от 1 до 78%, доля ювенильных растений и их приживаемость составляет от 0 до 20%. По нашим данным в ЦП 4, 5 отмечается значительное участие (около 50%) растений прегенеративного периода, в то же время ЦП 2, 3, 6, 7 состоят только из генеративных растений (рис. 4). Отсутствие молодых вегетативных растений в этих ЦП может быть связано с быстрыми темпами прохождения прегенеративных стадий в условиях повышенной влажности и богатства почвы. В работе Н.П. Крылова и Т.А. Работнова [22] отмечается, что благоприятные условия увлажнения, богатства почвы доступными для растений элементами, а также раннее скашивание сокращают продолжительность пребывания особей T. pratense в ювенильном и в виргинильном состояниях, при этом в благоприятных условиях T. pratense моно- или дикарпичное растение, в малоблагоприятных – поликарпик.
Рис. 4.
Соотношение прегенеративных и генеративных растений в ценопопуляциях Trifolium pratеnse. По горизонтали – доля участия, %; по вертикали – ценопопуляции. Fig. 4. The ratio of pregenerative and generative plants in the Trifolium pratеnse coenopopulations. X-axis – share of pregenerative and generative plants, %; y-axis –coenopopulation.

Выборочное изучение репродуктивных особенностей T. pratense (ЦП 1, 4) показало, что к-оэффициент семенной продуктивности, как соотношение числа выполненных семян и заложившихся семязачатков, находится в пределах 12–40% и его значения сопоставимы с литературными данными. Например, в горных районах Осетии этот показатель у дикорастущего T. pratense составляет 25–45% [25]. В ЦП 1 в одном соцветии образуется от 1 до 39 семян, из-за части невыполненных семян реальная семенная продуктивность соцветия в среднем составила 6 семян (табл. 6). Коэффициент продуктивности в этих условиях низкий, около 12%. В соцветиях растений ЦП 4 по сравнению с ЦП 1 формируется в 1.4 раза больше цветков и в 3.7 раз больше семян, при этом коэффициент семенной продуктивности составляет около 40%. Следовательно, значительное участие молодых растений прегенеративного периода и высокая плотность в ЦП 4 объясняется высокой семенной продуктивностью.
Таблица 6.
Показатели репродукции в изученных ценопопуляциях Trifolium pratеnse на территории Республики Татарстан Table 6. Trifolium pratense reproduction parameters in the studied coenopopulations from the Republic of Tatarstan
| Ландшафтная
подзона/ЦП Landscape subzone/ Coenopopulation (CP) |
Число цветков в соцветии Number of flowers in inflorescence |
Число семян в соцветии Number of seeds in inflorescence |
Число выполненных
семян в соцветии, экз. Number of filled seeds per inflorescence, pcs |
Коэффициент
продуктивности, % Coefficient of seed productivity,% |
||
|---|---|---|---|---|---|---|
| M | D | M | D | |||
| Южнотаежная/ЦП 1 South taiga/CP 1 |
51.4 | 10–104 | 8.9 | 1–39 | 6.0 | 12 |
| Широколиственна/ЦП 4 Broadleaf/CP 4 |
69.4 | 47–107 | 33.0 | 7–92 | 26.2 | 38 |
По нашим данным, в изученных ценопопуляциях различных ландшафтных подзон на территории РТ запасы сухой надземной фитомассы T. pratense варьируют от 170 до 432 кг/га (табл. 7). На территории Удмуртии (подзона южной тайги) при культивировании T. pratense многолетняя среднесортовая урожайность составляет 145–157 кг/га и может варьировать от погодных условий и количества укосов от 40 до 282 кг/га [26]. В то же время, в горах Кабардино-Балкарии в составе горных послелесных лугов на выщелочных черноземах урожайность надземной фитомассы T. pratense составляет 2.4–3.5 т/га [27], это в 6–8 раз выше по сравнению с РТ.
Таблица 7.
Запас растительного сырья Trifolium pratеnse в различных ландшафтных подзонах на территории Республики Татарстан Table 7. Stock of Trifolium pratens aboveground phytomass of in different landscape subzones of the Republic of Tatarstan
| Ландщафтная подзона / ЦП Landscape subzones / Coenopopulation (CP) |
Надземная фитомасса, воздушно-сухой вес Aboveground phytomass, air-dry weight |
|
|---|---|---|
| г/м2 g/m2 |
кг/га kg/ha |
|
| Южнотаежная/ЦП 1 South taiga/CP 1 |
33.8 | 338 |
| Подтаежная/ЦП 2 Coniferous-deciduous/CP 2 |
18.6 | 186 |
| Подтаежная/ЦП 3 Coniferous-deciduous/CP 3 |
29.8 | 298 |
| Широколиственная/ЦП 4 Broadleaf/CP 4 |
43.2 | 432 |
| Широколиственная/ЦП 5 Broadleaf/CP 5 |
41.3 | 413 |
| Широколиственная/ЦП 6 Broadleaf/CP 6 |
20.1 | 201 |
| Широколиственная/ЦП 7 Broadleaf/CP 7 |
17.0 | 170 |
| Лесостепная/ЦП 8 Forest-steppe/CP 8 |
28.8 | 288 |
Расчеты показали, что сравнительно низкая урожайность надземной фитомассы (170–201 кг/га) T. pratense характерна для ЦП 2, 3, 6 и 7, формирующихся на свежих почвах со средним содержанием минерального азота. Это связано с низкой плотностью и наличием в ЦП только генеративных растений высокого морфологического статуса. Наибольшая урожайность надземной фитомассы T. pratense (413–432 кг/га) выявлена в ЦП 4, 5 в условиях сухих периодически свежих и бедных почв, для которых характерны растения среднего морфологического статуса, высокая плотность, участие в составе как генеративных, так и молодых вегетативных растений.
ЗАКЛЮЧЕНИЕ
На территории Республики Татарстан частное проективное покрытие Trifolium pratense в составе опушечных сообществ (южно-таежная и подтаежная подзоны), вторичных вейниково-разнотравных и овсяницево-разнотравных лугов (широколиственная подзона), а также остепненных овсянициево-разнотравных лугов (лесостепная подзона) варьирует в пределах 15–35%. Условия местообитания ЦП T. pratense различаются преимущественно по параметрам почв. В соответствии с экологическими шкалами Г. Элленберга, почвы изученных сообществ по увлажнению составляют ряд от сухих периодически свежих до свежих (F = 3.78–4.87), по содержанию азота – от бедных до средних (N = 3.36–5.45), по кислотности – от нейтральных до слабокислых (R = = 6.01–7.36).
Исследования показали наличие достаточно тесной положительной связи между величиной общей фитомассы растений, числом генеративных побегов, массой репродуктивных органов T. pratense и влажностью почвы (R = 0.78–0.86), а также наличие отрицательной связи между плотностью ЦП и уровнем влажности и богатства почвы азотом (R = –0.87 и –0.79).
В сообществах на свежих почвах со средним содержанием азота почв в пределах РТ ЦП T. pratense имеют низкую плотность (3–5 экз./м2), в их составе преобладают генеративные растения высокого морфологического статуса, у которых число генеративных побегов выше в 2.5–3.5 раза, число соцветий – в 1.7–5.7 раз, надземная фитомасса – в 1.8–4 раза, чем у растений других исследованных ЦП.
На наиболее сухих и бедных минеральным азотом почвах в пределах РТ ЦП T. pratense образованы растениями низкого морфологического статуса. Однако, в этих условиях ЦП отличаются значительной плотностью (16.6–17.2 экз./м2) и большой долей участия растений прегенеративного периода (около 50%). Наличие молодых вегетативных растений объясняется высоким коэффициентом семенной продуктивности, значения которого составляют около 40%. По нашим данным для ЦП высокой плотности характерна наибольшая урожайность воздушно-сухой надземной фитомассы (413–432 кг/га). Территории с такими экологическими характеристиками в условиях РТ можно рекомендовать для заготовки сырья T. pratense.
Список литературы
Бакин О.В., Рогова Т.В., Ситников А.П. 2000. Сосудистые растения Татарстана. Казань. 496 с.
Безделев А.Б., Безделева Т.А. 2006. Жизненные формы семенных растений Российского Дальнего Востока. Владивосток. 295с.
Бобров Е.Г. 1987. Клевер Trifolium L. – В кн.: Флора Европейской части СССР. Л. Т. 6. С. 195–208.
Растительные ресурсы России: Дикорастущие цветковые растения, их компонентный состав и биологическая активность. 2010. Т.3. Семейства Fabaceae – Apiaceae. СПб.; М. 601с.
Дренин А.А., Ботиров Э.Х. 2017. Флавоноиды и изофлаваноиды растений рода Trifolium L. Структурное разнообразие и биологическая активность. – Химия растительного сырья. 3: 39–53. https://doi.org/10.14258/jcprm.2017031646
Зайцева Н.В., Погуляева И.А. 2017. Эколого-биохимические особенности растений рода Trifolium L., произрастающих в Южной Якутии (на примере г. Нерюнги). – Известия Самарского НЦ РАН. 19 (2): 441–447. http://www.ssc.smr.ru/media/journals/izvestia/2017/2017_2_441_447.pdf
Booth N.L., Overk C.R., Yao P., Burdette J.E., Nikolic D., Chen S.-N., Bolton J.L., Breemen R.B. van, Pauli G.F., Farnsworth N.R. 2006. The chemical and biologic profile of a red clover (Trifolium pratense L.). Phase II clinical extract – J. Altern. Complement. Med. 12(3): 133–139. https://doi.org/10.1089/acm.2006.12.133
Sabudak T., Guler N. 2009. Trifolium L. – a review on its phytochemical and pharmacological profile – Phytother. Res. 23(3): 439–446. https://doi.org/10.1002/ptr.2709
Kaurinovic B., Popovic M., Vlaisavljevic S., Schwartsova H., Vojinovic-Miloradov M. 2012. Antioxidant Profile of Trifolium pratense L. – Molecules. 17(9): 11156–11172. https://doi.org/10.3390/molecules170911156
Злобин Ю.А. 1989. Принципы и методы изучения ценотических популяций растений. Казань. 145 с.
Созинов О.В., Кузьмичева Н.А. 2003. Ценопопуляции Ledum palustre L. и их сырьевая характеристика в условиях Средненеманской низины (Республика Беларусь). – Растительные ресурсы. 39(3): 55–62. https://elibrary.ru/item.asp?id=17080755
Созинов О.В. 2015. Морфологические и ресурсные характеристики Calluna vulgaris (Ericaceae) в нарушенных фитоценозах северо-запада Республики Беларусь. – Растительные ресурсы. 51(4): 473–490. https://elibrary.ru/item.asp?id=24347081
Созинов О.В., Кузьмичева Н.А. 2016. Ресурсно-фитохимическая изменчивость и биоэкологическая характеристика Vaccinium vitis-idaea (Ericaceae) в сосняке мшистом на орографическом градиенте (Республика Беларусь). – Растительные ресурсы. 52 (2): 202–214. https://elibrary.ru/item.asp?id=25818688
Ландшафты Республики Татарстан. Региональный ландшафтно-экологический анализ. 2007. Казань. 411с.
Ермолаев О.П., Игонин М.Е. 2006. Ландшафтное районирование и картографирование региона Среднего Поволжья. – Георесурсы. 2(19): 20–23. https://geors.ru/media/pdf/06_Ermolaev.pdf
Голуб В.Б. 2011. Использование геоботанических описаний в качестве коллекции образцов для классификации растительности. – Растительность России. 17(18): 70–83. https://doi.org/10.31111/vegrus/2011.17-18.70
Ellenberg H., Weber H.E., Düll R., Wirth V., Werner W., Paulißen D. 1991. Zeigerwerte von Pflanzen in Mitteleuropa. – Scr. Geobot. 18: 1–248.
Schaffers A.P., Sýkora K.V. 2000. Reliability of Ellenberg indicator values for moisture, nitrogen and soil reaction: a comparison with field measurements. – Journal of Vegetation Science. 11(2): 225–244. https://doi.org/10.2307/3236802
Зубкова Е.В., Ханина Л.Г., Грохлина Т.И., Дорогова Ю.А. 2008. Компьютерная обработка геоботанических описаний по экологическим шкалам с помощью программы EcoscaleWin: учеб. пособие. Йошкар-Ола. 96 с.
Экологические шкалы и методы анализа экологического разнообразия растений. 2010. Йошкар-Ола. 368с.
Левина Р.Е. 1981. Репродуктивная биология семенных растений. М. 96 с.
Крылов Н.П., Работнов Т.А. 1975. Клевер луговой – В сб.: Биологическая флора Московской области. 2. М. С. 89–101.
The Plant List. 2013. http://www.theplantlist.org
Жукова Л.А. 1995. Популяционная жизнь луговых растений. Йошкар-Ола. 224 с.
Бекузарова С.А., Комжа А.Л., Соколова Л.Б. 2016. Репродуктивные особенности дикорастущих видов клевера горных фитоценозов. – Бюллетень ГНБС. 120: 49–54.
Касаткина Н.И., Нелюбина Ж.С. 2016. Продуктивность сортов клевера лугового в условиях Среднего Предуралья. – Аграрная наука Евро-Северо-Востока. 5(54): 31–36. https://www.agronauka-sv.ru/jour/article/view/66/66
Фисун М.Н., Тамахина А.Я., Гадиева А.А. 2011. Оценка ресурсов Galega orientalis, Trifolium prаtense и Lotus corniculatus (Fabaceae) в фитоценозах горной зоны Кабардино-Балкарии. – Раст. ресурсы. 47(3): 1–8. https://elibrary.ru/item.asp?id=16521924
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы