

Растительные ресурсы, 2021, T. 57, № 1, стр. 39-48
Влияние климатических факторов на радиальный прирост Pinus sylvestris и Picea abies (Pinaceae) на территории Брянской области
В. П. Иванов 1, *, С. И. Марченко 1, Д. И. Нартов 1, Л. П. Балухта 1
1 Брянский государственный инженерно-технологический университет
г. Брянск, Россия
* E-mail: ivpinfo@mail.ru
Поступила в редакцию 13.03.2020
После доработки 04.06.2020
Принята к публикации 10.12.2020
Аннотация
Исследованы особенности роста по диаметру Pinus sylvestris L. и Picea abies (L.) Н. Karst (Pinaceae) в сходных лесорастительных условиях (переходных от свежих боровых к свежим суборевым) на территории Брянской обл. Выполнена оценка влияния на величину радиального прироста среднемесячных температур, сумм осадков текущего года и предыдущих лет, величины прироста предыдущего года. Выявлена значимая корреляция величины радиальных приростов и индексов радиальных приростов у изучаемых видов, подтвержденная индексированными хронологиями. В отдельные годы наблюдаются выраженные синхронные изменения величины прироста: заметное снижение и увеличение (относительно среднего значения). Значимое влияние на величину радиального прироста в районе исследований оказывают прирост предыдущего года и средняя месячная температура мая текущего года. Глобальные изменения температурного режима за последние десятилетия могут негативно отражаться на характере ростовых процессов видов, сформировавшихся в условиях бореального климата. Существенного влияния количества осадков на ширину годичного кольца изучаемых видов не выявлено.
Погодные аномалии начала XXI в. в виде высоких температур воздуха, увеличения продолжительности засушливых периодов привели к значительному ослаблению позиций хвойных лесообразующих видов в центральной части России. На этом фоне возросло поражение еловых древостоев короедом типографом, особенно на южной границе ареала, а последующие санитарные рубки привили к значительному сокращению их площади. Происходящие изменения в динамике роста хвойных насаждений можно проследить на основе анализа радиальных приростов сосны обыкновенной Pinus sylvestris L. (Pinaceae) и ели европейской Picea abies (L.) Н. Karst (Pinaceae). На территории Брянской обл. ареалы этих древесных пород совпадают, однако южная граница ареала P. sylvestris проходит несколько южнее.
Радиальный прирост древесины – сложный интегральный показатель, источник длительной ретроспективной информации о характере ростовых процессов, поэтому использование методов дендрохронологии для оценки реакции древесных растений на воздействие внешних факторов получило заслуженное признание лесоводов [1–3]. Диапазон исследований последних лет в области дендрохронологии значительно расширился в нашей стране и за рубежом, хотя исследования носят большей частью региональный и локальный характер [4–7].
Помимо акцентирования внимания на периодах с минимальными значениями радиального прироста исследуемых пород и выявления климатических факторов, негативно на него влияющих, сделаны попытки более глубокого анализа особенностей радиального роста различных видов–лесообразователей [8]. Следует отметить анализ распределения клеток в древесине сосны по диаметру и толщине стенок, что, по мнению авторов, является индикатором адаптации структуры древесины к недостатку влаги [9] и оценку влияния климатических факторов на клеточное строение годичных колец лиственницы Гмелина в контрастных гидротермических условиях лесотундры [10]. Исследовано влияние климатических параметров на формирование радиального прироста сосны на северной границе ареала в Европейской части России [11]. Выполнен анализ многолетней изменчивости анатомических параметров годичных колец хвойных пород в криолитозоне Средней Сибири [12], прироста стволовой древесины хвойных в условиях промышленного загрязнения [13] и при отсутствии антропогенного воздействия на территории заповедника “Вишерский” [14].
В своих исследованиях авторы связывают величину радиального прироста с температурой воздуха, количеством осадков, солнечной активностью, различием сроков начала ростовых процессов. Показано, что топоэкологические условия произрастания трансформируют влияние регионального климатического сигнала на изменчивость радиального прироста различных древесных пород [15, 16]. Еще в 1974 г. Т.Т. Битвинскас [1] выделял два ведущих фактора, влияющих на ширину годичных колец: изменение возраста дерева и воздействие погодно-климатических факторов; иногда к ним добавляют взаимовлияние деревьев [17].
В отдельных работах отмечается связь величины текущего годичного прироста по диаметру с приростом предшествующего года [18]. Связь радиального прироста текущего года с приростом следующего года следует рассматривать в контексте непрерывного процесса роста дерева по диаметру, что позволяет более полно учитывать факторы воздействия на его формирование.
Условия окружающей среды оказывают на годичный прирост лишь модифицирующее воздействие, более значительные нарушения ритмики роста происходят в годы с погодными аномалиями. Г.Г. Хамидуллина с соавторами считают, что температура воздуха и осадки, как отдельно взятые климатические факторы, не играют существенной роли, но их сочетание значимо влияет на радиальный прирост и определяет его динамику [19]. Результаты работ многих исследователей значительно расширяют представления о характере ростовых процессов основных лесообразующих пород. Однако влияние климатических факторов (в широтном и региональном аспектах) на характер и величину радиальных приростов P. sylvestris и P. abies остается во многом дискуссионным [20, 21].
Цель исследований заключается в выявлении особенностей роста по диаметру P. sylvestris и P. abies, произрастающих в восточной части Брянской обл., в сходных лесорастительных условиях на различных геоэлементах рельефа и оценка влияния среднемесячных температур, сумм осадков текущего и предыдущих лет и прироста предыдущего года на величину радиального прироста.
МАТЕРИАЛ И МЕТОДЫ
Негативные последствия засушливых периодов 2002 и 2010 гг., массовое поражение короедом типографом, последующие санитарные рубки сократили на 4% долю участия Рicea abies в лесах Учебно-опытного лесхоза Брянского государственного инженерно-технологического университета (УОЛ БГИТУ), при этом доля P. sylvestris увеличилась на 3%. Чистые еловые древостои распадаются, сохранившиеся смешанные разновозрастные насаждения P. sylvestris и P. abies характеризуются большей биологической устойчивостью. Изучение характера формирования годичного радиального прироста в смешанных хвойных насаждениях, сохранивших биологическую устойчивость, представляет интерес для более глубокого понимания особенностей ростовых процессов лесообразователей хвойных пород.
Исследования проведены в восточной части Брянской обл., на территории Брянского лесного массива (II и III террасы левобережья р. Десны), на двух участках с совместным произрастанием P. sylvestris и P. abies. На участке № 1 в кв. 14, в. 2 (абсолютная высота над ур. моря – 171 м; II терраса р. Десны) произрастет смешанный, спелый древостой, имеющий следующие таксационные характеристики: I ярус – 7С3Е; средняя высота – 26 м; средний диаметр – 52 см; относительная полнота – 0.3; II ярус состав – 7Е3С; полнота – 0.6; тип леса – сосняк брусничный; тип лесорастительных условий – переходные от свежих боровых к свежим суборевым; подрост – 8Е1Д1Кл, редкий; подлесок из рябины обыкновенной – редкий.
На участке № 2 в кв. 63, в. 12 (абсолютная высота над ур. моря – 199 м; III терраса р. Десны) произрастает смешанный приспевающий древостой с преобладанием сосны. Состав – 9С1Е+Б; средняя высота – 25 м; средний диаметр – 28 см; относительная полнота – 0.9; тип леса – сосняк брусничный; тип лесорастительных условий – переходные от свежих боровых к свежим суборевым; подрост – 10Е средней густоты; подлесок из рябины обыкновенной и крушины ломкой – средней густоты. Санитарно-патологическое состояние древостоев удовлетворительное, без признаков ослабления болезнями и вредителями. Микрорельеф и гидрологические режимы участков идентичны; почвы подзолистые песчаные, высокой степени дренированности, сформированные на водно-ледниковых песчаных отложениях четвертичного возраста.
На каждом из участков c 30 учетных деревьев I яруса каждой исследуемой породы (сосны и ели) возрастным буравом Пресслера Haglof (Швеция) были взяты 60 кернов древесины на высоте ствола 1.3 м для изучения динамики радиальных приростов. Учетные деревья – здоровые, хорошо развитые, без механических повреждений, обладающие средними для исследуемых древостоев значениями таксационных показателей (высоты и диаметра). В работе использованы метеоданные (среднемесячные температуры и суммы осадков) Брянского центра по гидрометеорологии и мониторингу окружающей среды – филиала ФГБУ “Центрально-Черноземного управления по гидрометеорологии и мониторингу окружающей среды”.
Обработка собранного экспериментального материала выполнялась в камеральных условиях путем сканирования зачищенных кернов древесины (разрешение 300 dpi) с последующим выполнением измерений программным дигитайзером Surfer (Golden Software, Inc.), при этом точность измерений величины приростов составляла до 0.1 мм [22]. Последующие расчеты выполнены средствами электронной таблицы Microsoft Excel. Синхронный анализ хронологий P. sylvestris и P. abies выполнен за период с середины 1950-х по 2016 гг.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Исследования на участке № 1 выявили в целом синхронную динамику радиальных приростов у P. sylvestris и P. abies (рис. 1), что подтверждает корреляционный анализ (табл. 1). Согласно значениям критериев достоверности, полученные коэффициенты корреляции значимы при р = 0.001. Аналогичные результаты показал анализ индексированных хронологий после исключения влияния возрастных трендов, что свидетельствует о синхронности изменения параметров, как сходной нормы реакции совместно произрастающих видов. Варьирование величины радиального прироста у P. sylvestris находится в диапазоне 18.9–44.4%, P. abies – 13.2–76.3%. Диапазон варьирования признака у P. abies (63.1%) более чем в 2 раза превышает соответствующий показатель у P. sylvestris (25.5%). Этот результат можно объяснить морфо-биологическими особенностями видов, и, возможно, последствиями аномально засушливых периодов.
Рис. 1.
Динамика годичных приростов (a) и индексов радиального прироста (b) P. sylvestris и P. abies на участке № 1. По горизонтали – календарные годы; по вертикали – значения радиальных приростов (мм), значения индексов радиальных приростов. Fig. 1. Dynamics of the annual radial increment (a) and radial growth indices (b) of P. sylvestris and P. abies in the Sample plot 1. X-axis – calendar years; y-axis – value of the radial increment (mm) (a), value of the radial growth indices (b).
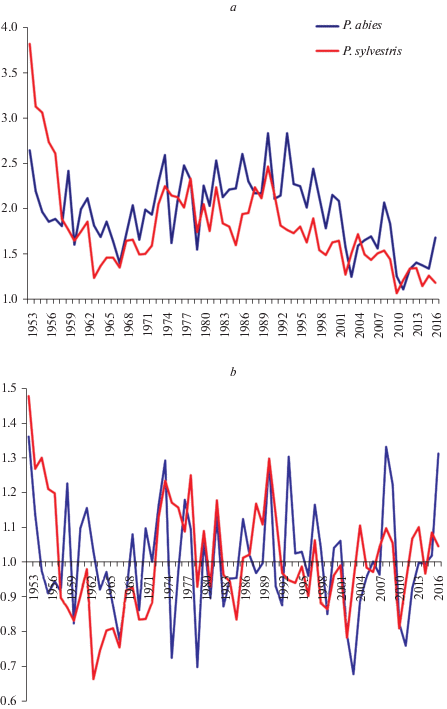
Таблица 1.
Корреляции радиальных приростов и индексов радиальных приростов P. sylvestris и P. abies на исследуемых участках Table 1. Correlation of the radial increment and radial growth indices of P. sylvestris and P. abies in the studied areas
| Номер участка Plot number |
Коэффициент корреляции (r) ± основная ошибка
коэффициента корреляции (mr) Correlation coefficient (r) ± standard error of the correlation coefficient (mr) |
Критерий достоверности (tr) Significance test (tr) |
|---|---|---|
| Абсолютные значения величины радиальных приростов, мм | ||
| Absolute values of the radial increment, mm | ||
| 1 | 0.58 ± 0.10 | 5.6 |
| 2 | 0.82 ± 0.07 | 11.2 |
| Индексы радиальных приростов | ||
| Radial growth indices | ||
| 1 | 0.43 ± 0.12 | 3.7 |
| 2 | 0.55 ± 0.11 | 5.2 |
Выполненный анализ позволил выявить годы с выраженной синхронной реакцией у хвойных пород по величине радиального прироста, причем в 1953, 1954, 1973, 1974, 1982, 1990 гг. наблюдается заметное увеличение, а в 1960, 1966, 1967, 1970, 1999, 2002 и 2010 гг. – заметное уменьшение величины радиальных приростов. Низкая синхронность роста по диаметру за 69-летний период у P. sylvestris и P. abies наблюдается лишь в 18.8% случаев, что, вероятно, связано с их индивидуальной нормой реакции на погодно-климатические условия.
На участке № 2 динамика радиальных приростов P. sylvestris и P. abies за анализируемый период также характеризуется довольно высокой синхронностью (рис. 2); варьирование величины признака у P. sylvestris наблюдается в диапазоне 19.4–52.5% (размах 33.1%), у P. abies – 19.7–60.2% (40.5%). Корреляционный анализ абсолютных значений радиальных приростов и индексированных хронологий подтверждает полученные результаты, хотя в литературе встречается информация о различной реакции P. sylvestris и P. abies на погодно-климатические условия. Например, указывается, что в зависимости от места произрастания, меняются сроки естественной спелости деревьев, ширина годичных колец и закономерности их уменьшения [1, 23, 24].
Рис. 2.
Динамика годичных приростов (a) и индексов радиального прироста (b) P. sylvestris и P. abies на участке № 2. По горизонтали – календарные годы; по вертикали – значения радиальных приростов (мм), значения индексов радиальных приростов. Fig. 2. Dynamics of the annual radial increment (a) and radial growth indices (b) of P. sylvestris and P. abies in the Sample plot 2. X-axis – calendar years; y-axis – value of the radial increment (mm) (a), value of the radial growth indices (b).
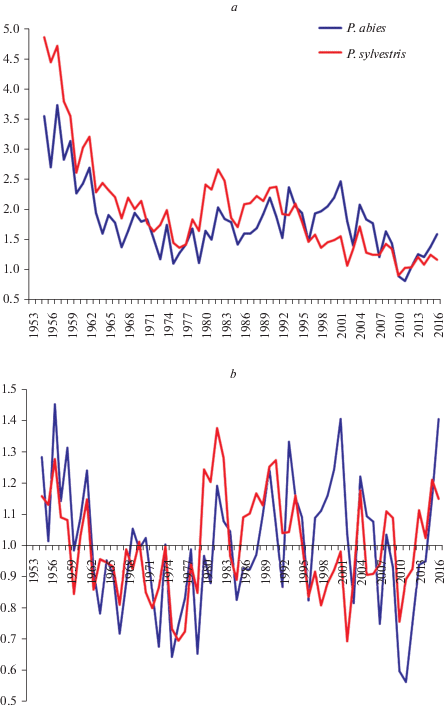
Существенное превышение величины радиальных приростов P. sylvestris по сравнению с P. abies отмечено в периоды 1955–1956, 1979–1983, 1991–1992 гг.; обратная тенденция – в периоды 1997–2002 и 2005–2006 гг., что может свидетельствовать об изменении характера конкурентных отношений между видами в древостое. Можно предположить, что это связано с их биологическими особенностями и доминированием P. sylvestris в молодом возрасте, когда при совместном произрастании она имела явное преимущество в темпах роста перед P. abies. Но с 1993 г. ситуация изменилась: средние радиальные приросты P. abies чаще оказывались выше, так как с возрастом у нее увеличивается темп роста [25].
В период аномально высокой температуры и длительного периода без осадков в метаболизме древесных пород происходят значительные изменения, которые отражаются на ростовых процессах, и, в частности, на радиальном приросте, значительная часть которого формируется за счет питательных веществ текущего года. Снижение степени оводненности цитоплазмы клеток, происходящее на фоне аномальных погодных явлений, по нашему мнению, вызывает торможение, а иногда меняет и направленность биохимичесих реакций, связанных с образованием органических веществ и их повышенный расход на транспирацию и дыхание растений.
В процессе исследований выявлены годы с выраженной синхронной реакцией у P. sylvestris и P. abies по величине радиального прироста на двух исследованных участках (на разных элементах рельефа). В 1955, 1957, 1962, 1982, 1990, 1994, 2004, 2015, 2016 гг. у обоих видов зафиксировано заметное увеличение (9 сезонов), а в 1967, 1973, 1975–1977, 1979, 1996 и 2010 гг. – заметное уменьшение средних радиальных приростов (8 сезонов). Синхронный годичный прирост P. sylvestris и P. abies наблюдается в 17 сезонах за 66-летний период, что составляет 25.7% изучаемого периода. Следовательно, можно предположить, что в четверти случаев, возможно, наблюдается значимое влияние неких факторов, оказывающих выраженное влияние на рост изучаемых видов растений.
Оценка влияния среднемесячных температур и сумм осадков текущего и предыдущего годов на величину радиального прироста показала достоверное влияние (при р < 0.05) только средних температур мая. Уменьшение и увеличение радиальных приростов у P. sylvestris наблюдалось при различии в 2.2 °С (15.1 ± 4.5 и 12.9 ± 4.3 °С); для P. abies – в 2.3 °С (15.3 ± 5.1 и 13.0 ± 4.6 °С). Следует отметить, что именно во второй половине мая обычно начинается прирост по диаметру у древесных растений, произрастающих в Брянском лесном массиве.
Анализ динамики среднегодовых температур атмосферного воздуха за последние 50–55 лет по данным Брянской метеостанции показал достоверное их повышение (примерно на 2.5 °С), что делает погодно-климатические условия менее комфортными для произрастания P. abies, южная граница ареала которой проходит по территории Брянской обл. Хотя в некоторых работах говорится, что “…дальнейшее потепление климата после 1998 г. явилось благоприятным фактором для сосны обыкновенной и ели европейской” [26]. Подобный вывод неоднозначен, так как современные ареалы видов сформировались в условиях бореального климата, и постепенное потепление неизбежно приведет к ослаблению роста и изменению положения южных границ их ареалов.
Анализ совместного влияния сумм осадков и среднемесячных температур текущего и предыдущего годов на величину радиального прироста хвойных лесообразователей показал влияние исключительно среднемесячных температур мая текущего года, хотя это расходится с взглядами Г.Г. Хамидуллиной с соавт., что только сочетание климатических факторов значимо для роста [19]. Выявление ведущей роли температурного воздействия является достаточно важным моментом в дендрохронологических исследованиях.
Результаты исследований показали, что величина радиального прироста предыдущего года (Zr – 1) у P. sylvestris и P. abies связана с аналогичным показателем текущего года (Zr). Коэффициент корреляции между абсолютными значениями радиальных приростов текущего и предыдущего годов у P. sylvestris составляет 0.90 ± 0.04 (р = 0.01). У P. abies корреляция между абсолютными значениями радиальных приростов текущего и предыдущего годов составляет 0.66 ± 0.07 (р = 0.01). Подобное явление можно объяснить расходом различного количества пластических веществ на построение годичных колец в разные годы. Этот фактор оказался наиболее значим для построения регрессионных моделей (табл. 2), с помощью которых возможно прогнозирование величины прироста текущего года еще до его окончания. Влияние радиального прироста предыдущего года и среднемесячной температуры мая (Т5) текущего года оцениваются для P. sylvestris в пределах 71–89%, P. abies – 45–63% дисперсии факторных признаков, включенных в модель прогноза величины радиального прироста текущего года. Представленные модели позволяют прогнозировать величину прироста текущего года до окончания его формирования с точностью 9–13%.
Таблица 2.
Модели множественной регрессии влияния прироста прошлого года и температуры мая текущего года на текущий радиальный прирост P. sylvestris и P. abies Table 2. The multiple regression models of the effect of the previous year radial incremnt and average May temperature of the current year on the current radial increment of P. sylvestris and P. abies
| Номер участка Plot number |
Модель Model |
Коэффициент
детерминации (R2) Determination coefficient (R2) |
Ошибка уравнения, % Error of equation, % |
|---|---|---|---|
| P. sylvestris | |||
| 1 | Zr = 1.266064 + 0.678129 × Zr – 1 – 0.053114 × T5 | 0.71 | 9.2 |
| 2 | Zr = 1.370531 + 0.808991 × Zr – 1 – 0.076313 × T5 | 0.89 | 10.8 |
| P. abies | |||
| 1 | Zr = 2.060188 + 0.491478 × Zr – 1 – 0.077810 × T5 | 0.45 | 11.6 |
| 2 | Zr = 1.845973 + 0.622550 × Zr – 1 – 0.086786 × T5 | 0.63 | 13.3 |
Древесина, как основной ресурс древесных растительных формаций, формируется за счет ежегодного накопления лигнин-целлюлозного комплекса, что находит отражение в годичных радиальных приростах. Синхронная реакция P. sylvestris и P. abies, сформировавшихся в условиях бореального климата, прослеживается по ряду показателей. Помимо близости южных границ ареалов на территории Брянской обл., наиболее значимое влияние на величину радиальных приростов, по результатам регрессионного анализа, оказывают среднемесячные температуры начала вегетационного периода, в частности, мая.
ЗАКЛЮЧЕНИЕ
Выявлена значимая корреляция (р < 0.05) величин текущего радиального прироста Pinus sylvestris L. и Picea abies (L.) Н. Karst (Pinaceae), произрастающих в идентичных типах лесорастительных условий (переходных от свежих боровых к свежим суборевым) восточной части Брянской обл. В отдельные годы исследованного периода (1953–2016 гг.) наблюдаются резко выраженные изменения величины прироста у обоих видов: его заметное уменьшение и увеличение. Синхронное изменение величин радиального прироста изученных видов подтверждается индексированными хронологиями. Изменения абсолютных значений радиальных приростов видов довольно значительны, что можно связать с воздействием климатических факторов. В результате выполненного исследования установлено, что значимое влияние на величину радиального прироста оказывает прирост предыдущего года и средняя месячная температура мая текущего года. Для P. sylvestris и P. abies влиянием этих факторов объясняется соответственно 71–89 и 45–63% общей дисперсии радиального прироста.
Глобальные изменения температурного режима атмосферного воздуха за последние десятилетия могут негативно отражаться на характере ростовых процессов P. sylvestris и P. abies, ареалы которых сформировались в условиях бореального климата, что наблюдается в условиях Брянской области, на территории, находящейся вблизи южной границы ареала ели и в южной части ареала сосны. Существенного влияния осадков на величину текущего радиального прироста изучаемых видов не выявлено, но влияние данного фактора требует дальнейших исследований.
Список литературы
Битвинскас Т.Т. 1974. Дендроклиматические исследования. Л. 172 с.
Николаева С.А., Савчук Д.А., Бочаров А.Ю. 2015. Влияние различных факторов на прирост деревьев Pinus sibirica в высокогорных лесах Центрального Алтая. – Журн. Сибирского федерального университета. Биология. 8(3): 299–318. https://doi.org/10.17516/1997-1389-2015-8-3-299-318
Charney N., Babst F., Poulter B., Record S., Trouet V., Enquist B., Evans M. 2016. Observed forest sensitivity to climate implies large changes in 21st century North American forest growth. – Ecology Letters. 19(9): 1119–1128. https://doi.org/10.1111/ele.12650
Матвеев С.М. 2014. Цикличность в динамике радиального прироста естественных и искусственных сосновых древостоев в борах центральной лесостепи. – Лесной вестник / Forestry Bulletin. 5: 110–115. https://www.elibrary.ru/item.asp?id=22399808
Foster J.R., Finley A.O., D’Amato A.W., Bradford J.B., Banerjee S. 2016. Predicting tree biomass growth in the temperate-boreal ecotone: Is tree size, age, competition, or climate response most important? – Global Change Biology. 22(6): 2138–2151. https://doi.org/10.1111/gcb.13208
Rollinson C.R., Kaye M.W., Canham C.D. 2016. Interspecific variation in growth responses to climate and competition of five eastern tree species. – Ecology. 97(4): 1003–1011. https://doi.org/10.1890/15-1549.1
Болботунов А.А., Дегтярева Е.В. 2019. Особенности формирования сезонной древесины хвойных пород деревьев в Беларуси. – Материалы II междунар. науч.-практ. конф. “Сохранение лесных экосистем: проблемы и пути их решения”. Киров. С. 27–32.
Кирдянов А.В. 2017 Радиальный прирост хвойных в лесотундре и северной тайге Средней Сибири. Роль факторов внешней среды. Автореф. дис. … д-ра биол. наук. Красноярск. 38 с. http://elib.sfu-kras.ru/handle/2311/68401
Бабушкина Е.А., Белокопытова Л.В., Костякова Т.В., Кокова В.И. 2018. Характеристики ранней и поздней древесины Pinus sylvestris в семиаридных природных зонах Южной Сибири. – Экология. 3: 174–183. https://doi.org/10.7868/S0367059718030022
Фахрутдинова В.В., Бенькова В.Е., Шашкин А.В. 2017. Влияние климатических факторов на клеточное строение годичных колец лиственницы Гмелина в контрастных гидротермических условиях лесотундры. – Известия Российской академии наук. Серия биологическая. 6: 665–674. https://doi.org/10.7868/S0002332917060108
Пинаевская Е.А. 2018. Влияние климатических параметров на формирование радиального прироста сосны на северной границе ареала Европейского Севера России. – Вестник КрасГАУ. 2: 208–214. http://www.kgau.ru/vestnik/2018_2/content/31.pdf
Фонти М.В., Фахрутдинова В.В., Калинина Е.В., Тычков И.И., Попкова М.И., Шишов В.В., Николаев А.Н. 2018. Многолетняя изменчивость анатомических параметров годичных колец хвойных пород в криолитозоне Средней Сибири. – Лесоведение. 6: 403–416. https://doi.org/10.1134/S0024114818050030
Уразгильдин Р.В., Полякова Г.Р., Аминева К.З., Галиахметов Р.Д., Кулагин А.Ю. 2018. Прирост стволовой древесины сосны обыкновенной, ели сибирской и лиственницы Сукачева в условиях промышленного загрязнения. – Бюллетень Московского общества испытателей природы. Отдел биологический. 123(2): 45–61. https://www.elibrary.ru/item.asp?id=32866522
Андреев Д.Н., Хотяновская Ю.В. 2016. Анализ изменения радиального прироста ели сибирской (Picea obovata) и пихты сибирской (Abies sibirica) на территории заповедника “Вишерский”. – Известия Самарского научного центра Российской академии наук. 18(2): 30–34. http://www.ssc.smr.ru/media/journals/izvestia/2016/2016_2_30_34.pdf
Демина А.В., Костякова Т.В., Кропачева Ю.А. 2016. Различия влияния температуры воздуха и почвы на ширину годичных колец разных видов хвойных, произрастающих в лесостепной зоне юга Сибири. – Инновационная наука. 3–4: 43–46. https://www.elibrary.ru/item.asp?id=25684242
Поршнева У.В. 2017. Сравнительный анализ динамики атмосферных осадков и солнечной активности с радиальным приростом можжевельника обыкновенного. – В сб.: Папанинские чтения. Архангельск. С. 352–356. https://narfu.ru/university/library/books/3229.pdf
Рогозин М.В., Разин Г.С. 2015. Развитие древостоев. Модели, законы, гипотезы. Пермь. 277 с.
Демаков Ю.П., Андреев Н.В. 2014. Закономерности радиального прироста деревьев сосны в приозерных биотопах национального парка “Марий Чодра”. – Экопотенциал. 3(7): 48–58. https://www.elibrary.ru/item.asp?id=22006156
Хамидуллина Г.Г., Исхаков Ф.Ф., Кулагин А.А., Зайцев Г.А., Давыдычев А.Н. 2014. Зависимость радиального прироста сосны обыкновенной (Pinus sylvestris L.) от топоэкологических условий произрастания. – Известия Оренбургского государственного аграрного университета. 1(45): 12–14. https://orensau.ru/ru/prochiedokumenty/doc_download/3139—45
Ерегина С.В. 2016. Влияние геофизических факторов среды на радиальный прирост хвойных. – Вестник научных конференций. 6–2(10): 31–35. https://ukonf.com/doc/cn.2016.06.02.pdf
Хуторной О.В., Жук Е.А., Бочаров А.Ю. 2018. Радиальный рост евроазиатских видов 5-хвойных сосен в клоновом архиве на юге Томской области. – Журн. сибирского федерального университета. 11(3): 260–274. https://doi.org/10.17516/1997-1389-0070
Марченко С.И. 2008. Техника выполнения измерительных работ с использованием компьютера: учеб. пособие. Брянск. 20 с.
Искандиров П.Ю., Тишин Д.В., Чижикова Н.А. 2019. Дендроклиматический потенциал сосны обыкновенной Приволжской вовышенности Ульяновской области. – Известия Самарского науч. центра РАН. 21(2): 127–130. http://www.ssc.smr.ru/media/journals/izvestia/2019/2019_2_127_130.pdf
Калинина Е.В., Кнорре А.А., Фонти М.В., Ваганов Е.А. 2019. Сезонное формирование годичных колец лиственницы сибирской и сосны обыкновенной в зоне Южной тайги Средней Сибири. – Экология. 3: 182–188. https://doi.org/10.1134/S0367059719030065
Тарасенко В.П., Родин С.А. 2007. Ель обыкновенная. М. 216 с.
Киселев В.Н., Матюшевская Е.В., Яротов А.Е., Митрахович П.А. 2016. Дендроклиматические исследования для понимания экологических проблем в лесном хозяйстве Беларуси. – Природопользование. 29: 50–56. http://nature-nas.by/resources/journals/default/PRIRODA_29_2016.pdf
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы