

Растительные ресурсы, 2021, T. 57, № 1, стр. 15-38
Фитомасса лишайников и зеленых растений в сообществах восточноевропейских тундр
С. А. Уваров 1, *, А. М. Лапина 2, О. В. Лавриненко 2, 3
1 Баренц-отделение Всемирного фонда дикой природы (WWF) России
г. Нарьян-Мар, Россия
2 Ботанический институт им. В.Л. Комарова РАН
г. Санкт-Петербург, Россия
3 Государственный природный заповедник “Ненецкий”
г. Нарьян-Мар, Россия
* E-mail: SUvarov@wwf.ru
Поступила в редакцию 10.04.2020
После доработки 11.09.2020
Принята к публикации 10.12.2020
Аннотация
В 10 районах восточноевропейских тундр выполнен отбор фитомассы растений в сообществах синтаксонов 4 основных классов растительности (Carici rupestris–Kobresietea bellardii, Carici arctisibiricae–Hylocomietea alaskanii cl. prov., Loiseleurio procumbentis–Vaccinietea, Oxycocco–Sphagnetea), распространенных в автоморфных условиях на водоразделах. В фитоценозах, отнесенных к 7 ассоциациям (включая 4 субассоциации) и 4 типам сообществ, общая надземная фитомасса изменялась от 130 до 1823 г/м2, при среднем значении – 794 г/м2. Наибольший вклад в ее запасы в ненарушенных сообществах вносили лишайники, в нарушенных выпасом оленей (на о-ве Колгуев) – мхи. На широтном градиенте с севера на юг этот показатель увеличивался, в основном, за счет лишайников.
Исследования кормовых запасов фитомассы особенно важны для тундр европейского Севера, почти вся территория которых (более 70%) является оленьими пастбищами. В 30–70-х гг. прошлого столетия это хорошо понимали руководители государственных органов Советской власти, по заказу которых были проведены геоботанические экспедиции с целью обследования оленьих пастбищ в разных районах восточноевропейских тундр. Непосредственное участие в них принимали крупные ученые-геоботаники – В.Д. Александрова (Новая Земля), В.Н. Андреев (Большеземельская тундра, п-ов Канин), И.Д. Богдановская-Гиенэф и З.Н. Смирнова (о-в Колгуев), К.Н. Игошина (Приуралье), Ф.В. Самбук и А.А. Дедов (Тиманская, Малоземельская и Большеземельская тундры). В.Н. Андреев [1, 2] разработал методику учета и картирования кормовых запасов фитомассы, которая впоследствии активно применялась. Расчеты оленеемкости геоботанических контуров и пастбищ разных сезонов выпаса, как основного критерия оценки качества пастбищ, основаны на реальных, измеряемых непосредственно в полевых условиях, значениях кормового валового запаса фитомассы трав, листьев кустарников и кустарничков (в том числе зимнезеленых) и лишайников в разных сообществах.
В настоящее время изменились подходы и к описанию и классификации растительности, и к картографированию, которое проводится с использованием современных технологий дистанционного зондирования и полевого дешифрирования спутниковых снимков. Однако основой для расчета оленеемкости по-прежнему являются подготовка точной карты растительности и получение достоверных данных о кормовом запасе фитомассы в разных сообществах.
В данной статье для восточноевропейских тундр впервые приведены актуальные данные о структуре надземной фитомассы в сообществах синтаксонов, описанных в традициях школы Браун-Бланке.
МАТЕРИАЛ И МЕТОДЫ
Геоботанические работы и отбор фитомассы в сообществах были проведены в 10 районах восточноевропейских тундр, расположенных в подзонах арктических, типичных и южных тундр и в северной лесотундре [3, 4] (рис. 1, 1–10).
Рис. 1.
Картосхема районов исследований. 1–10 – номера районов: 1 – бассейн р. Песчанки, о-в Колгуев; 2 – губа Долгая, о-в Вайгач; 3 – бухта Лямчина, о-в Вайгач; 4 – бухта Варнек, о-в Вайгач; 5 – долина р. Большая Светлая, Тиманский кряж; 6 – оз. Голодная Губа, Малоземельская тундра; 7 – устье р. Юшиной, Большеземельская тундра; 8 – бассейн р. Арвисью, Большеземельская тундра; 9 – бассейн р. Северная, Большеземельская тундра; 10 – бассейн р. Шапкина, Большеземельская тундра. Пунктирной линией обозначены границы подзон и полос: I – арктические тундры, II – типичные тундры, III – южные тундры, IV – северная лесотундра. Fig. 1. Map of study areas. 1–10 – Site numbers: 1 – Peschanka River basin, Kolguev Island; 2 – Dolgaya Bay, Vaygach Island; 3 – Lyamchina Bay, Vaygach Island; 4 – Varnek Bay, Vaygach Island; 5 – Bolshaya Svetlaya River basin, Timanskiy ridge; 6 – Golodnaya Guba Lake, Malozemelskaya tundra; 7 – Yushina River mouth, Bolshezemelskaya tundra; 8 – Arvisyu River basin, Bolshezemelskaya tundra; 9 – Severnaya River basin, Bolshezemelskaya tundra; 10 – Shapkina River basin, Bolshezemelskaya tundra. Boundary of subzones labeled by dotted line: I – arctic tundra, II – typical tundra, III – southern tundra, IV – northern forest-tundra.
Район 1 находится в центральной части о‑ва Колгуев, в бассейне р. Песчанки (69°15′–69°18′ с.ш., 48°58′ в.д.). Здесь в пределах наиболее высокой террасы 80–10011 расположены ландшафты ледниковой и водно-ледниковой аккумулятивной волнистой и мелкохолмисто-увалистой равнины. Отдельные холмы, сложенные с поверхности суглинками с галькой и валунами, достигают абсолютных отметок 140–150 м.
Районы 2–4 расположены на о-ве Вайгач: 2 – в окрестностях губы Долгая на севере острова (70°14′–70°16′ с.ш., 58°44′–58°49′ в.д.), 3 – в окрестностях бухты Лямчина вдоль западного берега острова (69°50′–69°54′ с.ш., 59°22′–59°40′ в.д.) и 4 – в окрестностях бухты Варнек на юге острова (69°45′ с. ш., 60°00′ в. д.). Вытянутые в северо-западном направлении гряды сложены палеозойскими глинистыми сланцами, глинистыми и песчанистыми известняками; на севере острова они высокие (до 157 м), на юге более низкие (100–130 м) и постепенно террасами понижаются к краям и окаймлены узкой полосой прибрежной равнины [5].
Район 5 находится в пределах северной части Тиманского кряжа (хребет Чайцынский камень) в долине р. Большая Светлая (67°09′–67°13′ с.ш., 49°02′–49°07′ в.д.). Вершины гряд, сложенных древними протерозойскими и палеозойскими песчаниками, известняком, доломитами, кварцитами, сланцами, интрузиями базальтов, хорошо выровнены и имеют высоту 240–280 м (максимальная – 310 м), глубина эрозионного расчленения – до 120 м [6].
Остальные районы расположены на востоке Малоземельской тундры (район 6) и на западе Большеземельской тундры (районы 7–10), которые разделены р. Печорой. Для этой равнинной территории, наклоненной в сторону моря, характерно чередование аккумулятивного, почти плоского рельефа ледникового, ледниково-морского и морского происхождения. На прилегающих к долине р. Печоры участках выражены аллювиальные и морские террасы до 60 м высотой, рельеф здесь формировался в результате деятельности рек и моря. Вдоль береговой линии моря развиты низменные сильно заболоченные и заозеренные террасы. К югу поверхность поднимается уступами нескольких древних террас, сложенных среднеплейстоценовыми ледниковыми и ледниково-морскими отложениями, представленными суглинками и глинами с частыми включениями валунно-галечного материала и на значительной части перекрытыми торфом небольшой (менее 0.5 м) толщины. Изолированные холмы и вытянутые в цепочки ледниковые гряды имеют средние высоты 140–180 м и разделены многочисленными долинами, болотами, озерными котловинами остаточно-ледникового и термокарстового происхождения. Максимальная высотная отметка в Малоземельской тундре – 171, в Большеземельской – 242 м; глубина эрозионного расчленения – 10–70 м [6]. Район 6 примыкает к северо-западному побережью оз. Голодная Губа (67°57′–67°58′ с.ш., 52°49′–52°50′ в.д.), 7 – находится на правобережье в нижнем течении р. Печоры (в месте впадения р. Юшиной) (68°07′–68°11′ с.ш., 54°11′–54°15′ в.д.), 8 – в бассейне р. Арвисью (правый приток р. Неруты) (67°48′ с.ш., 54°58′ в.д.), 9 – в бассейне р. Северная (67°37′–67°38′ с.ш., 54°01′–54°07′ в.д.), 10 – в бассейне р. Шапкина (67°31′–67°35′ с.ш., 55°01′–56°02′ в.д.).
Полевые работы проведены С.А. Уваровым в 2011–2014 гг. Геоботанические описания (для синтаксономического анализа использовано 175) выполняли на площадках 4 × 4 м, на которых описывали микро- и нанорельеф, горизонтальную и вертикальную структуру растительного покрова, выявляли все виды сосудистых растений и основные виды мохообразных и лишайников, оценивали их проективное покрытие (%) и обилие-встречаемость по шкале Браун-Бланке: r – единично; + – менее 1%; 1 – 1–5%; 2а – 6–12%; 2b – 13–25%; 3 – 26–50%; 4 – 51–75%; 5 – 76–100% [7]. Геоботанические описания из базы данных, сформированной в программе TURBOVEG [8], экспортировали в Juice [9], где выполняли автоматическую обработку таблиц. Последующую работу с таблицами проводили в программе Excel. Для каждого вида в каждом синтаксоне рассчитывали константность по шкале (%): I – >0–20, II – 21–40, III – 41–60, IV – 61–80, V – 81–100.
Определение принадлежности выделенных синтаксонов к уже описанным в системе эколого-флористической классификации выполнено при их табличном сравнении на основе сходства видового состава и дифференцирующих комбинаций видов, характерных видов высших единиц и с учетом других характеристик: структуры сообществ, занимаемых ими местообитаний, включая почвенные условия. При небольшом числе сделанных описаний (4–5) синтаксоны оставлены в ранге типа сообществ (com. type), как и ценозы, сильно нарушенные в результате чрезмерного выпаса оленей.
Учет надземной фитомассы выполнен на 74 геоботанических площадках по методике В.Д. Александровой и В.Ф. Шамурина [10]. Отбор проб проводили на площадках размером 1 × 1 м, которые закладывали внутри геоботанической площадки таким образом, чтобы на нее попадали все элементы микро- и нанорельефа, выраженные в данном фитоценозе. Собирали отдельно листья кустарников, кустарнички (зимнезеленые и листопадные), травянистые растения (разнотравье, злаки, осоковидные и хвощи), лишайники (кустистые и листоватые) и мхи (включая крупный печеночник Ptilidium ciliare). Отбирали те части растений, которые возвышаются над моховой дерниной и находятся в живом слое мхов, а также верхние части моховой дернины (живые части мхов). Разобранные образцы взвешивали во влажном состоянии в полевых условиях при помощи электронных весов “Tefal Easy” с ошибкой 0.1 г. В последующем в лабораторных условиях образцы высушивали до воздушно-сухого состояния и взвешивали при помощи электронных весов “AND ek-120i” с ошибкой 0.01 г.
Номенклатура таксонов сосудистых растений дана по Н.А. Секретаревой [11], мхов – по сводке O.M. Afonina, I.V. Czernyadjeva [12], печеночников – по Н.А. Константиновой с соавт. [13], лишайников – по R. Santesson et al. [14].
Номенклатура высших синтаксонов приведена в соответствии с “Vegetation of Europe…” [15]. Характерные виды высших синтаксонов, включая региональные, даны по работе O.V. Lavrinenko, I.A. Lavrinenko [16]. Названия ассоциаций и синтаксонов более низкого ранга соответствуют “International code of phytosociological nomenclature” (ICPN) [17].
Синтаксономический раздел занимает большую часть работы, поскольку это соответствует ее цели – оценить запасы фитомассы в сообществах разных синтаксонов эколого-флористической классификации.
Характеристика синтаксонов
Сообщества, в которых отобрана фитомасса, отнесены к 7 ассоциациям (включая 4 субассоциации) и трем типам сообществ из 4 классов растительности. Принадлежность еще одного типа сообществ высшим единицам не установлена. Большинство синтаксонов ранее были описаны на территории восточноевропейских тундр [18–21].
Продромус синтаксонов:
Класс Carici rupestris–Kobresietea bellardii Ohba 1974
Порядок Thymo arcticae–Kobresietalia bellardii Ohba 1974
Союз Kobresio–Dryadion Nordhagen 1943
Асс. Vulpicido tilesii–Dryadetum octopetalae Lavrinenko, Matveyeva et Lavrinenko 2014
Класс Carici arctisibiricae–Hylocomietea alaskanii Matveyeva et Lavrinenko 2016 cl. prov.
Порядок ?
Союз Dryado octopetalae–Caricion arctisibiricae Koroleva et Kulyugina in Chytrý et al. 2015 22
Асс. Dryado octopetalae–Hylocomietum splendentis Andreyev ex Lavrinenko et Lavrinenko 2018
Субасс. caricetosum redowskianae Lavrinenko et Lavrinenko 2018
Асс. Carici arctisibiricae–Hylocomietum splendentis Andreyev ex Lavrinenko et Lavrinenko 2018
Асс. Calamagrostio lapponicae–Hylocomietum splendentis Lavrinenko et Lavrinenko 2018
Класс Loiseleurio procumbentis–Vaccinietea Eggler ex Schubert 1960
Порядок Deschampsio flexuosae–Vaccinietalia myrtilli Dahl 1957
Союз Loiseleurio–Arctostaphylion Kalliola ex Nordhagen 1943
Асс. Loiseleurio–Diapensietum (Fries 1913) Nordhagen 1943
Субасс. typicum
Асс. Empetro–Betuletum nanae Nordhagen 1943
Тип сообщества Dicranum elongatum–Salix nummularia com. type
Тип сообщества Hierochloё alpina–Empetrum hermaphroditum com. type
Тип сообщества Stereocaulon rivulorum com. type
Класс Oxycocco–Sphagnetea Br.-Bl. et Tx. ex Westhoff et al. 1946
Порядок Sphagnetalia medii Kästner et Flössner 1933
Союз Rubo chamaemori–Dicranion elongati Lavrinenko et Lavrinenko 2015
Асс. Rubo chamaemori–Dicranetum elongati Dedov ex Lavrinenko et Lavrinenko 2015
Субасс. inops (Bogdanovskaya-Giyenef 1938) Lavrinenko et Lavrinenko 2015
Субасс. caricetosum rariflorae Lavrinenko et Lavrinenko 2015
Класс ?
Порядок ?
Союз ?
Тип сообщества Bistorta major–Salix myrsinites com. type
В классе Carici rupestris–Kobresietea bellardii описаны сообщества, отнесенные к одной ассоциации – асс. Vulpicido tilesii–Dryadetum octopetalae (табл. 1, синтаксон 1), занимающие слабо выпуклые плато на вершинах гряд и пологие участки по краям морских террас с карбонатными щебнисто-суглинистыми грунтами на о-ве Вайгач (районы 2–4). Общее проективное покрытие растительности – 35–50%. Горизонтальная структура покрова регулярно-циклическая или неявно сетчатая – слабовыпуклые или плоские щебнисто-суглинистые пятна обрамлены шпалерами кустарничка Dryas octopetala (10–40 см шириной и 3–5 см толщиной), в которых много осоки Carex rupestris, лишайников Flavocetraria nivalis и Thamnolia vermicularis и мхов Hypnum bambergeri, Syntrichia ruralis и Tomentypnum nitens. На пятнах образует подушки Silene acaulis. Дифференцирующий вид ассоциации – лишайник Vulpicida tilesii, малообильный, но хорошо заметный из-за яркой желтой окраски.
Таблица 1.
Сводная таблица синтаксонов основных классов растительности, в которых отобрана фитомасса Table 1. Synoptic table of basic classes of vegetation from which phytomass was collected
| Класс Class |
Carici–Kobresietea | Carici arctisibiricae–Hylocomietea alaskani cl. prov. | Loiseleurio procumbentis–Vaccinietea | Oxycocco–Sphagnetea | ? | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Синтаксон Syntaxon |
Ass. Vulpicido tilesii–Dryadetum octopetalae | Ass. Dryado octopetalae–Hylocomietum splendentis subass. caricetosum redowskianae | Ass. Carici arctisibiricae–Hylocomietum splendentis | Ass. Calamagrostio lapponicae–Hylocomietum splendentis | Ass. Loiseleurio–Diapensietum subass. typicum | Dicranum elongatum–Salix nummularia com. type | Ass. Empetro–Betuletum nanae | Hierochloё alpina–Empetrum hermaphroditum com. type | Stereocaulon rivulorum com. type | Rubo chamaemori–Dicranetum elongati subass. inops | Ass. Rubo chamaemori–Dicranetum elongati subass. caricetosum rariflorae | Bistorta major–Salix myrsinites com. type |
| Проективное покрытие Projective cover, %: | ||||||||||||
| Общее Total | 35–50 | 55–95 | 90–95 | 95–100 | 85–100 | 85–90 | 90–100 | 95–100 | 70–85 | 95–100 | 90–100 | 100 |
| Кустарники Shrubs | 1 | 1 | 1–5 | 1–15 | 1–20 | 1 | 1–30 | 0 | 0 | 0 | 1–10 | 10–20 |
| Кустарнички Dwarf shrubs | 25 | 20–35 | 15–25 | 20–35 | 3–15 | 25–25 | 5–40 | 25–30 | 15–25 | 10–20 | 20–40 | 20–30 |
| Травы Herbs | 5–10 | 5–15 | 5–10 | 1–10 | 1–5 | 3–5 | 1–5 | 1–5 | 5–10 | 5–15 | 1–10 | 5–10 |
| Мохообразны Mosses | 1–5 | 5–20 | 25–35 | 15–40 | 3–15 | 20–30 | 5–40 | 1–5 | 5–10 | 45–60 | 10–40 | 40–60 |
| Лишайники Lichens | 5–10 | 20–60 | 25–40 | 25–50 | 30–75 | 30–40 | 40–60 | 60–70 | 50–55 | 20–25 | 30–60 | 30–40 |
| Число описаний Number of relevés | 4 | 5 | 5 | 11 | 33 | 24 | 35 | 13 | 4 | 28 | 10 | 3 |
| Номер синтаксона Syntaxon number | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| Дифференцирующий вид асс. Vulpicido tilesii–Dryadetum octopetalae | ||||||||||||
| Differential species of ass. Vulpicido tilesii–Dryadetum octopetalae | ||||||||||||
| Vulpicida tilesii K-D | 4r,+ | . | . | . | . | . | . | . | . | . | . | . |
| Характерные виды союза Kobresio–Dryadion, пор. Thymo arcticae–Kobresietalia bellardii и кл. Carici rupestris–Kobresietea bellardii | ||||||||||||
| Characteristic species of Kobresio–Dryadion alliance, Thymo arcticae–Kobresietalia bellardii order and Carici rupestris–Kobresietea bellardii class | ||||||||||||
| Carex rupestris C-K | 41,2a | . | . | . | . | . | . | . | . | . | . | . |
| Androsace chamaejasme subsp. arctisibirica | 4+ | . | . | . | . | . | . | . | . | . | . | . |
| Carex fuligunosa subsp. misandraC-K | 3r,+ | . | . | . | . | . | . | . | . | . | . | . |
| Saxifraga oppositifolia s. str. | 3+ | . | . | . | . | . | . | . | . | . | . | . |
| Hedysarum hedysaroides subsp. arcticum | 2+ | I+ | . | . | . | . | . | . | . | . | . | . |
| Региональные характерные виды союза Kobresio–Dryadion | ||||||||||||
| Regional characteristic species of Kobresio–Dryadion alliance | ||||||||||||
| Hypnum bambergeri | 4+,1 | I+ | . | . | . | . | . | . | . | . | . | . |
| Syntrichia ruralis | 31 | . | . | . | . | . | . | . | . | . | . | . |
| Характерные виды союза Kobresio–Dryadion и класса Carici rupestris–Kobresietea bellardii, константные в зональных синтаксонах в восточноевропейских тундрах | ||||||||||||
| Characteristic species of Kobresio–Dryadion alliance and Carici rupestris–Kobresietea bellardii class, constant in zonal syntaxa in the East European tundra | ||||||||||||
| Dryas octopetala s. l. C-K | 4/2b,3 | V2b,3 | . | . | . | . | . | . | . | . | . | . |
| Pedicularis oederi C-K* | 4r,+ | V+ | IVr,+ | I+ | . | . | . | . | . | . | . | . |
| Silene acaulis C-K | 41 | V1 | I+ | . | . | . | . | . | . | . | . | . |
| Bistorta vivipara | 4+ | V+,1 | . | . | I+ | . | . | . | . | . | . | 2+,1 |
| Cladonia pocillum C-K* | . | IVr,+ | . | . | . | . | . | . | . | . | . | . |
| Дифференцирующая комбинация видов асс. Dryado octopetalae–Hylocomietum splendentis | ||||||||||||
| Differential combination of species of ass. Dryado octopetalae–Hylocomietum splendentis | ||||||||||||
| Salix reticulata K-D | 2+,1 | V+–2a | . | I+ | . | . | . | . | . | . | . | 1+ |
| Equisetum scirpoides | 1+ | V+ | III+,1 | I+ | . | . | . | . | . | . | . | 3+,1 |
| Дифференцирующая комбинация видов субасс. D. o.–H. s. caricetosum redowskianae | ||||||||||||
| Differential combination of species of subass. D. o.–H. s. caricetosum redowskianae | ||||||||||||
| Carex parallela subsp. redowskiana | . | V+ | . | . | . | . | . | . | . | . | . | . |
| Astragalus umbellatus | . | Vr–1 | . | . | . | . | . | . | . | . | . | . |
| Myxobilimbia lobulata | 3+,1 | V+,1 | . | . | . | . | . | . | . | . | . | . |
| Festuca vivipara | 3+,1 | IVr,+ | . | . | . | . | . | . | . | . | . | . |
| Дифференцирующая комбинация видов асс. Calamagrostio lapponicae–Hylocomietum splendentis | ||||||||||||
| Differential combination of species of ass. Calamagrostio lapponicae–Hylocomietum splendentis | ||||||||||||
| Calamagrostis lapponica | . | . | . | V+ | . | . | . | I1 | . | . | . | . |
| Nephroma arcticum | . | . | Ir | V1 | Ir | Ir–1 | I+ | . | . | . | I+,1 | . |
| Salix phylicifolia | . | . | . | IVr | . | . | . | . | . | . | I+ | . |
| Характерные виды высших единиц кл. Carici arctisibiricae–Hylocomietea alaskani cl. prov. | ||||||||||||
| Characteristic species of high syntaxa of Carici arctisibiricae–Hylocomietea alaskani class prov. | ||||||||||||
| Carex bigelowii subsp. arctisibirica | . | V+,1 | IV+,1 | IV+–2a | I+,1 | IV+,1 | II1 | III+,1 | . | . | . | . |
| Hylocomium splendens | . | V+,1 | V1,2a | V+–2b | I+–2a | Ir-1 | III+–2a | . | . | . | II1–2b | 31–2b |
| Aulacomnium turgidum | 11 | III+,1 | V1 | V+–2a | . | I+ | I+ | . | . | . | . | 31,2a |
| Salix polaris | 11 | V1–3 | V1,2a | . | . | I1 | . | . | . | . | . | . |
| Luzula arcuata | . | IV1 | V1 | Ir,+ | . | . | . | . | . | . | . | . |
| Tomentypnum nitens | 2+,1 | IV+–2a | . | II1–2b | . | . | Ir | . | . | . | . | . |
| Racomitrium lanuginosum | . | III1,2a | I+ | II+ | . | . | I1–2b | . | . | . | . | . |
| Deschampsia glauca | . | II1 | V+,1 | . | . | . | . | . | . | . | . | . |
| Petasites frigidus | . | . | IV1 | . | . | I+ | . | . | . | . | . | . |
| Salix glauca s. str. | . | . | I1 | V+–2a | I1 | . | IIr–1 | . | . | . | I+ | 21,2a |
| Pedicularis lapponica | . | . | . | III+,1 | I+ | . | . | . | . | . | . | . |
| Дифференцирующий вид асс. Loiseleurio–Diapensietum и субасс. L.-D. typicum | ||||||||||||
| Differential species of ass. Loiseleurio–Diapensietum and subass. L.-D. typicum | ||||||||||||
| Loiseleuria procumbens L-A | . | . | . | . | V+–2a | . | . | . | . | . | . | . |
| Дифференцирующий вид Dicranum elongatum–Salix nummularia com. type | ||||||||||||
| Differential species of Dicranum elongatum–Salix nummularia com. type | ||||||||||||
| Salix nummularia | 21 | . | . | . | I+,1 | V1–2b | I1 | . | 41 | . | . | . |
| Дифференцирующая комбинация видов асс. Empetro–Betuletum nanae | ||||||||||||
| Differential combination of species of ass. Empetro–Betuletum nanae | ||||||||||||
| Betula nana L-V* | . | . | . | V1–2b | V1–2b | Ir | V1–3 | . | . | . | IV1,2a | 31 |
| Ptilidium ciliare | . | . | . | . | I2a | IV–2a | V+–2b | V1 | . | . | V1–2b | 32a,2b |
| Stereocaulon paschale L-V* | . | . | . | . | III+–2b | IIr–1 | V+–3 | III+–2b | . | . | . | . |
| Дифференцирующий вид Hierochloё alpina–Empetrum hermaphroditum com. type | ||||||||||||
| Differential species of Hierochloё alpina–Empetrum hermaphroditum com. type | ||||||||||||
| Hierochloё alpina L-V | . | . | . | . | . | . | . | IV+,1 | . | . | . | . |
| Дифференцирующая комбинация видов Stereocaulon rivulorum com. type | ||||||||||||
| Differential combination of species of Stereocaulon rivulorum com. type | ||||||||||||
| Stereocaulon rivulorum | . | . | . | . | . | . | . | . | 44 | . | . | . |
| Polemonium boreale | . | . | . | . | . | . | . | . | 4r–1 | . | . | . |
| Solidago lapponica | . | . | . | I+ | I1 | I | Ir,+ | . | 3+,1 | . | . | . |
| Solorina crocea | . | . | Ir | . | I1 | . | Ir,+ | . | 3+,1 | . | . | . |
| Характерные виды союза Loiseleurio–Arctostaphylion, пор. Rhododendro ferruginei–Vaccinietalia и кл. Loiseleurio procumbentis–Vaccinietea | ||||||||||||
| Characteristic species of Loiseleurio–Arctostaphylion alliance, Rhododendro ferruginei–Vaccinietalia order and Loiseleurio procumbentis–Vaccinietea class | ||||||||||||
| Arctous alpina L-V | . | . | III1 | V+–2a | IV1,2a | IV1,2a | V1–3 | . | 42a | . | . | 31,2a |
| Alectoria ochroleuca L-V* | 1r | II+ | . | I+ | III+,1 | I1 | IV+–2a | IV1,2a | . | . | I+ | 1+ |
| Juncus trifidus | . | . | . | . | III+,1 | I+ | I+,1 | I1 | 11 | . | . | . |
| Arctostaphylos uva-ursi | . | . | . | . | I2a | . | . | . | 11 | . | . | . |
| Региональные характерные виды союза Loiseleurio–Arctostaphylion | ||||||||||||
| Regional characteristic species of Loiseleurio–Arctostaphylion alliance | ||||||||||||
| Polytrichum piliferum | . | . | . | I1 | IIIr–1 | I1 | III+,1 | IVr–2a | 41,2a | . | . | 1+ |
| Cetraria nigricans | . | . | . | . | II+,1 | . | II+,1 | I1 | . | . | . | . |
| Характерные виды союза Rubo chamaemori–Dicranion elongati и дифференцирующая комбинация видов асс. Rubo chamaemori–Dicranetum elongati | ||||||||||||
| Characteristic species of Rubo chamaemori–Dicranion elongati alliance and differential combination of species of ass. Rubo chamaemori–Dicranetum elongati | ||||||||||||
| Dicranum elongatum | . | I1 | IV2a–3 | II1,2a | I+,1 | V1–3 | I1,2a | II1 | . | V3–5 | V1–3 | 32a,2b |
| Rubus chamaemorus O-S | . | . | III1 | I+,1 | . | . | I1 | . | . | V2b,3 | V+–2b | . |
| Cladonia stellaris | . | . | IIr–1 | II+,1 | IIIr–1 | IIr,+ | III+,1 | II+,1 | . | IIr–1 | IVr–1 | 2r,+ |
| Дифференцирующий вид субасс. R. ch.–D. e. caricetosum rariflorae | ||||||||||||
| Differential species of subass. R. ch.–D. e. caricetosum rariflorae | ||||||||||||
| Carex rariflora | . | . | . | . | . | . | II1 | . | . | . | V1,2a | . |
| Характерные виды кл. Oxycocco-Sphagnetea, пор. Sphagnetalia medii | ||||||||||||
| Characteristic species of Oxycocco-Sphagnetea class, Sphagnetalia medii order | ||||||||||||
| Polytrichum strictum O-S | . | . | IV1–2b | II+–2a | I1 | . | Ir | I1 | . | V2a | III+–2a | . |
| Eriophorum vaginatum O-S | . | . | I1 | . | . | . | . | I1 | . | IV+–2a | III+–2a | . |
| Ledum palustre subsp. decumbensO-S | . | . | . | II1,2a | IV+–2a | . | V1–2b | III1,2a | . | . | V1–2b | 1+ |
| Andromeda polifolia subsp. pumilaO-S | . | . | . | I2a | I1–2b | . | I+ | I1 | . | . | III1,2a | . |
| Sphagnum balticum O-S | . | . | . | . | . | . | . | . | . | I+,1 | IV1–2b | . |
| S. russowii O-S* | . | . | . | . | . | . | . | . | . | I+,1 | II+,1 | . |
| Дифференцирующая комбинация видов Bartsia alpina–Salix myrsinites com. typ | ||||||||||||
| Differential combination of species of Bartsia alpina–Salix myrsinites com. type | ||||||||||||
| Salix myrsinites | . | . | . | . | . | . | . | . | . | . | . | 31–2b |
| Bartsia alpina | . | . | . | . | . | . | . | . | . | . | . | 3+,1 |
| Bistorta major | . | II+ | . | IIr–1 | I+,1 | I+ | I+,1 | II1 | . | . | . | 31 |
| Carex vaginata subsp. quasivaginata | . | . | . | . | . | . | . | . | . | . | . | 3+,1 |
| Equisetum pratense | . | . | . | IIIr–1 | . | . | . | II+,1 | . | . | . | 2+,1 |
| Pedicularis labradorica | . | . | . | Ir | . | . | I1 | . | . | . | . | 21 |
| Гипоарктические кустарнички, обычные в сообществах классов Loiseleurio procumbentis–Vaccinietea, Carici arctisibiricae–Hylocomietea alaskani и Oxycocco–Sphagnetea в южных и типичных тундрах | ||||||||||||
| Hypoarctic dwarf-shrubs, common in communities of Loiseleurio procumbentis–Vaccinietea, Carici arctisibiricae–Hylocomietea alaskani and Oxycocco–Sphagnetea classes in southern and typical tundra | ||||||||||||
| Vaccinium vitis-idaea subsp. minusL-V | . | III1,2a | V1,2a | V1,2a | IV1 | V1–2b | V1,2a | V1–2b | . | V1–2b | V1–2b | 3+–2a |
| Empetrum hermaphroditum L-V | . | . | V1–2b | V1–3 | V1–2b | IV+–2b | V1–2b | V2a–3 | 41–2b | V2a–3 | V1–2b | 32a,2b |
| Vaccinium uliginosum subsp. microphyllumL-V | . | . | II+–2a | V+–2a | III+,1 | I+ | V+–2a | IV1,2a | . | I1,2a | V+–2b | 31,2a |
| Константные виды синтаксонов | ||||||||||||
| Constant species | ||||||||||||
| Flavocetraria nivalis L-A, L-V* | 41,2a | V2a,2b | V1,2a | V+–2b | V1–3 | V1,2a | V1–3 | V2b–4 | 3+ | V1,2a | V+–3 | 3+,1 |
| F. cucullata L-V* | 4+ | V+,1 | V1 | V+,1 | V+–2a | V+,1 | V+–2a | V1 | 2+,1 | I1 | V+,1 | 3+,1 |
| Thamnolia vermicularis L-V* | 4+,1 | V+,1 | III+,1 | V+,1 | III+,1 | IV+,1 | V+,1 | V1,2a | . | IV+,1 | IV+,1 | 2+,1 |
| Cetraria islandica subsp. crispiformis | 4+,1 | V+,1 | V1 | IV+–2a | V1–2b | V+,1 | V+,1 | V1 | . | V1 | V+,1 | 31 |
| Sphaerophorus globosus L-V* | 1+ | V+,1 | IV1 | IV+–2b | IV+–2b | V1,2a | V+–2a | V+,1 | . | V1 | II1 | 21 |
| Alectoria nigricans | 1+ | Vr–1 | IV+,1 | III+,1 | IV1 | IVr–1 | V+–2b | V1,2a | 2+,1 | V+,1 | III+,1 | 3+,1 |
| Bryocaulon divergens | 1+ | Vr–1 | III1 | III1 | II+,1 | V+–2a | V+–2a | V1 | 41 | IV+,1 | III+,1 | 1+ |
| Cladonia arbuscula s. l. | . | V+–2b | V2a,2b | V1–3 | V1–2b | V1–2b | V1–2b | V1–3 | 1+ | V2b | V2a–3 | 32b,3 |
| C. amaurocraea | . | IVr–1 | V1 | V+,1 | I+ | Vr–1 | V+,1 | IV+ | . | V+,1 | V+,1 | 3+,1 |
| C. gracilis subsp. elongata | . | IV+,1 | V+,1 | V+,1 | III+,1 | V+,1 | V+,1 | V+,1 | . | V+,1 | V+,1 | 31 |
| Cetrariella delisei | 1+ | III+,1 | III1 | I+ | I/3 | III+–2a | I+ | III+,1 | . | I+,1 | III+,1 | . |
| Ochrolechia frigida L-V | . | . | IV+,1 | I+ | IIr–1 | IV+,1 | III+,1 | IIIr–1 | . | V+,1 | II+,1 | 3r,+ |
| Cladonia rangiferina | . | . | V2a | V+–3 | V1–2b | IIIr–2a | V+–2b | V1 | . | V+–2a | V1–2b | 32a |
| C. uncialis | . | . | . | III+ | III+,1 | I+ | II+,1 | IIIr | . | Ir,+ | IIIr–1 | 3+,1 |
| Polytrichastrum alpinum | 1+ | V+–2a | . | I+ | II+–2a | II+–2a | II1–2b | I+ | . | . | II+,1 | . |
| Polytrichum hyperboreum | . | I+ | . | IV+–2a | II1,2a | V1–2b | IV+–2b | IIIr–2a | 12a | II+–2a | . | 1+ |
| Pleurozium schreberi | . | . | IV+,1 | II+,1 | I+–2a | IIr–1 | I+ | I+ | . | . | I+ | 2+ |
| Festuca ovina | . | . | V1 | IV+,1 | III1 | IIIr–1 | IVr–1 | I1 | 31 | . | . | 2+ |
| Luzula confusa | . | . | V1 | . | Ir–1 | V+,1 | III+,1 | IV+,1 | . | . | I+ | 31 |
| Salix herbacea | . | . | V2a,2b | . | . | V1–2b | . | . | . | . | . | . |
| Stereocaulon alpinum | 11 | II+ | IIIr–1 | I+ | . | . | II+,1 | I1 | . | . | . | . |
| Sphenolobus minutus | . | . | III+ | IV+ | . | II+ | II+ | . | . | I+,1 | IV+ | 2+ |
Примечание. Виды с константность I–II, относящиеся к “Прочим”, в таблицу не включены. Сокращения: O-S – вид приведен в сводке L. Mucina et al. (2016) как диагностический класса Oxycocco–Sphagnetea, L-V – класса Loiseleurio procumbentis–Vaccinietea, C-K – класса Carici rupestris–Kobresietea bellardii; звездочка рядом с сокращением означает, что в этой сводке вид приведен не как характерный для одного класса, а имеет общую диагностическую ценность. L-A – характерный вид союза Loiseleurio–Arctostaphylion, K-D – союза Kobresio–Dryadion. Note. Species with a constancy I or II related to “Other” are omitted. Abbreviations: O-S – species is given in the paper by L. Mucina et al. (2016) as a diagnostic of Oxycocco–Sphagnetea class, L-V – Loiseleurio procumbentis–Vaccinietea class, C-K –Carici rupestris–Kobresietea bellardii class; an asterisk next to the abbreviation denote that in this paper the species is not shown as being characteristic of one class, but has general diagnostic value. L-A – characteristic species of Loiseleurio–Arctostaphylion alliance, K-D – Kobresio–Dryadion alliance.
Сообщества трех ассоциаций класса Carici arctisibiricae–Hylocomietea alaskanii cl. prov., предложенного для зональной тундровой растительности [22], занимают плакоры – слабонаклонные (1°–3°) суглинистые поверхности водоразделов. Почвы представлены глееземами.
Ценозы асс. Dryado octopetalae–Hylocomietum splendentis, зональной для типичных тундр, описаны на террасах 10–80 м высотой в северной и юго-западной частях о-ва Вайгач (районы 2, 3), где представлены субасс. caricetosum redowskianae (табл. 1, синтаксон 2). Общее проективное покрытие растительности варьирует от 55 до 95%. Горизонтальная структура регулярно-циклическая – кустарничково-мохово-лишайниковая дернина прерывается округлыми или вытянутыми пятнами суглинка, частично со щебнем на поверхности. В покрове содоминируют кустарнички Dryas octopetala, Salix polaris, лишайники Flavocetraria nivalis и реже Cladonia arbuscula и мхи Aulacomnium turgidum, Hylocomium splendens, Tomentypnum nitens, заметна осока Carex bigelowii subsp. arctisibirica. С невысоким обилием, но постоянно встречаются дифференцирующие виды ассоциации – Equisetum scirpoides, Salix reticulata и субассоциации – Astragalus umbellatus, Carex parallela subsp. redowskiana и Festuca vivipara.
В подзоне типичных тундр на о-ве Колгуев плакорные местообитания в верховьях р. Песчанки (район 1) заняты сообществами второй зональной ассоциации – асс. Carici arctisibiricae–Hylocomietum splendentis (табл. 1, синтаксон 3). Общее проективное покрытие растительности – 90–95%. Горизонтальная структура гетерогенная, обусловлена чередованием приподнятых округлых или овальных (5–15 см выстой, 0.5–2.0 м в поперечнике) участков, на которых имеются небольшие пятна выдавленного суглинка, и окружающих их ровных поверхностей. Сообщества олигодоминантные. В травяно-кустарничковом ярусе преобладают Empetrum hermaphroditum, Salix herbaceae, S. polaris, Vaccinium vitis-idaea subsp. minus, Carex bigelowii subsp. arctisibirica, Deschampsia glauca, Luzula arcuata. В напочвенном покрове содоминируют мхи – Aulacomnium turgidum, Dicranum elongatum, D. majus, Hylocomium splendens, Polytrichum hyperboreum, на повышенных элементах рельефа преобладают лишайники – Cladonia arbuscula, C. rangiferina, Flavocetraria nivalis.
Сообщества асс. Calamagrostio lapponicae–Hylocomietum splendentis (табл. 1, синтаксон 4), зональной для южных тундр, описаны в Тиманской, Малоземельской и Большеземельской тундрах (районы 5–7, 10). Общее проективное покрытие растительности – 95–100%. Дифференцирующие виды ассоциации – Calamagrostis lapponica, Nephroma arcticum и Salix phylicifolia. Сообщества отличает разреженный (до 30%) кустарниковый ярус из Betula nana, Salix glauca и S. phylicifolia. В травяно-кустарничковом ярусе активны гипоарктические кустарнички – Arctous alpina, Empetrum hermaphroditum, Vaccinium uliginosum subsp. microphyllum, V. vitis-idaea subsp. minus, реже Ledum palustre subsp. decumbens, и травянистые растения – Calamagrostis lapponica, Festuca rubra subsp. arctica, Pedicularis lapponica. Напочвенный покров лишайниково-моховой, многовидовой, в нем наиболее заметны Aulacomnium turgidum, Dicranum laevidens, Hylocomium splendens, Polytrichum hyperboreum, Ptilidium ciliare, Cladonia arbuscula, Flavocetraria nivalis, Stereocaulon paschale.
Ценозы двух ассоциаций (одна из которых представлена субассоциацией) и трех синтаксонов ранга типа сообществ из класса Loiseleurio procumbentis–Vaccinietea распространены на выположенных, хорошо дренированных, сухих летом и слабо укрытых снегом зимой поверхностях водоразделов с песчаными субстратами. Почвы представлены подбурами и псаммоземами.
Кустарничково-лишайниковые сообщества асс. Loiseleurio–Diapensietum субасс. typicum (табл. 1, синтаксон 5) описаны в Тиманской, Малоземельской и Большеземельской тундрах (районы 5, 6, 9). Общее проективное покрытие растительности – 85–100%. Есть очень разреженный (от 1 до 20% покрытия) кустарниковый ярус из Betula nana, иногда вместе с Ledum palustre subsp. decumbens. Бывают обильны кустарнички, образующие куртины, – Arctous alpina, Empetrum hermaphroditum и Loiseleuria procumbens, что обусловливает мозаичную горизонтальную структуру сообщества. В напочвенном покрове доминируют лишайники – Cetraria islandica subsp. crispiformis, Cladonia arbuscula, C. rangiferina, Flavocetraria cucullata, F. nivalis, Stereocaulon paschale. Мхов немного, в лишайниковый покров вкраплены в основном дерновинки и побеги политрихумов (Polytrichum hyperboreum и P. piliferum). Травы редки, чаще других встречаются Juncus trifidus и Festuca ovina.
При чрезмерной пастбищной нагрузке в кустарничково-лишайниковых тундрах не остается неповрежденных талломов кормовых лишайников, их подеции выдернуты из дернины, низко скусаны, обломаны или имеют вид лишайниковой крошки. Такие сильно измененные выпасом оленей сообщества описаны в ранге типа сообществ Dicranum elongatum–Salix nummularia com. type [21]. Мы отнесли к нему ценозы (табл. 1, синтаксон 6), описанные на о-ве Колгуев и в окрестностях оз. Голодная Губа (районы 1, 6). Общее проективное покрытие растительности – 85–90%. Сильно выбитые сообщества от других синтаксонов класса Loiseleurio procumbentis–Vaccinietea отличает постоянство кустарничков (Salix herbacea и S. nummularia), мхов (наиболее обильны Dicranum elongatum, Polytrichum hyperboreum, Ptilidium ciliare) и некоторых трав (Carex bigelowii subsp. arctisibirica, Luzula confusa). Горизонтальная структура мозаичная, кустарнички Arctous alpina, Empetrum hermaphroditum, Salix herbacea и S. nummularia разрастаются куртинами.
Сообщества асс. Empetro–Betuletum nanae (табл. 1, синтаксон 7) обычны на хорошо дренированных песчаных водораздельных поверхностях в Тиманской, Малоземельской и Большеземельской тундрах (районы 5, 6, 9, 10). Дифференцирующая комбинация видов ассоциации: Betula nana, Ledum palustre subsp. decumbens, Ptilidium ciliare и Stereocaulon paschale. Общее проективное покрытие растительности – 90–100%. Ерник растет кустами около 1 м в диам. и 25–30 (до 40) см выс. Обилие кустарничков (Arctous alpina, Empetrum hermaphroditum и Vaccinium uliginosum subsp. microphyllum, V. vitis-idaea subsp. minus) бывает высоким (до 40%). Напочвенный покров мозаичный: под кустами ерника – преимущественно мхи Hylocomium splendens и Ptilidium ciliare, на открытых пространствах – лишайники Alectoria nigricans, Bryocaulon divergens, Cladonia arbuscula, Flavocetraria nivalis и Stereocaulon paschale.
Кустарничково-лишайниковые тундры с доминированием Empetrum hermaphroditum, Vaccinium vitis-idaea subsp. minus, Cladonia arbuscula, Flavocetraria nivalis и с участием злака Hierochloё alpina описаны в ранге типа сообществ Hierochloё alpina–Empetrum hermaphroditum com. type (табл. 1, синтаксон 8). Они занимают пологие или плоские хорошо-дренированные участки на водоразделах в Малоземельской и Большеземельской тундрах (районы 6 и 9). Из характерных видов класса, кроме Hierochloё alpina, в них отмечены только Alectoria ochroleuca и Stereocaulon paschale. С высокой константностью присутствует Polytrichum piliferum – региональный характерный вид союза Loiseleurio–Arctostaphylion. Общее проективное покрытие растительности – 95–100%. Горизонтальная структура мозаичная, напочвенный покров сложен в основном светлоокрашенными лишайниками Cladonia arbuscula и Flavocetraria nivalis с вкраплениями подушек темноокрашенных Alectoria nigricans и Bryocaulon divergens и побегов кустарничков.
Пионерные сообщества на зарастающих песчаных обнажениях с доминированием лишайника Stereocaulon rivulorum описаны в Большеземельской тундре (район 9) в ранге типа сообществ Stereocaulon rivulorum com. type (табл. 1, синтаксон 9). Общее проективное покрытие растительности – 70–85%. Стереокаулон покрывает поверхность почвы тонким (3–4 см выс.) светло-серым ковром, который прерывается куртинами кустарничков Arctous alpina и Empetrum hermaphroditum и дерновинками Polytrichum piliferum. Встречаются редкие травы (Polemonium boreale, Solidago lapponica) и накипной лишайник Solorina crocea.
К классу Oxycocco–Sphagnetea, объединяющему в тундровой зоне растительность торфяных бугров плоскобугристо-топяных и полигонально-трещиноватых комплексных болот, отнесены сообщества асс. Rubo chamaemori–Dicranetum elongati, представленной двумя субассоциациями – обедненной на о-ве Колгуев и субасс. caricetosum rariflorae в материковых тундрах.
Ценозы субасс. R. ch.–D. e. caricetosum rariflorae (табл. 1, синтаксон 11) описаны в Малоземельской и Большеземельской тундрах (районы 6, 8). Дифференцирующий вид субассоциации – Carex rariflora. Общее проективное покрытие растительности – 90–100%. Поверхность пологобугорковая. Есть разреженный ярус Betula nana, 10–15 см выс. Травяно-кустарничковый ярус сформирован Rubus chamaemorus, Andromeda polifolia subsp. pumila, Empetrum hermaphroditum, Ledum palustre subsp. decumbens, Vaccinium uliginosum subsp. microphyllum, V. vitis-idaea subsp. minus. В мохово-лишайниковом покрове доминируют Dicranum elongatum, Ptilidium ciliare, кустистые кладонии (Cladonia arbuscula, C. rangiferina) и Flavocetraria nivalis, часто, но не обильно встречаются Cladonia stellaris и по понижениям – Cetrariella delisei и Sphagnum balticum.
Сообщества субасс. R. ch.–D. e. inops (табл. 1, синтаксон 10), описанные на торфяниках в северной половине о-ва Колгуев (район 1), отличает отсутствие кустарника Betula nana, кустарничков Andromeda polifolia subsp. pumila и Ledum palustre subsp. decumbens (характерных видов класса Oxycocco–Sphagnetea), низкая константность Vaccinium uliginosum subsp. microphyllum. Общее проективное покрытие растительности – 95–100%. Постоянны и бывают обильны Empetrum hermaphroditum, Vaccinium vitis-idaea subsp. minus, Rubus chamaemorus, встречаются кочки Eriophorum vaginatum. В напочвенном покрове над лишайниками преобладают мхи – Dicranum elongatum и Polytrichum strictum, что говорит о сильной нарушенности сообществ под влиянием нерегламентированного выпаса оленей.
Ценозы, оставленные в ранге типа сообществ Bistorta major–Salix myrsinites com. type (табл. 1, синтаксон 12), описаны на востоке Малоземельской тундры в окрестностях оз. Голодная Губа (район 6), где занимают нижние части склонов сопок, хорошо укрытые снегом зимой. Это кустарничково-лишайниково-моховые сообщества с пологобугорковой поверхностью и разреженным (10–20%) ярусом кустарников Betula nana, Salix glauca и S. myrsinites. Дифференцирующая комбинация видов синтаксона включает травянистые растения – Bartsia alpina, Bistorta major, Carex vaginata subsp. quasivaginata, Equisetum pratense, Pedicularis labradorica и иву Salix myrsinites. Общее проективное покрытие растительности – 100%. Кустарничково-лишайниково-моховая дернина мозаичная, в ней наиболее обильны Empetrum hermaphroditum, Aulacomnium turgidum, Dicranum elongatum, Hylocomium splendens, Ptilidium ciliare, Cladonia arbuscula.
Запасы фитомассы
По всем образцам фитомассы в воздушно-сухом состоянии, отобранным в сообществах разных синтаксонов восточноевропейских тундр, минимальные значения составили 130.2 г/м2, максимальные – 1822.7 г/м2, при среднем значении – 794.0 г/м2 (табл. 2). Пропорция “зеленые сосудистые растения : мохообразные : лишайники” (средняя по всем пробам) равна 1.0 : 1.5 : 4.4, т.е. наибольший вклад в запасы фитомассы вносят лишайники. Максимальные значения на 1 м2 площади для лишайников достигали 1324.0 г, для мхов – 1220.3 г, для зеленых частей сосудистых растений – 500.0 г.
Таблица 2.
Значения воздушно-сухой фитомассы растений разных жизненных форм (г/м2) в сообществах разных синтаксонов восточноевропейских тундр Table 2. Air-dry plant phytomass in plants of different life forms (g/m2) in the syntaxon communities of the East European tundra
| Авторский номер описания Relevé number by the author |
Номер района Site number |
Кустарники Shrubs |
Кустарнички Dwarf-shrubs |
Травы Herbs |
Зеленая фитомасса Green phytomass |
Мхи Mosses |
Лишайники Lichens |
Общая фитомасса Total phytomass |
|---|---|---|---|---|---|---|---|---|
| Класс Carici rupestris–Kobresietea bellardii | ||||||||
| Асс. Vulpicido tilesii–Dryadetum octopetalae | ||||||||
| 8 ЛЯМ | 3 | – | 11.6 | 56.1 | 67.7 | 7.5 | 26.6 | 191.9 |
| 9 ЛЯМ | 3 | – | 146.5 | 5.9 | 152.4 | 4.0 | 82.0 | 283.4 |
| Среднее | – | 79.1 ± 67.4 | 31.0 ± 25.1 | 110.1 ± 42.3 | 5.8 ± 1.8 | 54.3 ± 27.7 | 237.6 ± 45.8 | |
| Класс Carici arctisibiricae–Hylocomietea alaskani | ||||||||
| Асс. Dryado octopetalae–Hylocomieteum splendentis субасс. caricetosum redowskianae | ||||||||
| 20ДОЛ | 2 | – | 91.9 | 36.6 | 128.5 | 33.5 | 182.3 | 344.3 |
| 22ДОЛ | 2 | – | 248.1 | 13.2 | 261.3 | 400.8 | 717.6 | 1379.8 |
| Среднее | – | 170.0 ± 78.1 | 24.9 ± 11.7 | 194.9 ± 66.4 | 217.2 ± 183.6 | 450.0 ± 267.7 | 862.0 ± 517.7 | |
| Асс. Carici arctisibiricae–Hylocomietum splendentis | ||||||||
| 66КОЛ | 1 | 0.1 | 45.6 | 5.8 | 51.5 | 535.1 | 147.7 | 779.3 |
| 74КОЛ | 1 | 8.2 | 78.5 | 17.1 | 103.8 | 335.0 | 101.4 | 540.2 |
| Среднее | 2.0 ± 4.1 | 62.0 ± 16.5 | 11.4 ± 5.7 | 77.6 ± 26.2 | 435.0 ± 100 | 124.5 ± 23.2 | 659.7 ± 119.5 | |
| Асс. Calamagrostio lapponicae–Hylocomieteum splendentis | ||||||||
| 1ШАП* | 10 | 29.6 | 0.5 | 9.3 | 39.4 | 106.2 | 1301.1 | 1446.6 |
| 1ШАП* | 10 | 12.5 | 75.2 | 11.1 | 98.8 | 177.9 | 1292.4 | 1569.1 |
| 13ШАП* | 10 | – | 189.3 | 5.5 | 194.8 | 87.7 | 582.0 | 864.5 |
| 13ШАП* | 10 | 9.2 | 159.3 | 3.3 | 171.8 | 57.4 | 521.5 | 750.6 |
| 14ШАП | 10 | 47.8 | 17.4 | 0.5 | 65.7 | 264.2 | 414.5 | 833.2 |
| 10ШАП | 10 | 1.7 | 15.5 | 10.0 | 27.2 | 117.5 | 170.2 | 450.0 |
| 12ШАП | 10 | 14.2 | 6.2 | 37.5 | 57.9 | 848.7 | 845.2 | 1751.8 |
| Среднее | 19.2 ± 7.5 | 66.2 ± 32.0 | 11.0 ± 5.0 | 93.7 ± 26.8 | 237.1 ± 113.6 | 732.4 ± 177.5 | 1095.1 ± 199.7 | |
| Класс Loiseleurio–Vaccinietea | ||||||||
| Асс. Loiseleurio–Diapensietum субасс. typicum | ||||||||
| 1СЕВ | 9 | 4.5 | 71.9 | 0.4 | 76.8 | 2.1 | 125.2 | 204.1 |
| 18СЕВ | 9 | 0.7 | 118.7 | – | 119.4 | – | 471.9 | 591.3 |
| 14СЕВ | 9 | – | 172.6 | 0.6 | 173.2 | – | 1187.5 | 1360.7 |
| 15СЕВ | 9 | 12.5 | 149.2 | 0.3 | 162.0 | – | 713.6 | 875.6 |
| 7СЕВ | 9 | 63.1 | 2.8 | 3.2 | 69.1 | 311.0 | 615.6 | 995.8 |
| 9СЕВ | 9 | – | 67.4 | – | 67.4 | – | 641.7 | 709.1 |
| 21СЕВ | 9 | 6.6 | 21.1 | 49.0 | 76.7 | – | 815.1 | 891.1 |
| 15СЕВ | 9 | 31.2 | 4.2 | 4.0 | 39.4 | 8.1 | 902.8 | 950.3 |
| 18СЕВ | 9 | 17.2 | 144.5 | 0.1 | 161.8 | – | 214.9 | 376.8 |
| 28ТИМ | 5 | 2.0 | 39.0 | 0.5 | 41.5 | 28.0 | 334.0 | 691.5 |
| 29ТИМ | 5 | 2.0 | 25.0 | 9.0 | 36.0 | 194.0 | 930.0 | 1178.0 |
| 30ТИМ | 5 | 26.0 | 7.0 | – | 33.0 | 55.0 | 1324.0 | 1475.0 |
| 36ТИМ | 5 | 4.0 | 48.2 | 3.0 | 489.0 | 5.0 | 543.0 | 1037.0 |
| 39ТИМ | 5 | 22.0 | 83.0 | – | 105.0 | 42.0 | 900.0 | 1047.0 |
| Среднее | 16.0 ± 5.2 | 69.7 ± 17,3 | 7.0 ± 4.7 | 117.9 ± 31.4 | 80.7 ± 39.6 | 694.2 ± 92.6 | 884.5 ± 93.8 | |
| Dicranum elongatum–Salix nummularia com. type | ||||||||
| 22КОЛ | 1 | – | 34.2 | 1.1 | 35.3 | 22.7 | 89.7 | 147.6 |
| 25КОЛ | 1 | – | 19.3 | 8.5 | 27.8 | 40.8 | 65.0 | 133.6 |
| 29КОЛ | 1 | – | 13.6 | 2.9 | 16.5 | 16.9 | 107.8 | 141.1 |
| 41КОЛ | 1 | – | 6.1 | 1.2 | 7.3 | 54.4 | 42.3 | 167.0 |
| 64КОЛ | 1 | – | 37.4 | 14.0 | 51.4 | 68.1 | 106.7 | 225.8 |
| 78КОЛ | 1 | – | 37.5 | 7.0 | 44.5 | 516.9 | 116.7 | 678.0 |
| 24ГОЛ | 6 | – | 171.1 | 16.8 | 187.9 | 43.8 | 621.2 | 852.9 |
| Среднее | – | 45.6 ± 21.4 | 7.4 ± 2.4 | 53.0 ± 23.2 | 109.0 ± 68.3 | 164.2 ± 76.8 | 335.1 ± 113.3 | |
| Асс. Empetro–Betuletum nanae | ||||||||
| 8СЕВ | 9 | 1.9 | 114.3 | 1.9 | 118.1 | 57.0 | 759.7 | 934.9 |
| 7СЕВ | 9 | 24.4 | 1.7 | 3.6 | 29.7 | 75.0 | 617.6 | 821.3 |
| 3ШАП | 10 | 11.3 | 67.5 | 0.9 | 79.7 | – | 369.3 | 449.0 |
| 11ШАП | 10 | 37.6 | 236.2 | 3.4 | 277.2 | 55.5 | 473.3 | 805.8 |
| 35 ТИМ | 5 | 22.0 | 116.0 | 0.5 | 138.5 | 7.0 | 386.0 | 531.5 |
| 61ГОЛ | 6 | – | 496.9 | 3.0 | 499.9 | 3.5 | 597.5 | 1100.0 |
| 6ШАП | 10 | – | 17.9 | 38.7 | 56.6 | 32.5 | 110.6 | 199.7 |
| 9ШАП | 10 | 4.5 | 137.5 | – | 142.0 | 103.1 | 380.8 | 625.9 |
| 8ШАП | 10 | 27.8 | 135.0 | – | 162.8 | 197.0 | 379.7 | 739.6 |
| 23ГОЛ | 6 | 29.2 | 68.6 | – | 97.8 | – | 1101.6 | 1199.5 |
| 34ГОЛ | 6 | 33.2 | 182.9 | 0.3 | 216.4 | 30.4 | 861.7 | 1108.5 |
| 60ГОЛ | 6 | – | 18.9 | 4.4 | 23.3 | 421.2 | 612.7 | 1219.2 |
| 2ШАП* | 10 | 8.8 | 256.4 | 0.3 | 265.5 | 36.4 | 527.9 | 829.8 |
| 2ШАП* | 10 | 9.7 | 18.5 | 11.4 | 39.6 | 38.8 | 724.7 | 965.1 |
| 7ШАП | 10 | 17.3 | 96.5 | 5.4 | 119.2 | 29.8 | 372.8 | 521.7 |
| Среднее | 19.0 ± 3.4 | 131.0 ± 32.9 | 6.2 ± 3.1 | 151.1 ± 32.2 | 83.6 ± 31.4 | 551.7 ± 63 | 803.4 ± 76.7 | |
| Hierochloё alpina–Empetrum hermaphroditum com. type | ||||||||
| 10СЕВ | 9 | – | 151.3 | 5.4 | 156.7 | 8.9 | 1162.9 | 1328.3 |
| 50ГОЛ | 6 | – | 137.8 | 19.6 | 157.4 | 20.7 | 681.9 | 860.0 |
| 53ГОЛ | 6 | – | 265.5 | 9.4 | 274.9 | 31.0 | 771.6 | 1077.4 |
| 28ГОЛ | 6 | – | 212.0 | 4.7 | 216.7 | 12.8 | 747.5 | 976.9 |
| 40ГОЛ | 6 | – | 14.7 | 2.9 | 17.6 | 26.1 | 637.0 | 770.7 |
| 42ГОЛ | 6 | – | 75.4 | 2.7 | 78.1 | – | 798.0 | 875.7 |
| 57ГОЛ | 6 | – | 16.9 | 7.2 | 24.1 | – | 932.4 | 1100.3 |
| Среднее | – | 124.8 ± 36 | 7.4 ± 2.2 | 132.2 ± 36.7 | 19.9 ± 4.1 | 818.7 ± 67.4 | 998.5 ± 71 | |
| Stereocaulon rivulorum com. type | ||||||||
| 3СЕВ | 9 | – | 29.9 | 3.6 | 33.5 | 0.2 | 318.9 | 352.6 |
| 14СЕВ | 9 | – | 27.7 | 3.7 | 31.4 | 7.9 | 163.3 | 219.7 |
| Среднее | – | 28.8 ± 1.1 | 3.7 ± 0.1 | 32.5 ± 1.1 | 4.0 ± 3.8 | 241.1 ± 77.8 | 286.2 ± 66.5 | |
| Класс Oxycocco–Sphagnetea | ||||||||
| Асс. Rubo chamaemori–Dicranetum elongati субасс. inops | ||||||||
| 32КОЛ | 1 | – | 26.8 | 21.5 | 48.3 | 614.5 | 197.4 | 860.1 |
| 33КОЛ | 1 | – | 47.9 | 12.4 | 60.3 | 351.0 | 204.9 | 616.2 |
| 46КОЛ | 1 | – | 66.4 | 14.0 | 80.4 | 587.7 | 455.4 | 1123.5 |
| 53КОЛ | 1 | – | 39.2 | 15.0 | 54.2 | 404.1 | 186.2 | 644.4 |
| 58КОЛ | 1 | – | 34.7 | 34.4 | 69.1 | 428.9 | 218.5 | 713.5 |
| Среднее | – | 43.0 ± 6.8 | 19.4 ± 4.1 | 62.5 ± 5.7 | 477.2 ± 52.3 | 252.5 ± 51 | 791.5 ± 93.1 | |
| Асс. Rubo chamaemori–Dicranetum elongati субасс. caricetosum rariflorae | ||||||||
| 4ШАП | 10 | 1.6 | 141.5 | 22.4 | 165.5 | 104.7 | 314.0 | 584.2 |
| 19АРВ | 8 | – | 43.6 | 17.5 | 61.1 | 457.7 | 101.8 | 620.6 |
| 15ШАП* | 10 | 4.2 | 135.2 | 16.0 | 155.4 | 83.0 | 289.9 | 528.2 |
| 15ШАП* | 10 | 6.3 | 166.8 | 14.2 | 187.3 | 235.8 | 538.3 | 961.3 |
| 36ГОЛ | 6 | – | 198.8 | 21.1 | 219.9 | – | 436.5 | 655.7 |
| 37ГОЛ | 6 | – | 15.8 | 3.4 | 19.2 | 53.2 | 377.0 | 538.7 |
| 31ГОЛ | 6 | – | 99.4 | 9.7 | 109.1 | 69.4 | 592.7 | 771.3 |
| 33ГОЛ | 6 | 5.0 | 85.7 | 15.0 | 105.7 | 27.0 | 754.6 | 887.2 |
| 55ГОЛ | 6 | 5.9 | 184.8 | 14.4 | 205.1 | 191.4 | 783.9 | 1180.2 |
| Среднее | 4.6 ± 0.8 | 119.0 ± 20.9 | 14.8 ± 1.9 | 136.5 ± 22.5 | 152.8 ± 50.2 | 465.4 ± 74.7 | 747.5 ± 74.2 | |
| Класс ? | ||||||||
| Bistorta major–Salix myrsinites com. type | ||||||||
| 45ГОЛ | 6 | 21.9 | 119.2 | 3.2 | 144.3 | 1220.3 | 458.1 | 1822.7 |
| 46ГОЛ | 6 | 44.4 | 248.7 | 6.7 | 299.8 | 799.2 | 283.3 | 1382.3 |
| Среднее Mean |
33.2 ± 11.2 | 183.9 ± 64.8 | 5.0 ± 1.7 | 222.1 ± 77.8 | 1009.7 ± 210.6 | 370.7 ± 87.4 | 1602.5 ± 220.2 | |
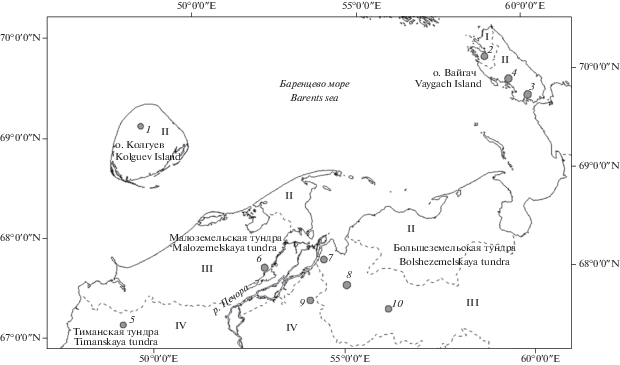
В сообществах асс. Vulpicido tilesii–Dryadetum octopetalae (класс Carici rupestris–Kobresietea bellardii) на о-ве Вайгач запасы общей фитомассы в среднем составили лишь 237.6 г/м2, около 65% из них приходилось на зеленую фитомассу (в основном листья кустарничков) и меньше всего было мхов (рис. 2а, b).
Рис. 2.
а и b. Запасы надземной фитомассы в сообществах разных классов в восточноевропейских тундрах. По вертикали: а – средние значения воздушно-сухой фитомассы (г/м2) со стандартной погрешностью, b – относительные значения воздушно-сухой фитомассы; по горизонтали – краткие названия классов растительности. Жизненные формы: штриховка – зеленые сосудистые растения, темная заливка – мхи, светлая заливка – лишайники. Fig. 2. а and b. Stock of the above-ground phytomass in the East European tundra communities of different classes. Y-axis: a – average values of air-dry phytomass (g/m2) with standard error, b – relative values of air-dry phytomass; Х-axis: – short names of vegetation classes. Life forms: hatching – green vascular plants, dark solid – mosses, light solid – lichens.
В сообществах остальных трех классов растительности значения общей фитомассы достоверно не различались. На зеленую фитомассу сосудистых растений приходилось чуть больше 10–15%, а остальное составляли споровые.
В (редкоивово)-осоково-кустарничково-лишайниково-моховых сообществах класса Carici arctisibiricae–Hylocomietea alaskani средние значения общей фитомассы были наибольшими – 903.3 г/м2, из них около 60% составляли лишайники и 30% – мхи. В сообществах двух других классов – Loiseleurio procumbentis–Vaccinietea и Oxycocco–Sphagnetea, средние значения фитомассы примерно равные – 757.3 и 780.0 г/м2, но структура запасов различается. В (ерниково)-кустарничково-лишайниковых сообществах на песчаных субстратах (класс Loiseleurio procumbentis–Vaccinietea) наибольшая доля запасов (около 75%) приходится на лишайники, а в кустарничково-морошково-мохово-лишайниковых ценозах на торфяниках (класс Oxycocco–Sphagnetea) лишайников примерно половина и около 35% мхов (рис. 2а, b).
В классе Carici arctisibiricae–Hylocomietea alaskani наименьшие значения общей фитомассы отмечены в сообществах асс. Carici arctisibiricae–Hylocomietum splendentis на о-ве Колгуев (в среднем 659.7 г/м2), в том числе 66% запасов составляли мхи и лишь около 20% – лишайники (табл. 2, рис. 3а, b). Полагаем, что это результат длительного нерегламентированного выпаса оленей. В ценозах субасс. Dryado octopetalae–Hylocomieteum splendentis caricetosum redowskianae на о-ве Вайгач и асс. Calamagrostio–Hylocomietum splendentis в южных тундрах при бóльших значениях общей средней фитомассы (862.0 и 1095.1 г/м2) на мхи приходилось немногим больше 20%, а на лишайники – 52 и 68% соответственно. Запасы зеленой фитомассы в сообществах зональных синтаксонов составляли около 10–20%.
Рис. 3.
а и b. Запасы надземной фитомассы в сообществах ассоциаций и подчиненных синтаксонов на островах и в материковых тундрах. По вертикали: а – средние значения воздушно-сухой фитомассы (г/м2) со стандартной погрешностью, b – относительные значения воздушно-сухой фитомассы; по горизонтали – названия синтаксонов. Жизненные формы: штриховка – зеленые сосудистые растения, темная заливка – мхи, светлая заливка – лишайники. Fig. 3. а and b. Stock of the above-ground phytomass in communities of associations and subordinate sytaxa on the islands and in the mainland tundra. X-axis: a – average values of air-dry phytomass (g/m2) with standart error, b – relative values of air-dry phytomass; y-axis – syntaxa names. Life forms: hatching – green vascular plants, dark solid – mosses, light solid – lichens.
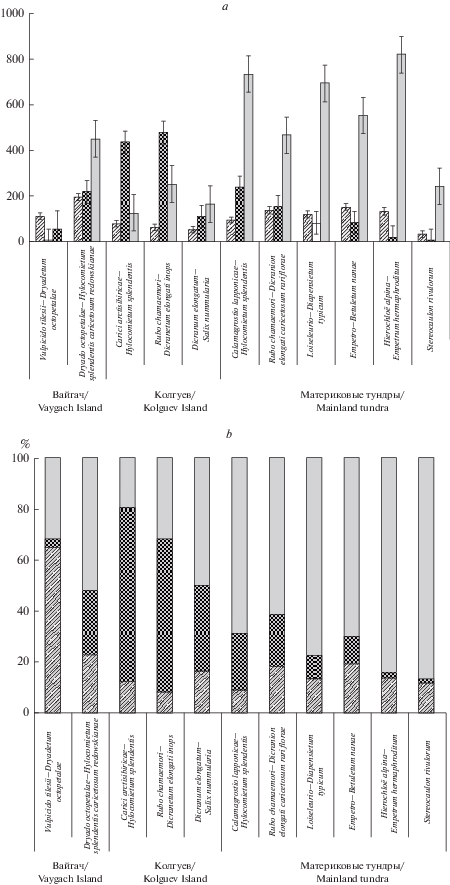
В классе Loiseleurio procumbentis–Vaccinietea наименьшая общая фитомасса (в среднем 286.2 г/м2) была в сообществах Stereocaulon rivulorum com. type (табл. 2) на зарастающих песчаных обнажениях, где на лишайники, доминирующие в покрове, приходилось 85% запасов (241.1 г/м2). Невелики запасы фитомассы и в нарушенных выпасом оленей сообществах Dicranum elongatum–Salix nummularia com. type на о-ве Колгуев – в среднем 335.1 г/м2, из них лишайники составляли около половины, но в абсолютном выражении лишь 164.2 г/м2. В кустарничково-лишайниковых сообществах субасс. Loiseleurio–Diapensietum typicum, асс. Empetro–Betuletum nanae и Hierochloё alpina–Empetrum hermaphroditum com. type в материковых тундрах общая средняя фитомасса составила 884.5, 803.4 и 998.5 г/м2 соответственно, что почти в 2.5–3.0 раза выше, чем на о-ве Колгуев, в том числе 70–85% этих запасов составляли лишайники (табл. 2, рис. 3а, b).
Влияние чрезмерного выпаса оленей на структуру запасов фитомассы хорошо видно и при сравнении двух субассоциаций асс. Rubo chamaemori–Dicranetum elongati на торфяниках (класс Oxycocco–Sphagnetea). При близких средних значениях общей фитомассы в сообществах субасс. R. ch.–D. e. caricetosum rariflorae в материковых тундрах и субасс. R. ch.–D. e. inops на о-ве Колгуев (747.5 и 791.5 г/м2), доминирующими по фитомассе в первых были лишайники (62%), а во вторых – мхи (60%).
Сомкнутые сообщества с хорошо развитым моховым покровом, отнесенные к типу Bistorta major–Salix myrsinites com. type, отличались наибольшими запасами фитомассы (в среднем 1602.5 г/м2), 63% которой (чуть больше 1000 г/м2) приходилось на бриофиты.
По запасам фитомассы прослеживаются различия на широтном градиенте. Сообщества синтаксонов, распространенных в подзоне южных тундр и северной лесотундре, по общей фитомассе превосходят синтаксоны, ареал которых находится на о-вах Колгуев и Вайгач (типичные и арктические тундры). Изменяется и вклад разных групп растений в структуру фитомассы (рис. 3а). Такие различия обусловлены не только климатическими факторами, но и влиянием на растительность нерегламентированного оленеводства, в результате которого на о-ве Колгуев к 2012–2013 гг. покров из поедаемых лишайников (кустистых кладоний и флавоцетрарий) в разных сообществах оказался сильно нарушенным [23]. При средней общей фитомассе в сообществах на о-ве Колгуев равной 545 г/м2, на лишайники приходилось 190 г/м2, или 35% (11% – зеленые сосудистые и 53% – мхи). На о-ве Вайгач средняя общая фитомасса равнялась 550 г/м2, лишайников – 252 г/м2, или 46% (28% – зеленые сосудистые и 20% – мхи). В материковых тундрах (южные тундры и лесотундра) в среднем накапливалось 886 г/м2, из них лишайники составляли 612 г/м2, или 69% (14% – зеленые сосудистые растения и 17% – мхи). Полученные значения по фитомассе лишайников укладываются в пределы, выявленные ранее для ненарушенных сообществ с доминированием кустистых кладоний и Flavocetraria nivalis в припечорских тундрах, – 490–1550 г/м2 [24]. Максимальные показатели общей фитомассы, определенные нами для сообществ асс. Empetro–Betuletum nanae в подзоне южных тундр (1100–1219 г/м2) близки к значениям для ерников в бассейне р. Усы на востоке Большеземельской тундры (в среднем 1423 г/м2) [25].
Восточноевропейские тундры по общей надземной фитомассе в сообществах сопоставимы с другими районами Арктики, но отличаются от них по структуре запасов. В малонарушенных ценозах синтаксонов зональной растительности восточноевропейских тундр (на о-ве Вайгач и в материковых тундрах) наибольший вклад в запасы фитомассы вносят лишайники (59%), тогда как в сообществах плакорных местообитаний восточного Таймыра – мхи: при общей фитомассе 594 г/м2 на них приходилось почти 60%, а на лишайники – только 8% [26]. В подзоне арктических тундр на о-ве Большой Ляховский в сообществах на плакорах общие запасы фитомассы составляли 147 г/м2, из которых около половины приходилось на долю разнотравья и кустарничков [27]. В двух образцах фитомассы, отобранных на аналогичных позициях на севере о-ва Вайгач в сообществах асс. Dryado octopetalae–Hylocomieteum splendentis субасс. caricetosum redowskianae, значения сильно варьировали (344 и 1379 г/м2), но половину в обоих составляли лишайники.
В ненарушенных болотных сообществах в южных тундрах Якутии фитомасса растений – около 730 г/м2 [28], что аналогично средним ее значениям в ценозах класса Oxycocco–Sphagnetea в материковой части восточноевропейских тундр – 747 г/м2. Запасы фитомассы в болотных ценозах Чукотки ниже: для плоскобугристого болота – 450 г/м2 и для пушицево-кочкарного – 167 г/м2 [29]. По соотношению компонентов фитомассы они отличаются от болотных сообществ восточноевропейских тундр: в фитомассе торфяников Чаунской низменности на долю споровых (представленных преимущественно мхами) приходилось около 50% от общих запасов, тогда как в сообществах класса Oxycocco–Sphagnetea восточноевропейских тундр половину запасов составляли лишайники.
В пятнистых тундрах на супесчаных грунтах и в сообществах торфяных болот в полосе лесотундры (окрестности г. Лабытнанги) накапливалось около 700 г/м2 надземной фитомассы, при том что доля лишайников в них была минимальной – 23 и 4% соответственно [30]. Л.М. Морозова с соавторами [31] приводят другие, ближе к нашим, значения для предгорных и горных районов Полярного Урала. Так, в лишайниковых тундрах при общей средней фитомассе 691 г/м2 (114–1101 г/м2) доля лишайников составляла 67% (47–92%), что сопоставимо с таковой в фитоценозах класса Loiseleurio–Vaccinietea восточноевропейских тундр – 757 г/м2 и 73% соответственно.
На Аляске, где образцы надземной фитомассы были отобраны вдоль трансекты от мыса Барроу на севере до границы леса на юге, ее значения увеличивались от 400 до 932 г/м2 в тундровых сообществах на кислых почвах и от 400 до 658 г/м2 – на не кислых. При продвижении с севера на юг продуктивность фитоценозов на кислых почвах росла за счет увеличения общей надземной фитомассы сосудистых растений (от 82 у побережья до 719 г/м2 на южном конце градиента), а на не кислых – мхов (от 100 до почти 500 г/м2). На всем широтном градиенте в структуре запасов фитомассы доля лишайников была очень низкой – около 5% (максимальные значения – около 100 г/м2) [32].
ВЫВОДЫ
1. Впервые для территории восточноевропейских тундр осуществлена привязка значений фитомассы лишайников, мхов и зеленых частей сосудистых растений к сообществам синтаксонов, установленных в традициях классификации Браун-Бланке. В ценозах четырех основных классов растительности, распространенных в автоморфных условиях на водоразделах, запасы фитомассы изменялись от 130.2 до 1822.7 г/м2, при среднем значении – 794.0 г/м2.
2. Среднее значение фитомассы было наименьшим в сообществах класса Carici rupestris–Kobresietea bellardii на о-ве Вайгач – 237.6 г/м2; в сообществах остальных классов растительности эти значения в три и более раз выше и достоверно не различались: Carici arctisibiricae–Hylocomietea alaskanii cl. prov. – 903.3 г/м2, Loiseleurio procumbentis–Vaccinietea – 757.3 г/м2 и Oxycocco–Sphagnetea – 763.2 г/м2.
3. В сообществах класса Carici rupestris–Kobresietea bellardii около 65% запасов приходилось на зеленую фитомассу (в основном листья кустарничков), в ненарушенных сообществах остальных трех классов наибольший вклад в общую фитомассу вносили лишайники (50–75%), в нарушенных выпасом оленей на о-ве Колгуев – мхи (33–66%).
4. На широтном градиенте с севера на юг значения фитомассы увеличивались, и это происходит в основном за счет лишайников.
5. Восточноевропейские тундры по общей надземной фитомассе в сообществах сопоставимы с другими районами Арктики (Таймыр, Якутия, Чукотка, Аляска), но отличаются от них значительно более высокой долей лишайников.
Синтаксономический анализ позволяет отчетливо увидеть различия в запасах фитомассы в ненарушенных и трансформированных сообществах, ее изменение на широтном градиенте в ценозах разных синтаксонов, приуроченных к разным местообитаниям. Примененный нами подход открывает возможности для создания точных карт продуктивности тундровых сообществ и оленьих пастбищ всех сезонов выпаса на основе геоботанического картографирования территории с использованием фитосоциологического метода [33, 34].
Список литературы
Андреев В.Н. 1948. Корма и пастбища северных оленей. – В сб.: Северное оленеводство. М. С. 100–157.
Андреев В.Н. 1971. Методика учета и картирования кормовых запасов фитомассы Субарктики. – Растительные ресурсы. 7(3): 439–444.
Геоботаническое районирование Нечерноземья европейской части РСФСР. 1989. Л. 64 с. 9(3):
Лавриненко И.А. 2012. Использование дистанционных методов при геоботаническом районировании восточно-европейских тундр. – Современные проблемы дистанционного зондирования Земли из космоса. 9(3): 269–276. 269–276. http://d33.infospace.ru/d33_conf/sb2012t3/269-276.pdf
Атлас Архангельской области. 1976. М. 72 с.
Ненецкий автономный округ. Современное состояние и перспективы развития. 2005. СПб. 512 с.
Barkman J.J., Doing H., Segal S. 1964. Kritische Bemerkungen und Vorschlӓge zur quantitativen Vegetationsanalyse – Acta Bot. Neerl. 13(3): 394–419. https://doi.org/10.1111/j.1438-8677.1964.tb00164.x
Hennekens S.M., Schaminée J.H.J. 2001. TURBOVEG, a comprehensive data base management system for vegetation data. – J. Veg. Sci. 12(4): 589–591. https://doi.org/10.2307/3237010
Tichý L. 2002. JUICE, software for vegetation classification. – J. Veg. Sci. 13(3): 451–453. https://doi.org/10.1111/j.1654-1103.2002.tb02069.x
Александрова В.Д., Шамурин В.Ф. 1972. Методика определения запасов и структуры фитомассы тундровых сообществ – В кн: Изучение биогеоценозов тундры и лесотундры. Л. С. 60–64.
Секретарева Н.А. 2004. Сосудистые растения Российской Арктики и сопредельных территорий. М. 131 с.
Афонина О.М., Чернядьева И.В. 1995. Мхи Российской Арктики: список видов и библиография. – Arctoa. 5: 99–142. https://doi.org/10.15298/arctoa.05.07
Константинова Н.А., Потемкин А.Д., Шляков Р.Н. 1992. Список печеночников и антоцеротовых территории бывшего СССР. – Arctoa. 1(1–2): 87–127. https://doi.org/10.15298/arctoa.01.02
Santesson R., Moberg R., Nordin A., Tønsberg T., Vitikainen O. 2004. Lichenforming and lichenicolous fungi of Fennoscandia. Museum of Evolution. Uppsala University. 359 p.
Mucina L., Bültmann H., Dierßen K., Theurillat J.-P., Raus T., Čarni A., Šumberová K., Willner W., Dengler J., Gavilán García R., Chytrý M., Hájek M., Di Pietro R., Iakushenko D., Pallas J., Daniëls F.J.A., Bergmeie, E., Santos Guerra A., Ermakov N., Valachovič M., Schaminée J.H.J., Lysenko T., Didukh Ya. P., Pignatti S., Rodwell J.S., Capelo J., Weber H.E., Solomeshch A., Dimopoulos P., Aguiar C., Freitag H., Hennekens S. M., Tichý L. 2016. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities – Applied Vegetation Science. 19(1): 1–264. https://doi.org/10.1111/avsc.12257
Лавриненко О.В., Лавриненко И.А. 2020а. Характерные виды высших синтаксонов в равнинных восточноевропейских тундрах. – Бот. журн. 105(4): 315–347. https://doi.org/10.31857/S0006813620040055
Weber H.E., Moravec J., Theurillat J.-P. 2000. International code of phytosociological nomenclature. 3rd edition. – J. Veg. Sci. 11(5): 739–768. https://doi.org/10.2307/3236580
Лавриненко О.В., Матвеева Н.В., Лавриненко И.А. 2014а. Дриадовые сообщества на востоке европейской части Российской Арктики. – Растительность России. 24: 38–63. https://doi.org/10.31111/vegrus/2014.24.38
Лавриненко О.В., Лавриненко И.А. 2015. Сообщества класса Oxycocco-Sphagnetea Br.-Bl. et R. Tx. 1943 в восточноевропейских тундрах. – Растительность России. 26: 55–84. https://doi.org/10.31111/vegrus/2015.26.55
Лавриненко О.В., Лавриненко И.А. 2018. Зональная растительность равнинных восточноевропейских тундр. – Растительность России. 32: 35–108. https://doi.org/10.31111/vegrus/2018.32.35
Лавриненко О.В., Лавриненко И.А. 2020б. Растительность класса Loiseleurio procumbentis–Vaccinietea Eggler ex Schubert 1960 в восточноевропейских тундрах. – Растительность России. 38: 27–84. https://doi.org/10.31111/vegrus/2020.38.27
Лавриненко О.В., Матвеева Н.В., Лавриненко И.А. 2016. Предварительные итоги классификации растительности восточноевропейских тундр и новый класс для зональных местообитаний. – В сб.: Разнообразие и классификация растительности. Ялта. С. 95–105. (Сборник научных трудов ГНБС. Т. 143).
Лавриненко О.В., Лавриненко И.А. 2014б. Растительный покров оленьих пастбищ острова Колгуев: преемственность исследований и современные подходы. – Труды международной научной конференции “Ботаника: история, теория, практика (к 300-летию Ботанического института им. В.Л. Комарова Российской академии наук)”. СПб. С. 124–130. https://www.binran.ru/files/publications/Proceedings/Proceedings_300-years/Proceedings_300-years_Lavrinenko.pdf
Lavrinenko I.A., Lavrinenko O.V., Sivkov M., Leummens H.J.L. 2000. Vegetation cover and productivity. – Pechora Delta: Structure and dynamics of the Pechora Delta ecosystems (1995–1999). Lelystad. P. 83–97.
Патова Е.Н., Кулюгина Е.Е., Пастухов А.В. 2015. Почвенный и растительный покров территории Усинского угольного месторождения (Большеземельская тундра) – Известия Коми научного центра УрО РАН. 3(23): 16–23. http://www.izvestia.komisc.ru/archive/i23_ann.files/patova.pdf
Поспелова Е.Б., Орлов М.В. 1987. Фитомасса растительных сообществ центральной части Восточного Таймыра и особенности ее пространственного размещения – Экология. 5: 28–37.
Николин Е.Г. 2017. К продуктивности растительных сообществ арктических тундр о-ва Большой Ляховский (Новосибирские острова, Якутия). – Наука и образование. 1: 113–121. http://no.ysn.ru/attachments/article/1857/20%20Николин%20113-121.pdf
Карпов Н.С. 2005. Надземная фитомасса и продуктивность растительности южных субарктических тундр Якутии – Природные ресурсы Арктики и Субарктики. 2: 18–23.
Пугачев А.А. 2008. Биологический круговорот в экосистемах болот тундровой зоны Чукотки – Вестник Дальневосточного отделения Российской академии наук. 4: 77–82.
Радченко Т.А., Валдайских В.В., Некрасова О.А. 2014. Биологическая продуктивность сообществ некоторых природных экосистем лесотундры Ямала. – В сб.: Современные исследования природных и социально-экономических систем. Инновационные процессы и проблемы развития естественнонаучного образования. Сборник трудов международной научно-практической конференции. Т. 2. Екатеринбург. С. 81–89. https://www.elibrary.ru/item.asp?id=23581641&pf=1
Морозова Л.М., Магомедова М.А., Эктова С.Н., Дьяченко А.П., Князев М.С., Рябицева Н.Ю., Шурова Е.А. 2006. Растительный покров и растительные ресурсы Полярного Урала. Екатеринбург. 796 с.
Walker D.A., Epstein H.E., Jia G.J., Balser A., Copass C., Edwards E.J., Gould W.A., Hollingsworth J., Knudson J., Maier H.A., Moody A., Raynolds M.K. 2003. Phytomass, LAI, and NDVI in northern Alaska: Relationships to summer warmth, soil pH, plant functional types, and extrapolation to the circumpolar Arctic – J. Geophysical Research: Atmospheres. 108(D2): 8169. https://doi.org/10.1029/2001JD000986
Лавриненко И.А. 2015. Типология территориальных единиц растительности для целей крупномасштабного картографирования (на примере острова Колгуев) – Геоботаническое картографирование. СПб. С. 95–119. https://doi.org/10.31111/geobotmap/2015.94
Лавриненко И.А. 2020. Типология и синтаксономический состав территориальных единиц растительности: новый подход на примере изучения арктических маршей – Растительность России. 39: 100–148. https://doi.org/10.31111/vegrus/2020.39.100
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы