

Растительные ресурсы, 2021, T. 57, № 1, стр. 3-14
Энергозатраты на формирование фитомассы древостоев хвойных лесов в различных эдафических и климатических условиях
Ю. В. Карпечко 1, *, А. В. Туюнен 1
1 Институт леса – обособленное подразделение Федерального государственного бюджетного учреждения науки Федерального исследовательского центра “Карельский научный центр Российской академии наук”
г. Петрозаводск, Россия
* E-mail: yuvkarp@onego.ru
Поступила в редакцию 10.04.2020
После доработки 27.07.2020
Принята к публикации 10.12.2020
Аннотация
Для 100-летних сосняков и ельников зеленомошной, долгомошной и сфагновой групп типов леса в Европейской части России (ЕЧР) получены зависимости запасов стволовой древесины от радиационного баланса лесных участков. Отмеченное повышение продуктивности древостоев с ростом радиационного баланса при продвижении от северной границы лесной зоны к южной, связано с увеличением массы фотосинтезирующего аппарата. Более высокая интенсивность возрастания продуктивности отмечена для древостоев, произрастающих в лучших лесорастительных условиях (сосняки и ельники кисличные). Затраты энергии при создании единицы фитомассы увеличиваются с ухудшением эдафических условий и зависят от широты местности. Наименьшими затратами энергии для создания единицы объема стволовой древесины 100-летних сосняков и ельников всех типов леса характеризуется территория между 61°–62° и 53°–55° с.ш. ЕЧР. Минимальные удельные энергозатраты на создание стволовой древесины 100-летних хвойных древостоев различных типов леса могут использоваться в качестве количественных критериев при классификации лесов.
Леса являются крупнейшей экосистемой суши на планете, они играют ведущую роль не только в снабжении народного хозяйства древесиной, но и в формировании почвенного покрова, водного баланса, оказывают влияние на химический состав, тепловой режим и влагосодержание нижних слоев атмосферы. Проблемы, связанные с промышленным использованием лесов, их подверженность влиянию экологических факторов, в том числе и изменения климата, а также необходимость сохранения в возможно большем объеме экосистемных функций и услуг волнует в настоящее время многих специалистов лесной отрасли. Большое внимание уделяется изучению влияния лесов на процессы в климатической системе, в том числе их роли в глобальном углеродном цикле, влагообороте, тепловом и радиационном балансах участков суши [1–8]. При этом до настоящего времени остаются мало изученными особенности энерго- и массообмена в различных по продуктивности лесах, при различном поступлении энергии, хотя эти факторы в значительной степени определяют интенсивность протекающих в лесу процессов и объем экосистемных функций и услуг. Изучение влияния климата и условий произрастания на интенсивность физических и биологических процессов позволит более объективно оценивать природоформирующую роль лесов в различающихся по географическому расположению регионах.
Климатические характеристики и условия произрастания, характеризуемые геологическим строением местности, рельефом, гидрологическими характеристиками и плодородием почв, являются основными факторами, определяющими интенсивность функционирования растительного покрова и его продуктивность. А.А. Григорьевым и М.И. Будыко было показано, что основными климатическими факторами, определяющими географическую зональность растительности, являются радиационный баланс и радиационный индекс сухости (отношение годового радиационного баланса к затратам энергии на испарение всех выпавших за год осадков) [9]. Для регионов, занятых преимущественно лесами, радиационный индекс сухости, как это отмечено в работе М.И. Будыко [9], изменяется от 0.3 до 1. При этом соответственно его возрастанию при продвижении с севера на юг увеличивается радиационный баланс. Значения индекса сухости в пределах лесной зоны свидетельствуют о достаточном ее увлажнении, что позволяет принимать в качестве основного фактора интенсивности функционирования древостоя из всех климатических показателей энергетический фактор. Интересно отметить, что из всего многообразия причин, объясняющих зональное положение древесных растений, в качестве одной из значимых исследователями рассматривается именно оптимальное увлажнение [10].
Количество поступающей энергии, наряду с условиями произрастания, оказывает влияние на продуктивность растительного покрова в соответствии с географическим положением. При этом представляет интерес выявление степени и путей влияния этих факторов на энерго- и массообмен леса и продуктивность древостоя.
На участках с однородными условиями произрастания формируются сообщества, относящиеся к определенному типу леса, который является ведущим показателем, качественно иллюстрирующим продуктивность древостоя в условиях конкретного региона. В связи с этим совершенствование типологии леса с введением новых индикаторов выделения типов леса является важной задачей лесного хозяйства [11].
Целью настоящей работы является анализ зависимости продуктивности древостоев хвойных лесов от радиационного баланса и эдафических условий, а также получение количественных оценок, характеризующих тип леса.
МАТЕРИАЛ И МЕТОДЫ
Достижение поставленной цели основано на использовании данных о пространственном распределении радиационного баланса [12] и опубликованных сведений о распределении сосняков и ельников по типам леса (с указанием их осредненного класса бонитета) в каждом административном регионе в пределах Европейской части России (ЕЧР) [13, 14]. Из-за трудностей, возникающих при измерении элементов радиационного баланса, данных о них для лесных участков немного. Используемые для построения карт и приведенные в справочниках значения радиационного баланса получены на метеостанциях, характеризующих энергетический баланс безлесной территории, к которой можно отнести, в частности, луг. Вместе с тем, известно, что радиационный баланс участка местности в большой степени зависит от характеристик деятельной поверхности. Наибольшие различия в годовых суммах радиации, поглощенной разными геосистемами, получены при сравнении метеоплощадок и лесных участков [15].
Основное влияние на соотношение величин радиационного баланса лесного массива и безлесного участка в конкретном регионе оказывают различия альбедо и длинноволнового излучения деятельной поверхности. Наблюдаемое увеличение радиационного баланса лесного массива обусловлено, в основном, пониженным поглощением коротковолновой радиации безлесным участком за счет повышенных значений альбедо [16, 17]. Значения альбедо для луга составляют 15–25%, для участков хвойного леса – 10–15%, лиственного леса – 15–20% [18]. Пространственное распределение соотношений радиационного баланса лесных и безлесных территорий зависит от продолжительности периода с положительными значениями радиационного баланса, что наблюдается при отсутствии снежного покрова [16, 19]. В результате специального анализа было установлено наличие линейной связи между значениями элементов радиационного баланса лесного участка и метеоплощадки [16, 18]. Эта связь достаточно тесная, и на нее мало влияют различия типов леса, хотя снижение плотности древостоя способствует некоторому снижению альбедо [16]. Тесная линейная связь между радиационными балансами безлесных территорий и леса, а также небольшое влияние типов леса на их радиационный баланс позволило Ю.Л. Раунеру [16] предложить метод для расчета балансов лесных участков по данным метеостанций, на основе осредненных параметров линейного уравнения. Этот метод использовался нами для определения радиационного баланса лесных участков, расположенных на территориях административных районов ЕЧР.
Количественным показателем продуктивности леса является класс бонитета. Существует тесная связь между классом бонитета и радиационным балансом территории [20]. Однако, с нашей точки зрения, большой интерес представляет исследование роли энергетического фактора в формировании запаса (объема) древесины. Поэтому в качестве характеристики продуктивности нами использовались производные класса бонитета и тесно связанные с ним запасы древесины 100-летних сосняков и ельников, которые определялись по следующей схеме. Для каждого класса бонитета конкретного типа леса, соответствующего определенному радиационному балансу административного региона, была установлена средняя высота 100-летнего древостоя по бонитировочным таблицам М.М. Орлова [21]. По высоте древостоя нами были рассчитаны запасы древесины с использованием приведенной в табличном виде зависимости между высотой и запасом древесины для насаждений с полнотой равной единице [22]. Вычисленные таким образом величины радиационного баланса лесных участков и соответствующие величины запасов 100-летних древостоев использовались нами для дальнейшего анализа.
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Влияние радиационного баланса на продуктивность древостоя
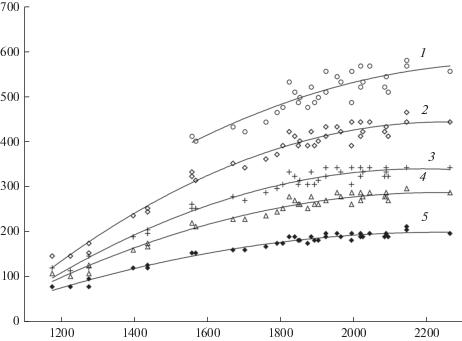
Полученные данные, характеризующие зависимость запасов 100-летних древостоев от радиационного баланса и типов леса, приведены на рис. 1, 2. Каждая точка графиков (рис. 1, 2) отражает осредненную для типа леса величину запаса 100-летнего древостоя, соответствующую энергообеспеченности участка. Нужно отметить, что такие зависимости, той или иной степени надежности, получены для типов леса, встречающихся в лесной зоне ЕЧР [13, 14]. В указанных нормативных справочниках представлено 20 типов сосновых и 18 типов еловых лесов. Их ограниченное количество на рис. 1, 2 обусловлено тем, что для построения зависимостей были использованы типы, которые имеют наибольшее пространственное распространение и, следовательно, кривые подкреплены большим количеством точек. Из-за существующей изменчивости характеристик каждого типа леса конкретного региона величина запаса стволовой древесины колеблется, однако, эти колебания происходят в узком интервале. Приведенные зависимости свидетельствуют, что изменение запаса древесины конкретного типа леса с изменением географического положения в лесной зоне зависит в основном от энергетического фактора, который является лимитирующим. Это обусловлено достаточной увлажненностью территории и достаточной обеспеченностью растений влагой.
Рис. 1.
Связь между запасом стволовой древесины 100-летнего сосняка, радиационным балансом и типом леса. 1 – сосняк кисличный, 2 – сосняк черничный, 3 – сосняк брусничный, 4 – сосняк долгомошный, 5 – сосняк багульниковый, 6 – сосняк сфагновый. По горизонтали – радиационный баланс, МДж/м2; по вертикали – запас древесины, м3/га. Fig. 1. Relationship between the stem wood stock in 100-year-old pine stands, radiation balance and forest type. 1 – Pinetum oxalidosum, 2 – Pinetum myrtillosum, 3 – Pinetum vacciniosum, 4 – Pinetum politrichosum, 5 – Pinetum ledumosum, 6 – Pinetum sphagnosum. X-axis – radiation balance, MJ/m2; y-axis – stem wood stock, m3/ha.
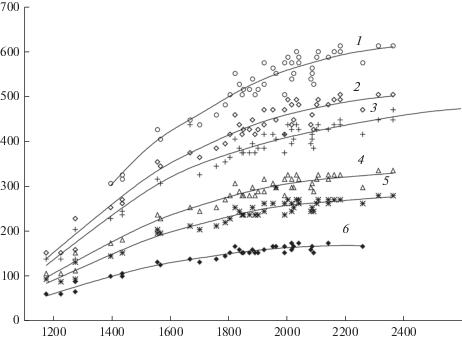
Рис. 2.
Связь между запасом стволовой древесины 100-летнего ельника, радиационным балансом и типом леса. 1 – ельник кисличный, 2 – ельник черничный, 3 – ельник брусничный, 4 – ельник долгомошный, 5 – ельник сфагновый. По горизонтали – радиационный баланс, МДж/м2; по вертикали – запас древесины, м3/га. Fig. 2. Relationship between the stem wood stock in 100-year-old spruce stands, radiation balance and forest type. 1 – Piceetum oxalidosum, 2 – Piceetum myrtillosum, 3 – Piceetum vacciniosum, 4 – Piceetum politrichosum, 5 – Piceetum sphagnosum. X-axis – radiation balance, MJ/m2; y-axis – stem wood stock, m3/ha.
Нужно отметить, что обычно в качестве характеристики энерго- и теплообеспеченности территории используется температура воздуха и ее производные (сумма температур, среднегодовая и среднемесячные величины и т.д.) из-за доступности этих метеорологических показателей. К настоящему времени выполнено много исследований, показывающих положительное влияние температуры воздуха на продуктивность лесной растительности, как в конкретном регионе, так и при изменении географического положения [23–27]. Однако, с нашей точки зрения, при анализе продуктивности растительного покрова более полную информацию можно получить, используя радиационный баланс.
Учитывая хорошее соответствие аппроксимирующих кривых эмпирическим точкам, данные зависимости могут использоваться для расчетов среднего для региона запаса 100-летнего древостоя. Кроме того, полученные зависимости позволяют сделать соответствующие выводы об особенностях формирования продуктивности древостоя в различных географических и лесорастительных условиях. Анализ траекторий кривых, характеризующих зависимости запасов стволовой древесины 100-летних древостоев от радиационного баланса, свидетельствует о том, что запасы древостоя интенсивнее возрастают с ростом величины поступающей энергии в тех типах леса, которые произрастают в более благоприятных эдафических условиях (сосняки и ельники кисличные). С ухудшением лесорастительных условий реакция растений на энергетический фактор замедляется, наименьшим изменением продуктивности характеризуются сосняки и ельники сфагновые (рис. 1, 2).
Влияние лесорастительных условий на продуктивность древостоя конкретного региона определяется в большой степени содержанием элементов питания в почве. Улучшение лесорастительных условий (характеризуемых в конкретном регионе классом бонитета) способствует увеличению концентрации элементов питания, лимитирующих рост и развитие растений, в переносимой транспирационным потоком древостоя воде и снижению удельной массы ассимиляционных органов [28, 29]. Следовательно, затраты воды и энергии на прирост единицы фитомассы древостоя будут снижаться с улучшением экологических условий, а более экономный расход этих ресурсов создает возможности для роста общих запасов фитомассы.
С увеличением радиационного баланса (что соответствует продвижению от северных границ лесной зоны к южным) продуктивность древостоя каждого типа леса возрастает. Наиболее интенсивно она увеличивается, как это следует из полученных нами зависимостей, до достижения значений радиационного баланса, равных для леса 1900–2000 МДж/м2 (рис. 1, 2), это соответствует полученным на метеостанциях значениям, составляющим 1400–1500 МДж/м2. Анализ показывает, что при этих и меньших значениях радиационного баланса радиационный индекс сухости на территории ЕЧP, как правило, не превышает 1, что свидетельствует о достаточном увлажнении рассматриваемой территории. Это обеспечивает тесную связь между продуктивностью древостоя и энергетическим фактором.
Значения радиационного баланса, соответствующие 1400–1500 МДж/м2 по данным метеостанций в пределах территории ЕЧР, отмечаются на 55°–57° с.ш. [12]. По приведенным в работе Цельникер и др. [30] результатам, продуктивность березы, лиственницы, ели европейской и сосны сибирской возрастает при продвижении с севера на юг, и максимальная продуктивность древостоя наблюдается на 56°–60° с.ш. Для этих же, примерно, широт (54°–58° с.ш.) характерна и максимальная величина листового индекса ряда древесных пород [31]. Следует отметить, что в работе Цельникер и др. [31] анализируются породы древостоя, произрастающего, как это отмечено авторами, в обеспечивающих максимальную продуктивность растений условиях. Это дает возможность исключать в исследованиях влияние условий произрастания на величину листового индекса и принимать в качестве определяющего фактора солнечную радиацию. Следовательно, полученные нами зависимости продуктивности древостоя от радиационного баланса вполне согласуются с приведенными в указанных работах выводами, и отмечается соответствие между изменениями продуктивности древостоя и величины листового индекса.
Важно отметить, что каждому типу леса соответствуют однородные лесорастительные условия [32] с определенными почвенными, гидрологическими, геологическими и орографическими характеристиками. На основании этого можно предположить, что содержание элементов питания в почвах каждого типа леса мало зависит от его географического положения, следовательно, мало меняется концентрация элементов в транспирационном потоке. При этом при продвижении с севера на юг увеличивается поступление солнечной энергии, что может сопровождаться возрастанием транспирации. В этом случае увеличение продуктивности древостоя (рис. 1, 2) может обеспечиваться в большой степени за счет возрастания поступления минеральных элементов в растения при росте переносящего эти элементы объема транспирируемой воды. Для определения транспирации древостоем предложен достаточно простой метод расчета [33–35], увязывающий основные факторы этого процесса:
где Et – транспирация древостоем, мм; m – масса листвы, т/га; Kt – коэффициент транспирационной активности.Из уравнения следует, что увеличение объема транспирации и, следовательно, перенос минеральных элементов с потребляемой растением водой возможен за счет возрастания коэффициента транспирационной активности и, при его постоянстве, за счет увеличения массы листвы. Соответствие между ростом листового индекса (массы листвы) [31] и ростом продуктивности древостоя [30], а также увеличением запасов стволовой древесины различных типов леса (рис. 1, 2) при продвижении с севера на юг, примерно, до одних и тех же широт, позволяет объяснить возрастание продуктивности древостоя увеличением массы листвы. Увеличение массы листвы для каждого типа леса создает возможность большего поглощения солнечной радиации, значительная часть которой в лесу расходуется на транспирацию древостоем. Соответствие же между транспирацией и количеством листвы позволяет допустить, как следует из формулы, что изменение географического положения древостоя мало отражается на коэффициенте транспирационной активности. Это согласуется со сделанными ранее многими исследователями выводами о независимости для конкретной древесной породы величины этого коэффициента от географического положения.
Энергетическая “цена” запаса стволовой древесины разных типов леса в различных климатических условиях
Затраты радиационного баланса на производство единицы запаса стволовой древесины (удельные величины радиационного баланса, полученные как отношение радиационного баланса к запасам стволовой древесины) зависят от лесорастительных условий и поступления солнечной энергии. Чем хуже условия роста растений, тем выше энергетическая “цена” единицы запаса древесины. Кроме того, величина энергии, затрачиваемая на производство единицы запаса древесины каждого типа леса, меняется в широтном направлении в соответствии с изменением радиационного баланса. Это следует из приведенных на рис. 3, 4 зависимостей.
Рис. 3.
Зависимость удельной величины радиационного баланса сосняка (отношение радиационного баланса леса к запасу древесины) от радиационного баланса для различных типов леса. 1 – сосняк кисличный; 2 – сосняк черничный; 3 – сосняк вересковый; 4 – сосняк багульниковый; 5 – сосняк сфагновый. По горизонтали – радиационный баланс, МДж/м2; по вертикали – удельная величина радиационного баланса, МДж/м3. Fig. 3. Dependence of the pine stand radiation balance specific value (the ratio of the forest radiation balance to the stem wood stock) on the radiation balance in different types of forest. 1 – Pinetum oxalidosum, 2 – Pinetum myrtillosum, 3 – Pinetum callunosum, 4 – Pinetum ledumosum, 5 – Pinetum sphagnosum. X-axis – radiation balance, MJ/m2; y-axis – specific value of the radiation balance, MJ/m3.
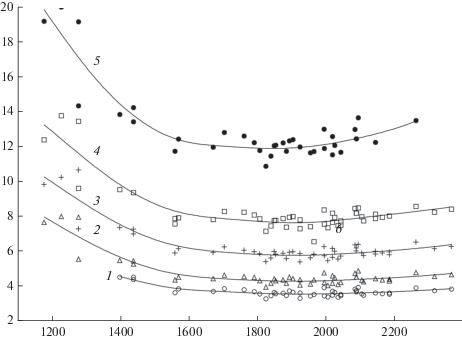
Рис. 4.
Зависимость удельной величины радиационного баланса ельника (отношение радиационного баланса леса к запасу древесины) от радиационного баланса для различных типов леса. 1 – ельник кисличный; 2 – ельник черничный; 3 – ельник брусничный; 4 – ельник долгомошный; 5 – ельник сфагновый. По горизонтали – радиационный баланс, МДж/м2; по вертикали – удельная величина радиационного баланса, МДж/м3. Fig. 4. Dependence of the spruce stand radiation balance specific value (the ratio of the forest radiation balance to the stem wood stock) on the radiation balance in different types of forest. 1 – Piceetum oxalidosum, 2 – Piceetum myrtillosum, 3 – Piceetum vacciniosum, 4 – Piceetum politrichosum, 5 – Piceetum sphagnosum. X-axis – radiation balance, MJ/m2; y-axis – specific value of the radiation balance, MJ/m3.
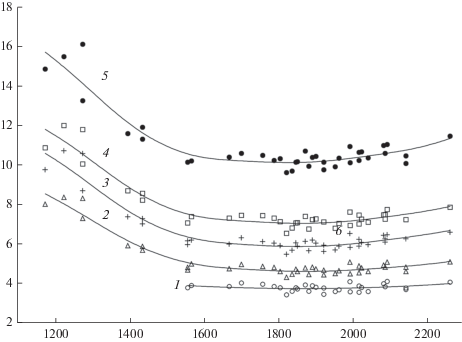
Наибольшими удельными затратами энергии на производство стволовой древесины характеризуется северная часть лесной зоны. Интенсивное снижение энергозатрат при продвижении с севера на юг происходит до значений радиационного баланса леса 1600–1700 МДж/м2 (рис. 3, 4). Для безлесной территории эти значения соответствуют 1150–1200 МДж/м2, таким образом южная граница интенсивного снижения энергозатрат находится примерно на 62°–63° с.ш. [12].
Это снижение удельных энергозатрат каждого типа леса может объясняться изменениями, которые происходят в различных компонентах экосистем. Известно, что с увеличением температуры воздуха возрастает скорость разложения опада и в целом круговорота химических веществ, что может усиливать интенсивность поступления питательных веществ к сосущим окончаниям корневых систем [26]. При изучении эктомикоризных сообществ в бореальных лесах [36], были отмечены изменения их структуры в различных абиотических условиях, что также может влиять на эффективность взаимодействия симбионтов. Некоторые исследователи отмечают изменение структуры биомассы лесных сообществ в зависимости от их географического положения [37], в частности, увеличение соотношения листва/тонкие корни при перемещении с севера на юг, что может создавать возможность большего потребления энергии на прирост стволовой древесины, чем на прирост тонких корней.
Значения минимальных затрат радиационного баланса на рост сосновых и еловых лесов заключены в интервале 1600–1700…2000–2100 МДж/м2 (соответственно 1150–1200…1500–1600 МДж/м2 для безлесных участков). Эта территория в ЕЧР расположена между 61°–62° и 53°–55° с.ш. Малое изменение удельных энергозатрат для конкретных типов леса в указанном интервале радиационного баланса можно объяснить тем, что устанавливается соответствие между увеличением поступления энергии и увеличением массы листового аппарата, обеспечивающей рост транспирации и, следовательно, рост потребления минеральных веществ растением. При этом можно предположить, что на этой территории структура биомассы древостоя (соотношение массы корней, листвы, ветвей и ствола) мало меняется. Минимальные значения удельных энергозатрат можно принимать, по-видимому, как свидетельство оптимальных условий произрастания.
Приведенные на рис. 3, 4 зависимости, характеризующие влияние условий роста на удельные энергозатраты, могут быть объяснены при анализе процессов в лесных биогеоценозах. К наиболее энергозатратным процессам теплового баланса территории с достаточным увлажнением относится испарение. Доля радиационного баланса, затрачиваемая на испарение, для лесной зоны составляет 70–80% [38]. Транспирация, составляющая большую часть суммарного испарения с леса, является важным физиологическим процессом, участвующим в снабжении растения и распределении в нем минеральных элементов. Ее величина связана с плодородием почв [29]. Уменьшение концентрации минеральных элементов в ксилемном соке, составляющем транспирационный поток, с ухудшением лесорастительных условий и снижение продуктивности древостоя выявлено в исследовании, основанном на прямых измерениях [28]. Авторами показано, что в вересковом сосняке, наиболее продуктивном из исследуемых сообществ (III, 7 класс бонитета), концентрация азота в ксилемном соке составляла в разные годы 41.4 и 42.5 мг/л. В менее продуктивных сосняках кустарничково-долгомошном (IV, 4 класс бонитета) и лишайниковом (IV, 4 класс бонитета) этот показатель составлял соответственно 28.6 и 31.7 мг/л. Снижение концентрации минеральных элементов в транспирационном потоке предполагает увеличение удельного объема этого потока. Этим объясняется увеличение энергозатрат на транспирацию, обеспечивающую жизнедеятельность и рост единицы фитомассы, и соответствующий рост энергетической “цены” единицы объема стволовой древесины с ухудшением лесорастительных условий.
Усредненные удельные значения радиационного баланса, относящиеся к интервалу оптимальных условий произрастания древостоев, могут использоваться в качестве количественной характеристики типов леса. Для некоторых типов леса, взятых из вышеуказанных работ [13, 14], эти значения, полученные по приведенным выше методам, приведены в табл. 1.
Таблица 1.
Удельные энергозатраты на создание стволовой древесины 100-летнего древостоя в разных типах леса Table 1. Specific energy consumption for the stem wood buildup in 100-year-old stands of different forest types
| Тип леса Forest type |
Удельные энергозатраты, МДж/м3 Specific energy consumption, MJ/m3 |
Тип леса Forest type |
Удельные энергозатраты, МДж/м3 Specific energy consumption, MJ/m3 |
|---|---|---|---|
| Pinetum corylosum | 3.28 | Piceetum tiliosum | 2.99 |
| Pinetum tiliosum | 3.40 | Piceetum oxalidosum | 3.78 |
| Pinetum oxalidosum | 3.61 | Piceetum pteridiosum | 4.16 |
| Pinetum pteridiosum | 3.81 | Piceetum pleuroziosum | 4.72 |
| Pinetum pleuroziosum | 4.10 | Piceetum myrtillosum | 4.72 |
| Pinetum myrtillosum | 4.38 | Piceetum polypodophytosum | 5.14 |
| Pinetum herbosum | 4.59 | Piceetum politrichoso-myrtillosum | 5.64 |
| Pinetum vacciniosum | 4.88 | Piceetum vacciniosum | 6.04 |
| Pinetum callunosum | 5.88 | Piceetum politrichosum | 7.21 |
| Pinetum politrichosum | 6.57 | Piceetum fruticuloso-sphagnosum | 8.42 |
| Pinetum cladinosum | 7.62 | Piceetum carioso-sphagnosum | 9.08 |
| Pinetum ledumosum | 7.82 | Piceetum sphagnosum | 10.40 |
| Pinetum cariosum | 8.98 | ||
| Pinetum sphagnosum | 12.2 |
Даже при отсутствии в настоящее время достаточно строгих критериев выделения типов леса, на что указывает С.К. Глушко [11], следует отметить, что энергетическая “цена” 1 м3 древесины однородных по напочвенной растительности участков мало зависит от лесообразующей породы. Это дает дополнительное основание для рассмотрения удельных энергозатрат в качестве возможных количественных критериев типов леса.
ЗАКЛЮЧЕНИЕ
Основным показателем, иллюстрирующим продуктивность древостоя конкретного региона, является тип леса. В пределах лесной зоны, характеризуемой достаточным увлажнением, продуктивность конкретного типа леса определяется энергетическим фактором. При продвижении с севера на юг до 55°–57° с.ш. с увеличением радиационного баланса продуктивность древостоев возрастает, что обусловлено увеличением массы ассимиляционных органов. При этом запас древостоя увеличивается с ростом величины поступающей энергии более интенсивно в тех типах леса, которые произрастают в более благоприятных эдафических условиях. Интенсивность возрастания запаса снижается от сосняков и ельников кисличных к соснякам и ельникам сфагновым.
С ухудшением лесорастительных условий затраты энергии на создание единицы объема стволовой древесины возрастают. Энергозатраты каждого типа леса зависят от широты местности. Минимальными затратами энергии на создание единицы объема стволовой древесины 100-летнего хвойного древостоя, определяющими оптимальные условия произрастания, характеризуется территория между 61°–62° и 53°–55° с.ш. Величину этих затрат можно использовать в качестве количественного критерия при типологической классификации лесов.
Список литературы
Ваганов Е.А., Ведрова Э.Ф., Верховец С.В., Ефремов С.П., Ефремова Т.Т., Круглов В.Б., Онучин А.А., Сухинин А.И., Шибистова О.Б. 2005. Леса и болота Сибири в глобальном цикле углерода. – Сибирский экологический журнал. 4: 631–649. https://www.elibrary.ru/item.asp?id=9130850
Чебакова Н.М., Выгодская Н.Н., Арнет А., Белелли Маркезини Л., Колле О., Курбатова Ю.А., Парфенова Е.И., Валентини Р., Ваганов Е.А., Шульце Е.Д. 2013. Энерго- и массообмен и продуктивность основных экосистем Сибири (по результатам измерений методом турбулентных пульсаций). 1. Структура теплового баланса за период вегетации. – Известия РАН. Серия биологическая. 6: 728–737. https://doi.org/10.7868/S0002332913060039
Герасимов Ю., Хетемяки Л, Йонсон Р., Катила П., Келломяки С., Коскела Т, Кранкина О., Люндмарк Т., Моен Й., Месьер К., Мийликяйнен К., Наскали А., Нордин А., Саастамойнен О., Ванханен Х. 2014. Бореальные леса для человека и биоразнообразия. – Устойчивое лесопользование. 2(39): 2–12. https://wwf.ru/upload/iblock/18c/01-_7_.pdf
Курганова И.Н., Лопес де Гереню В.О., Мякшина Т.Н., Сапронов Д.В., Кудеяров В.Н. 2017. Оценка климаторегулирующей функции лесных экосистем южного Подмосковья. – В сб.: Бореальные леса: состояние динамика, экосистемные услуги: Тез. докл. Всероссийской научной конференции. Петрозаводск. С. 160–162. http://resources.krc.karelia.ru/forestry/doc/publ/borealnye_lesa_il_2017.pdf
Blanken P.D., Black T.A., Yang P.C., Neumann H.H., Nesic Z., Staebler R., den Hartog G., Novak M.D., Lee X. 1997. Energy balance and canopy conductance of a boreal aspen forest: Partitioning overstory and understory components. – J. Geophys. Res. Atmos. 102(D24): 28915–28927. https://doi.org/10.1029/97JD00193
Mahli Y., Baldocchi D.D., Jarvis P.G. 1999. The carbon balance of tropical, temperate and boreal forests. – Plant Cell Environ. 22(6): 715–740. https://doi.org/10.1046/j.1365-3040.1999.00453.x
Kurtén T., Kulmala M., Dal Maso M., Suni T., Reissell A., Vehkamäki H., Hari P., Laaksonen A., Viisanen Y., Vesala T. 2003. Estimation of different forest-related contributions to the radiative balance using observations in southern Finland. – Boreal Env. Res. 8(4): 275–285. http://www.borenv.net/BER/archive/pdfs/ber8/ber8-275.pdf
Pan Y., Birdsey R.A., Fang J., Houghton R., Kauppi P.E., Kurz W.A., Phillips O.I. 2011. A large and persistent carbon sink in the world’s forests. – Science. 333(6045): 988–993. https://doi.org/10.1126/science.1201609
Будыко М.И. 1971. Климат и жизнь. Л. 472 с.
Абатуров Б.Д., Лопатин В.Н. 2002. Связь зональной структуры растительного покрова с особенностями углеродного обмена и энергетики деревьев, трав и кустарников. – Поволжский экологический журнал. 1: 3–18. http://www.sevin.ru/volecomag/issues/2002_1/PEJ_2002_1_3-18.pdf
Глушко С.Г. 2017. Роль типологии леса в современном лесном хозяйстве. – В сб.: Бореальные леса: состояние динамика, экосистемные услуги: Тез. докл. Всероссийской научной конференции. Петрозаводск. С. 78–80. http://resources.krc.karelia.ru/forestry/doc/publ/borealnye_lesa_il_2017.pdf
Атлас СССР. 1985. М. 260 с.
Производительность сосновых насаждений по типам леса (экологические нормативы). 1990. Петрозаводск. 43 с.
Производительность еловых насаждений по типам леса (экологические нормативы). 1991. Петрозаводск. 44 с.
Максютова Е.В. 2006. Оценка радиационного баланса геосистем Минусинской котловины. – География и природные ресурсы. 3: 69–75. http://www.izdatgeo.ru/pdf/gipr/2006-3/69.pdf
Раунер Ю.Л. 1972. Тепловой баланс растительного покрова. Л. 210 с.
Wicke W., Bernhofer Ch. 1996. Energy balance comparison of the Hartheim forest and an adjacent grassland site during the HartX experiment. – Theor. App. Climatol. 53(1–3): 49–58. https://link.springer.com/article/10.1007/BF00866410
Руднев Н.И. 2003. Средообразующая роль растительности тропических и умеренных широт Евразии. М. 307 с.
Lafleur P.M., Renzetti A.V., Bello R. 1993. Seasonal changes in the radiation balance of subarctic forest and tundra. – Arctic and Alpine Research 25(1): 32–36. https://www.tandfonline.com/doi/pdf/10.1080/00040851.1993.12002977
Карпечко Ю.В., Кондратьев С.А., Родионов В.З., Шмакова М.В. 2020. Особенности формирования испарения в различных по возрасту, условиям произрастания и продуктивности лесах. – Гидрометеорология и экология. Ученые записки РГГМУ. 58: 49–67. https://doi.org/10.33933/2074-2762-2020-58-49-67
Лесотаксационный справочник. 1980. М. 288 с.
Лесотаксационные таблицы. 1976. Петрозаводск. 33 с.
Молчанов А.А. 1976. Дендроклиматические основы прогнозов погоды. М. 168 с.
Кокорин А.О., Назаров И.М. 1994. Оценка влияния потепления климата и роста потока фотосинтетически активной радиации на бореальные леса. – Метеорология и гидрология. 5: 44–54.
Книзе А.А., Романюк Б.Д., Федорчук В.Н. 2000. Динамика продуктивности таежных и подтаежных лесов ЕТР за последнее столетие. – Изв. РГО. 132(3): 63–66.
Шанин В.Н., Михайлов А.В., Быховец С.С., Комаров А.С. 2010. Глобальные изменения климата и баланс углерода в лесных экосистемах бореальной зоны: иммитационное моделирование как инструмент прогноза. – Известия РАН. Серия биологическая. 6: 719–730. https://www.elibrary.ru/item.asp?id=15523640
Myneni R.B., Keeling C.D., Tucker C.J., Asrar G., Nemani R.R. 1997. Increased plant growth in the northern high latitudes from 1981 to 1991. – Nature. 386(6626): 698–702. https://doi.org/10.1038/386698a0
Сазонова Т.А., Придача В.Б., Колосова С.В. 2009. О содержании элементов минерального питания в ксилемном соке Pinus sylvestris (Pinaceae). – Растительные ресурсы. 45(1): 113–121. https://www.elibrary.ru/item.asp?id=17013594
Карпечко Ю.В., Мясникова Н.А. 2014. Особенности формирования листового аппарата и транспирации хвойного древостоя. – Ученые записки Петрозаводского государственного университета. Сер. “Естественные и технические науки”. 4(141): 78–84. http://uchzap.petrsu.ru/files/n141.pdf
Цельникер Ю.Л., Корзухин М.Д., Семенов С.М. 2010. Модельный анализ широтного распределения продуктивности лесных пород России. – Лесоведение. 2: 36–45. https://www.elibrary.ru/item.asp?id=13754348
Цельникер Ю.Л., Малкина И.С., Завельская Н.А. 2002. Географические аспекты фотосинтеза у лесных деревьев России. – В сб.: Проблемы экологического мониторинга и моделирования экосистем. СПб. Т. XVIII. С. 81–108. http://downloads.igce.ru/publications/pemem/pemem2002_18.pdf
Стандарт отрасли ОСТ 56-108-98 “Лесоводство. Термины и определения” (утв. приказом Рослесхоза от 3 декабря 1998 г. N 203).
Хильми Г.Ф. 1966. Основы физики биосферы. Л. 299 с.
Кайбияйнен Л.К. 1986. Сбалансированность системы водного транспорта у сосны обыкновенной. IV. Общие характеристики водного режима в разных экологических условиях. – Лесоведение. 4: 70–75. https://www.elibrary.ru/item.asp?id=21838205
Крестовский О.И. 1986. Влияние вырубок и восстановления лесов на водность рек. Л. 118 с.
Defrenne C.E., Philpott T.J., Guichon S.H.A., Roach W.J., Pickles B.J., Simard S.W. 2019. Shifts in Ectomycorrhizal Fungal Communities and Exploration Types Relate to the Environment and Fine-Root Traits Across Interior Douglas-Fir Forests of Western Canada. – Front. Plant Sci. 10: 643. https://doi.org/10.3389/fpls.2019.00643
Helmisaari H.-S., Derome J., Nöjd P., Kukkola M. 2007. Fine root biomass in relation to site and stand characteristics in Norway spruce and Scots pine stands. – Tree Physiology. 27(10): 1493–1504. https://doi.org/10.1093/treephys/27.10.1493
Несмелова Е.И., Филиппова Б.Г. 1995. Микроклиматология. Условия формирования микроклиматических режимов. М. 165 с.
Дополнительные материалы отсутствуют.
Инструменты
Растительные ресурсы