

Российский физиологический журнал им. И.М. Сеченова, 2019, T. 105, № 5, стр. 608-618
Влияние кратковременного и длительного неконтролируемого стресса на животных доминантного и субординантного социального статуса с различным типом стрессорной реакции
В. В. Немец 1, *, В. И. Шмурак 2, В. Е. Соболев 2, В. В. Гарнюк 2, Е. Д. Рован 2, Е. П. Виноградова 3
1 Национальный медицинский исследовательский центр им. В.А. Алмазова
Санкт-Петербург, Россия
2 2Научно-исследовательский институт гигиены, профпатологии и экологии человека
г.п. Кузьмоловский, ст. Капитолово, Ленинградская область, Россия
3 Санкт-Петербургский государственный университет, биологический факультет
Санкт-Петербург, Россия
* E-mail: seva_nemets@list.ru
Поступила в редакцию 15.08.2018
После доработки 08.02.2019
Принята к публикации 05.03.2019
Аннотация
Цель данной работы – изучение влияния кратковременного и длительного неконтролируемого стрессорного воздействия на поведение животных различного типа стрессорной реакции и социального статуса. Кратковременный стресс (неизбегаемое плавание в течение 5-ти минут) приводит к угнетению двигательной активности у субординантов с неактивным типом стрессорной реакции, в то время как у активных доминантов проявляется лишь один поведенческий компонент стрессорного ответа – груминг. При действии длительного неконтролируемого стресса (иммобилизация в тесных пеналах 5 дней/4 ч) у активных доминантов в отличие от неактивных субординантов наблюдаются признаки депрессивно-подобного состояния, выраженного в достоверном снижении показателей динамики массы тела, увеличении уровня агедонии, тревожности, снижении двигательной и исследовательской активности на 5-й день воздействия, увеличении болевого порога по сравнению с фоном. У субординантов с неактивным типом стрессорной реакции на 5-й день воздействия наблюдается стрессорная реакция, выраженная в снижении двигательной и исследовательской активности и увеличении тревожности. Кратковременный и длительный неконтролируемый стресс по-разному влияет на животных с различным социальным статусом и стратегией поведения. Эти данные могут быть полезны для индивидуального подбора соответствующих антидепрессантов, исходя из психофизиологических характеристик.
Стрессорные воздействия различной длительности и силы оказывают разные влияния на организм. Кратковременные стрессорные воздействия, с одной стороны, могут активировать когнитивные функции особи [1, 2] с другой – могут привести к депрессивным проявлениям при их хроническом воздействии [3].
Хроническое стрессорное воздействие может нанести существенный вред организму. Чтобы противостоять такому стрессору (аверсивному воздействию) в организме запускаются процессы адаптации, но в случае если организм не в состоянии справиться со стрессором, наступает фаза истощения и гибели [4]. Длительное субхроническое (3–5 дней) и хроническое (40–60 дней) стрессорное воздействие сопровождается активацией как симпатоадреналовой (САС), так и гипофизадреналовой системы (ГАС) и негативно влияет на процессы обучения и памяти, вследствие разрушения нейронов в гиппокампе под действием глюкокортикоидов [5].
Наибольшее неблагоприятное воздействие на организм оказывает неконтролируемый стресс – воздействие, к которому невозможно приспособиться, избежать или избавиться от него, также невозможно предсказать его начало и конец [6].
Как кратковременный [7], так и длительных стресс [8] приводят к подъему кортикостерона в плазме крови мышей [9] и изменению моноаминов в ЦНС у крыс [10], угнетению поведенческой активности, кроме того, показана положительная корреляция с исходным уровнем двигательной активности животного [11].
Показано, что хронический умеренный неконтролируемый стресс (модель стресса повседневности), когда животных в течение 1 месяца ежедневно подвергали не сильным, но постоянно изменяющимся условиям среды (мокрая подстилка, инверсия освещения, наклон клетки и т.п.) приводил к развитию депрессивно-подобного состояния [12]. Важно отметить, что степень выраженности депрессивно-подобного состояния зависела от стратегии поведения животных, и была также более выражена у активных, но не пассивных животных [3].
В процессе адаптации к стрессорным воздействиям животное демонстрирует различные генетически детерминированные стратегии поведения – “борьба”, “бегство” или “затаивание” [13]. Данные стратегии определяют тип стрессорной реакции организма [14].
У животных с полярным (“активным” и “пассивным”) типом стрессорной реакции наблюдается различный уровень активации ГАС и САС во время стресса, отражающийся в уровне стрессорных гормонов (кортизола, адреналина) и нейромедиаторов (дофамин, серотонин) [15].
Существует несколько подходов к разделению животных по поведенческим типам. Ключевым в процессе рандомизации является признак, по которому будет происходить отбор. Тест “УРАИ” позволяет выявить животных с различной стратегией поведения в условиях электроболевого стресса. Тест “резидент-интрудер” позволяет оценить уровень агрессивности животных. Существует альтернативный тест “водная депривация”, позволяющий быстро и эффективно выявить доминантов и субординантов в популяции крыс. Известно, что доминанты часто демонстрируют территориальную агрессию, а также агрессию в процессе борьбы за витальные ресурсы, что характерно для носителей “активного” поведенческого типа [16]. В литературе показано, что животные доминантного социальным ранга демонстрируют достоверно большую активность в челночной камере, чем субординанты, однако не было обнаружено достоверных различий в уровне кортикостерона между группами [17].
Таким образом, целью данной работы послужило изучение воздействия кратковременного и длительного неконтролируемого стресса на животных с “активным” и “неактивным” поведенческим типом стрессорной реакции и различным социальным статусом.
МЕТОДЫ ИССЛЕДОВАНИЯ
Экспериментальные исследования проводили на белых беспородных крысах-самцах (n = 60) массой 280–320 г., питомник “Пущино”, возраст на момент начала эксперимента – 3 мес. Животных содержали в виварии, в стандартных условиях (5 крыс в клетке) при световом режиме (12/12 ч), со свободным доступом к воде и корму. Работа с лабораторными животными осуществлялась согласно протоколу исследований в соответствии с Женевской конвенцией 1985 г. о “Международных принципах биомедицинских исследований с использованием животных” и Хельсинкской декларацией 2000 г. о гуманном отношении к животным [18], (протокол закрытого заседания этического комитета ФГУП НИИГПЭЧ № 001010/18 05.11.2017).
Перед началом эксперимента были произведены фоновые измерения поведенческих реакций и физиологических показателей. Для этого были использованы следующие тесты: “приподнятый крестообразный лабиринт” (ПКЛ), тест “потребление сладкого раствора”, тест “распознавание нового объекта”, тест “отдергивание хвоста”.
В настоящее время тест ПКЛ считается наиболее адекватным для оценки тревожности у лабораторных животных [19]. Установка состоит из двух открытых рукавов длиной 50 см, шириной 10 см, и двух закрытых рукавов такой же длины и ширины, высотой не менее 40 см над поверхностью лабиринта. Размеры входов в закрытые рукава составляют 10 × 10 см. Они расположены друг напротив друга на расстоянии 10 см. Пол каждого из рукавов расчерчен на квадраты 10 × 10 см, по 5 квадратов на каждом рукаве. ПКЛ располагается на высоте не менее 75 см над полом.
Всех животных сажали носом по направлению к центру лабиринта. После тестирования каждого из животных лабиринт протирался 11%-ным раствором перекиси водорода для уничтожения запаховых сигналов от предыдущей крысы. С помощью данного теста оценивали уровень тревожности, двигательную и исследовательскую активность. Регистрацию поведенческих показателей осуществляли с помощью программы “Поведение крыс” © Nicolai Kamyshev 2007. Программа регистрировала длительность каждого поведенческого акта в % за 100 с измерения. Количество пройденных квадратов регистрировали количественно, M ± m. Процедура подробно описана ранее [3].
Для выявления изменения гедонистической мотивации использовали тест “потребление сладкого раствора” (двухбутылочный тест). В течение 18-ти часов, преимущественно в ночное время суток (16:00–10:00) крысам, рассаженным в одиночные клетки, был одновременно предоставлен доступ к выбору между двумя бутылками, одна из которых содержала 1%-ный раствор сахарозы, а другая — воду. Потребление воды и раствора сахарозы было оценено путем взвешивания бутылки. Предпочтение раствора сахарозы вычисляли по формуле: коэффициент предпочтения = масса раствора сахара/(масса раствора сахара + масса раствора воды)) × 100%.
Тест “распознавание нового объекта” является диагностическим и распространен для выявления нарушений зрительной памяти и когнитивных процессов у лабораторных животных. Тест направлен на естественную исследовательскую активность грызунов в процессе изучения окружающей среды. В распознавании нового объекта участвует зрительная кора области V1 (часть затылочной коры, ответственной за визуальное восприятие) и периринальная кора больших полушарий. В экспериментах показано, что удаление этих отделов головного мозга приводит к невозможности идентифицировать новый объект в сравнении с ранее изученным. В первой части теста в течение 8–10 мин животным предъявляли зрительный стимул в виде объекта круглой, квадратной или другой формы. Через 60 мин объект предъявляли повторно (старый объект – “С”) в течение 3-х мин вместе с новым объектом (новый объект – “Н”), отличающимся по цвету, форме и текстуре. Регистрировали время изучения каждого из объектов. Рейтинг предпочтения нового объекта высчитывали, исходя из разницы времени изучения объектов по формуле (Н – С/С + Н). Эксперимент проводился в одноместных клетках для исключения неофобии у животных. Процедура тестирования подробно описана ранее [20].
Для измерения болевого порога у крыс применялся тест “отдергивание хвоста” с использованием аппарата PANLAB 7160, Испания. Адаптации к установке не производилась, учитывалось одно измерение, нагревание прибора происходило в течение 20 мин, финальная температура 50°С. В процессе тестирования регистрировали латентный период реакции “отдергивания” хвоста в ответ на тепловое раздражение.
Для выявления особенностей действия кратковременного и длительного неконтролируемого стресса, производили рандомизацию животных по типу стрессорной реакции (активные и пассивные) и по социальному статусу в клетке (доминанты, субдоминанты, субординанты).
Для тестирования стратегии поведения в ситуации электроболевого стресса использовали тест условного рефлекса активного избегания “УРАИ”. Установка была создана коллективом НИИ ГПЭЧ [21]. Тестирование производили в течение 2-х дней (25 подходов/день). Сила тока при электроболевом подкреплении определялась, исходя из индивидуальной чувствительности животного – до 1 мА. По данным, полученным ранее [3], эффективность выработки навыка реакций “избегания” на условный сигнал – звук на второй день тестирования – определяет активность стратегии поведения животного.
Тест “конкуренция за воду” использовался для выявления социального статуса животных в клетке [7, 22]. Крыс, находящихся в домашних клетках (n = 5) с устоявшейся иерархией, подвергали водной депривации в течение 24-х ч. После этого устанавливали поилку с водой на специальном штативе. По данным литературы и по опыту в данной лаборатории, иерархия у крыс устанавливается обычно в течение 7-ми дней после помещения в новую клетку. Стабильность иерархии определялась незначительным количеством социальных конфликтов в клетке в процессе лабораторных манипуляций (чистка клеток и т.д.), а также отсутствием конкуренции за какие-либо ресурсы (пища и вода в свободном доступе) и других стрессорных воздействий. Поведение крыс регистрировали с помощью видеокамеры. Время каждого тестирования составляло 10 мин. При анализе видеозаписи для каждого животного, предварительно помеченного яркой меткой, подсчитывали латентное время подхода к поилке, число и эффективность агрессивных атак, количество выпитой воды. Социальный статус вычислялся по эффективности конкуренции за витальный ресурс (воду) по совокупности поведенческих показателей [23]. Тестирование социального статуса проводили три раза с интервалом раз в 2 нед., к соответствующим поведенческим группам относили животных с результативностью 2/3 и 3/3.
По окончании процедур рандомизации из общей группы крыс-самцов (n = 60) были выделены следующие подгруппы: “активные доминанты” АД (n = 15) и “неактивные субординанты” СН (n = 15). Отобранных для дальнейшего эксперимента животных содержали в их домашних клетках для сохранения иерархического статуса.
Через неделю после окончания тестирования индивидуальных поведенческих характеристик животных подвергали процедурам стрессорного воздействия. Для создания острого кратковременного неконтролируемого воздействия использовали тест неизбегаемого плавания (тест Порсолта), подробно см. [3]. Процедура производилась в течение 5-ти мин. Через 4 ч после воздействия оценивали уровень тревожности животных в тесте ПКЛ. Данное время наиболее адекватно для оценки тревожности после стрессорного воздействия [24]. Для реализации процедуры длительного неконтролируемого стресса использовали иммобилизацию в специальных пеналах. Длительность воздействия составляла 4 ч (с 10.00 до14:00) ежедневно в течение 5-ти суток, после этого производили оценку поведенческого компонента стрессорного ответа.
Статистический анализ полученных данных осуществляли с помощью программного пакета Statistica 6.0. Использовали непараметрические методы статистического анализа, U-критерий Манна–Уитни и коэффициент корреляции Спирмена.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
В группе крыс-самцов (n = 60) с помощью теста “УРАИ” были выделены животные с высокой и низкой активностью в стрессогенных условиях, а при помощи теста “конкуренция за воду” был определен их социальный статус в клетке. В результате сопоставления данных двух тестов было показано, что 42% высокоактивных животных, выявленных по тесту “УРАИ”, являлись носителями доминантного социального статуса, а 75% животных с пассивной стратегией поведения занимали субординантный социальный статус (табл. 1).
Таблица 1.
Соотношение животных различного социального статуса и стратегий поведения в группе самцов крыс
| Стратегии поведения в стрессорной ситуации | Социальные группы |
|---|---|
| Высоко активные животные (n = 23) | 42% доминанты |
| 28% субдоминанты | |
| 28% субординанты | |
| Низко активные животные (n = 37) | 5% доминанты |
| 20% субдоминанты | |
| 75% субординанты |
Здесь и в табл. 2 для выявления стратегий поведения в стрессорной ситуации использовали тест “УРАИ”, социальный статус животных определяли с помощью теста “конкуренция за воду”.
С использованием тестов “УРАИ” и “конкуренция за воду” (рис. 1) были сформированы экспериментальные группы (по 15 животных в группе): “активные доминанты” АД и “неактивные субординанты” СН. Критерий отбора – более 70% реакций “избегания” – “активные животные” (тест “УРАИ”), высокое количество агрессивных контактов, латентный период подхода к поилке – менее 10% от общего времени тестирования. В результате отбора количество доминантов в сформированной группе АД составило 84%, а субординантов в группе СН – 88% (табл. 2).
Рис. 1.
Поведенческие характеристики животных групп “активные доминанты” и “неактивные субординанты” в тестах “УРАИ” (А, Б) и “конкуренция за воду” (В). На рис. 1А, В по вертикали указано количество поведенческих элементов (N). На рис. 1Б по вертикали – время в с., ЛП – латентный период подхода к поилке. Темные столбцы – “активные доминанты”, светлые столбцы – “неактивные субординанты”. * – различия между группами статистически значимы. р ≤ 0.05. M ± m.
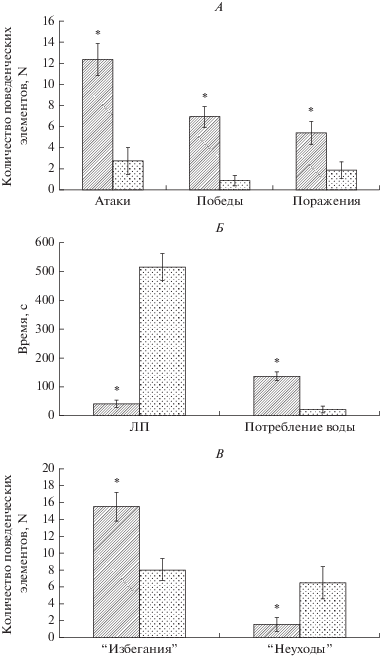
Таблица 2.
Соотношение животных различного социального статуса и стратегии поведения в экспериментальных группах
| Группы животных | Стратегии поведения в стрессорной ситуации | Социальные группы |
|---|---|---|
| АД (n = 15) | 100% Высоко активные животные |
84% доминанты |
| 6% субдоминанты | ||
| 10% субординанты | ||
| СН (n = 15) | 100% Низко активные животные |
0% доминанты |
| 12% субдоминанты | ||
| 88% субординанты |
После кратковременного неизбегаемого стресса обнаружено статистически значимое снижение двигательной активности в группе СН по сравнению с фоновыми значениями, в группе АД различия не достоверны (табл. 3). В группе АД обнаружено статистически значимое увеличение длительности груминга, в группе СН различия не достоверны.
Таблица 3.
Поведенческие показатели в тесте ПКЛ у крыс после кратковременного неконтролируемого стресса (M ± m)
| Сроки | Группы животных | Количество пройденных квадратов | Длительность стоек, % | Длительность груминга, % | Время в открытых рукавах, % | Время в закрытых рукавах, % |
|---|---|---|---|---|---|---|
| До стресса | АД | 26.4 ± 3.4 | 3.5 ± 0.9 | 7.2 ± 1.6 | 29.4 ± 6.2 | 70.6 ± 6.1 |
| СН | 26.3 ± 3.5 | 4.5 ± 0.8 | 14.6 ± 2.6 | 16.6 ± 2.5# | 83.3 ± 5.1# | |
| После стресса | АД | 17.6 ± 3.0 | 4.0 ± 1.0 | 20.3 ± 4.0* | 15.9 ± 5.8 | 78.2 ± 7.5 |
| СН | 13.8 ± 2.1** | 4.7 ± 1.1 | 24.2 ± 5.6 | 22.9 ± 7.7 | 77.1 ± 7.7 |
После длительного иммобилизационного стресса у крыс обеих групп обнаружена стрессорная реакция, более выраженная у неактивных крыс: показано статистически значимое снижение количества пройденных квадратов, увеличение длительности груминга, снижение времени нахождения в открытых рукавах (табл. 4).
Таблица 4.
Поведенческие показатели у крыс в тесте ПКЛ после 5-дневного иммобилизационного стресса
| Сроки | Группы животных | Количество пройденных квадратов | Длительность стоек, % | Длительность груминга, % | Количество болюсов | Время в открытых рукавах, % | Время в закрытых рукавах, % |
|---|---|---|---|---|---|---|---|
| До стресса | АД | 26.4 ± 3.4 | 3.5 ± 0.9 | 7.2 ± 1.6 | 2.3 ± 0.3 | 29.4 ± 6.2 | 70.6 ± 6.2 |
| СН | 26.3 ± 3.5 | 4.5 ± 0.8 | 14.6 ± 2.6 | 3.3 ± 0.5# | 16.6 ± 2.5# | 83.3 ± 5.1# | |
| После стресса | АД | 14.2 ± 2.8** | 3.8 ± 0.7 | 31.1 ± 6.9** | 3.2 ± 1.2 | 11.7 ± 6.2* | 88.3 ± 6.2* |
| СН | 13.4 ± 3.0** | 5.4 ± 1.6 | 30.1 ± 3.4** | 2.4 ± 0.8 | 3.7 ± 1.2** | 96.3 ± 3.7** |
У группы АД после 5-дневного иммобилизационного стресса статистически значимо снижается болевая чувствительность (рис. 2), коэффициент предпочтения сладкого раствора (рис. 3), общее время исследования предъявляемых объектов (табл. 5), при этом изменений рейтинга предпочтения нового объекта по сравнению фоновыми значениями не обнаружено (табл. 5).
Рис. 2.
Болевая чувствительность у крыс в норме и после 5-дневного иммобилизационного стресса. По горизонтали – группы животных, по вертикали – латентный период (ЛП) отдергивания хвоста, с. Светлые столбцы – фоновые показатели, темные – данные после 5-дневного иммобилизационного стресса; * – различия статистически значимы по сравнению с фоном р ≤ 0.05. M ± m.
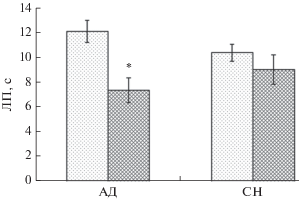
Рис. 3.
Коэффициент предпочтения сладкого раствора в двухбутылочном тесте у крыс в норме и после 5-дневного иммобилизационного стресса. Светлые столбцы – фоновые показатели, темные – данные после 5-дневного иммобилизационного стресса. По горизонтали – группы животных, по вертикали – коэффициент предпочтения. * – различия статистически значимы по сравнению с фоном р ≤ 0.05. M ± m.
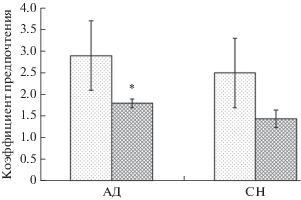
Таблица 5.
Когнитивный тест (предпочтение нового объекта) у крыс до и после 5-дневного иммобилизационного стресса
| Исследовательская активность, с | Рейтинг предпочтения, с | |||
|---|---|---|---|---|
| группы животных: | действие стресса | |||
| до | после | до | после | |
| АД | 83.9 ± 12.1 | 19.9 ± 7.3** | 0.3 ± 0.1 | 0.4 ± 0.1 |
| СН | 62.6 ± 15.9 | 29.2 ± 13.2 | 0.1 ± 0.1 | 0.3 ± 0.1 |
Также показано достоверное уменьшение массы тела у животных группы АД на 3-и и 5-е сутки иммобилизационного стресса (p ≤ 0.05). В группе СН не наблюдается статистически значимого изменения массы.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
На основании анализа поведения животных двух исследованных групп (активные доминанты и неактивные субординанты) можно заключить, что существующие поведенческие различия между данными группами становятся наиболее выраженными после стрессорного воздействия. У животных группы СН по сравнению с группой АД наблюдается более высокий уровень тревожности и более низкий уровень агрессивного поведения. Вероятно, такое состояние животных является следствием хронических агонистических конфликтов в иерархически организованных группах грызунов [25]. Эти результаты согласуются с данными литературы, которые показывают, что хронический социальный стресс оказывает большее влияние на уровень тестостерона и кортикостерона у субординантов по сравнению с доминантами [26].
Животные, занимающие низкий социальный ранг, имеют меньший репродуктивный успех [27]. Вследствие пониженной пролиферации клеток гиппокампа [28] в когнитивных тестах субординанты демонстрируют пониженные показатели рабочей памяти в отличие от доминантов [29].
Активные доминанты менее чувствительны не только к социальному, но и кратковременному физическому стрессору. По нашим данным, в отличие от субординантов у доминантов не наблюдается значимых изменений поведения в тесте ПКЛ после 5-минутного теста неизбегаемого плавания, однако увеличивается продолжительность груминга, что является нормальной смещенной активностью в ответ на стресс у доминантных животных [14].
Длительный иммобилизационный стресс оказывал более значимое влияние на всех животных. Снижение двигательной и исследовательской активности вместе с увеличением тревожности свидетельствует о развитии выраженного стрессорного ответа [16].
При действии неконтролируемого стрессорного воздействия (длительная иммобилизация), “неактивные субординанты” проявляли типичную стратегию “затаивания”, выраженную в значительном снижении двигательной, исследовательской активности, увеличении тревожности и груминга. С помощью такой пассивно-оборонительной стратегии животные проявляют приспособленность к изменяющимся условиям внешней среды, на которое они не могут повлиять, тем самым сберегая ресурсы организма для поддержания гомеостаза.
У животных из группы АД достоверно снижается уровень потребления сладких растворов. Снижение потребления сладких растворов свидетельствует о развитии агедонии [16, 30]. Изменения вышеуказанного параметра в модели стресс-индуцированной агедонии на крысах, по данным литературы, коррелируют с молекулярными, электрофизиологическими, поведенческими и биохимическими сдвигами, характерными для депрессивно-подобных состояний [31]. Также, у животных группы АД снижается мотивация к активному исследованию окружающего пространства (исследование незнакомого предмета), увеличивается болевая чувствительность, достоверно снижается масса тела. Все эти данные свидетельствуют о наличии выраженного депрессивно-подобного состояния и значительных нарушениях функционирования ЦНС у группы АД, в отличие от группы СН.
Таким образом, показано, что в ответ на длительное неконтролируемое воздействие у животных группы АД обнаружены признаки выраженного депрессивно-подобного состояния. В то же время у животных из группы СН наблюдается лишь сильная стрессорная реакция без признаков депрессивно-подобного состояния.
На основании полученных данных можно заключить, что “активные доминанты”, устойчивые к хроническому эмоциональному и кратковременным стрессам, более подвержены действию острого длительного неконтролируемого стрессора с последующим развитием депрессивно-подобного состояния. Напротив, “неактивные субординанты”, неустойчивые к хроническому эмоциональному и кратковременным стрессам, проявляют большую устойчивость к длительному неконтролируемому стрессу без неблагоприятных воздействий на психо-эмоциональное состояние.
Таким образом, зная поведенческий тип животного, возможно спрогнозировать его стрессорную реакцию в ответ на неконтролируемое или контролируемое стрессорное воздействие.
Благодарности: авторы выражают благодарность лаборантам лаборатории № 61, П.П. Бельтюкову, проф. А.А. Карцовой, Бессоновой Елене.
Список литературы
Dhabhar F.S. The short-term stress response – Mother nature’s mechanism for enhancing protection and performance under conditions of threat, challenge, and opportunity. Front Neuroendocrinol. 49: 175–192. 2018.
McIntyre C.K., Roozendaal B. Adrenal stress hormones and enhanced memory for emotionally arousing experiences. CRC Press/Taylor & Francis. 13. 2007.
Виноградова Е.П., Немец В.В., Жуков Д.А. Активная стратегия поведения как фактор риска депрессивноподобных нарушений после хронического умеренного стресса. Журн. высшей нерв. деятельности им. И.П.Павлова. 5: 589–596. 2013. [Vinogradova Е.P., Nemets V.V., Zhukov D.A. Active Coping Style as a Risk Factor of Depressive-Like Disorders after Cronic Mild Stress. Zh. Vyssh. Nerv. Deiat. Im. I. P Pavlova. 5: 589–596. 2013. (In Russ.)].
Selye H. The General-Adaptation-Syndrome. Ann. Rev. 1956.
Mizoguchi K., Kunishita T., Chui D.H. Stress Induces neuronal death in the hippocampus of castrated rats. Neurosci. Lett. 138(1): 157–160. 1992.
Cabib S. Stefano Puglisi-Allegra. Stress, depression and the mesolimbic dopamine system. Psychopharmacology. 128(4): 331–342. 1996.
Jameel M.K., Joshi A.R., Dawane J.A. Effect of various physical stress models on serum cortisol level in wistar rats. JCDR. 8(3): 181–183. 2014.
Gomaa A.M., Galal H., Abou-Elgait A.T. Neuroprotective effects of melatonin administration against chronic immobilization stress in rats. Int. J. Physiol. Pathophysiol. Pharmacol. 9(2): 16–27. 2017.
Kim J.G., Jung H.S., Kim K.J., Min S.S., Yoon B.J. Basal blood corticosterone level is correlated with susceptibility to chronic restraint stress in mice. Neurosci. Lett. 555: 137–142. 2013.
Ahmad A., Rasheed N., Chand K., Maurya R., Banu N., Palit G. Restraint stress-induced central monoaminergic & oxidative changes in rats & their prevention by novel Ocimum sanctum compounds. Indian J. Med. Res. 135(4): 548–554. 2012.
Фролова Г.А., Богданова С.А., Поведенческие эффекты женских половых гормонов различной природы у иммобилизированных самцов белых крыс. Живые и биокосные системы. 14. 2015. [Frolova G.A., Bogdanova S.A., Behavioral effects of female sex hormones of different nature in immobilized male white rats. Live and biokosy systems. 14. 2015. (In Russ.)].
Willner P. Validity, reliability and utility of the chronic mild stress model of depression: A 10-year review and evaluation. Psychopharmacology. 134(4): 319–329. 1997.
Cools A.R., Brachten R., Heeren D., Willemen A., Ellenbroek B. Search after neurobiological profile of individual-specific features of wistar rats. Brain Res. Bull. 24: 49–69. 1990.
Немец В.В., Виноградова Е.П. Стресс и стратегии поведения. Нац. псих. жур. 2(26): 59–72. 2017. [Nemets V.V., Vinogradova E.P. Stress and neurobiology of coping styles. Nat. Psychol. J. 2(26): 59–72. 2017 (In Russ.)].
de Boer S.F., Buwalda B., Koolhaas J.M. Untangling the neurobiology of coping styles in rodents: towards neural mechanisms underlying individual differences in disease susceptibility. Neurosci. Biobehav. Rev. 74: 401–422. 2017.
Жуков Д.А. Психогенетика стресса. Поведенческие и эндокринные корреляты генетических детерминант стресс-реактивности при неконтролируемой ситуации. СПб. 1997. [Zhukov D.A. Psichogenetika stressa. Povedenchskie i endocrinnie korrelyati geneticheskich determinant stress-reactivnosti pri nekontroliruemoy situazii. [Psychogenetics of stress. Behavioral and endocrine correlates of genetic determinants of stress reactivity in an uncontrolled situation]. Sankt-Petersburg. 1997].
Жуков Д.А., Виноградова Е.П., Вековищева О.Ю. Крысы с пассивной стратегией приспособления обладают средним, а не низшим социальным рангом. Журн. высш. нервн. деят. 52(2): 175–182. 2002. [Zhukov D.A., Vinogradova K.P., Vekovisheva O.U. Rats with passive coping style have a medium rather than lower social rank. Zh. Vyssh. Nerv. Deiat. Im I. P. Pavlov. 52(2): 175–182. 2002. (In Russ.)].
Taylor K., Gordon N., Langley G., Higgins W. Estimates for worldwide laboratory animal use in 2005. ATLA. 36(3): 327–342. 2008.
Griebel G., Holmes A. 50 years of hurdles and hope in anxiolytic drug discovery. Nat. Rev. Drug Discov. 12(9): 667–687. 2013.
Mathiasen J.R., Di Camillo A. Novel object recognition in the rat: A facile assay for cognitive function. Curr. Protoc. Pharmacol. 5(5): 59. 2010.
Немец В.В., Николаев А.И., Пшенов А.Б., Соболев В.Е., Виноградова Е.П. Новая модификация аппарата “челночная камера”. Лаб. животные для научных исследований. 1: 92–99. 2018. [Nemets V.V., Nikolaev A.I., Pshenov A.B., Sobolev V.E., Vinogradova E.P. A new modification of the shuttle box device. Lab. Animals for Scient. Res. 1: 92–99. 2018. (In Russ.)].
Иванов Д.Г., Семенов Н.А., Зайцева М.С. Методика определения социального статуса самцов крыс в триадах. Успехи соврем. естествознания. 9(1): 43–46. 2014. [Ivanov D.G., Semenov N.A., Zaitseva M.S. Methods for determining the social status of male rats in triads. Suc. Mod. Sci. 9(1): 43–46. 2014. (In Russ.)].
Пошивалов В.П. Этологический атлас для фармакологических исследований на лабораторных грызунах. Деп. В ВИНИТИ. 3164–3178. 1978. [Poshivalov V.P. Ethological atlas for pharmacological research in laboratory rodents. Dep. VINITI. 3164–3178. 1978. (In Russ.)].
Lewis J.M., Hori T.S., Rise M.L., Walsh P.J. Transcriptome responses to heat stress in the nucleated red blood cells of the rainbow trout (Oncorhynchus mykiss). Physiol. Genomics. 42: 361–373. 2010.
Blanchard D.C., Sakai R.R., McEwen B., Weiss S.M., Blanchard R.J. Subordination Stress: Behavioral, Brain, and Neuroendocrine Correlates. Behav. Brain Res. 58(1–2): 113–121. 1993.
Hardy M.P., Sottas C.M., Ge R., McKittrick C.R. Trends of reproductive hormones in male rats during psychosocial stress: Role of glucocorticoid metabolism in behavioral dominance. Biol. Reprod. 67(6): 1750–1755. 2002.
Cart W.J., Kimmel K.R., Anthony S.L., Schlocker D.E. Female rats prefer to mate with dominant rather than subordinant males. Bull. Psychonormic. Soc. 20. 89–91. 1982.
Hoshaw B.A., Evans J.C., Mueller B., Valentino R.J., Lucki I. Social Competition in Rats: Cell Proliferation and Behavior. Behav. Brain Res. 175(2): 343–351. 2006.
Blanchard R.J., McKittrick C.R., Blanchard D.C. Animal models of social stress: Effects on behavior and brain neurochemical systems. Physiol. Behav. 73(3): 261–271. 2001.
Grønli J., Murison R., Bjorvatn B., Sørensen E., Portas C.M., Ursin R. Chronic mild stress affects sucrose intake and sleep in rats. Behav. Brain Res. 150(1–2): 139–147. 2004.
Moreau J.L., Scherschlicht R., Jenck F., Martin J.R. Chronic mild stress-induced anhedonia model of depression; sleep abnormalities and curative effects of electroshock treatment. Behav. Pharmacol. 6(7): 682–687. 1995.
Дополнительные материалы отсутствуют.
Инструменты
Российский физиологический журнал им. И.М. Сеченова