

Российский физиологический журнал им. И.М. Сеченова, 2020, T. 106, № 1, стр. 121-132
Методы определения социальных рангов животных в малых группах
И. С. Матюлько 1, 2, *, Е. П. Муртазина 1, Б. В. Журавлев 1
1 НИИ нормальной физиологии им. П.К. Анохина
Москва, Россия
2 Национальный исследовательский университет “Высшая школа экономики”
Москва, Россия
* E-mail: irinamatulko@gmail.com
Поступила в редакцию 04.07.2019
После доработки 16.10.2019
Принята к публикации 21.10.2019
Аннотация
Представлены методы определения социальных рангов крыс, основанные на количественном анализе социометрических матриц, построенных с использованием данных об антагонистических отношениях животных, возникающих во время конкуренции за получение подкрепления. Проведены сравнение и оценка существующих методов, а также предложена новая экспериментальная модель изучения доминантного поведения крыс в малых группах в лабораторных условиях с учетом питьевой активности каждой особи и диадических конкурентных взаимодействий. Предложены алгоритмы анализа полученных данных на основе построения гистограмм распределения количеств и длительностей питьевых актов у крыс в группах, учета всех диадических соотношений побед и поражений особей в борьбе за доступ к поилке, а также оценки приоритетности особей в удовлетворении питьевой мотивации во временной динамике экспериментальных сессий. Результаты апробации предложенных методов показали, что их комплексное использование позволяет провести точную оценку социального ранга каждой крысы, а также проследить динамику становления, стабильности и пластичности иерархической социальной организации групп на длительном протяжении их существования. Таким образом, применение данной комплексной методики может позволить выявлять взаимосвязи иерархической структуры групп социальных животных и индивидуальных социальных рангов особей с различными физиологическими, фармакологическими, средовыми и другими факторами.
Многие виды животных живут в группах, образуя сложные системы, внутри которых осуществляется адаптация к окружающей среде, охота, миграция, размножение и эволюция. В процессе взаимодействия особей внутри социальных групп происходит формирование многоуровневой иерархической организации, в основе которой лежат определенные физиологические и нейрохимические процессы, связанные с участием нейроэндокринной, дофаминергической и серотонинергической систем в формировании доминантной структуры группы за счет регуляции уровня агрессии, устойчивости к стрессу и кооперативного поведения [1, 2]. Показано, что активность многочисленных отделов головного мозга, таких как миндалина, гиппокамп, стриатум, префронтальная и париетальная корковые области участвуют в формировании и восприятии доминантности, а также различных видов зоосоциального поведения [3, 4].
Иерархическая структура подразумевает распределение обязанностей и ролей между членами группы, каждый из которых обладает определенным социальным рангом (иерархическим статусом), установившимся в результате социального опыта взаимодействий – как антагонистических, так и кооперативных. Кооперативные взаимоотношения, как правило, направлены на достижение общей цели и зачастую характерны для таких процессов, как охота, защита от врагов, забота о потомстве. Под антагонистическими взаимодействиями понимается конкурентное и агрессивное поведение особей по отношению друг к другу, целью которого является установление доминантности и определяет ранг особи в группе [4]. Успешные особи (победители) являются доминантами, в то время как менее успешные (проигравшие) – субординантами. В случае, если антагонистические взаимоотношения основаны исключительно на конкурентных способностях, наблюдается корреляция социального ранга с физическими характеристиками особей, а сам ранг обладает свойством транзитивности: если особь А доминирует над особью Б, а та в свою очередь над особью С, то особь А также доминирует над С [5]. Однако гораздо чаще встречаются ситуации, когда иерархическая структура характеризуется нелинейностью и лабильностью, а особи формируют не диады (доминант–субординант), а триады, и даже тетрады, в которых позиции победителей и проигравших могут меняться для различных взаимодействующих между собой животных. По этой причине оценка доминантности особей и иерархической структуры групп зачастую оказывается затруднительной и требует комплексного подхода, включающего создание экспериментальных установок, обеспечивающих объективное наблюдение за поведением животных и регистрацию паттернов взаимодействий, а также разработку программных средств для последующего количественного анализа многосторонних взаимоотношений.
Для исследования доминантности животных в лабораторных условиях применяются поведенческие модели, подразумевающие контакт между животными из разных групп (модель конфликта) или же из одной. В последнем случае используют модели конкуренции, основанные на применении естественных физиологических подкрепляющих стимулов, таких как привлекательная для животных пища или сладкая вода. Для повышения мотивации животных до проведения эксперимента их ограничивают в доступе к пище или воде.
Одним из тестов, используемых для изучения диадических отношений, а именно уровня агрессии и доминантности, является “Стандартный тест оппонентов” (Standard Opponent Test). Тестируемое животное помещается в нейтральную среду с незнакомой особью своего вида – оппонентом. Далее проводится количественная оценка поведенческих актов обнюхивания тела и гениталий, преследования, встряхивания хвостом, количества укусов [6].
Разновидностью теста, не подразумевающего наличие конфликта между животными, является тест “Труба” [7], в котором две особи одного пола помещаются на противоположных концах прозрачной трубы. В процессе проявления исследовательской активности животные начинают двигаться по направлению к центру до момента встречи друг с другом. Доминантной считается та особь, которая при встрече будет иметь склонность двигаться вперед, в то время как подчиненная особь будет отступать назад до тех пор, пока не будет вытолкнута первой.
Для исследования социальных рангов крыс также используется “Система открытой норы” (Visible Burrow System) [8]. Экспериментальная установка состоит из нескольких камер, соединенных коридорами. Измеряемыми параметрами для выявления доминантности являются время, проведенное вне установки, ранение животных, количество конфликтов между особями в диадах и потеря массы тела. В дополнение к поведенческим параметрам, также измеряются уровни кортикостерона, кортизола [1], тестостерона [2] и окситоцина в плазме крови или слюне [9].
Исследование социального статуса крыс в триадах в условиях конкуренции животных за доступ к поилке после 24-часовой питьевой депривации представлено в работе Д.Г. Иванова с соавт. [10]. В экспериментальную камеру, по центру которой находилась поилка с водой, помещали трех особей и в течение 8 мин регистрировали латентное время первого подхода к поилке, общее число подходов, а также длительность их питьевого поведения. Для каждой крысы также регистрировали количество контактов, когда особь отгоняла других от поилки (А) и число раз, когда другие особи отгоняли ее (Б). Социальный статус рассчитывался как вероятность доминирующего поведения (ВДП), равную отношению числа доминирования крысы над другими к сумме числа событий, когда животное отгоняло других и когда отгоняли его (ВДП = А/(А + Б)). Доминантами считались животные, вероятность доминирующего поведения которых превышала или была равна 0.65; субдоминантами считались особи со значением индекса между 0.35 и 0.65; субординанты характеризовались значениями ВДП меньшими или равными 0.35.
П.Д. Шабанов и А.А. Лебедев [11] в своей работе исследовали модель конкурентных взаимоотношений крыс (группы по 8–10 особей) за доступ к поилке. После суточной питьевой депривации животных помещали в экспериментальную клетку, где им было необходимо достигнуть поилки, находящейся на расстоянии 22 см от пола и соединенной с полом клетки наклонной лестницей. Конкурентоспособность (Кп) крыс определяли как соотношение времени питья (в минутах) из поилки (Тп) к общему времени эксперимента (То) и выражали через индекс Кп (Кп = Тп/То). Более высокие значения коэффициента соответствовали более высокой конкурентоспособности крысы, т.е. большей успешности в доступе к воде.
Данные, полученные в результате наблюдений за взаимоотношениями между животными в группах, обрабатываются с помощью математических методов, включая построение и анализ социометрических матриц для выявления иерархической структуры и доминантности особей [12, 13]. Часто в работах используются индексы линейности [14], отражающие, насколько внутригрупповые отношения соответствуют линейному типу иерархии. Однако при использовании подобных подходов важно учитывать, что организация сообществ зачастую является нелинейной [7].
Исходя из вышеизложенного, разработка новых подходов к исследованию индивидуальных социальных рангов (доминантности) особей и иерархической структуры групп млекопитающих, а также создание новых экспериментальных лабораторных комплексов и их программного обеспечения является актуальной современной задачей физиологии и социоэтологии.
Целью настоящей работы была разработка и апробация лабораторных методов определения социальных рангов особей и иерархической структуры в малых группах крыс в процессе конкуренции за подкрепление на фоне доминирующего мотивационного состояния.
МЕТОДЫ ИССЛЕДОВАНИЯ
Эксперименты проводили на самцах крыс линии Вистар (n = 16) с исходной массой на момент начала исследования 150–200 г. Крысы содержались в группах по 5–6 особей (3 группы) в условиях вивария при температуре 22°С и регулярном световом цикле (включение света в 8:00, выключение в 22:00). На протяжении всего периода исследования животные находились в условиях свободного доступа к воде и корму (“Полнорационный комбикорм для лабораторных животных ПК 120-1240” в виде экструдированных гранул, ЗАО "Гатчинский ККЗ”, Россия) за исключением дней, предшествующих тестированию доминантности. Содержание пищевых веществ, витаминов и минералов в корме соответствовало международным стандартам и позволяло полностью обеспечить все физиологические потребности животных. Исследование было выполнено с соблюдением принципов гуманности, изложенных в директиве европейского сообщества (86/609/ЕС) и в соответствии с принципами Базельской декларации и требованиями, указанными в Директиве 2010/63/EU Европейского парламента и Совета Европейского союза от 22.09.2010 г. по охране животных, используемых в научных целях. Проведение эксперимента было одобрено этическим комитетом ФГБНУ “НИИ нормальной физиологии им. П.К. Анохина”.
Нами был создан экспериментальный комплекс для исследования иерархического статуса крыс в малых группах, схожий по конструкции с использованным П.Д. Шабановым и А.А. Лебедевым [11]. Установка включала камеру (52 × 30 × 19 см) с прозрачными стенами, к одной из которых прилегала приподнятая платформа в форме куба (10 × 10 см), на которую могла забраться только одна крыса. Над платформой располагался кончик поилки, закрепленной на наружной стороне камеры на регулируемой высоте, так, чтобы крысы могли достичь поилки в вертикальной стойке.
Для дистанционного наблюдения за крысами, записи и регистрации поведенческих паттернов над экспериментальным боксом была установлена веб-камера. Каждой крысе присваивался свой номер, который наносился маркером на спинку.
Перед экспериментальной сессией животные подвергались суточной питьевой депривации. Крыс одной группы помещали в экспериментальную камеру на 30 мин, где они вступали в конкуренцию за доступ поилке, поочередно забираясь на платформу и сталкивая друг друга.
Поведенческие паттерны регистрировали дистанционно на компьютере с помощью разработанной нами программы “Этограф”. Посредством нажатия на клавишу, соответствующую номеру крысы, экспериментатор отмечал, какая именно особь забралась на платформу. Если крыса слезала сама, за ее номером следовал ноль, если же ее сталкивала другая особь – отмечали номер столкнувшего животного. После окончания наблюдения в программе “Этограф” автоматически создавался файл, содержащий данные об эксперименте (дата и время проведения, количество животных, масса тела, возраст и пол каждой крысы), суммарное количество (N) и общая длительность (T) питьевых актов для каждой крысы, а также полная временная динамика (время начала и длительность) зарегистрированных событий.
Тестирование доминантности и оценку иерархической организации групп проводили один раз в неделю на протяжении 9 недель, начиная с момента получения крыс из питомника и формирования групп с использованием процедуры рандомизации.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ И ИХ ОБСУЖДЕНИЕ
На основе данных зарегистрированной поведенческой активности строили гистограммы распределения количеств и длительностей питьевых актов всех крыс внутри группы за экспериментальную сессию. На рис. 1 представлен пример гистограмм, построенных для всех крыс одной из экспериментальных групп.
Рис. 1.
Пример гистограмм распределения количеств и длительностей питьевых актов всех животных из одной группы в течение 30-минутной экспериментальной сессии. Данные представлены в процентах от общей активности (N %) и общего времени питья (T %) за данную сессию. Fig. 1. Histograms of the distribution of the number and duration of drinking patterns of all animals of one group within a 30-min experimental session. The data are presented as a percentage of the total activity (N %) and total drinking time (T %) within a given session.
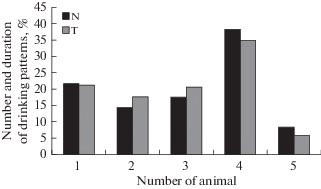
Расчет доли времени питья Т по каждой крысе (в % от общего времени сессии) совпадает с определением коэффициента конкурентоспособности, предложенным П.Д. Шабановым и А.А. Лебедевым [11]. В нашей работе мы предлагаем учитывать не только суммарную длительность питья каждой особи, но и количество ее питьевых актов N (в % от общего количества питьевых актов всех крыс в группе), так как показатель Т напрямую не отражает конкурентные взаимодействия между особями.
В дополнение к статистическим показателям питьевой активности мы предлагаем определять доминантность крыс, рассчитывая соотношение числа побед и поражений во всех диадах с учетом результативности каждой крысы относительно суммарной активности группы. На основе данных динамики поведенческих паттернов составляли социометрические матрицы, содержащие количество побед и поражений каждой особи в результате ее диадических взаимодействий с каждым из представителей группы. В таблице также учитывались акты, когда животное без борьбы залезало на пустую платформу и самостоятельно покидало ее. Пример социометрической матрицы, построенной на основе результатов одной экспериментальной сессии, приведен в табл. 1.
Таблица 1.
Пример социометрической матрицы, содержащей количество побед и поражений каждой крысы в конкуренции за поилку после суточной питьевой депривации, а также количество актов, когда животное без борьбы залезло на пустую платформу или покинуло ее Table 1. Sociometric matrix containing the total number of wins and losses of each rat in the competition for the access to a drinking bowl after 24-h water deprivation and the number of events when animals climbed to or left the platform without competition.
| Номер крысы (i) Number of animal (i) |
1 | 2 | 3 | 4 | 5 | W | O1i | IAi = Wi + O1i | HS |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 4 | 1 | 5 | 1 | 11 | 39 | 50 | 0.40 | |
| 2 | 2 | 0 | 0 | 1 | 3 | 30 | 33 | 0.07 | |
| 3 | 1 | 1 | 3 | 0 | 5 | 35 | 40 | 0.29 | |
| 4 | 3 | 1 | 2 | 0 | 6 | 81 | 87 | 0.25 | |
| 5 | 0 | 0 | 0 | 1 | 1 | 18 | 19 | 0.04 | |
| L | 6 | 6 | 3 | 9 | 2 | GA = ∑ IAi = 229 | |||
| O2i | 44 | 27 | 37 | 78 | 17 | ||||
| IAi = Li + O2i | 50 | 33 | 40 | 87 | 19 | ||||
Для определения иерархического статуса каждой особи нами был разработан индекс HS, рассчитываемый по следующей формуле:
Индекс HS может принимать любые значения от 0 до 1, а сумма значений HS всех животных в группе внутри одной сессии равняется единице. Иерархическая организация группы определяется путем сопоставления рангов. Крысы, характеризующиеся наибольшей величиной индекса HS, являются доминантными, в то время как особи с меньшим значениям ранга занимают подчиненное положение. При близких значениях индекса у двух особей, оценивается результат их диадического взаимодействия, а именно отношение их побед друг над другом (рис. 2).
Рис. 2.
Схематичное представление иерархической организации группы животных на основе данных Таблицы 1. В каждой ячейке отображен номер животного и соответствующее значение социального ранга HS. Стрелки направлены в сторону от “победителя” к “проигравшему” с указанием соотношения побед к поражениям. Fig. 2. Schematic presentation of the hierarchical organization of the group according to the data shown in Table 1. Each cell contains the number of the animal and a corresponding value of social rank (HS). The arrows point from the “winner” toward the “loser” with the win to loss ratio indicated above.
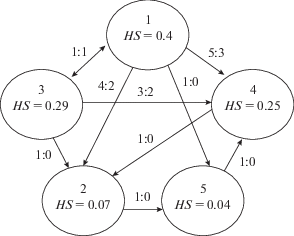
Индекс HS характеризуется высокой лабильностью и обладает чувствительностью к изменению внешних факторов, включая социальный контекст и обучение. Предполагается, что после становления и в отсутствии внешних воздействий социальная структура группы относительно стабильна и изменения индексов у животных одной группы незначительны.
В отличие от гистограмм социометрический метод определения доминантного статуса животных более предпочтителен, так как индекс HS является нормированным показателем и позволяет объективно оценить ранги особей на основе их антагонистических отношений с другими субъектами в диадах, в то время как гистограммы распределения количества питьевых актов и их длительностей не оценивают взаимоотношения между особями, а отражают суммарную способность удовлетворения жажды каждым индивидуумом за все время экспериментальной сессии, аналогично показателю конкурентоспособности, описанному в работе П.Д. Шабанова и А.А. Лебедева. Более того, зачастую доминантность животного, рассчитанная по показателю количеств питьевых актов, не всегда совпадает с ее оценкой по длительности питья и наоборот. Индекс HS позволяет оценить ранг животного в группе на основе соотношения количеств побед и поражений с учетом доли его индивидуальной питьевой активности в общей активности группы за 30-минутный период исследования.
Сопоставление результатов гистограмм распределения количества и длительностей питьевых актов и значений индекса HS, рассчитанного по социометрической матрице, свидетельствует о том, что конкурентоспособность животного, определенная по гистограммам, может быть не тождественна социальному рангу. Например, по приведенным выше данным видно, что питьевая активность крысы 4 была ваше остальных (рис. 1), в то время как результаты социометрической матрицы (табл. 1) свидетельствуют о том, что эта особь удовлетворяла питьевую мотивацию преимущественно избегая антагонистических контактов (т.е. залезая на пустую платформу). Самый высокий ранг в группе был у крысы 1 как результат преобладания ее побед над всеми другими особями группы.
Предложенная нами экспериментальная модель отличается от предложенной Д.Г. Ивановым по ряду параметров. В работе Д.Г. Иванова наблюдения проводились только за группами из трех животных, а их время составляло 8 мин, что, по нашему мнению, недостаточно для того, чтобы полноценно оценить динамику питьевого и конкурентного поведения особей. Предложенный Д.Г. Ивановым индекс ВДП (вероятность доминантного поведения) имеет один существенный недостаток: он может принимать равные значения у особей, проявляющих разную суммарную активность. Например, значение индекса ВДП = 0.66, определенное автором как доминантное, может быть присвоено как крысе с высокой активность (соотношение 8 побед особи к ее 12 контактам со всеми другими крысами), так и крысе с низкой активностью (соотношение 2/3 соответственно). Предложенный нами индекс позволяет избежать этой проблемы за счет учета доли индивидуальной результативности каждой крысы в суммарной активности группы.
Существуют разнообразные методы определения социальных рангов особей в группах [15], основанные на построении социометрических матриц, которые содержат количественные значения побед и поражений между всеми возможными парами особей группы. T.H. Clutton-Brock с соавт. [16] изучали поведение самцов благородных оленей во время поединков в естественной среде обитания и фиксировали соотношения побед и поражений. На основе полученных данных рассчитывали индекс CBI (Clutton-Brock’s Index), учитывающий прямые и непрямые победы и поражения. Под прямыми победами в данном случае понимается количество побежденных данным самцом особей, в то время как непрямые победы определяются как общее количество особей, побежденных теми, над которыми одержал победу данный самец. Таким образом, непрямые победы могут характеризоваться, согласно работе M.C. Appleby [5], как линейные: самец, доминирующий над другими самцами, также доминирует над теми, над кем доминируют им побежденные. Это может быть выражено в виде схемы: A > B > C, A > C. Расчет индекса CBI проводится согласно следующей формуле [17]:
Однако недостатком данного индекса является то, что он не учитывает количество побед и поражений, а проводит только качественную оценку диадических взаимоотношений внутри каждой пары особей (над сколькими особями доминирует данный самец и сколько особей доминирует над ним), рассчитывая количество животных, но не количество антагонистических взаимодействий. По этой причине индекс CBI больше подходит для изучения взаимоотношений внутри больших групп животных в естественных условиях на уровне целой популяции. Предложенный нами индекс HS обладает бóльшей силой при изучении поведения лабораторных животных в малых группах, существующих совместно на протяжении длительного периода времени и характеризующихся многократными диадическими антагонистическими контактами.
H.A David [18, 19] предложил индекс DS (David’s Score), который учитывает количество побед и поражений особей посредством расчета доли диадических взаимодействий каждого субъекта группы. В социометрическую таблицу вносятся значения соотношений побед особи к сумме побед и поражений. Для расчета используются взвешенные доли побед и поражений. Индекс рассчитывается по следующей формуле:
где w – суммарная доля побед данной особи; w2 – сумма взвешенных долей побед индивидуумов, побежденных данной особью; l – суммарная доля проигрышей; l2 – сумма взвешенных долей проигрышей индивидуумов, победивших данную особь. Данные для расчета показателей w2 и l2 получаются в результате умножения долей побед и поражений каждого противника на соответствующие значения w и l [17].Значения индекса DS могут принимать как положительное, так и отрицательное значения. Недостатком данного метода является то, что в случае отсутствия активности животного (когда абсолютное значение его активности равно нулю), индекс DS может превзойти индексы с отрицательным значением, что приведет к неправильной интерпретации результатов. К тому же, не учитывается общая активность животного помимо непосредственного взаимодействия с другими особями. В отличие от индекса DS, предложенный нами индекс HS не принимает отрицательные значения и позволяет выявить животных, чья активность была равна нулю.
В дополнение к статистическому и социометрическому методам оценки рангов нами предлагается проводить дополнительный анализ динамики питьевой активности животных на протяжении сессии, который позволяет выявить более ранний доступ особи к поилке по сравнению с другими крысами и индивидуальный временной паттерн ее активности в течение эксперимента (рис. 3).
Рис. 3.
Пример динамик паттернов питьевых актов всех крыс одной из групп в течение 30-минутной экспериментальной сессии. По оси абсцисс – время экспериментальной сессии (мин), по оси ординат – длительность питьевых актов каждой крысы. Fig. 3. The dynamics of drinking patterns of all rats of one group within a 30-min experimental session. The X axis shows the time of experimental session (min); the Y axis shows the duration of each rat’s drinking patterns.
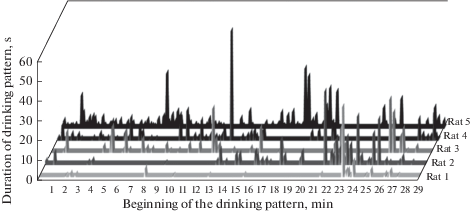
На рис. 3 представлен график, отображающий индивидуальные динамики моментов осуществления и длительностей питьевых актов крысами одной из групп за 30-минутную экспериментальную сессию. Он иллюстрирует, что животные удовлетворяли питьевую мотивацию в разные периоды времени – раньше или позже других особей. Так, например, пятая крыса смогла попить лишь в конце эксперимента, в то время как первая и четвертая крысы достигали поилки как в начале, так и на протяжении всей сессии. Третья крыса была наиболее активна во второй половине сессии, а вторая особь демонстрировала эпизодическую активность на протяжении всего опыта. Такой анализ позволяет получить дополнительные качественные критерии для уточнения социальной позиции животного в группе, особенно при равных значениях ранее рассчитанных социальных рангов. Индекс HS и гистограммы распределения количеств и длительностей питьевых актов являются интегральными показателями, отражающими социальное положение каждой крысы в целом и общую питьевую активность животных, в то время как динамические характеристики питьевых актов внутри сессии позволяют оценить индивидуальный питьевой паттерн каждой крысы.
Более раннее удовлетворение мотивации жажды можно также выявить с помощью построения кумулятивных кривых длительностей доступа к поилке различными крысами в процессе эксперимента, а проведение по ним регрессионного анализа позволяет получить количественные характеристики паттернов питьевой активности крыс и сопоставить их значения между различными животными (рис. 4).
Рис. 4.
Пример кумулятивных кривых длительностей питьевых актов в течение 30-минутного экспериментального сеанса у крыс одной из групп. По оси абсцисс – время экспериментальной сессии (мин), по оси ординат – текущие значения накопленной суммы длительностей осуществленных ранее питьевых актов. Для двух крыс представлены тренды и формулы с вычисленными характеристиками линейного регрессионного анализа. y – длительность питьевых актов, накопленная в течение экспериментальной сессии; x – время эксперимента; R2 – величина достоверности аппроксимации. Fig. 4. Cumulative curves representing the duration of drinking patterns of all animals of one group within a 30-min experimental session. The abscissa shows the time of experimental session (min); the ordinate shows current values of accumulated sum of drinking pattern durations. The trends and formulas with the parameters of linear regression are calculated and presented for two rats. y – the duration of drinking patterns, accumulated throughout the experimental session; x – time of the experiment; R2 – reliability index.
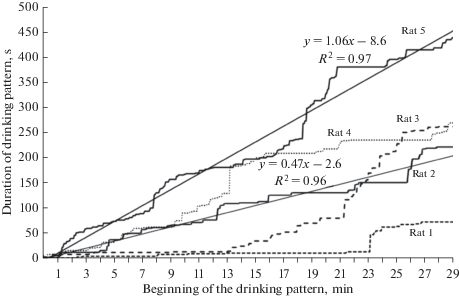
По кумулятивным графикам и показателям линейной регрессии уi = Аiх + Вi (Аi – наклон, Вi – пересечение с осью ординат) можно определить интенсивность и начало момента удовлетворения крысами питьевой мотивации. Так, на рис. 4 можно видеть, что четвертая крыса достигала поилки быстрее остальных и была активна на всем протяжении экспериментальной сессии, а временные промежутки, в которые эта крыса была неактивна, возникали лишь в конце наблюдения. Кумулятивные кривые для крыс 1 и 2 иллюстрируют меньшую для них доступность к осуществлению питьевых актов, что прослеживается на всем протяжении 30-минутной сессии. Третья и пятая крысы, как демонстрируют их кривые на рис. 4, смогли начать удовлетворять питьевую мотивацию в конкурентной среде своей группы только в середине или конце эксперимента соответственно.
Отличительной особенностью и преимуществом нашей модели является комплексный подход, позволяющий определить социальный статус каждого животного не только в данный период времени, но и проследить динамику формирования, стабильность или лабильность социальных рангов всех особей, которые содержатся совместно в течение длительного периода времени и могут при этом подвергаться различным экспериментальным воздействиям.
Таким образом, продемонстрировано, что разработанный нами метод является комплексным и не только учитывает и объединяет показатели, предложенные другими авторами, но и позволяет выявить индивидуальные характеристики питьевой и конкурентной активности каждой особи посредством построения графиков динамики активности животных за каждую экспериментальную сессию, и проследить процесс становления, а также оценить устойчивость иерархической организации групп в течение длительного периода исследования. Он может быть успешно применен в исследованиях нейробиологических коррелятов социальной структуры в группах животных, а также для выявления эффектов фармакологических агентов на социальное поведение.
Известно, что активация префронтальной коры, являющейся основным центром исполнительного контроля, непосредственно связана с обработкой информации о социальной иерархии [20]. Хирургическое разрушение этой области вызывает изменения социального поведения животного и приводит к снижению его иерархического статуса. Ранее было показано [21], что крысы, у которых проводилось разрушение области медиальной префронтальной коры, проявляют субординантное поведение в процессе антагонистических взаимодействий и характеризуются снижением мотивации в процессе конкуренции за подкрепление. Разрушение передней поясной коры, активность которой связана с соревновательной способностью [21], приводит к снижению интереса крысы к другим особям группы и ухудшению социальной памяти, а также нарушению процесса принятия решений в группе [22]. Важный вклад в модуляцию доминантного поведения у животных вносит эффективность синаптических связей в медиальной префронтальной коре [23], связанная с уровнем экспрессии генов Ras или субъединицы GluR4 глутаматных AMPA-рецепторов. В частности, показано, что воздействия, приводящие к увеличению или снижению экспрессии этих генов, сопровождались ростом или падением индексов доминантности соответственно. Фармакологическое ингибирование дофаминовых D2-рецепторов введением антагониста сульпирида приводит к снижению социального ранга у доминантных крыс. С другой стороны, введение антагониста дофаминовых D2-рецепторов животным с более низким социальным рангом не оказывает влияние на социальный статус особей [24].
ЗАКЛЮЧЕНИЕ
Представленные в данной работе методы и подходы к определению социальных рангов лабораторных животных, содержащихся в группах в течение длительного периода времени, позволяют проводить оценку иерархической структуры малых групп животных в лабораторных условиях с учетом индивидуальной активности каждой особи и временной динамики изменения паттернов питьевого поведения. Предложенный нормированный показатель социального ранга HS позволяет учитывать диадические взаимодействия крыс и их активность. Гистограммы распределения количеств и длительностей питьевых актов, паттерны питьевого поведения каждой крысы в группе, а также кумулятивные кривые длительностей питьевых актов позволяют проанализировать конкурентное поведение и индивидуальные характеристики каждой крысы как внутри одной сессии, так и на протяжении длительного периода. Разработанные модели и методы могут быть применены для изучения изменений социальной организации групп при различных этологических и нейрохимических воздействиях.
Список литературы
Creel S. Dominance, aggression, and glucocorticoid levels in social carnivores. J. Mammal. 86(2): 255–64. 2005.
Selmanoff M.K., Goldman B.D., Ginsburg B.E. Serum testosterone, agonistic behavior, and dominance in inbred strains of mice. Horm. Behav. 8(1): 107–19. 1977.
Watanabe N., Yamamoto M. Neural mechanisms of social dominance. Front. Neurosci. 9: 154. 2015.
Hobson E.A., DeDeo S. Social Feedback and the Emergence of Rank in Animal Society. PLoS Comput. Biol. 11(9): e1004411. 2015.
Appleby M.C. The probability of linearity in hierarchies. Anim. Behav. 31(2): 600–608. 1983.
Al-Maliki S., Brain P.F. Effects of food deprivation on fighting behaviour in standard opponent tests by male and female “TO” strain albino mice. Anim. Behav. 27: 562–566. 1979.
Lindzey G., Winston H., Manosevitz M. Social Dominance in Inbred Mouse Strains. Nature. 191(4787): 474–476. 1961.
Blanchard D.C., Spencer R.L., Weiss S.M., Blanchard R.J., McEwen B., Sakai R.R. Visible burrow system as a model of chronic social stress: behavioral and neuroendocrine correlates. Psychoneuroendocrinology. 20(2): 117–134. 1995.
Lee W., Hiura L., Yang E., Broekman K.A., Ophir A.G., Curley J. Social status in mouse social hierarchies is associated with variation in oxytocin and vasopressin 1a receptor densities. bioRxiv (online). 2019.
Иванов Д.Г., Семёнов Н.А., Зайцева М.С. Методика определения социального статуса самцов крыс в триадах. Успехи современ естествознания. 9(1): 43–46. 2014. [Ivanov D.G., Semenov N.A., Zaitseva M.S. Methods for determining the social status of male rats in triads. Suc. Mod. Sci. 9(1): 43–46. 2014.(In Russ)].
Шабанов П.Д., Лебедев А.А. Зоосоциальное поведение крыс. Обзоры по клинической фармакологии и лекарственной терапии. 5: 31. 2007. [Shabanov P.D., Lebedev A.A. Zoosocial behavior of rats. Rev. Clin. Pharmacol. Drug. Therapy. 5: 31. 2007. (In Russ)].
Altmann J. Observational study of behavior: sampling methods. Behaviour. 49(3): 227–267. 1974.
Norscia I., Palagi E. The socio-matrix reloaded: from hierarchy to dominance profile in wild lemurs. Peer. J. 3: e729. 2015.
Landau H. G. On dominance relations and the structure of animal societies: I. Effect of inherent characteristics. Bull. Math. Biophys. 13(1): 1–19. 1951.
Bayly K.L., Evans C.S., Taylor A. Measuring social structure: A comparison of eight dominance indices. Behav. Processes. 73(1): 1–12. 2006.
Clutton-Brock T.H., Albon S.D. The roaring of red deer and the evolution of honest advertisement. Behaviour. 69(3–4): 145–170. 1979.
Bang A., Deshpande S., Sumana A., Gadagkar R. Choosing an appropriate index to construct dominance hierarchies in animal societies: a comparison of three indices. Anim. Behav. 79(3): 631–636. 2010.
David H.A. Ranking from Unbalanced Paired-Comparison Data. Biometrika. 74(2): 432–6. 1987.
David H.A. The method of paired comparisons. New York, Hafner Pub. Co. 1963.
Wang F., Kessels H.W., Hu H. The mouse that roared: neural mechanisms of social hierarchy. Trends Neurosci. 37(11):674–682. 2014.
Hillman K.L., Bilkey D.K. Neural encoding of competitive effort in the anterior cingulate cortex. Nat. Neurosci. 15(9): 1290–1297. 2012.
Rudebeck P.H., Walton M.E., Millette B.H.P., Shirley E., Rushworth M.F.S., Bannerman D.M. Distinct contributions of frontal areas to emotion and social behaviour in the rat. Eur. J. Neurosci. 26(8): 2315–2326. 2007.
Wang F., Zhu J., Zhu H., Zhang Q., Lin Z., Hu H. Bidirectional control of social hierarchy by synaptic efficacy in medial prefrontal cortex. Science. 334(6056): 693–697. 2011.
Yamaguchi Y., Lee Y-A., Kato A., Jas E., Goto Y. The Roles of Dopamine D2 Receptor in the Social Hierarchy of Rodents and Primates. Sci. Rep. 7: 43348. 2017.
Дополнительные материалы отсутствуют.
Инструменты
Российский физиологический журнал им. И.М. Сеченова