

Российский физиологический журнал им. И.М. Сеченова, 2021, T. 107, № 10, стр. 1264-1273
Реакции нейронов соматосенсорной коры на стимуляцию области вибрисс у крыс линии WAG/Rij, генетически предрасположенных к абсанс-эпилепсии
Д. А. Цветаева 1, В. В. Раевский 1, *
1 Институт высшей нервной деятельности и нейрофизиологии РАН
Москва, Россия
* E-mail: vraevsky@mail.ru
Поступила в редакцию 11.03.2021
После доработки 05.05.2021
Принята к публикации 06.05.2021
Аннотация
У крыс линии WAG/Rij, генетически предрасположенных к абсанс-эпилепсии (АЭ) и демонстрирующих депрессивно-подобное поведение, исследовали особенность функционирования соматосенсорной системы в периоды, когда АЭ еще не сформирована (2–3 мес.) и когда патология хорошо выражена (6–7 мес.). Контрольными животными являлись здоровые крысы линии Wistar, у которых регистрировали внеклеточные ответы одиночных нейронов соматосенсорной коры на одиночное электрическое раздражение области вибрисс. Установлено, что вызванная активность нейронов соматосенсорной коры на афферентную стимуляцию, вне зависимости от возраста и вида животных, включает четыре компонента: первичные (коротко-латентные) возбуждение и торможение, и вторичные (длинно-латентные) возбуждение и торможение. У крыс WAG/Rij, в отличие от здоровых крыс Wistar, в 2–3-месячном возрасте число коротко-латентных тормозных реакций значимо меньше, а число длинно-латентных тормозных реакций значимо больше. Постулируется, что нарушение баланса тормозных механизмов разной природы может обусловливать развитие как АЭ, так и депрессии у крыс линии WAG/Rij. У 6–7-месячных крыс линии WAG/Rij (в период, когда эпилептическая активность сформирована) обнаружено, что вторичная активационная реакция на афферентное раздражение представлена преимущественно многокомпонентной высокочастотной фазной реакцией (чередование коротких по продолжительности разрядов нейрона и тормозных периодов). У крыс линии Wistar в этот возрастной период вторичное возбуждение представлено тонической реакцией. Частота фазных разрядов сопоставима с частотой пик-волновой активности, являющейся электроэнцефалографическим признаком АЭ. Предполагается, что фазную реакцию можно рассматривать как нейронный эквивалент эпилептической активности, и пик-волновая активность у крыс WAG/Rij может быть провоцирована афферентацией от вибрисс.
ВВЕДЕНИЕ
Крысы линии WAG/Rij генетически предрасположены к абсанс-эпилепсии (АЭ) [1] и демонстрируют депрессивно-подобное поведение [2]. Электроэнцефалографическим проявлением эпилептического приступа является генерализованная пик-волновая активность (Spike-wave discharges), которая формируется у этих животных в 5–6-месячном возрасте [3]. Эпилептический приступ продолжается в течение нескольких секунд, а частота пик-волновых разрядов у крыс WAG/Rij уменьшается от начала приступа до его окончания с 15 до 6–7 Гц [4]. Существенная роль в формировании АЭ принадлежит сенсорным факторам. Было установлено, что ограничение видоспецифической афферентации за счет выстригания вибрисс в течение первых трех недель жизни крысенка провоцирует более раннее развитие АЭ [5]. В противоположность этому, ежедневная дополнительная тактильная стимуляция крысят линии WAG/Rij в течение первых 20-ти дней жизни препятствует развитию как АЭ, так и депрессивно-подобного поведения в 6-месячном возрасте у этих животных [6]. На формирование АЭ могут оказывать влияние и видоспецифические сенсорные факторы раннего онтогенеза. В экспериментах с перекрестным воспитанием новорожденных крысят показано, что замена самок WAG/Rij, генетически предрасположенных к АЭ, здоровыми самками линии Wistar в период молочного вскармливания замедляет развитие патологии [7]. Установлено, что данная процедура оказывает влияние и на проявление депрессии у этих животных [8].
Особая роль в возникновении эпилептического приступа отводится соматосенсорной коре. Предполагается, что она является триггерной зоной генерации пик-волновой активности [9]. Сформированный в этой области первичный разряд распространяется кортикофугально, активируя таламические ядра [10].
Относительно недавно появились сведения о том, что заднее ядро таламуса (РО) может выполнять функцию ревербератора, обеспечивая циркуляцию возбуждения между неокортексом и таламусом [11]. Нами установлено, что электрическая стимуляция РО вызывает в сенсомоторной коре у 6-месячных крыс WAG/Rij фазную реакцию (чередование коротких по продолжительности разрядов нейрона и тормозных периодов), которую было предложено рассматривать в качестве нейронного эквивалента пик-волновой активности [12]. В связи с тем, что РО опосредует паралемнисковый вход в соматосенсорную кору от вибрисс, допустимо предположить, что афферентация из области вибрисс способна инициировать эпилептический приступ у крыс линии WAG/Rij.
Таким образом, результаты исследования роли соматической афферентации в формировании АЭ свидетельствуют в пользу наличия функциональных особенностей соматосенсорного анализатора у крыс WAG/Rij, которые обусловливают как раннее развитие депрессии, так и формирование эпилептического статуса.
Задачей настоящего исследования было выявление специфики реакций одиночных нейронов соматосенсорной зоны коры на стимуляцию области вибрисс у крыс линии WAG/Rij 2–3-месячного возраста, когда АЭ еще не сформирована и 6–7-месячного возраста, когда АЭ хорошо выражена.
МЕТОДЫ ИССЛЕДОВАНИЯ
Использовали самцов крыс линии Wistar и линии WAG/Rij (собственное разведение). Все эксперименты выполняли в соответствии с протоколом использования лабораторных животных Института высшей нервной деятельности и нейрофизиологии РАН, основанного на директиве Совета Европейского Сообщества (86/609/ЕЕС).
В остром опыте под уретановым наркозом (0.15 г на 100 г массы тела животного) с помощью стеклянных микроэлектродов, заполненных 2 М раствором NaCl (диаметр кончика ≤1 мкм) регистрировали внеклеточную активность одиночных нейронов. Регистрирующий электрод ориентировали по координатам АР +2.5; L 5.5 и погружали вертикально. В этом случае он входил в соматосенсорную кору тангенциально под углом 45°. Ориентируясь на морфометрические исследования [13], было рассчитано, что IV слой коры в этом случае занимает пространство на глубине от 550 до 1400 мкм. Регистрацию активности нейронов осуществляли в этом диапазоне.
Как было отмечено, основанием данного исследования, в том числе, были полученные нами данные о возникновении в соматосенсорной коре фазной реакции на электрическую стимуляцию РО и высказанное нами предположение, что афферентация из области вибрисс способна инициировать приступ эпилепсии. В связи с тем, что при стимуляции РО активируются множественные афферентные входы в кору, мы сочли целесообразным использовать в настоящем исследовании не механическую стимуляцию отдельных вибрисс, позволяющую анализировать рецептивные поля сенсорных нейронов, а электрическое раздражение этой области, полагая, что результаты стимуляции множественных входов из области вибрисс более сопоставимо с результатами, полученными при электрическом раздражении РО.
Для стимуляции области вибрисс использовали игольчатые электроды с межэлектродным расстоянием 4 мм, которые вводили вдоль ряда С вибрисс. Стимуляцию осуществляли одиночным прямоугольным импульсом тока силой 0.5 мкА, длительностью 0.2 мс, что, как правило, в 2 раза превышало силу раздражения, вызывающую минимальное подергивание вибрисс.
Импульсную активность нейрона регистрировали в течение 4 с: 2 с до и 2 с после стимула. Результаты сохраняли в памяти компьютера. Для анализа реакций по результатам 10 раздражений строили перистимульные гистограммы с разрешением бина от 1 до 16 мс. Анализ осуществляли с помощью компьютерной программы, созданной по нашему заказу фирмой L-Card. На основе сравнения средней частоты потенциалов действия в фоне и после раздражения программа выделяла предполагаемые зоны реакции (возбуждение/торможение) по критерию 30%-ного увеличения или уменьшения средней частоты разрядов. После установления значимого отличия частоты потенциалов действия в предполагаемой зоне реакции по сравнению с фоном (использовали непараметрический критерий Вилкоксона) вычисляли латентный период (от момента нанесения раздражения до начала зоны реакции), продолжительность реакции и частоту потенциалов действия в зоне реакции.
Статистическую значимость межгрупповых различий определяли с использованием параметрического t-теста (все анализируемые показатели имели нормальное распределение – тест Шапиро–Уилка) и теста двух пропорций (two-sided).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Реакция нейронов соматосенсорной коры на стимуляцию области вибрисс у 2–3-месячных крыс линии Wistar
В экспериментах на 6 крысах линии Wistar зарегистрированы 48 нейронов. Из них 37 нейронов реагировали на одиночную электрическую стимуляцию области вибрисс. Реакция состояла из нескольких активационных и тормозных компонентов. По критериям латентного периода и длительности реакции нами выделено 4 компонента ответа на стимуляцию области вибрисс: первичное возбуждение (1 E), первичное торможение (1 I), вторичное возбуждение (2 E) и вторичное торможение (2 I) (рис. 1a). Латентный период первичного возбуждения составлял 27.8 ± 8.3 мс, а продолжительность 51.5 ± 44 мс. Латентный период вторичного возбуждения был 209.3 ± 67.5 мс а продолжительность 313 ± 224.7 мс. По обоим показателям первичный и вторичный ответ значимо отличались (t (13.1) – 45, p < 0.001; t (5.6) – 45, p < 0.001 соответственно). Первичное торможение возникало с латентным периодом 70.6 ± 23.4 мс, а латентный период вторичного торможения составлял 697.5 ± 146 мс. Отличия были значимы (t (15.3) – 26, p < 0.001).
Рис. 1.
Реакция нейронов соматосенсорной зоны коры на стимуляцию области вибрисс у 2–3-месячных крыс Wistar. Суммарные перистимульные гистограммы (10 предъявлений). Эпоха анализа 2с. 1 бин – 8 мс. По оси ординат – количество спайков в бине. Стрелка – момент нанесения раздражения. a – сложный ответ, включающий первичные возбуждение 1E и торможение 1I, вторичные возбуждение 2E и торможение 2I; b – вторичное возбуждение; с – первичное торможение; d – первичные возбуждение и торможение; e – соотношение активационных и тормозных компонентов ответа на стимуляцию области вибрисс у крыс Wistar и WAG/Rij; 1 – первичное возбуждение, 2 – первичное торможение, 3 – вторичное возбуждение, 4 – вторичное торможение. *p < 0.05 по критерию двух пропорций (two-sided).
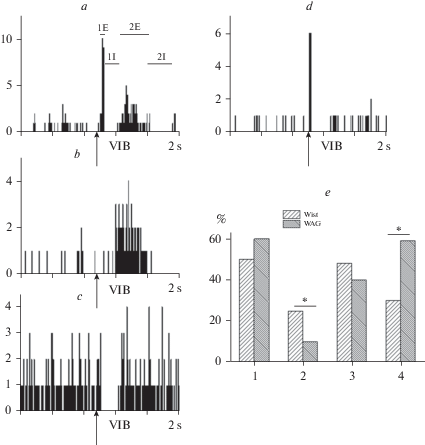
В ряде случаев реакция нейронов соматосенсорной коры на стимуляцию области вибрисс состояла из отдельных компонентов (рис. 1b–d). В связи с этим частота встречаемости перечисленных активационных и тормозных реакций была разной. Первичное возбуждение выявлено у половины всех зарегистрированных нейронов. Первичное торможение обнаружено у 25% нервных клеток. Вторичное возбуждение выявили у 48%, а вторичное торможение у 30% нейронов соматосенсорной коры (рис. 1e).
Реакция нейронов соматосенсорной коры на стимуляцию области вибрисс у 2–3-месячных крыс линии WAG/Rij
В этой серии экспериментов у 8 крыс WAG/Rij зарегистрировали 58 нейронов соматосенсорной коры. Из них 49 нейронов реагировали на одиночную электрическую стимуляцию области вибрисс. Компонентный состав реакций нейронов у крыс линии WAG/Rij не отличался от обнаруженного у крыс Wistar. Временные параметры ответов в основном не различались. Отличия были выявлены только для латентных периодов первичного возбуждения и вторичного торможения. У крыс линии WAG/Rij эти параметры были значимо меньше, чем у крыс линии Wistar (табл. 1).
Таблица 1.
Временные параметры реакций нейронов соматосенсорной коры в мс (латентные периоды и длительность) на стимуляцию области вибрисс у 2–3-месячных крыс линий Wistar и WAG/Rij
| Возбуждение 1. Лат. | Возбуждение 1 Длит. | Торможение 1. Лат. | Торможение 1. Длит. | Возбуждение 2. Лат. | Возбуждение 2. Длит. | Торможение 2. Лат. | Торможение 2. Длит. | |
|---|---|---|---|---|---|---|---|---|
| Wistar | 27.8 ± 8.3 n = 24 24 (24–28) |
51.5 ± 44 n = 24 40 (24–48) |
70.6 ± 23.4 n = 13 64 (48–56) |
136.4 ± 40.7 n = 13 136.4 (112–176) |
209.3 ± 67.5 n = 23 200 (164– 240) |
313 ± 224.7 n = 23 272 (144–370) |
697.5 ± 146 n = 15 640 (592– 840) |
327.7 ± 271.7 n = 15 232 (160–312) |
| WAG/Rij | 24.7 ± 3.0* n = 35 24 (24–24) |
51.1 ± 26 n = 35 48 (37–72) |
52 ± 8.4 n = 6 52 (48–56) |
110.6 ± 24.5 n = 6 108 (56–120) |
180.8 ± 44.2 n = 23 160 (144– 208) |
302.8 ± 240.2 n = 23 192 (112–416) |
531.1 ± 173 n = 34* 504 (400–660) |
307.8 ± 174.4 n = 34 272 (208–352) |
Существенные отличия реакций нейронов соматосенсорной коры на афферентную стимуляцию у крыс линии WAG/Rij были обнаружены по критерию встречаемости тормозных компонентов ответа. Число нейронов, реагирующих первичной тормозной реакцией у крыс линии WAG/Rij было значимо меньше (p = 0.04 по тесту двух пропорций – two-sided). (рис. 1e, 2). В отличие от первичного торможения вторичное торможение у крыс линии WAG/Rij зарегистрировано у большего числа нейронов соматосенсорной коры (59% у WAG/Rij, 30% у Wistar) (рис. 1e, 4). Различие значимо – р = 0.03 по тесту двух пропорций (two-sided).
Реакция нейронов соматосенсорной коры на стимуляцию области вибрисс у 6–7-месячных крыс линии Wistar
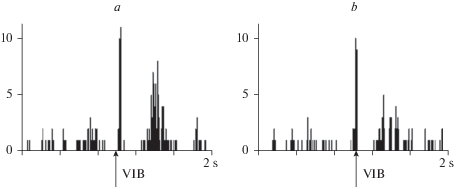
У 6-ти крыс линии Wistar зарегистрирована активность 52-х нейронов соматосенсорной коры, из которых 49 нейронов реагировали на одиночную электрическую стимуляцию области вибрисс. Как правило, реакция была сложной: включала активационные и тормозные компоненты и в целом была схожей с выявленной нами у крыс линии Wistar более раннего возраста. Наиболее существенное отличие заключалось в том, что у части нейронов вторичный активационный компонент мог быть представлен не только тоническим разрядом (рис. 2a), но и двумя более короткими по продолжительности фазами, а у 4-х нейронов был обнаружен 3-компонентный ответ (рис. 2b).
Рис. 2.
Реакции нейронов соматосенсорной зоны коры на стимуляцию области вибрисс у 6–7-месячных крыс Wistar: a – сложный ответ, включающий вторичное тоническое возбуждение; b – сложный ответ, включающий вторичную фазную активацию. Суммарные перистимульные гистограммы (10 предъявлений). Эпоха анализа – 2 с; 1 бин – 8 мс. По оси ординат – количество спайков в бине. Стрелка – момент нанесения раздражения.
Реакция нейронов соматосенсорной коры на стимуляцию области вибрисс у 6–7-месячных крыс линии WAG/Rij
У 6-ти крыс линии WAG/Rij в возрасте, когда АЭ уже хорошо выражена, исследована реакция 51-ного нейрона соматосенсорной коры на стимуляцию области вибрисс. В этой серии экспериментов 46 нейронов реагировали на афферентную стимуляцию. Компонентный состав реакций был таким же, как и у крыс Wistar. Наиболее выраженное отличие реакции у крыс линии WAG/Rij заключается в появлении многокомпонентной фазной реакции (рис. 3a), которая представлена серией из 6–8 коротких по продолжительности (56.0 ± 20.7 мс) высокочастотных разрядов нейрона и тормозных пауз, а также в уменьшении числа вторичных активационных тонических реакций (рис. 3b, 3). Период генерации этих разрядов составлял 79.7 ± 16.7 мс. Как было отмечено в предыдущем разделе работы, у крыс линии Wistar также были обнаружены фазные реакции нейронов коры на стимуляцию вибрисс. Однако они по целому ряду показателей отличаются от фазных реакций, зарегистрированных у крыс WAG/Rij. Во-первых, фазные реакции у крыс линии Wistar обнаружены только у 4-х нейронов, в то время как фазный ответ у крыс линии WAG/Rij был характерен для 33% нервных клеток (рис. 3b, 4). Во-вторых, частота потенциалов действия в фазном разряде у WAG/Rij значимо больше (t (9.4) – 28, p < 0.001) (рис. 3c). В-третьих, фазный ответ у крыс Wistar представлен максимум тремя разрядами, в то время как у крыс WAG/Rij количество разрядов может достигать восьми. В результате общая продолжительность вторичного компонента реакции у крыс WAG/Rij (366.1 ± 95 мс) значимо превышает длительность вторичного возбуждения у крыс Wistar (t (3.6) – 53, p < 0.001) (табл. 2).
Рис. 3.
Особенности реакций нейронов соматосенсорной коры на стимуляцию области вибрисс у 6–7-месячных крыс WAG/Rij. a – фазная реакция нейрона у крысы линии WAG/Rij (Суммарная перистимульная гистограмма, 10 предъявлений). Эпоха анализа – 2 с; 1 бин – 8 мс. По оси ординат количество спайков в бине. Стрелка – момент нанесения раздражения. b – соотношение активационных и тормозных компонентов ответа нейронов соматосенсорной коры у крыс линии Wistar (Wist) и WAG/Rij (WAG). 1 – первичное возбуждение, 2 – первичное торможение, 3 – вторичная тоническое возбуждение, 4 – вторичная фазная активация, 5 – вторичное торможение; *p < 0.05 по критерию двух пропорций (two-sided). с – сравнительный анализ частоты потенциалов действия в фазном компоненте ответа у крыс Wistar и WAG/Rij (по оси ординат частота в Гц); *p < 0.001 по t-критерию.

Таблица 2.
Временные параметры реакций нейронов соматосенсорной коры (мс) на стимуляцию области вибрисс у 6–7-месячных крыс линий Wistar и WAG/Rij
| Возбуждение 1 Длит | Торможение 1 Лат | Возбуждение 2 Лат | Возбуждение 2 Длит | Торможение 2. Лат | |
|---|---|---|---|---|---|
| Wistar | 189 ± 79.5 n = 20 168 (144–208) |
68.6 ± 27.6 n = 20 60 (52–76) |
252.2 ± 67.5 n = 37 232 (208–304) |
228.7 ± 144.9 n = 37 184 (128–320) |
623.2 ± 109.6 n = 10 592 (536–728) |
| WAG/Rij | 37 ± 21.4 * n = 31 32 (16–56) |
43.2 ± 21.1 * n = 15 32 (32–64) |
192 ± 67 * n = 23 192 (152–268) |
366.1 ± 95* n = 18 351 (280–424) |
740.8 ± 121.8 * n = 15 736 (664–848) |
Изменение структуры вызванной реакции сопровождается изменением временных параметров компонентов ответа (табл. 2).
Обращает на себя внимание меньший латентный период первичного торможения и вторичного возбуждения и меньшая продолжительность активационных ответов у крыс WAG/Rij. Больший латентный период вторичного торможения у крыс линии WAG/Rij является, по-видимому, следствием большей продолжительности вторичного возбуждения у крыс WAG/Rij по сравнению со вторичным возбуждением у крыс Wistar.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Исследование реакции нейронов соматосенсорной коры на стимуляцию вибрисс у крыс линии WAG/Rij, генетически предрасположенных к АЭ, и у здоровых животных линии Wistar позволило установить, что одиночное афферентное раздражение вызывает сложную реакцию нейронов коры у этих животных. Она представлена коротко- и длинно-латентными активационными и тормозными ответами. Вероятнее всего, каждый компонент ответа отражает вовлечение разных структур в реакцию на сенсорный стимул. Если первичное возбуждение и торможение можно рассматривать как реакцию на короткий электрический стимул, то вторичные длинно-латентные компоненты ответа, вероятнее всего, являются следствием вовлечения некой внутрикорковой системной организации нейронов, принимающих участие в анализе сенсорной информации или реакции нейронов на афферентное возбуждение, опосредованное ростральным и каудальным ретикулярными ядрами [10].
Первичное возбуждение оказалось наиболее стабильной реакцией, присущей животным обеих исследованных нами линий, и не зависело от возраста. Первичное торможение, которое связывают с вовлечением тормозных интернейронов коры, у 2–3-месячных крыс линии WAG/Rij, когда пик-волновая активность еще не сформирована, было выявлено у ограниченного числа нейронов и было значимо меньше, чем у крыс линии Wistar. Это позволяло сделать заключение о дефиците тормозной регуляции у крыс WAG/Rij. Однако, по всей видимости, речь может идти только об афферентном (первичном) торможении, так как вторичное длинно-латентное торможение выявлено у большего числа нейронов крыс линии WAG/Rij, чем у крыс линии Wistar. Достаточно неожиданным оказалось восстановление баланса между первичным и вторичным торможениями к 6–7-месячному возрасту у крыс WAG/Rij – казалось бы, что структурно-функциональное формирование неокортекса должно было завершиться к 2-месячному возрасту. Возможно, обнаруженное нами явление реализуется за счет перестройки внутрикорковых связей у крыс WAG/Rij к периоду формирования АЭ, что могло бы отражать один из компенсаторных механизмов, препятствующих развитию патологии. Исследование этого вопроса требует продолжения, но мы полагаем, что нарушение баланса между первичным и вторичным торможениями у 2–3-месячных крыс WAG/Rij можно рассматривать, как одно из оснований развития у этих животных комбинированной патологии: АЭ и депрессии.
У 6–7-месячных крыс линии WAG/Rij, когда АЭ хорошо проявляется, выявлены существенные изменения вызванной активности нейронов соматосенсорной коры. Это проявляется в изменении структуры вторичного возбуждения ответа. У 33% нейронов этот компонент представлен фазными реакциями: короткими высокочастотными разрядами. По своей структуре эти разряды весьма схожи со вспышечной активностью нейронов, зарегистрированных в разных отделах мозга и синхронизированных с появлением пик-волновой активности [14, 15]. Существенным является тот факт, что период генерации обнаруженных нами фазных разрядов составляет 79.7 ± 16.7 мс, что соответствует диапазону частот пик-волновой активности, характерной для крыс линии WAG/Rij [4]. Это позволяет нам заключить, что фазная реакция является нейронным эквивалентом пик-волновой активности, являющейся электроэнцефалографическим признаком АЭ. Тот факт, что эта активность может быть инициирована стимуляцией вибрисс, подчеркивает значение соматосенсорной системы для патогенеза АЭ. При рассмотрении возможных механизмов генерации фазной активности следует иметь в виду, что она возникает в ответ на короткий одиночный стимул и представлена серией разрядов, количество которых может достигать восьми. Это, по нашему мнению, свидетельствует о вовлечении в процесс достаточно сложной нейронной организации, которая может включать таламо-кортикальные связи, а также и внутрикорковую системную организацию нейронов. Ранее было высказано предположение о существовании реверберационного механизма генерализации эпилептической активности, который опосредован задним ядром таламуса (PO) [11]. Нами установлено, что период генерации фазных разрядов в среднем составляет 80 мс. Такой временной интервал достаточен для реализации корково-таламо-кортикального возбуждения, что допускает участие механизма реверберации в генерации фазных разрядов, выявленных нами. Альтернативным механизмом может быть функциональное изменение в системе внутрикорковых связей, обусловливающее генерацию пейсмейкерной активности нейронов. Уточнение механизмов требует дальнейших исследований, но, принимая заключение, что фазная реакция является нейронным эквивалентом пик-волновой активности, мы вправе постулировать, что афферентное возбуждение от вибрисс способно инициировать приступ АЭ у крыс линии WAG/Rij.
Список литературы
White H, Löscher W (2014) Searching for the ideal antiepileptogenic agent in experimental models: single treatment versus combinatorial treatment strategies. Neurotherapeutics 11: 373–384. https://doi.org/10.1007/s13311-013-0250-1
Sarkisova K, van Luijtelaar G (2011) The WAG/Rij strain: a genetic animal model of absence epilepsy with comorbidity of depression [corrected]. Progr Neuropsychopharmacol Biol Psychiatry 35: 854–876. https://doi.org/10.1016/j.pnpbp. 2010.11.010
Coenen A, van Luijtelaar G (2003) Genetic animal models for absence epilepsy: a review of the WAG/Rij strain of rats. Behav Genetics 33: 635–655.https://doi.org/10.1023/a:1026179013847
Bosnyakova D, Gabova A, Zharikovae A, Gnezditski V, Kuznetsova G, van Luijtelaar G (2007) Some peculiarities of time-frequency dynamics of spike-wave discharges in humans and rats. Clin Neurophysiol 118: 1736–1743. https://doi.org/10.1016/j.clinph.2007.04.013
Sitnikova E (2011) Neonatal sensory deprivation promotes development of absence seizures in adult rats with genetic predisposition to epilepsy. Brain Res 1377: 109–118.https://doi.org/10.1016/j.brainres.2010.12.067
Aymen Balikci, Gul Ilbay, Nurbay Ates (2020) Neonatal tactile stimulations affect genetic generalized epilepsy and comorbid depression-like behaviors. Front Behav Neurosci 23(14): 132. https://doi.org/10.3389/fnbeh.2020.00132
Sitnikova E, Rutskova E, Raevsky V (2015) Reduction of epileptic spike-wave activity in WAG/Rij rats fostered by Wistar dams. Brain Res 1594: 305–309.https://doi.org/10.1016/j.brainres.2014.10.067
Sarkisova K, Gabova A (2018) Maternal care exerts disease-modifying effects on genetic absence epilepsy and comorbid depression. Genes, Brain and Behavior 17: e12477.https://doi.org/10.1111/gbb.12477
Sitnikova E, van Luijtelaar G (2004) Cortical control of generalized absence seizures: effect of lidocaine applied to the somatosensory cortex in WAG/Rij rats. Brain Res 1012: 127–137.https://doi.org/10.1016/j.brainres.2004.03.041
Lüttjohann A, Jan-Mathijs S, van Luijtelaar G (2014) Termination of ongoing spike-wave discharges investigated by cortico–thalamic network analyses. Neurobiol of Disease 70: 127–137. https://doi.org/10.1016/j.nbd.2014.06.007
Lüttjohann A, van Luijtelaar G (2015) Dynamics of networks during absence seizure’s on- and offset in rodents and man. Front Physiol 6: 16.https://doi.org/10.3389/fphys.2015.00016
Tsvetaeva D, Sitnikova E, Raevsky V (2019) Cortical Somatosensory Neurons in WAG/Rij Rats Transform Firing Evoked by Simulation of Posterior Thalamic Nucleus from Tonic to Phasic at Age of 6 Months. Bull Exp Biol Med 168: 1–4. https://doi.org/doi 10.1007/s10517-019-04632-8
Shubina O, Teltsov L, Komusova O (2015) Cytological and morphometric features of the cortex of white rats. Modern Problems Science and Education Issue № 2 (part 1).
Pinault D, O’Brien T (2005) Cellular and network mechanisms of genetically-determined absence seizures. Thalamus and Related Systems 3: 181–203. https://doi.org/doi: . Epub 2007 Jan 22https://doi.org/10.1017/S1472928807000209
Zheng T, O’Brien T, Morris M, Reid C, JovanovskaV, O’Brien P, Leena van Raay, Gandrathi A, Pinault D (2012) Rhythmic neuronal activity in S2 somatosensory and insular cortices contribute to the initiation of absence related spike and wave discharges. Epilepsia 53: 1948–1958. https://doi.org/doi:10.1111/j.1528-1167.2012.03720.x
Дополнительные материалы отсутствуют.
Инструменты
Российский физиологический журнал им. И.М. Сеченова