

Российский физиологический журнал им. И.М. Сеченова, 2021, T. 107, № 3, стр. 267-287
Возможности и перспективы поведенческого теста “Водный лабиринт Морриса”
Д. П. Чернюк 1, *, А. В. Большакова 1, О. Л. Власова 1, И. Б. Безпрозванный 1, 2, **
1 Санкт-Петербургский политехнический университета Петра Великого
Санкт-Петербург, Россия
2 Отделение физиологии Юго-западного медицинского центра Техасского университета
Даллас, США
* E-mail: dashachernuk@gmail.com
** E-mail: mnlabspb@gmail.com
Поступила в редакцию 30.11.2020
После доработки 18.12.2020
Принята к публикации 19.12.2020
Аннотация
Поведенческий тест “Водный лабиринт Морриса” является универсальным способом изучения когнитивных функций у подопытных грызунов, особенно он эффективен при выявлении отклонений функций памяти и обучения, что делает его незаменимым при исследовании старения, инсульта, нейродегенеративных заболеваний, влияния терапевтических препаратов и т.д. Однако данный тест может быть намного более информативным инструментом анализа поведения подопытных животных, чем кажется на первый взгляд. Формирование, консолидация памяти и обучение – довольно сложные процессы, которые главным образом задействуют гиппокамп, но и кроме него в этих процессах участвуют многие зоны мозга. Различные протоколы этого теста настолько чувствительны к изменениям нормальной функции различных областей мозга, не только гиппокампа, что их можно использовать в качестве “индикатора” нормального функционирования когнитивных функций. Таким образом, разные модификации “Водного лабиринта Морриса” включают различные механизмы навигации, обучения и памяти, а результаты этих тестов и их правильный анализ могут дать очень много информации о поведении лабораторных животных. В данной обзорной статье мы приводим перечень наиболее популярных методик проведения процедуры “Водного лабиринта Морриса” и разбираем, какие информативные параметры могут помочь исследователю в анализе результатов данного теста.
ВВЕДЕНИЕ
Головной мозг – самая таинственная и загадочная структура человеческого организма. Несмотря на то, что его строение уже достаточно хорошо изучено, до сих пор функционирование мозга в большей степени остается для человечества загадкой. Обучение и память являются одними из самых сложных и малопонятных функций человеческого мозга. А самой важной для формирования и консолидации памяти частью мозга считается гиппокамп. Британский нейробиолог Ричард Грэм Майкл Моррис посвятил свою научную деятельность изучению функционирования гиппокампа и в начале 1980-х первым описал процедуру водного лабиринта в открытом поле для грызунов [1, 2]. Идея данного поведенческого теста достаточно проста: животных, обычно крыс или мышей, помещают в большой круговой бассейн с водой, и им необходимо выбраться на скрытую под поверхностью воды платформу. Новизна данной методики заключалась в том, что пройти этот тест возможно только с использованием пространственной памяти, так как никаких локальных сигналов нет – грызунам необходимо определить по памяти местоположение объекта, который они никогда не видели, не чуяли и не могли услышать. После первых публикаций эта методика быстро набрала популярность в нейробиологических исследованиях, в том числе благодаря исследованиям канадской группы ученых во главе с Яном Уишоу [3], стала именоваться “Водным лабиринтом Морриса” (Morris Water Maze, MWM) и приобрела множество различных технических вариаций [4–9]. К преимуществам данной методики можно отнести высокую надежность в широком диапазоне конфигураций водных резервуаров и процедур проведения испытаний, его межвидовую утилитарность (крысы, мыши и даже люди – в виртуальном лабиринте [10]), многочисленные свидетельства его эффективности в качестве способа изучения гиппокамп-зависимой пространственной навигации и “референс” памяти [11–13], а также его относительную невосприимчивость к мотивационным факторам проводимых экспериментов [5]. Таким образом, “Водный лабиринт Морриса” стал золотым стандартом для изучения пространственной памяти и процессов обучения, он входит в перечень необходимых тестов для фенотипирования мутантных и трансгенных мышей [14], а также часто используется в качестве общего анализа когнитивной функции [4], например, для тестирования воздействия различных нарушений нервной системы, таких как модели инсульта на животных [15], старения [16], нейродегенеративных заболеваний [17–22] или потенциального воздействия новых терапевтических препаратов [23]. Главным параметром оценки процессов обучения и памяти во всех этих протоколах является латентный период нахождения платформы испытуемым животным, кроме того, показательным параметром служит и сама траектория плавания до платформы. Так как в век цифровых технологий визуальная регистрация является достаточно необъективной и неточной методикой изучения поведения, широкое распространение получила автоматическая трассировка перемещения животного с помощью присоединенной к компьютеру цифровой видеокамеры [24–26 ]. Значительным плюсом автоматизации видеонаблюдения и анализа является надежность и последовательность алгоритма, чего сложно достичь при визуальной и ручной обработке данных, тем более, если речь идет о длительных периодах времени. Кроме того, использование автоматизированного анализа дает возможность более качественно и всесторонне анализировать полученные экспериментальные данные, например, путем более сложного обсчета траектории движения подопытного животного, тем самым позволяя выйти за рамки первоначальной поставленной перед MWM-тестом задачи исследования только пространственной памяти. Настоящий обзор нацелен на обсуждение наиболее часто применяемых протоколов проведения поведенческого теста MWM, анализ его классических и современных параметров, а также возможностей, которые можно получить с использованием автоматического анализа полученных данных (в том числе, с помощью разработанного авторами программного продукта).
ВОДНЫЙ ЛАБИРИНТ МОРРИСА И ЕГО МОДИФИКАЦИИ
Процедура “Водного лабиринта” используется для изучения процесса обучения с начала XX века [27–29]. Температура воды и ее повышение, когда грызуны выбирают из всех секторов более теплый [30], а также “способность к плаванию” влияют на скорость обучения в простых тестах [31]. В этих и более усовершенствованных процедурах “Водного лабиринта Морриса” животных не нужно лишать пищи, чтобы мотивировать обучение, или проводить обширную предварительную подготовку, также исключается использование удара током для мотивирования, как, например, в поведенческих тестах, направленных на формирование условных рефлексов, таких как “Условный рефлекс активного избегания” (“active avoidance”) и тест “Условного рефлекса страха” (“fear conditioning”). Крысы и мыши являются естественными пловцами и легко выполняют задачу нахождения скрытой под водой спасательной платформы, даже если они перенесли мозговые поражения различного типа [2]. Вода обеспечивает хороший контроль возможных обонятельных сигналов и позволяет проводить разнообразные обучающие процедуры, чтобы разделить эффекты обучения и работоспособности. Типичные испытания в водном лабиринте ограничены 60-ю с, в то время как испытания в сухом лабиринте часто длятся намного дольше. Это позволяет повысить пропускную способность и эффективность “Водного лабиринта Морриса” [32]. Однако он также имеет определенные ограничения, одним из которых является тот факт, что различные компоненты памяти, то есть “референс” и рабочая память, не могут быть проверены одновременно [33]. Кроме того, для данного теста важным фактором является стабильное положение достаточного количества визуальных ориентиров, так как основная задача MWM – это изучение пространственной памяти. Благодаря своим особенностям “Водный лабиринт Морриса” является универсальным средством для решения множества различных задач [34].
Самая простая задача – это обучение поиску скрытой под водой спасательной платформы в одном фиксированном месте, часто включается в серию иногда довольно сложных протоколов обучения и тестирования для исследования конкретных теоретических вопросов. Разные протоколы задействуют разные механизмы навигации, обучения и памяти. Далее рассмотрим основные из них.
1. Пространственная “референс” память
Самый простой и широко используемый протокол проведения поведенческого теста “Водный лабиринт Морриса” заключается в использовании замутненной воды и спрятанной под ее поверхностью (на 0.5–2.0 см) платформы, которая находится в фиксированном месте по отношению к внешним визуальным сигналам, обычно это середина одного из четырех квадрантов бассейна (рис. 1 ). Во время обучающих испытаний животных помещают в воду в одной из четырех точек у стенки бассейна, головой от центра. Местоположение этих точек выбирается произвольно, но аналогично направлениям сторон света, т.е. крестообразно, и обычно обозначаются N, E, S и W (рис. 1B). Часто стартовые позиции совмещаются с местоположением дополнительных контрастных черно-белых ориентиров четырех типов (ключей MWM), которые используются для стандартизации визуальных сигналов при проведении “Водного лабиринта Морриса”. С животными проводят серию ежедневных испытаний с использованием случайного или полуслучайного набора начальных местоположений. Полуслучайные наборы стартовых позиций являются наиболее распространенными: используются четыре позиции с тем ограничением, что один тест каждый день проводится с каждой из четырех позиций [5]. Встречаются исследования, где используются восемь стартовых локаций [35]. Подопытные быстро учатся находить фиксированную платформу: с каждым испытанием уменьшается время ее поиска, а траектории плавания становятся более короткими и прямолинейными (рис. 1C). В процессе испытания система слежения фиксирует сокращение времени поиска платформы и уменьшение длины пути, а также изменения таких параметров как скорость плавания, направленность относительно местоположения платформы и так далее. Наблюдение за животными показывает, что, забравшись на спасательную платформу, они часто встают и оглядываются, словно пытаясь определить свое местоположение в пространстве. Обучение проводится ежедневно в течение от 2 до 14 дней и состоит из 3–6 процедур для каждого животного в день.
Рис. 1.
Стандартный протокол проведения “Водного лабиринта Морриса”. А – конфигурация оборудования и основные параметры бассейна. B – воображаемое разделение площади бассейна на квадранты (NW, NE, SE и SW) и начальные положения испытуемого животного (N, E, S и W). Показан пример расположения скрытой платформы в центре одного из четырех квадрантов бассейна. C – пример изменения траектории плавания грызуна в ходе обучающих испытаний. D – пробное испытание в стандартном протоколе проведения “Водного лабиринта Морриса”. Воображаемое разделение площади бассейна на квадранты (NW, NE, SE и SW) и пример положения, где во время тренировочных испытаний располагалась платформа. E – пример траектории плавания грызуна в ходе пробного испытания. Fig. 1 . Standard protocol for conducting the “Morris Water Maze” test. A – Equipment configuration and basic parameters of the pool. B – The imaginary division of the pool into quadrants (NW, NE, SE и SW) and the initial positions of the test animal (N, E, S и W). An example of the location of a hidden platform in the center of one of the four pool quadrants. C – An example of changes in the rodent swimming trajectory during training trials. D – Probe trial in the standard protocol for conducting the “Morris Water Maze”. The imaginary division of the pool into quadrants (NW, NE, SE и SW) and an example of the position where the platform was located during training trials. E – An example of a rodent trajectory during a trial test.
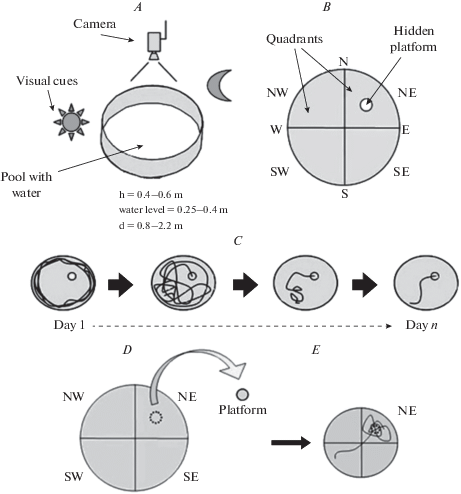
После окончания обучения через 24 ч проводится однократное тестирование поведения животного в бассейне в условиях отсутствия спасательной платформы – животное запускается в бассейн на определенное время, после истечения которого испытание заканчивается (рис. 1D, E). В данной процедуре происходит оценка так называемой “силы памяти”, т.е. насколько хорошо животное запомнило место локализации платформы из предыдущих обучающих испытаний. Для измерения чаще всего используют параметр времени, проведенного грызуном в квадранте, где раньше находилась платформа. Более сложные измерения включают среднее расстояние положения животного от местоположения платформы (индекс обучения) [36, 37] или число пересечений местоположения платформы и др. Многие из этих измерений включают видеозапись поведения грызуна и применение компьютерного программного обеспечения для анализа полученных данных [32]. Иногда проводятся промежуточные пробные испытания, например, после каждых 2–3 дней обучения. Тогда результаты этих пробных испытаний можно использовать в качестве показателя скорости обучения [38].
Нередко, как и в исследовании Морриса в 1981 г., в течение 1–2 дней проводятся предварительные ознакомительные процедуры, для чего грызунов помещают на 180 с в бассейн с замутненной водой, но без платформы. В этих предварительных испытаниях делаются только качественные наблюдения [1], например, тестируется зрение и умение животных плавать.
2. Рабочая память
Данная модификация проведения теста отличается от базовой тем, что платформа помещается в одну из четырех новых позиций каждый день тестирования, при этом проводятся только 2 испытания в день для каждого животного, таким образом создается так называемая процедура “отложенного сопоставления с местом” (“delayed matching to place”, DMP) (рис. 2 ). Главный вопрос, которым задается исследователь, заключается в том, может ли грызун показать улучшенный результат в испытании № 2, основываясь на опыте из испытания № 1 [4, 39]. Такая процедура обучения считается аналогичной быстрому процессу приобретения воспоминаний, задействует рабочую память. По сути, эта процедура позволяет изучать повторяющиеся случаи обучения за одно испытание. Интервал между испытаниями № 1 и № 2 (intertrial interval) может затем систематически изменяться, что помогает выявить, насколько хорошо работает кратковременная пространственная память [8].
Рис. 2.
Протокол проведения “Водного лабиринта Морриса” для изучения рабочей памяти подопытного животного. А – конфигурация оборудования. B – воображаемое разделение площади бассейна на квадранты и начальное положение скрытой платформы в центре одного из четырех квадрантов. C – пример изменения местоположения скрытой платформы в каждый из дней проводимых испытаний. Fig. 2 . The protocol of the “Morris Water Maze” to study the working memory of the experimental animal. A – Equipment configuration. B – The imaginary division of the pool into quadrants and the example of hidden platform initial position in the center of one of the four quadrants. C – An example of a change in the location of a hidden platform in each of the testing days.
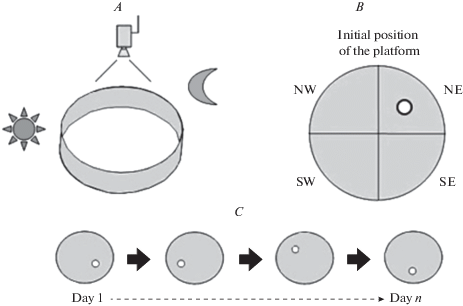
К конфигурации “Водного лабиринта Морриса”, в которой испытуемые вынуждены использовать рабочую память, можно отнести “on demand” процедуру (или процедура “по требованию”). Данная разновидность проведения теста была представлена в публикации чешских ученых 1985 г. [6]. Стандартная подводная платформа была заменена складной (платформа “по требованию”, или платформа Atlantis, названная в честь потерянного города), чтобы предотвратить случайное обнаружение скрытой цели. Платформу поднимали под поверхность воды только после того, как животное оставалось в целевой области в течение заранее определенного времени. Если это время истекало, но критерий не был достигнут, платформу поднимали и помещали на нее подопытную крысу.
В работах Бурешовой выполнение задания со складной платформой происходило со скоростью, аналогичной скорости с жесткой платформой, когда условия критерия были легкими (расстояние до целевой области – 15 см, время – 2.5 с), но постепенно ухудшались, так как время в целевой области увеличивалось до 10 с. Бурешова и соавторы пришли к выводу, что применение модифицированной методики позволяет исследовать тонкую структуру когнитивных карт и их изменений, вызванных различными вмешательствами [4, 6]. Также эта процедура приводит к приобретению высокосфокусированной стратегии поиска, ориентированной на целевое местоположение во время обучения [40].
3. Обратное обучение
Многие из протоколов проведения теста включают в себя скрытое перемещение спрятанной платформы. Так, например, в процедуре обратного обучения (или переобучения) после проведения ряда обучающих испытаний с фиксированной платформой она перемещается в другой сектор бассейна. Поскольку платформа остается спрятанной, для животного не очевидно, что что-то изменилось, до тех пор, пока оно не обнаруживает отсутствие платформы на своем обычном месте. Основное внимание уделяется тому, как животное реагирует на это изменение и как быстро оно находит новое место. Повторное обучение, которое происходит в протоколе обратного обучения, использовалось в анализе основных генетических факторов, определяющих поведение разных линий мышей [41].
4. Обучение распознаванию
Эта процедура была разработана для изучения процесса пространственного и непространственного распознавания. В этой конфигурации используются две видимые платформы: одна стандартная, а другая плавающая. При обучении пространственному распознаванию стандартная платформа определялась ее положением в пространстве, в то время как при обучении визуальному распознаванию стандартная платформа отличалась своей яркостью или рисунком, при этом исключались лишние сигналы лабиринта. В дополнение к обычным параметрам также учитывается мера первого выбора между двумя платформами. Было обнаружено, что обучение пространственному распознаванию происходит быстрее, чем обучение визуальному распознаванию, в отличие от данных более ранних работ Морриса [1], в которых обучение по меткам происходило быстрее, чем обучение по локализации (следует помнить, что процедура обучения и характеристики платформ различны в двух задачах). Также была предпринята попытка объединить протоколы, направленные на изучение пространственной памяти и процессов распознавания: рабочую память изучали в двухплатформенном обучающем протоколе, в котором местоположение “правильной” платформы ежедневно изменялось в произвольном порядке [42].
5. Некартографические стратегии
Изначально “Водный лабиринт Морриса” был разработан для изучения пространственной памяти грызунов, однако у подопытных животных есть и другие, не связанные с пространственным ориентированием стратегии, при помощи которых осуществляется решение задачи навигации [43]. Например, крыса может научиться делать определенную последовательность движений – рабочая стратегия. Эта стратегия эффективна только в том случае, если отправная точка и цель всегда фиксированы относительно друг друга. В качестве альтернативы крыса может научиться подходить к определенным локальным (проксимальным) сигналам, связанным с целью – таксис-стратегия. Однако в этом случае крыса может достичь непосредственно цели только с места, откуда видны локальные сигналы. Используя различные варианты процедуры проведения тестирования, можно продемонстрировать приобретение этих стратегий, а также стратегию картографирования: изучение относительного местоположения объектов в ближайшем окружении и прямой переход к цели из любой начальной точки. Крысы могут приобретать как картографические, так и некартографические стратегии и применять их, когда этого требуют особые ситуации, например, если расположение платформы случайное, если недостаточно локальных сигналов, а иногда в качестве дополнительной стратегии к стратегии картографирования. Было высказано предположение, что грызуны одновременно используют ряд поведенческих стратегий, включая ответы по положению, ответы по меткам и ответы по месту [7]. Последние исследования показывают, что поисковая стратегия мыши, выражающаяся в определенном паттерне движения, несет важную информацию о ее когнитивных функциях, но тем не менее, все еще малое количество исследований используют данный параметр в своих работах [44].
6. Другие модификации
Другие варианты проведения “Водного лабиринта Морриса” включают в себя такие изменения, как ограничение траектории плавающего животного для минимизации навигационных требований (например, кольцевой водный лабиринт [9]), уменьшение количества доступных внешних сигналов между обучением и пробным тестированием [8], использование плавающих платформ и другие манипуляции. Также был разработан радиальный водный лабиринт, сочетающий достоинства радиального лабиринта с простотой подготовки и проведения водного лабиринта. Такая методика часто используется при тестировании трансгенных мышей, имеющих наследственную форму болезни Альцгеймера [45].
Краткая информация об особенностях упомянутых в этом разделе модификаций проведения поведенческого теста “Водный лабиринт Морриса” представлена в табл. 1 .
Таблица 1.
Общая краткая информация различных модификаций “Водного лабиринта Морриса” Table 1 . General summary of various modifications of the “Morris Water Maze”
| Задача Task |
Модификации MWM Modification of MWM |
Особенности протокола Protocol features |
Задействованные зоны мозга Involved brain areas |
|---|---|---|---|
| “Референс” память "Reference" memory |
Стандартный протокол [1, 5] Standard protocol [1, 5] |
Несколько дней обучающих испытаний, в конце – одно пробное. Во время обучающих – платформа
фиксирована в одном месте, не видна, вода непрозрачна. Во время пробного – платформа
изымается Several days of training trials, at the end – one of probe trial. During training– the platform is fixed in one place, not visible, the water is opaque. During the probe trial – the platform is removed |
Гиппокамп, неокортекс, кора (лобная доля) Hippocampus, neocortex, cortex (frontal lobe) |
| Рабочая память Working memory |
“Отложенное сопоставление с местом”
(DMP) [8, 39] “Delayed matching to place” (DMP) [8, 39] |
Платформа перемещается на новую позицию каждый день тестирования, проводятся только
2 испытания в день The platform is moved to a new position every day of testing, only 2 tests per day are performed |
Гиппокамп, кора (затылочная, теменная доли) Hippocampus, cortex (occipital, parietal lobe) |
| “По требованию” [6, 40] “On demand” [6, 40] |
Стандартная платформа заменяется складной и предъявляется при нахождении животного
в целевой области в течение определенного времени The standard platform is replaced by a folding one and is presented when the animal is in the area for a set time |
Гиппокамп, кора (затылочная, теменная доли) Hippocampus, cortex (occipital, parietal lobe) |
|
| Обратное обучение Retraining |
Протокол переобучения [41] Retraining protocol [41] |
После проведения обучающих испытаний с фиксированной платформой она перемещается на
другую позицию After training tests with a fixed platform, it is moved to another position |
Дорсо-медиальный таламус,
кора (лобная доля), хвостатое ядро стриатума Dorso-medial thalamus, cortex (frontal lobe), caudate nucleus of the striatum |
| Обучение распознаванию Recognition training |
Двухплатформенный обучающий
протокол [1, 42] Two-platform training protocol [1, 42] |
Используются две видимые платформы: одна стандартная, а другая плавающая. Они могут
выглядеть идентично, или быть визуально разными для анализа пространственного или
визуального распознавания There are two visible platforms, one standard and the other floating. They may look identical or be visually different for analysis of spatial or visual recognition |
Гиппокамп, кора, вентральная область покрышки, неокортекс, базальные ядра Hippocampus, cortex, ventral tegmental region, neocortex, basal nuclei |
| Некартографические стратегии Non-cartographic strategies | Различные варианты процедур Various options for procedures |
Например, случайное расположение платформы или скудность локальных сигналов [7] For example, random platform arrangement or scarcity of local signals [7] |
Различные зоны мозга Different areas of the brain |
| Другие Others |
Множество различных модификаций Many different modifications |
Например, кольцевой водный лабиринт [9] или уменьшение внешних сигналов между обучением и пробным испытанием [8] For example, a circular water maze [9] or a decrease in external signals between training and probe trials [8] |
Различные зоны мозга Different areas of the brain |
Как видно из вышесказанного, поведенческий тест “Водный лабиринт Морриса” имеет значительное число модификаций, мы упомянули лишь самые популярные из них. Такое разнообразие протоколов проведения теста направленно на решение множества исследовательских задач, однако кроме правильной процедуры проведения теста важно иметь возможность максимально эффективно и информативно анализировать полученные при помощи выбранного протокола данные. Для этой цели существуют специальные параметры, некоторые из них довольно просты и их вычисление возможно вручную, другие же – требуют специальных сложных вычислений, для которых используют программные продукты, зато отличаются большей точностью и информативностью.
ПАРАМЕТРЫ
Первым и наиболее часто используемым параметром для оценки процессов обучения и запоминания во всех модификациях протокола проведения “Водного лабиринта Морриса” является время, за которое подопытное животное находит платформу. Этот параметр обычно называют временем задержки, или латентным временем поиска платформы. Он является самым простым и легко регистрируемым параметром, для которого необходимо только наличие таймера, измеряющего время от начала плавания животного до нахождения им платформы. В этом случае регистрация траектории плавания грызуна, скорости и пройденного им расстояния не обязательна, и на данный момент многие исследователи из-за недоступности трекинговых программ вынуждены ограничивать свои работы только этим параметром. Однако для более объективной оценки поведения и различных когнитивных процессов латентного времени поиска платформы недостаточно, и многие ученые отмечали, что на само время задержки влияют другие переменные, кроме мнемонических функций, и оно может варьироваться по причинам, не связанным со способностью животного к обучению [32, 36, 46]. Кроме того, использование данного параметра невозможно при пробных испытаниях, когда платформы вообще нет в бассейне. Некоторые современные исследования стали использовать вместо латентного времени нахождения животным платформы длину проделанного им пути, что стало возможным с использованием трекинговых программ, отслеживающих движение животного. Однако и этот параметр сильно зависим от индивидуальных особенностей локомоторных функций животного.
Вторым по популярности использования параметром является процентное время плавания грызуна в квадранте, где находится спрятанная платформа. Чаще данную меру используют в пробных испытаниях, когда после обучения платформа из бассейна изымается. Также вместо квадранта нередко используют круговую зону вокруг места, где стояла платформа. Равносильным этому параметру считают количество пересечений животным зоны, где раньше находилась поверхность платформы. Эти меры более характерны для целевого поиска местоположения, но они все же не являются достаточно надежными. Например, две крысы могут проводить одно и то же время в целевом квадранте, но одна может искать более близко к фактическому местоположению платформы [36, 46]. Аналогично, крыса может искать в узком круге очень близко к месту локализации платформы, но не может пересечь это место. Кроме того, для этих параметров недостаточно вариабельности внутри испытуемой группы животных, необходимое для надежного корреляционного анализа с другими поведенческими или нейробиологическими маркерами.
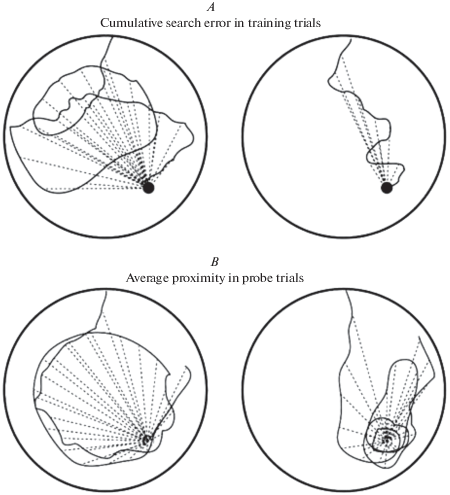
Чтобы лучше охарактеризовать стратегию поиска, которую животное использует в обучающих испытаниях, был введен индекс обучения (learning index или близость Галлахера, Gallagher’s proximity) [37] – средняя мера близости, которая опирается на постоянную оценку расстояния между местоположением платформы и животным, в пробных испытаниях – между животным и местом, где платформа раньше находилась (рис. 3 А). В оригинальной статье 1993 г. близость вычислялась десять раз в секунду, а среднее значение принималось каждую секунду. В настоящее время повышения частоты вычисления и, соответственно, качественного измерения можно добиться с использованием компьютерного анализа. Из этого параметра можно получить дополнительную переменную – кумулятивную ошибку поиска [36] (рис. 3B). В обучающих или пробных испытаниях кумулятивная ошибка поиска вычисляется путем суммирования средней секундной близости животного к платформе. Кумулятивная ошибка поиска является более чувствительным показателем пространственной памяти. Расстояние между начальной точкой плавания грызуна и местоположением платформы иногда вычитается из кумулятивной ошибки поиска, чтобы нормализовать параметр по различным начальным местоположениям. Чувствительность этих параметров позволяет лучше определить даже небольшие различия в поведении.
Рис. 3.
Примеры обучающих (А) и пробных (B) испытаний с использованием расчета индекса обучения и кумулятивной ошибки поиска. Сплошные линии – траектория движения грызуна. Пунктирные линии – средние ежесекундные значения близости животного к месту расположения платформы. Fig. 3 . Examples of training (A) and probe (B) trials using the calculation of the learning index and cumulative search error. Solid lines – the trajectory of the rodent. Dotted lines – 1-s averages of the proximity of the animal to the platform location.
Также для контроля динамики обучения подопытных животных можно воспользоваться индексами приобретения (acquisition index) и сохранения (savings index) [47, 48]. Индекс приобретения – мера обучения в течение дня, высчитанная с использованием первого и последнего латентного времени поиска платформы в испытаниях обучающей сессии, и усредненная за все дни обучения. Индекс сохранения – мера консолидации и сохранения памяти животного, основанная на вычислении разности латентного времени между последним испытанием данного дня обучения и первого испытания следующего дня, усредненное по всем дням.
Еще одним интересным и показательным параметром для оценки поведения животного в “Водном лабиринте Морриса” является ошибка Уишоу (Whishaw’s e-rror) – отклонение направления плавания грызуна от корректного направления к платформе [49, 50]. Для расчета данного параметра строится воображаемый коридор с произвольной шириной, которая определяется исследователем. Центральная линия этого коридора является прямой, соединяющей начальное положение испытуемого животного с местоположением платформы (рис. 4 А). Рассчитывается процент времени, проведенного животным в воображаемом коридоре. Основываясь на данном параметре, можно разделить испытания на удачные, в которых животное не выходило за пределы коридора, и неудачные, что упрощает итоговое сравнение экспериментальных групп.
Рис. 4.
Визуализация расчета ошибки Уишоу (А) и ошибки углового направления (B). Fig. 4 . Visualization of calculation of Whishaw’s error (А) and heading angle error (B).
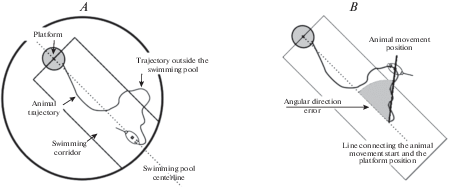
Ошибка углового направления (heading angle error) – достаточно эффективный, но редко используемый параметр. Он определяется как величина угла между направлением движения животного в определенной точке и прямым путем из стартового положения до платформы (рис. 4B). Иногда вместо первоначального положения животного сравнивают изменение угла ориентации животного в пространстве относительно платформы между каждой первой и второй секундой испытания [51]. Задержка в 1 с необходима, чтобы животное имело возможность поменять курс своего плавания. Но чаще всего используется однократное измерение ошибки углового направления на каждое испытание, для чего экспериментатором задается определенное условие положения животного, например, 10 см пройденного пути, 10% процентов от него, или местоположение грызуна через 10 с от начала движения [52].
С момента создания “Водного лабиринта Морриса” были предложены различные методы анализа полученных с его помощью данных, в том числе и сложные методы классификации траекторий плавания животных по поведенческим классам, известным как поисковые стратегии (или стратегии исследования). Методы классификации обеспечивают дополнительное понимание различных типов поведения животных, но только ограниченное число исследователей использует их [53]. Также классификация целых траекторий может привести к потере важной информации [44], так как не дает возможности выявить животных со смешанным поведением, поэтому наиболее качественный метод выявления стратегий исследования заключается в разделении траектории на сегменты, а затем классификации этих сегментов в поведенческие стратегии, часть из которых будет повторяться, пропадать или появляться в каждом из последующих испытаний. Таким образом, можно отследить изменения в поведении животных с каждым испытанием, а путь плавания животных в целом разделяется на более чем одну стратегию, раскрывая, как изменяется их поведение. Кроме того, кумулятивную ошибку поиска, ошибку Уишоу и ошибку углового направления можно отнести к и-ндивидуальным особенностям поведения испытуемого животного, и на их основании вывести стратегию поиска платформы. Пример определения стратегий поиска представлен на рис. 5 [53]. Эффективная и точная классификация подразумевает полное отсутствие ручной настройки и должна опираться только на компьютерные алгоритмы обработки данных, полученных в результате проведения испытаний. Поэтому наличие качественного программного обеспечения, способного измерить параметры более сложные, чем латентное время поиска платформы, является актуальной проблемой современных нейробиологических исследований.
Рис. 5.
Качественное определение стратегии поиска платформы подопытного грызуна в поведенческом тесте “Водный лабиринт Морриса”. А – определение различных качественных аспектов траектории плавания, необходимых для различия стратегий друг от друга. B – примеры различных стратегий поиска. Fig. 5 . Qualitative determination of the search platform strategies of the experimental rodent in the behavioral test “Morris Water Maze”. A – Identification of the various qualitative aspects of a trial’s swim path needed to distinguish the strategies from each other. B – Examples of the various search strategies.

Таблица 2.
Параметры “Водного лабиринта Морриса” Table 2 . Parameters of “Morris Water Maze”
| Параметры Parameters |
Определение Definition |
|---|---|
| Латентное время (время задержки) Latency |
Время, за которое подопытное животное находит платформу [1, 32] The time during which the experimental animal finds the platform [1, 32] |
| Процентное время плавания в квадранте Percentage swimming time in the quadrant |
Процентное время плавания грызуна в воображаемом квадранте, где находится (или находилась)
спрятанная платформа [46] Percentage swimming time of the rodent in the imaginary quadrant where the hidden platform is (or was) [46] |
| Количество пересечений
зоны платформы The number of platform area crossings |
Количество пересечений подопытным животным зоны, где раньше находилась платформа [46] The number of crossings by the experimental animals of the area where the platform was previously located [46] |
| Индекс обучения (близость Галлахера) Learning Index (Gallagher’s proximity) |
Средняя мера близости, которая опирается на постоянную оценку расстояния между местоположением
платформы и животным [37] An average measure of proximity that relies on a constant estimate of the distance between the platform location and the animal [37] |
| Кумулятивная ошибка поиска Cumulative search error |
Вычисляется путем суммирования средней секундной близости животного к платформе [36] Calculated by summing the average second proximities of an animal to the platform [36] |
| Индекс приобретенияAcquisition index | Разность между латентным временем поиска платформы в первом и последнем испытаниями тренировочной сессии, усредненное по всем дням обучения [47, 48]The difference between the latency in the first and last trials of the training session, averaged over all training days [47, 48] |
| Индекс сохранения Savings index |
Разность между латентным временем поиска платформы в последнем испытании первого дня
и первом испытании следующего дня, усредненное по всем дням обучения [47, 48] The difference between the latency in the last trial of the one day and the first trial of the next day, averaged over all training days [47, 48] |
| Ошибка Уишоу Whishaw's error |
Угол отклонения направления плавания грызуна от прямого пути от стартовой позиции
к платформе, который отражает корректность траектории движения животного по отношению
к платформе [49, 50] The deviation angle of the swimming direction of the rodent from the straight path from the starting position to the platform, which reflects the correctness of the animal trajectory in relation to the platform [49, 50] |
| Ошибка углового направления Heading angle error |
Величина отклонения угла между ориентацией животного в определенной точке своего плавания
в бассейне и прямым путем начала движения животного к платформе. Одна из вариаций
– сравнение не начального положения животного, а между каждой первой и второй секундой
испытания [51] The value of the angle deviation between the animal’s orientation at a certain point of its swimming in the pool and the direct path of the animal’s beginning of movement towards the platform. One of the variations is to compare not the initial position of the animal, but between every first and second seconds of the trial [51] |
| Поисковые стратегии
(стратегии исследования) Search strategies (research strategies) |
Сложные методы классификации траекторий плавания животных по поведенческим классам
[44, 53] (рис. 5
) Complex methods of classification of animal swimming trajectories by behavioral classes [44, 53] (fig. 5 ) |
Краткая информация о рассмотренных в этой главе параметрах оценки когнитивных функций в поведенческом тесте “Водный лабиринт Морриса” представлена в табл. 2 .
АППАРАТУРА И УСЛОВИЯ
Аппаратура, используемая при проведении теста “Водный лабиринт Морриса”, имеет широкий диапазон разных конфигураций: круглый бассейн может быть сделан из различных материалов, диаметр его 0.80–2.20 м, высота 0.40–0.60 м, при этом уровень воды устанавливается на высоте 0.25–0.40 м над основанием, температура воды в бассейне 18–26°C, а размеры платформы, на которую грызуны должны взбираться из воды, 9 × 9–13 × 13 см или, если она круглая, – диаметром 9–13 см. В первых своих экспериментах Моррис использовал большой круглый бассейн из ДВП, покрытый изнутри стекловолокном [1]. Размеры бассейна составляли 1.30 м в диаметре и 0.60 м в высоту, а уровень воды устанавливался на высоте 0.40 м над основанием. Температура воды в бассейне поддерживалась на уровне 26 ± 1°C. Бассейн помещали на опору с металлическим каркасом таким образом, чтобы уровень воды был 0.80 м над уровнем пола, а весь аппарат помещали в центр лабораторного помещения размером 3 × 4 м. Видеокамера с широкоугольным объективом была установлена над центром бассейна, и изображение передавалось на записывающее оборудование в соседней комнате.
Определенная постоянная температура воды также является очень важным условием проведения данного теста. Существует обратная U-образная зависимость, связывающая стресс с когнитивными функциями, согласно этой зависимости температура воды в бассейне на 13°C ниже температуры тела животного (обычно около 38°C) является достаточным стимулом для грызуна, чтобы начать активно искать скрытую платформу, но недостаточным стрессовым фактором, чтобы препятствовать функции обучения [54]. При понижении температуры воды до 19°C когнитивные способности испытуемого животного увеличиваются, но уменьшаются стимулирующие условия для нахождения спасательной платформы, а при достижении 12°C, стрессовые условия становятся превалирующими над когнитивными функциями грызуна.
Бассейн должен быть размещен в комнате с достаточным количеством визуальных ориентиров. Эти дистальные сигналы могут быть преднамеренными (например, использование ключей) или эндогенными для комнаты, при этом очень важно, чтобы сигналы не перемещались во время тестирования, поскольку они являются навигационными ориентирами животного для поиска цели. Ключи могут размещаться как в некотором отдалении от бассейна, так и непосредственно на его стенках над водой, что облегчает обучение животных. Расстояние сигналов от внешней стены лабиринта также может иметь важное значение [55]. Полезным дополнением являются шторы вокруг бассейна, с их помощью можно контролировать наличие или отсутствие необходимых дистальных сигналов, а также регулировать освещенность [5]. Освещаться помещение с бассейном должно рассеянным светом без преобладания одной конкретной точки – источника света, тем более, если производится запись эксперимента для дальнейшей обработки программным обеспечением. Камера и программное обеспечение для отслеживания чувствительны к отражениям света от поверхности воды, бликам и пересветам. На отражающие способности воды и блики может также повлиять степень замутненности воды в бассейне. На данный момент существует много способов приведение воды в непрозрачное состояние – разведение сухого молока или сухих сливок, добавление мела, водорастворимой нетоксичной краски или специальных красителей [18, 56].
Влияние экспериментатора в MWM главным образом заключаются в контакте с испытуемым животными и эффекте видимости экспериментатора. Влияние данных факторов на чистоту проводимого эксперимента и качество получаемых результатов аналогично, как и для любого поведенческого теста. Экспериментатор может: (а) покинуть комнату во время тестирования, что не всегда выполнимо; (б) стоять за визуальным барьером, например, шторами, либо сидеть на стуле, оставаясь ниже верхнего края стенки бассейна, будучи таким образом невидимым для животного; или (в) оставаться в фиксированном положении. Последнее является наиболее распространенным, но не следует забывать, что экспериментатор становится одним из дистальных сигналов. У животных, которые тестируются в разных квадрантах, особенно в дистальной и проксимальной половине лабиринта, иногда наблюдаются разные показатели латентного времени, в зависимости от того, плывут ли они к экспериментатору или от него [57].
Для качественной записи и обработки эксперимента необходимо соответствующее оборудование. Существует целый ряд коммерческих поставщиков, предоставляющих системы видеосъемки и отслеживания, которые можно использовать для “Водного лабиринта Морриса”. Например, HVS Image, San Diego Instruments, Accuscan, View Point, Clever Systems, Noldus, Columbus Instruments, Watermaze Software, Coulbourn Instruments и другие. Обычно коммерческие системы видеосъемки включают обязательную покупку отслеживающего программного обеспечения.
Поведение животных обычно регистрируется либо вручную, либо полуавтоматически. Традиционно принято наблюдать за поведением животного в лабиринте; если исследователь считает, что воспроизводится определенная модель поведения, экспериментатор регистрирует данный факт, записывая его или вводя данные в программу регистрации событий [24]. Ручная запись поведения может быть реализована с относительно небольшими затратами, и для некоторых моделей поведения это может быть единственным способом обнаружения и записи их возникновения. Однако автоматическое наблюдение может обеспечить значительные преимущества. Поведение регистрируется более надежно, потому что компьютерный алгоритм работает таким же образом, но система не зависит от усталости или невнимательности наблюдателя. Например, в отличие от ручного наблюдения автоматизированное отслеживание выполняет шаблонный анализ видеоизображения исследуемых животных, чтобы извлечь количественные измерения их поведения. Автоматическое наблюдение с использованием видеонаблюдения особенно подходит для измерения двигательной активности и локомоторной функции, выраженной в пространственных измерениях (расстояние, скорость, поворот и т.д.), которые наблюдатель-человек не может точно оценить [58].
В настоящее время поведенческий тест “Водный лабиринт Морриса” практически не используется без автоматизированного анализа полученных данных, так как единственный параметр, который возможно зарегистрировать вручную – это латентное время нахождения платформы. А он, как уже было упомянуто, является малоинформативным и упускает многие показательные аспекты сложного поведения животных. Поэтому автоматизация теста практически неизбежна. Такая компьютеризированная система должна обеспечивать быстрое и точное отслеживание с вычислением в реальном времени определенных параметров. У современного ученого есть два очевидных способа получить доступ к автоматизированному анализу MWM:
(I) Можно воспользоваться общедоступными программами, разработанными учеными, работающими в этой области, и выложенными ими для безвозмездного пользования любого желающего, например, Tracking [13], MouBeAT [58] или ToxTrac [59]. Плюсы такой программы – абсолютно простой и незатратный способ приобретения. А вот к минусам можно отнести возможное низкое качество анализа, сложный для обычного пользователя интерфейс, избирательную специализацию и низкую апробацию таких программ. Также для некоторых общедоступных опций программного обеспечения требуется лицензия MATLAB [60], что приводит к увеличению стоимости, занимает намного больше времени или ограничено операционными системами. Существует набор инструментов для поведенческого анализа мыши с открытым исходным кодом (MouBeAT) [58], основанный на программном обеспечении для анализа изображений ImageJ. Нами была сделана попытка воспользоваться одной из наиболее популярных и высоко оцениваемых общедоступных трекинговых программ – ToxTrac, которая была разработана группой шведских ученых в 2017 г. [59]. Но, к сожалению, при обработке отснятых нами видеофайлов траектория плавания подопытного животного не всегда определялась правильно, и нам не удалось настроить данную программу на безошибочное определение трека. Поэтому от использования данного программного продукта мы были вынуждены отказаться.
(II) Можно воспользоваться проверенной и удобной коммерческой программой. Такие программы обычно очень просты в использовании, имеют интуитивный интерфейс, заточены на конкретный поведенческий тест, в том числе на различные модификации “Водного лабиринта Морриса”, и часто дополнительно имеют подробную инструкцию от производителя. Примерами таких коммерческих автоматизированных программ для анализа поведения у грызунов могут служить EthoVision [24], Smart Video Tracking Software (www.panlab.com), VideoTrack (www.viewpoint.fr) и AnyMaze (www.stoeltingco.com). Значительный недостаток у них – очень высокая стоимость. Большинство коммерческих продуктов продаются в виде аппаратного и программного пакета, что делает их недоступными для многих лабораторий.
Ввиду перечисленных проблем, нами было принято решение воспользоваться третьим вариантом – разработать программное обеспечение самостоятельно. Первые полученные нами результаты свидетельствуют о том, что разработанная нами программа Minopontikos не уступает популярным коммерческим продуктам [52]. Мы рассчитываем, что в перспективе разработанная нами программа, способная высококачественно анализировать такие параметры, как латентное время, процентное время в квадрантах, скорость движения, длина трека, близость Галлахера, кумулятивная ошибка поиска, ошибка Уишоу, ошибка углового направления и различные стратегии поиска, может стать мощным инструментом анализа поведенческого теста MWM.
ЗАКЛЮЧЕНИЕ
В данной статье были рассмотрены различные модификации процедур проведения “Водного лабиринта Морриса”, а также перспективы его использования в лабораторной практике и результаты, которые можно получить с его помощью. Первоначальная задача MWM – это анализ пространственной памяти у подопытных грызунов, однако после изобретения данного теста в 80-х годах прошлого столетия его развитие и совершенствование не стоит на месте, и сейчас при помощи “Водного лабиринта Морриса” можно решать задачи, выходящие за рамки первоначальной. Изучение особенностей индивидуального поведения, поисковых стратегий – это относительно новые и малоизученные информационные возможности, которые дает MWM. Отчасти это проблема обусловлена отсутствием доступного программного обеспечения, способного качественно обрабатывать полученные данные. Без сомнения, популярные коммерческие продукты доказали свою эффективность, однако и у них есть ограничения по информативности – предварительное знакомство с некоторыми из них показало, что из перечисленных нами в данной статье сложных параметров единицы способны к полноценному их анализу, но в большинстве своем ограничиваются латентным временем, процентным временем в квадранте и некой угловой ошибкой (упрощенный вариант ошибки углового направления, для которой вычисление происходит только в одном положении из всей траектории подопытного животного). Таким образом, разработка качественного программного продукта для эффективного автоматизированного анализа “Водного лабиринта Морриса” все еще является актуальной задачей современной нейронауки.
Список литературы
Morris R.G.M. Spatial localization does not require the presence of local cues. Learning and Motivation. 12 (2): 239–260. 1981.
Morris R.G.M. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods. 11 (1): 47–60. 1984.
Sutherland R.J., Whishaw I.Q., Kolb B. Contributions of cingulate cortex to two forms of spatial learning and memory. J. Neurosci. 8 (6): 1863–1872. 1988.
Brandeis R., Brandys Y., Yehuda S. The use of the Morris Water Maze in the study of memory and learning. Int. J. Neurosci. 48(1–2): 29–69. 1989.
Vorhees C.V., Williams M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 1(2): 848–858. 2006.
Buresova O., Krekule I., Zahalka A., Bures J. On-demand platform improves accuracy of the Morris water maze procedure. J. Neurosci. Methods. 15(1): 63–72. 1985.
Whishaw I.Q., Mittleman G. Visits to starts, routes and places by rats (Rattus norvegicus) in swimming pool navigation tasks. J. Compar. Psychol. 100. 422–443. 1987.
Nakazawa K., Quirk M.C., Chitwood R.A., Watanabe M., Yeckel M.F., Sun L.D., Kato A., Carr C.A., Johnston D., Wilson M.A., Tonegawa, S. Requirement for hippocampal CA3 NMDA receptors in associative memory recall. Science. 297 (5579): 211–218. 2002.
Brun V.H., Otnass M.K., Molden S., Steffenach H.A., Witter M.P., Moser M.B., Moser E.I. Place cells and place recognition maintained by direct entorhinal-hippocampal circuitry. Science. 296 (5576): 2243–2246. 2002.
Kallai J., Makany T., Karadi K., Jacobs W.J. Spatial orientation strategies in Morris-type virtual water task for humans. Behav. Brain Res. 159(2): 187–196. 2005.
Morris R.G.M. An attempt to dissociate ‘spatial-mapping’ and ‘working-memory’ theories of hippocampal function. Neurobiology of the Hippocampus. New York. Acad. Press. 1993.
Lindner M.D., Balch A.H., VanderMaelen C.P. Short forms of the “reference-” and “working-memory” Morris water maze for assessing age-related deficits. Behav. Neural. Biol. 58 (2): 94–102. 1992.
Smolensky I.V., Zubareva O.E., Kalemenev S.V., Lavrentyeva V.V., Dyomina, A.V., Karepanov A.A., Zaitsev A.V. Impairments in cognitive functions and emotional and social behaviors in a rat lithium-pilocarpine model of temporal lobe epilepsy. Behav. Brain Res. 372 112044. 2019.
Crawley J.N. Behavioral phenotyping strategies for mutant mice. Neuron. 57(6): 809–818. 2008.
Nunn J.A., LePeillet E., Netto C.A., Hodges H., Gray J.A., Meldrum B.S. Global ischaemia: hippocampal pathology and spatial deficits in the water maze. Behav. Brain Res. 62 (1): 41–54. 1994.
Gallagher M., Rapp P.R. The use of animal models to study the effects of aging on cognition. Annu. Rev. Psychol. 48. 339–370. 1997.
Hsiao K., Chapman P., Nilsen S., Eckman C., Harigaya Y., Younkin S., Yang F. Cole G. Correlative memory deficits, Abeta elevation, and amyloid plaques in transgenic mice. Science. 274(5284): 99–102. 1996.
Bromley-Brits K., Deng Y., Song W. Morris water maze test for learning and memory deficits in Alzheimer’s disease model mice. J. Vis. Exp. (53): 2011.
Edwards S.R., Hamlin A.S., Marks N., Coulson E.J., Smith M.T. Comparative studies using the Morris water maze to assess spatial memory deficits in two transgenic mouse models of Alzheimer’s disease. Clin. Exp. Pharmacol. Physiol. 41(10): 798–806. 2014.
Shariatpanahi M., Khodagholi F., Ashabi G., Bonakdar Yazdi B., Hassani S., Azami K., Abdollahi M., Noorbakhsh F., Taghizadeh G., Sharifzadeh M. The involvement of protein kinase G inhibitor in regulation of apoptosis and autophagy markers in spatial memory deficit induced by Abeta. Fundam. Clin. Pharmacol. 30(4): 364–375. 2016.
Wu M.N., Zhou L.W., Wang Z.J., Han W.N., Zhang J., Liu X.J., Tong J.Q., Qi J.S. Colivelin ameliorates amyloid beta peptide-induced impairments in spatial memory, synaptic plasticity, and calcium homeostasis in rats. Hippocampus. 25(3): 363–372. 2015.
Ge M., Zhan Y., Hao Q., Zhao Y., Dong B. Effects of mesenchymal stem cells transplantation on cognitive deficits in animal models of Alzheimer’s disease: A systematic review and meta-analysis. Brain Behav. 8(7): e00982. 2018.
D'Hooge R., De Deyn P.P. Applications of the Morris water maze in the study of learning and memory. Brain Res. Brain Res. Rev. 36(1): 60–90. 2001.
Noldus L.P., Spink A.J., Tegelenbosch R.A. EthoVision: a versatile video tracking system for automation of behavioral experiments. Behav. Res. Methods Instrum. Comput. 33(3): 398–414. 2001.
Spink A.J., Tegelenbosch R.A., Buma M.O., Fursenko D.V., Kulikov A.V. Conducting and automating Morris water maze test under SPF conditions. Russ. J. Genet. Appl. Res. 6. 394–399. 2016.
Khotskin N.V., Kulikov V.A., Zavyalov E.L., Fursenko D.V., Kulikov A.V. Conducting and automating Morris water maze test under SPF conditions. Russ. J. Genet. Appl. Res. 6: 394–399. 2016.
Glaser O.C. The formation of habits at high speed. J. Compar. Neurol. Psychol. 20(3): 165–184. 1910.
Wever E.G. Water temperature as an incentive to swimming activity in the rat. J. Compar. Psychol. 14(2): 219–224. 1932.
Waller M.B., Waller P.F., Brewster L.A. A Water Maze for Use in Studies of Drive and Learning. Psychol. Rep. 7(1): 99–102. 1960.
Woods P.J., Davidson E.H., Peters R.J. Jr. Instrumental Escape Conditioning in a Water Tank: Effects of Variations in Drive Stimulus Intensity and Reinforcement Magnitude. J. Compar. Physiol. Psychol. 57. 466–470. 1964.
Woods P.J., Holland C.H. Performance on a black-white discrimination problem in water-maze as influenced by water temperature and swimming ability. Psychol. Reports. 9. 433–439. 1961.
Morgan D. Water Maze Tasks in Mice: Special Reference to Alzheimer’s Transgenic Mice. Methods Behav. Anal. Neurosci. 2. 14. Boca Raton (FL). 2009.
Bryan K.J., Lee H., Perry G., Smith M.A., Casadesus G. Transgenic Mouse Models of Alzheimer’s Disease: Behavioral Testing and Considerations. Methods of Behavior Analysis in Neuroscience. 2. 1. Boca Raton (FL). 2009.
Morris R.G.M. Morris water maze. Scholarpedia. 6315. 2008.
Kim J.J., Lee H.J., Han J.S., Packard M.G. Amygdala is critical for stress-induced modulation of hippocampal long-term potentiation and learning. J. Neurosci. 21(14): 5222–5228. 2001.
Tomas Pereira I., Burwell R.D. Using the spatial learning index to evaluate performance on the water maze. Behav. Neurosci. 129 (4): 533–539. 2015.
Gallagher M., Burwell R., Burchinal M. Severity of spatial learning impairment in aging: development of a learning index for performance in the Morris water maze. Behav. Neurosci. 107 (4): 618–626. 1993.
Westerman M.A., Cooper-Blacketer D., Mariash A., Kotilinek L., Kawarabayashi T., Younkin L.H., Carlson G.A., Younkin S.G., Ashe K.H. The relationship between Abeta and memory in the Tg2576 mouse model of Alzheimer’s disease. J. Neurosci. 22(5): 1858–1867. 2002.
Steele R.J., Morris R.G. Delay-dependent impairment of a matching-to-place task with chronic and intrahippocampal infusion of the NMDA-antagonist D-AP5. Hippocampus. 9(2): 118–136. 1999.
Spooner R.I., Thoson A., Hall J., Morris R.G., Salter S.H. The Atlantis platform: a new design and further developments of Buresova’s on-demand platform for the water maze. Learn. Mem. 1(3): 203–211. 1994.
Lipp H.P., Wolfer D.P. Genetically modified mice and cognition. Curr. Opin. Neurobiol. 8(2): 272–280. 1998.
Morris R.G., Hagan J.J., Rawlins J.N. Allocentric spatial learning by hippocampectomised rats: a further test of the “spatial mapping” and “working memory” theories of hippocampal function. Q. J. Exp. Psychol. B. 38 (4): 365–395. 1986.
Sutherland R.J., Dyck R.H. Place navigation by rats in a swimming pool. Canad. J. Psychol. 38. 322–347. 1984.
Vouros A., Gehring T.V., Szydlowska K., Janusz A., Tu Z., Croucher M., Lukasiuk K., Konopka W., Sandi C., Vasilaki E. A generalised framework for detailed classification of swimming paths inside the Morris Water Maze. Sci. Rep. 8 (1): 15089. 2018.
Morgan D., Diamond D.M., Gottschall P.E., Ugen K.E., Dickey C., Hardy J., Duff K., Jantzen P., DiCarlo G., Wilcock D., Connor K., Hatcher J., Hope C., Gordon M., Arendash G.W. A beta peptide vaccination prevents memory loss in an animal model of Alzheimer’s disease. Nature. 408(6815): 982–985. 2000.
Maei H.R., Zaslavsky K., Teixeira C.M., Frankland P.W. What is the Most Sensitive Measure of Water Maze Probe Test Performance? Front. Integr. Neurosci. 3 4. 2009.
Whiting M.D., Kokiko-Cochran O.N. Assessment of Cognitive Function in the Water Maze Task: Maximizing Data Collection and Analysis in Animal Models of Brain Injury. Methods Mol. Biol. 1462. 553–71. 2016.
Kokiko-Cochran O., Ransohoff L., Veenstra M., Lee S., Saber M., Sikora M., Teknipp R., Xu G., Bemiller S., Wilson G., Crish S., Bhaskar K., Lee Y.S., Ransohoff R.M., Lamb B.T. Altered Neuroinflammation and Behavior after Traumatic Brain Injury in a Mouse Model of Alzheimer’s Disease. J. Neurotrauma. 33 (7): 625–40. 2016.
Whishaw I.Q. Cholinergic receptor blockade in the rat impairs locale but not taxon strategies for place navigation in a swimming pool. Behav. Neurosci. 99 (5): 979–1005. 1985.
Harker, K.T. and Whishaw, I.Q. Impaired spatial performance in rats with retrosplenial lesions: importance of the spatial problem and the rat strain in identifying lesion effects in a swimming pool. J. Neurosci. 22 (3): 1155–1164. 2002.
Bye C.M., Hong N.S., Moore K., Deibel S.H., McDonald R.J. The effects of pool shape manipulations on rat spatial memory acquired in the Morris water maze. Learn. Behav. 47 (1): 29–37. 2019.
Chernyuk D.P., Zorin A.G., Derevtsova K.Z., Efimova E.V., Prikhodko V.A., Sysoev Y.I., Vlasova O.L., Bolsunovskaia M.V., Bezprozvanny I.B. Automatic analysis of the “Morris Water Maze” behavioral test data. I.P. Pavlov J. Higher Nerv. Activity. 71(1): 121–130. 2021.
Rogers J., Churilov L., Hannan A.J., Renoir T. Search strategy selection in the Morris water maze indicates allocentric map formation during learning that underpins spatial memory formation. Neurobiol. Learn. Mem. 139. 37–49. 2017.
Morris R. Neurobiology of Learning and Memory. Neurosci. in the 21st Century. 2173–2211. 2013
Maurer R., Derivaz V. Rats in a transparent morris water maze use elemental and configural geometry of landmarks as well as distance to the pool wall. Spatial Cognit. Comput. 2 135–156. 2000.
Nunez J. Morris Water Maze Experiment. J. Vis. Exp. (19): 897. 2008.
Hodges H. Maze procedures: the radial-arm and water maze compared. Brain Res. Cogn. Brain Res. 3(3–4): 167–181. 1996.
Bello-Arroyo E., Roque H., Marcos A., Orihuel J., Higuera-Matas A., Desco M., Caiolfa V.R., Ambrosio E., Lara-Pezzi E., Gomez-Gaviro M.V. MouBeAT: A New and Open Toolbox for Guided Analysis of Behavioral Tests in Mice. Front. Behav. Neurosci. 12. 201. 2018.
Rodriguez A., Zhang H., Klaminder J., Brodin T., Andersson P.L., Andersson M. ToxTrac: A fast and robust software for tracking organisms. Methods Ecol. and Evolut. 9 (3): 460–464. 2018.
Aragao Rda S., Rodrigues M.A., de Barros K.M., Silva S.R., Toscano A.E., de Souza R.E., Manhaes-de-Castro, R. Automatic system for analysis of locomotor activity in rodents–a reproducibility study. J. Neurosci. Methods. 195(2): 216–221. 2011.
Дополнительные материалы отсутствуют.
Инструменты
Российский физиологический журнал им. И.М. Сеченова