

Российский физиологический журнал им. И.М. Сеченова, 2021, T. 107, № 8, стр. 955-972
Фоновые показатели мощности ЭЭГ как корреляты вербальных и невербальных компонентов креативности и интеллекта
О. М. Разумникова *
Новосибирский государственный технический университет
Новосибирск, Россия
* E-mail: razoum@mail.ru
Поступила в редакцию 17.01.2021
После доработки 23.02.2021
Принята к публикации 07.03.2021
Аннотация
Интенсивные исследования нейрофизиологических коррелятов креативности последних лет выявили связь фоновой активности мозга (в том числе активности DMN – “сети в режиме по умолчанию”) и показателей креативности. Однако не ясны пока специфические компоненты такой активности, определяющие высокий уровень вербальной или невербальной креативности, и значение интеллектуальных способностей для успешного решения экспериментального творческого задания. Баланс фоновой активности фронтальных и задних отделов коры может отражать индивидуальный стиль решения проблемы, в качестве индикаторов такого баланса рассматриваются осцилляции разных частотных диапазонов. В связи с этим нами был выполнен анализ частотно-пространственной организации фоновой ЭЭГ, который выявил различия в мощности дельта-, тета-, альфа 2- и бета 2-ритмов в группах, дифференцированных по показателям оригинальности ответов при тестировании вербальной и образной креативности. Более высоким креативным способностям соответствуют большие значения мощности низкочастотных биопотенциалов в передней части коры и снижение мощности альфа-ритма в задних отделах. “Преднастройка” активности коры к вербальной оригинальности проявляется преимущественно в височных и центрально-париетальных областях коры, а к образной – для париетально-окципитальных. Вклад зрительно-пространственного компонента интеллекта в связанные с образной креативностью изменения активности коры представлен в большей степени на частоте дельта- и альфа 2-ритма, а вклад вербального компонента интеллекта в “преднастройку” нейронных систем коры для вербальной креативности – на частоте тета- и бета 2-диапазона. Следовательно, анализ частотно-пространственной организации активности коры головного мозга может быть полезным инструментом для выявления роли интеллектуальных способностей и эмоционально-мотивационной регуляции при формировании разных стратегий достижения высокого уровня креативности.
Интенсивные исследования нейрофизиологических коррелятов креативности, особенно активно развернувшиеся в последние годы, показали информационное значение организации сетей покоя (или фоновой активности мозга) для успешного выполнения экспериментальных задач, требующих оригинального решения [1–5]. Особое внимание в этом направлении уделяется изучению функций фронто-париетальной системы (FPN), фронтальные отделы которой связывают с организацией исполнительного контроля при выборе оригинального ответа, а париетальные – с поиском разнообразных идей решения поставленной проблемы [1, 6–8]. Кооперация функций FPN и DMN (default mode network) рассматривается как основа индивидуального разнообразия стратегий поиска ответа при тестировании как вербальной, так и образной креативности [5, 9]. Динамичная реорганизация этих систем с включением регионарно широко представленных нейронных структур, в том числе левой средней височной извилины и слуховой системы, характерна для вербальной креативности [10]. Данные других исследований указывают, что FPN является медиатором связи вербальной креативности с активностью передней части DMN, а визуальной креативности – с ее задней частью [5] или с латеральными изменениями показателя функциональной связности в дорзолатеральной префронтальной коре [11].
Представление о доминировании правого полушария при решении экспериментальных творческих заданий (например, [12]) изменилось в ходе накопления данных о функциональной активности мозга. Показано, например, участие левой дорзолатеральной префронтальной коры [13] или левой передней сингулярной коры [14] при выборе оригинального решения в ходе тестирования невербальной креативности.
Следовательно, для понимания закономерностей формирования разных функциональных нейронных систем, обеспечивающих разные формы креативности, требуются дополнительные исследования.
Имеется множество доказательств функциональной реорганизации FPN и DMN как основы дальнейшей реализации разных когнитивных функций, в том числе и креативности, и интеллектуальных способностей [1, 15–19]. Специализация взаимодействия этих систем заключается в том, что интеллект отражает систему “легкого” достижения функционального состояния с включением правой верхней париетальной области при низких интеграционных способностях левой ретросплениальной коры, а креативность – систему “сложного” переключения связности корковых областей с центром в правой дорзолатеральной префронтальной коре при высоких интеграционных способностях сенсомоторной коры [20].
Для изучения функционального значения фонового состояния мозга используются не только его томографические, но и энцефалографические характеристики [11, 21–24]. Среди разных частотных диапазонов ЭЭГ чаще других рассматривается синхронизация/десинхронизация альфа-колебаний, отражающая процессы торможения/активации в нейронных сетях, связанные как с показателями креативности, так и интеллекта (IQ) [15, 21, 26, 27]. Показано, что вариативность флюидного IQ связана с обновлением рабочей памяти, тогда как предикторами креативности является не только этот компонент исполнительной системы, но и тормозные функции [25]. Баланс фоновой активности фронтальных и задних отделов коры может отражать индивидуальный стиль решения проблемы, в том числе предпочтение инсайтной или аналитической стратегии [25, 28, 29], причем в качестве индикаторов такого баланса рассматриваются как альфа-, так и низкочастотные дельта-, тета-, и высокочастотные бета-осцилляции [8, 23, 30–32]. Дельта-ритм представляет интерес для анализа его связи с креативностью, так как увеличение его мощности отражает подавление усвоенного доминантного поведения и указывает на эффективность нового обучения, а тета- и бета-осцилляции рассматриваются как энцефалографические корреляты функций DMN и FPC [30, 33].
Ранее нами было показано усиление взаимодействия нейронных ансамблей передних областей коры с задними отделами левого полушария у лиц, характеризующихся сравнительно высокими показателями интеллекта и креативности по сравнению с теми, кто обладал их меньшими значениями [34]. Дальнейшее исследование фоновой активности коры головного мозга показало, что фронтальные дельта- и бета-ритмы могут служить предикторами как невербальной креативности, так и вербального интеллекта [35].
Целью настоящего исследования стало выяснение частотно-пространственной специфики паттернов нейронных осцилляций, отражающей и вербальную, и невербальную креативность, и ее потенциальной связи со значениями вербальных и зрительно-пространственных компонентов интеллекта. Учитывая изложенные выше сведения, были сформулированы следующие гипотезы:
• Вербальные и зрительно-пространственные компоненты креативности и интеллекта имеют сходные ЭЭГ корреляты активности фронтальных областей коры.
• Вербальная креативность связана с активностью преимущественно височных областей коры, а невербальная – теменно-затылочных областей.
• Высоким показателям креативности соответствует “преднастройка” синхронизации низкочастотных и высокочастотных осцилляций ЭЭГ.
МЕТОДЫ ИССЛЕДОВАНИЯ
В исследовании принимали участие 37 человек (практические здоровые студенты, правши 18 ± 1.1 лет; 27 девушек и 10 юношей), не проходившие медикаментозного лечения. Исследование было одобрено Этическим комитетом Факультета гуманитарного образования и информированное согласие было получено у всех участников исследования в соответствии с этическими нормами Хельсинской декларации 2013 г.
Для определения вербального (IQv) и зрительно-пространственного (IQs) компонентов интеллекта использовали тест структуры интеллекта Амтхауэра: средние значения выполнения двух вербальных (субтесты 2 и 3) и двух невербальных заданий (субтесты 7 и 8). Для оценки образной креативности применяли субтесты Торренса “Круги” и “Незавершенные фигуры”, вербальной – методику Гилфорда “Необычное использование обычного предмета” и составление осмысленного предложения с включением трех слов-стимулов (существительных из отдаленных семантических категорий). Показатели оригинальности ответов при выполнении трех первых методик вычисляли на основе соответствующей базы данных как число, обратное количеству таких же идей [36]. Оригинальность предложений оценивали подготовленные эксперты, имеющие опыт работы с данной методикой (альфа Кронбаха для полученных оценок составила 0.82).
Регистрацию ЭЭГ в состоянии спокойного бодрствования с закрытыми глазами выполняли с помощью аппаратуры и программного обеспечения “Мицар-201” в 19-ти отведениях (Fp1, Fp2, F7, F3, Fz, F4, F8, T3, C3, Cz, C4, T4, T5, P3, Pz, P4, T6, O1, O2), расположенных согласно системе 10/20, с объединенным референтным ушным электродом. Частота дискретизации 500 Гц, полоса пропускания 0.5–70 Гц. Для анализа активности мозга выбирали 2-секундные безартефактные отрезки ЭЭГ (с перекрытием 50%) общей длительностью 60 с. Для каждого отведения методом быстрого преобразования Фурье вычисляли спектральную плотность ЭЭГ в шести частотных диапазонах: дельта (1–4 Гц), тета (4–7 Гц), альфа1 (7–10 Гц), альфа 2 (10–13 Гц), бета 1 (13–20 Гц) и бета 2 (20–30 Гц).
Для статистического анализа был использован пакет программ Statistica 13.3 (SN: JPZ912J057923CNET2ACD-K). Для нормализации показателей мощности ЭЭГ применяли натуральный логарифм их значений.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Анализ частотно-пространственной организации фоновой активности коры головного мозга выполняли на основе сравнения групп, которые были сформированы делением выборки согласно полученным средним значениям показателей креативности и интеллекта для каждого субтеста. Выделенные таким образом группы с высоким (Гр1) или (Гр0) низким значением, соответственно для вербальной ГрВК1 и ГрВК0 и невербальной ГрОК1 и ГрОК0 креативности и для вербального ГрIQv1 и ГрIQv0 и невербального ГрIQs1 и ГрIQs0 интеллекта, различались по этим классификационным параметрам (p < 0.01) и составу индивидов в группах (табл. 1).
Таблица 1.
Средние значения показателей оригинальности при тестировании образных и вербальных компонентов креативности и интеллекта в сформированных группах, различающихся их уровнем
| Группа | Образная | Вербальная | Интеллект | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| n | Круги | Фигуры | n | НИОП | СП | n | IQv | n | IQs | |
| Гр0 | 22 | 0.8 | 2.8 | 20 | 0.8 | 2.8 | 20 | 100.0 | 20 | 100.9 |
| Гр1 | 15 | 1.8 | 4.4 | 17 | 2.0 | 4.8 | 17 | 109.2 | 17 | 112.4 |
Дисперсионный анализ показателей мощности ЭЭГ в каждом из шести выделенных диапазонов выполняли с независимой переменной ГРУППА (ВК(2), ОК(2), IQv(2) и IQs(2)). Зависимыми переменными были разные регионы коры в соответствии с гипотезами исследования: для изучения вклада исполнительной системы FPN – фронтальные (Fp1, Fp2, F3, F4, F7, F8), DMN – расположенные по центральной линии (Fz, Cz, Pz) и центрально-париетальные (C3, C4, P3, P4) области, а для анализа значения вербальной или образной природы креативности и интеллекта – темпоральные (T3, T4, T5, T6) и париетально-окципитальные (P3, P4, O1, O2) области левого и правого полушария (т.е. переменные ОБЛАСТЬ и ЛАТЕРАЛЬНОСТЬ). Анализ ANOVA выполняли с поправкой Гринхауза–Гейсера. Для выяснения причин обнаруженных эффектов использовали плановое сравнение показателей и рost-hoc анализ данных с поправкой Бонферрони для множественных сравнений.
Рис. 1.
Регионарные изменения мощности дельта-ритма, связанные с уровнем креативности (a) и интеллекта (b). ГрВК1, ГрВК0, ГрОК1 и ГрОК0 – группы с высоким или низким уровнем вербальной и образной оригинальности соответственно, и ГрIQs1 и ГрIQs0 – высокого и низкого зрительно-пространственного интеллекта.
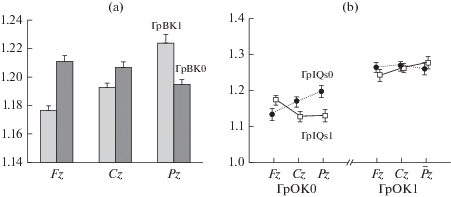
Обнаруженные эффекты, свидетельствующие о влиянии факторов ГРУППА или их взаимодействии (в том числе групп, различающихся по креативности и интеллекту), с переменными ОБЛАСТЬ и ЛАТЕРАЛЬНОСТЬ, сведены в табл. 2. Достоверные межгрупповые различия и регионарные особенности мощности биопотенциалов в каждом из шести диапазонов ЭЭГ, характеризующие Гр1, показаны на рис. 2.
Таблица 2.
Результаты анализа ANOVA с факторами ГРУППА, ОБЛАСТЬ, ЛАТЕРАЛЬНОСТЬ для разных частотных диапазонов
| Параметры | F | df | p | η | Эффект |
|---|---|---|---|---|---|
| Диапазон Дельта | |||||
| Область коры Фронтальные (Fp1, Fp2, F3, F4, F7, F8) | |||||
| ГрОК | 5.35 | 1.33 | 0.027 | 0.14 | ГрОК1 > ГрОК0 (1.24 ± 0.03 и 1.14 ± 0.03) |
| ГрIQv | 4.36 | 1.33 | 0.045 | 0.12 | ГрIQv1 < ГрIQv0 (1.13 ± 0.03 и 1.22 ± 0.03) |
| Центральные (Fz, Cz, Pz) | |||||
| ГрВК | 3.82 | 2.66 | 0.034 | 0.10 | ГрВК1: Fz < Pz. ГрВК0: Fz >Pz (рис. 1a) |
| ГрОК | 4.86 | 1.33 | 0.035 | 0.13 | ГрОК1 > ГрОК0 (1.26 ± 0.04 и 1.16 ± 0.03) |
| ГрОК × ГрIQs × ОБЛ | 4.05 | 2.66 | 0.022 | 0.11 | ГрОК0: Fz > Cz = Pz в ГрIQs1, но Fz < Cz < Pz в ГрIQs0; в ГрОК1 Fz = Cz = Pz независимо от IQs (рис. 1b) |
| Височные (T3, T4, T5, T6) | |||||
| ГрВК × ОБЛ | 6.67 | 1.33 | 0.014 | 0.17 | ГрВК0: T3, T4 < T5, T6 (0.83 ± 0.06 и 1.00 ± 0.07); ГрВК1: T3, T4 = T5, T6 (0.91 ± 0.07 и 0.92 ± 0.08) |
| Центрально-париетальные (C3, C4, P3, P4) | |||||
| ГрВК × ОБЛ | 6.36 | 1.33 | 0.017 | 0.16 | ГрВК0: C3, C4 < P3, P4 (1.06 ± 0.05 и 1.16 ± 0.06); ГрВК1: C3, C4 = P3, P4 (1.09 ± 0.06 и 1.11 ± 0.06) |
| ГрОК × ГрIQs × ОБЛ | 9.89 | 1.33 | 0.004 | 0.23 | ГрIQs0: P3, P4 > C3, C4 в ГрОК0; ГрIQs1: P3, P4 > C3, C4 в ГрОК1 (рис. 2) |
| ГрIQs × ОБЛ × ЛАТ | 4.39 | 1.33 | 0.044 | 0.12 | ГрIQs0: P4 > P3 (1.18 ± 0.04 и 1.06 ± 0.04) и C4 < C3 (1.09 ± 0.04 и 1.13 ± 0.04); ГрIQs1: P4 = P3 и C4 = C3 (1.13 ± 0.04 и 1.11 ± 0.04 и 1.08 ± 0.04 и 1.06 ± 0.04 соответственно) |
| Тета | |||||
| Фронтальные (Fp1, Fp2, F3, F4, F7, F8) | |||||
| ГрОК | 4.18 | 1.33 | 0.049 | 0.11 | ГрОК1 > ГрОК0 (0.70 ± 0.05 и 0.56 ± 0.04) |
| ГрВК × IQv × ОБЛ × ЛАТ | 3.99 | 2.66 | 0.023 | 0.11 | ГрIQv0: ГрВК1 > ГрВК0; IQv1: ГрВК1 < ГрВК0 во всех отведениях, за исключением F8 (рис. 2) |
| Центральные (Fz,Cz,Pz) | |||||
| ГрОК | 6.10 | 1.33 | 0.019 | 0.16 | ГрОК1 > ГрОК0 (0.94 ± 0.05 и 0.76 ± 0.05) |
| ГрВК × ОБЛ | 3.25 | 2.66 | 0.050 | 0.09 | ГрВК1: Cz > Pz. ГрВК0: Cz = Pz |
| ГрIQv × ОБЛ | 3.07 | 2.66 | 0.050 | 0.09 | ГрIQv0: Pz < Cz = Fz. ГрIQv1 Fz = Cz = Pz |
| Височные (T3, T4, T5, T6) | |||||
| ГрВК × ОБЛ | 4.52 | 1.33 | 0.041 | 0.12 | ГрВК0: T3, T4 < T5, T6; ГрВК1: T3, T4 = T5, T6 (рис. 2) |
| Центрально-париетальные (C3, C4, P3, P4) | |||||
| ГрОК | 4.31 | 1.33 | 0.046 | 0.12 | ГрОК1 > ГрОК0 (0.80 ± 0.06 и 0.63 ± 0.05) |
| ГрВК × ОБЛ | 8.35 | 2.66 | 0.007 | 0.20 | ГрВК0: C3, C4 < P3, P4; ГрВК1: C3, C4 > P3, P4 |
| ГрВК × ГрIQv × ОБЛ | 7.00 | 2.66 | 0.012 | 0.17 | ГрIQv0: C3, C4 > P3P4 в ГрВК0 и C3, C4 < P3, P4 в ГрВК1; ГрIQv1: C3, C4 = P3, P4 в ГрВК1 и C3, C4 < P3, P4 в ГрВК0 |
| Париетально-окипитальные (P3, P4, O1, O2) | |||||
| ГрОК × ОБЛ × ЛАТ | 6.51 | 1.33 | 0.016 | 0.16 | ГрОК1: O1 > O2. ГрОК0: O1 < O2 |
| Альфа 1 | |||||
| Височные (T3, T4, T5, T6) | |||||
| ГрВК × ОБЛ | 7.14 | 1.33 | 0.011 | 0.18 | ГрВК1 < ГрВК0 в T5, T6 (рис. 2); ГрВК0 = ГрВК1 в T3, T4 |
| Центрально-париетальные (C3, C4, P3, P4) | |||||
| ГрВК × ОБЛ | 5.05 | 1.33 | 0.031 | 0.13 | ГрВК1 < ГрВК0 в P3, P4 (рис. 2); ГРВК0: C3, C4 < P3, P4 |
| Париетально-окипитальные (P3, P4, O1, O2) | |||||
| ГрОК × ЛАТ | 4.45 | 1.33 | 0.043 | 0.12 | ГрОК1: P3, O1 > P4, O1 (рис. 2); ГрОК0: P3, O1 < Р4, O1 |
| ГрОК × ОБЛ × ЛАТ | 4.71 | 1.33 | 0.037 | 0.12 | ГрОК1: O1 > O2; ГрОК0: O1 < O2 |
| Альфа 2 | |||||
| Фронтальные (Fp1, Fp2, F3, F4, F7, F8) | |||||
| ГрОК | 4.10 | 1.33 | 0.050 | 0.11 | ГрОК1 > ГрОК0 (0.73 ± 0.07 и 0.55 ± 0.06) |
| Центральные (Fz,Cz,Pz) | |||||
| ГрОК | 4.77 | 1.33 | 0.036 | 0.13 | ГрОК1 > ГрОК0 (1.16 ± 0.08 и 0.93 ± 0.07) |
| Височные (T3, T4, T5, T6) | |||||
| ГрВК × ОБЛ | 4.67 | 1.33 | 0.038 | 0.12 | ГрВК1 > ГрВК0 в T3, T4; ГрВК1 < ГрВК0 в T5, T6 |
| Центрально-париетальные (C3, C4, P3, P4) | |||||
| ГрОК | 3.98 | 1.33 | 0.050 | 0.11 | ГрОК1 > ГрОК0 (1.21 ± 0.08 и 0.98 ± 0.07) |
| ГрОК × ГрIQs × ЛАТ | 5.55 | 1.33 | 0.025 | 0.14 | ГрОК1: C3, P3< C4, P4 в ГрIQs0 и C3, P3> C4, P4 в ГрIQs1 |
| ГрВК × ОБЛ | 4.03 | 1.33 | 0.050 | 0.11 | ГрВК1 > ГрВК0 в C3, C4, ГрВК1 = ГрВК0 в P3, P4 |
| Париетально-окипитальные (P3, P4, O1, O2) | |||||
| ГрОК × ГрIQs × ЛАТ | 4.26 | 1.33 | 0.047 | 0.11 | ГрОК1: O1, P3 < O2, P4 в ГрIQs0 и O1, P3 > O2, P4 в ГрIQs1 |
| ГрОК × ОБЛ × ЛАТ | 6.39 | 1.33 | 0.016 | 0.16 | ГрОК1: O1 > O2, ГрОК0: O1 < O2 |
| Бета 1 | |||||
| Центральные (Fz,Cz,Pz) | |||||
| ГрВК × ОБЛ | 6.97 | 2.66 | 0.002 | 0.17 | ГрВК0: Pz > Cz > Fz, ГрВК1: Pz > Cz = Fz |
| Бета 2 | |||||
| Височные (T3, T4, T5, T6) | |||||
| ГрОК × ЛАТ | 4.26 | 1.33 | 0.047 | 0.11 | ГрОК1: T3, T5 > T4, T6 ГрОК0: T3, T5 < T4, T6 |
| Центрально-париетальные (C3, C4, P3, P4) | |||||
| ГрВК × IQv × ЛАТ | 9.04 | 1.33 | 0.005 | 0.22 | ГрВК1: C3, P3 < C4, P4 в ГрIQv0 и C3, P3 > C4, P4 в ГрIQv1 |
| ГрВК × ОБЛ × ЛАТ | 4.33 | 1.33 | 0.045 | 0.12 | ГрВК0: С3 > С4, Р3 < Р4; ГрВК1: C3 = C4 и P3 = P4 |
| Париетально-окипитальные (P3, P4, O1, O2) | |||||
| ГрОК × ЛАТ | 10.55 | 1.33 | 0.003 | 0.24 | ГрОК1: P3, O1 > P4, O2; ГрОК0: P3, O1 < P4, O2 |
| ГрОК × ОБЛ × ЛАТ | 8.49 | 1.33 | 0.006 | 0.20 | ГрОК1: O1 > O2, ГрОК0: O1 < O2 |
ГрВК и ГрОК – группы с высоким или низким уровнем вербальной и образной оригинальности соответственно, ГрIQv и ГрIQs – высокого и низкого вербального и зрительно-пространственного интеллекта; ОБЛ и ЛАТ – зависимые переменные область коры и латеральность для мощности биопотенциалов в шести частотных диапазонах. В правом столбце таблицы представлено описание полученных эффектов.
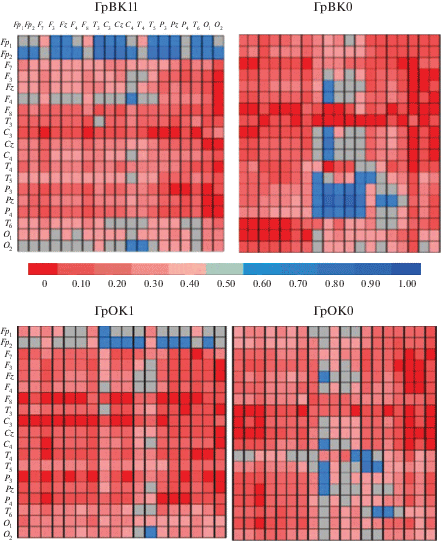
Рис. 2.
Карты разности мощности биопотенциалов при сравнении групп с высокой и низкой вербальной (ВК) и образной (ОК) креативностью в шести частотных диапазонах (delta–beta2). Белыми овалами отмечены области для общих эффектов ВК и ОК, кружками – отведения, для которых обнаружены регионарные различия в группах (в том числе регионарные эффекты для ГрВК1 и ГрОК1), стрелки указывают на области коры, для которых получен эффект взаимодействия креативности и соответствующего компонента интеллекта.

Post-hoc анализ приведенных в табл. 2 эффектов показал, что на частоте дельта-колебаний ГрОК1 (отличающаяся высокими значениями образной оригинальности) характеризуется большей фоновой мощностью дельта-ритма в передней части коры (отведения Fp1, Fp2, F3, F4, F7, F8 и Fz, Cz, Pz) по сравнению с ГрОК0 (пример этого эффекта для Fz, Cz, Pz показан на рис. 1a).
Для ГрВК1 обнаружено, напротив, снижение мощности дельта-ритма по оси Fz, Cz, Pz. Взаимодействие ГрВК × ОБЛ для височных и центрально-париетальных областей коры было связано с большей мощностью дельта-ритма в задневисочных и париетальных отведениях по сравнению с передне-височными и центральными в ГрВК0 при отсутствии достоверных различий для ГрВК1 (см. табл. 2).
Взаимодействие ГрОК × ГрIQs × ОБЛ для отведений Fz, Cz, Pz обусловлено разнонаправленными градиентами мощности дельта-ритма в ГрОК0: с его повышением в заднем направлении при низком интеллекте (ГрIQs0), но снижении при высоком (рис. 1b); при этом эффект большей мощности дельта-колебаний в ГрОК1 не зависел от уровня IQ. Взаимодействие ГрОК × ГрIQs × ОБЛ для центрально-париетальных областей было связано с тем, что ГрОК1 характеризовалась большими значениями дельта-ритма в париетальных по сравнению с центральными отведениями при высоких значениях IQs, сопровождающихся отсутствие асимметрии показателей мощности (рис. 2), которая отмечалась у лиц из ГрIQs0 (взаимодействие IQs × ОБЛ × ЛАТ).
В тета-диапазоне для ГрОК1 сохраняется эффект больших значений мощности биопотенциалов в сравнении с ГрОК0 для передних, центральных и центрально-теменных регионов коры (табл. 2, рис. 2). Латеральный эффект для ГрОК был представлен в затылочной коре: ГрОК1 характеризовалась большей левополушарной мощностью тета-ритма, а ГрОК0 – правополушарной.
На этой частоте выявлены регионарные эффекты фоновой активности коры для ГрВК в височных и центрально-париетальных областях коры: ГрВК1 характеризовалась большим уровнем тета-осцилляций в Сz, чем в Рz, а ГрВК0 – в передне-височных, чем в задневисочных отведениях. Связанные с вербальной креативностью регионарные особенности тета-ритма зависимы от уровня IQv во фронтальных и центрально-париетальных областях коры с более выраженным градиентом в ГрIQv0 (см. табл. 2). На рис. 2 стрелками указаны эти области, и отмечены отведения, для которых обнаружены различия между ГрВК1 и ГрВК0, регионарные особенности показателей мощности приведены для ГрВК1.
По альфа 1-активности ГрВК различались в височной и центрально-париетальной коре с меньшими значениями мощности в ГрВК1, чем ГрВК0, а ГрОК – асимметрией мощности альфа-биопотенциалов в парието-окципитальных областях при больших левополушарных значениях мощности в ГрОК1, но правополушарных для ГрОК0 (табл. 2, указаны отведения, для которых обнаружены межгрупповые различия для ГрВК1 и ГрВК0, и эффект полушарной асимметрии для ГрОК1).
В альфа 2-диапазоне ГрОК1 характеризовалась большим уровнем мощности биопотенциалов в передних областях коры, а в центральных, париетальных и окципитальных связанная с образной креативностью фоновая активация коры менялась зависимым от IQs образом: характеризующая ГрОК1 мощность альфа 2-ритма для ГрIQs0 была ниже в левом, чем в правом полушарии, а для ГрIQs1 – наоборот, в правом (на рис. 2 показана карта мощности с асимметрией альфа 2 для лиц, принадлежащих к ГрIQs1 из ГрОК1). ГрВК1 отличалась от ГрВК0 большим уровнем альфа 2-ритма в центральных и передне-височных отведениях, а в задневисочных – напротив, меньшим (на рис. 2 отмечены эти отведения, интенсивность темного цвета указывает на большие значения альфа в Гр1, чем в Гр0; светлого – наоборот, большие значения мощности биопотенциалов в Гр0).
В бета 1-диапазоне взаимодействие ГрВК × ОБЛ обусловлено более выраженным градиентом мощности биопотенциалов в ГрВК0 с большими значениями в Pz, чем Fz.
Обнаруженные эффекты для бета 2-диапазона связаны с фактором ЛАТЕРАЛЬНОСТЬ: для ГрОК – в височных и парието-окципитальных областях, а для ГрВК – в центрально-париетальных (табл. 2). ГрОК1 отличается большими значениями мощности левополушарного бета 2-ритма (рис. 2), а ГрОК0 – правополушарного; в ГрВК1 полушарная специфика латерального эффекта связана с уровнем IQv: доминирование левого полушария согласно большей мощности высокочастотных бета-биопотенциалов в ГрВК1 наблюдается при условии принадлежности к ГрIQv1 (рис. 2), а правого – к ГрIQv0. Взаимодействие ГрВК × ОБЛ × ЛАТ обусловлено разнонаправленным латеральным эффектом в мощности бета 2-колебаний в центральных и париетальных отведениях для ГрВК0 при отсутствии асимметрии в ГрВК1 (табл. 2).
Таким образом, полученные данные указывают, что большая часть различий в фоновой активности коры для групп, сформированных на основе разного уровня показателя оригинальности при тестировании креативности, представлена в низкочастотных дельта- и тета-диапазонах и высокочастотных альфа 2- и бета 2-диапазонах. Регионарная специфика фоновой ЭЭГ подтверждает гипотезу исследования: “преднастройка” к вербальной оригинальности преимущественно касается височных и центрально-париетальных областей коры, а к образной – париетально-окципитальных (см. рис. 2). Что касается градиента активности нейронных сетей в передне-заднем направлении, то можно заключить, что более высоким креативным способностям соответствуют большие значения низкочастотных биопотенциалов в передней части коры при относительно большей активации задних отделов на альфа-частоте.
Вклад IQs в связанные с образной креативностью изменения частотно-пространственной активности представлен на частоте дельта- и альфа 2-ритма, а вклад IQv в “преднастройку” нейронных систем коры для вербальной креативности – в тета- и бета 2-диапазонах.
Учитывая выявленную информативность дельта- и альфа2-биопотенциалов в дифференциации высокой и сравнительно низкой оригинальности ответов при тестировании креативности, нами выполнен корреляционный анализ показателей мощности этих ритмов для всех отведений биопотенциалов. Его результаты показаны на рис. 3.
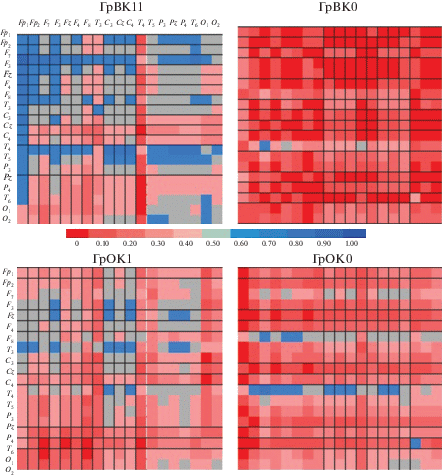
Рис. 3.
Карты корреляций между дельта- (отведения по вертикали) и альфа 2- (отведения по горизонтали) ритмами для групп, различающихся более высокой (ГрВК1-вербальной, ГрОК1-образной) или низкой (ГрВК1, ГрОК0) оригинальностью ответов.
Приведенные на рис. 3 карты корреляций указывают на разные формы организации взаимосвязи мощности дельта и альфа 2 в группах, различающихся уровнем вербальной или образной креативности (достоверные связи на приведенных картах начинаются для r > 0.45 при p < 0.05). Результаты межгруппового сравнения регионарных особенностей средних значений показателей корреляции свидетельствуют, что ГрВК1 отличается более выраженной положительной связью мощности дельта-колебаний в переднефронтальных отведениях и генерализованно представленного альфа 2 по сравнению с ГрВК0 (r = 0.62 и 0.24, p = 0.03), тогда как в ГрВК0 связь этих ритмов представлена в задних областях коры. Сравнительный анализ коэффициентов корреляции в отведениях C3, Cz, C4, P3, Pz, P4) показывает большие значения в ГрВК0, чем ГрВК1 (r = 0.55 и 0.23, p = 0.05). Такая же закономерность, но менее выраженная, прослеживается при сравнении корреляционных карт для ГрОК1 и ГрОК0 (r = 0.57 и 0.33, p = 0.10 и r = 0.44 и 0.22, p = 0.20 соответственно).
Аналогично выполненный корреляционный анализ тета- и бета 2-мощности выявил их тесную связь для ГрВК1: мощность тета-ритма в передней части коры, преимущественно в отведениях Fp1, F7, а также в T4 и T5, положительно связана с широко представленными по коре бета 2-осцилляциями (рис. 4, корреляции значимы при r > 0.50, p < 0.05). В ГрОК1 корреляции тета и бета 2 не такие высокие, в большей степени они представлены в центрально-париетальных отведениях. В ГрВК0 отмечены единичные связи показателей мощности этих ритмов, а в ГрОК0 – только корреляция тета- в Т4 и Р3 с диффузно представленной по коре мощностью бета 2-колебаний.
Рис. 4.
Карты корреляций между тета- (отведения по вертикали) и бета 2- (отведения по горизонтали) ритмами для групп, различающихся более высокой (ГрВК1-вербальной, ГрОК1-образной) или низкой (ГрВК1, ГрОК0) оригинальностью ответов.
Таким образом, фоновая активность мозга по-разному представлена в группах с высокой или низкой оригинальностью ответов при тестировании вербальной или образной креативности и связана с уровнем IQ.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Полученные в результате межгруппового сравнения различия в фоновой мощности ЭЭГ указывают на более широко частотно- и регионарно представленные эффекты, связанные с уровнем креативности, чем IQ. Действительно, данные многочисленных исследований креативности последних лет отражают необходимость интеграции широко распределенных нейронных сетей, охватывающих не только корковые области, но и подкорковые структуры [3, 6, 9, 37]. При формировании ГрОК и ГрВК были использованы показатели оригинальности дивергентного мышления в разных условиях тестирования креативности: требующих в первую очередь отказа от стереотипного ответа (Круги и НИОП), либо скорее способностей к формированию отдаленных ассоциаций (Фигуры и СП). Следовательно, сформированные группы различаются не только средним уровнем оригинальности ответов, но и стратегиями его поиска вследствие разных условий тестирования, влияние которых было показано нами ранее при исследовании возрастных особенностей креативности [38].
Взаимодействие нейронных ансамблей распределенной системы FPN и DMN как основы для генерации концептуально нового решения при тестировании креативности показано ранее с применением томографических методов [39]. Эффективную координацию таких ансамблей может обеспечивать синхронизация медленноволновой активности в дельта- и тета-диапазонах согласно нашим данным, а также результатам другого исследования, в котором отмечена положительная связь дельта-колебаний и оригинальности дивергентного мышления [40]. Имеются, однако, данные о противоположном эффекте: десинхронизации дельта-осцилляций при создании уникального образа [41]. Такие противоречия могут быть связаны с различиями в субъективно оцениваемой сложности задачи и соответствующих изменениях в балансе активационных и тормозных процессов в коре [34, 42].
Сравнительно более локально регионарно и частотно представленную связь фоновой ЭЭГ и IQ можно объяснить в соответствии с гипотезой нейронной эффективности [43], то есть точно скоординированной и соответственно экономно функционирующей нейронной сетью для извлечения логически верного ответа при тестировании IQ. Другой причиной может быть менее выраженная по сравнению с креативностью разница по IQ в Гр0 и Гр1 (по уровню IQ группы различаются примерно на 10%, тогда как по показателю оригинальности – в 1.5–2 раза).
Обнаруженные эффекты взаимодействия креативности и IQ (ГрОК × ГрIQs × ОБЛ, ГрВК × ГрIQv× ОБЛ, ГрВК × IQv × ОБЛ × ЛАТ или ГрВК × IQv × ЛАТ) были представлены в передней части коры для низкочастотных дельта- и тета-диапазонов, функциональное значение которых связывают с процессами мотивации и эмоциональной регуляции когнитивной деятельности [44–46], и в задних участках коры для высокочастотных альфа 2 и бета 2, которые ассоциируются с семантической обработкой информации [47–50]. Можно предположить, что связь интеллекта и креативности опирается на два разных процесса: исполнительный контроль деятельности со стремлением к ее лучшему результату (функции FPN) и способности к поиску отделенных ассоциаций с вовлечением обширных информационных ресурсов (функции DMN и париетальной системы внимания).
Разнонаправленные в ГрОК и ГрВК эффекты для дельта-колебаний в отведениях Fz, Cz, Pz (проекция DMN) можно объяснить большим вкладом вербальных интеллектуальных способностей в “преднастройку” нейронных сетей коры для организации дальнейшей когнитивной деятельности, сопровождающейся снижением активности DMN при ее повышении в FPN [51, 52]. В свою очередь обнаруженный разнонаправленный градиент мощности дельта-колебаний по оси Fz–Cz–Pz: с увеличением в заднем направлении для ГрВК1, но снижением для ГрВК0 (рис. 1A) с учетом регионарных особенностей взаимосвязи мощности дельта- и альфа2-ритмов (рис. 3) может отражать “преднастройку” к успешной креативной деятельности в ГрВК1 за счет ее мотивационного и исполнительного компонентов и соответствующей синхронизации дельта-биопотенциалов фронтальной коры и генерализованного по коре альфа 2-ритма (взаимосвязь систем FPN и DMN), тогда как в ГрВК0 такой эффект характерен для центрально-париетальных областей, что представляет, по-видимому, “автономное” состояние DMN. Подобие корреляционных карт для ГрВК1 и ГрОК1 как на частотах дельта–альфа 2, так и тета–бета 2 (рис. 3 и 4) можно рассматривать как подтверждение общего эффекта взаимодействия FPN и DMN, необходимого для поиска оригинального решения поставленной проблемы [1, 8, 9, 39].
ГрВК1 характеризуется более широко представленными и устойчивыми по сравнению с ГрОК1 паттернами корреляций низкочастотных и высокочастотных ритмов со сравнительно большим вовлечением височных отделов коры как специализированной основы вербальной креативности, тогда как анализ групп, различающихся образной креативностью, указывает на информативное значение функций задних отделов коры (см. рис. 2–4).
Обнаруженная на альфа 1,2-частоте большая синхронизация биопотенциалов в передней части коры для Гр1 по сравнению с Гр0 отражает, по-видимому, “преднастройку” фонового состояния мозга [29], ведущую к успешным процессам интернального внимания и торможения нерелевантной информации при дивергентном мышлении [26]. А эффект большей десинхронизации альфа-биопотенциалов в задних отделах коры применительно к вербальной или образной природе задания также подтверждает гипотезу исследования: вербальная креативность связана с изменениями активации преимущественно височных областей коры, а невербальная – теменно-затылочных областей (рис. 2).
Что касается эффекта ЛАТЕРАЛЬНОСТЬ, то на низких частотах тета- и альфа 1-диапазона он указывает на большую активацию правого полушария как предпосылку к образной креативности (табл. 2, рис. 2). В более высокочастотных альфа 2 и бета 2 выраженность полушарной асимметрии связана с IQ: для лиц из ГрВК1 и ГрIQv1 фоновая активность центрально-париетальных отделов коры характеризуется большими значениями бета 2-ритма в левом полушарии, а для представителей ГрОК1 и IQs1 – это левополушарное доминирование отмечается не только согласно показателям бета 2-, но и альфа 2-ритма. Следовательно, спорный вопрос о связанном с креативностью полушарном доминировании [12–14] можно решить, учитывая вклад разных компонентов интеллекта.
Таким образом, анализ частотно-пространственной организации активности коры головного мозга может быть полезным инструментом для выявления вклада интеллектуальных способностей и эмоционально-мотивационной регуляции стратегий достижения высокого уровня креативности. Обнаруженное в нашей работе информативное значение альфа- и бета 2-осцилляций при сравнении лиц, различающихся уровнем креативности, согласуется с данными классификации вербальной креативности на основе машинного обучения [53]. Оценка фоновой активности мозга и психометрических показателей разных компонентов интеллекта и креативности представляет особый интерес при выборе режима транскраниальной стимуляции для тренировки креативных способностей. В последнее время появилось множество таких работ (например, [54–56]), согласно результатам которых констатируется положительный эффект тренировки, однако механизм его достижения остается неясным [57], так как используются не только разные участки коры (чаще префронтальные области левого или правого полушария), но и совершенно разные частотные параметры воздействия на мозг.
ЗАКЛЮЧЕНИЕ
Связанные с креативностью особенности частотно-пространственной организации фоновой ЭЭГ в максимальной степени представлены в диапазоне дельта-, тета-, альфа 2-ритмов и бета 2-ритмов. Более высоким креативным способностям соответствуют большие значения низкочастотных биопотенциалов в передней части коры при относительно большей активации задних отделов на альфа-частоте. Связанная с дифференциацией креативности фоновая активность коры на частоте бета 2-диапазона отражается изменениями в доминировании активности левого или правого полушария. “Преднастройка” активности коры к вербальной оригинальности проявляется преимущественно в височных и центрально-париетальных областях коры, а к образной – в париетально-окципитальных.
Взаимодействие зрительно-пространственного компонента интеллекта и образной креативности представлено в большей степени в фоновой активности коры на частоте дельта- и альфа-ритма, а вербального компонента интеллекта и вербальной креативности – на частоте тета- и бета 2-диапазона.
Следовательно, определение индивидуальных характеристик фоновой частотно-пространственной организации активности коры головного мозга может быть полезно для выяснения ресурсов креативности и оптимальных стратегий инновационной деятельности.
Список литературы
Beaty RE, Benedek M, Wilkins RW, Jauk E (2014) Creativity and the default network: A functional connectivity analysis of the creative brain at rest. Neuropsychologia 64: 92–98. https://doi.org/10.1016/j.neuropsychologia.2014.09.019
Schuler A-L, Tik M, Sladky R, Luft CDB, Hoffmann A, Woletz M, Zioga I, Bhattacharya J, Windischberger C (2019) Modulations in resting state networks of subcortical structures linked to creativity. Neuroimage 195: 311–319. https://doi.org/10.1016/j.neuroimage.2019.03.017
Shi L, Sun J, Xia Y, Ren Z, Chen Q, Wei D, Yang W, Qiu J (2018) Large-scale brain network connectivity underlying creativity in resting-state and task fMRI: Cooperation between default network and frontal-parietal network. Biol Psychol 135: 102–111. https://doi.org/10.1016/j.biopsycho.2018.03.005
Sun J, Liu Z, Rolls ET, Chen Q, Yao Y, Yang W, Wei D, Zhang Q, Zhang J, Feng J (2019) Verbal creativity correlates with the temporal variability of brain networks during the resting state. Cerebr Cortex 29 (3): 1047–1058. https://doi.org/10.3389/fpsyg.2019.00894
Zhu W, Chen Q, Xia L, Beaty RE, Yang W, Tian F, Sun J, Cao G, Zhang Q, Chen X, Qiu J (2017) Common and distinct brain networks underlying verbal and visual creativity. Hum Brain Mapp 38(4): 2094–2111. https://doi.org/10.1002/hbm.23507
Beaty RE, Seli P, Schacter DL (2019) Network neuroscience of creative cognition: Mapping cognitive mechanisms and individual differences in the creative brain. Curr Opin Behav Sci 27: 22–30. https://doi.org/10.1016/j.cobeha.2018.08.013
Gulbinaite R, van Rijn H, Cohen MX (2014) Fronto-parietal network oscillations reveal relationship between working memory capacity and cognitive control. Front Human Neurosci 8: 761. https://doi.org/10.3389/fnhum.2014.00761
Heinonen J, Numminen J, Hlushchuk Y, Antell H, Taatila V, Suomala J (2016) Default Mode and Executive Networks areas: Association with the serial order in divergent thinking. PLoS ONE 11(9): e0162234. https://doi.org/10.1371/journal.pone.0162234
Beaty RE, Kenett YN., Christensen AP, Rosenberg MD, Benedek M, Chen Q, Fink A, Qiu J, Kwapil TR, Kane MJ., Silvia PJ (2018) Robust prediction of individual creative ability from brain functional connectivity. Proc Nat Acad Sci 115(5): 1087–1092. https://doi.org/10.1073/pnas.1713532115
Feng Q, He L, Yang W, Zhang Y, Wu X, Qiu J (2019) Verbal creativity is correlated with the dynamic reconfiguration of brain networks in the resting state. Front Psychol 10: 894. https://doi.org/10.3389/fpsyg.2019.00894
Li W, Yang J, Zhang Q, Li G, Qiu J (2016) The association between resting functional connectivity and visual creativity. Sci Rep 6: 25395. https://doi.org/10.1038/srep25395
Mihov KM, Denzler M, Förster J (2010) Hemispheric specialization and creative thinking: a meta-analytic review of lateralization of creativity. Brain Cogn 72(3): 442–448. https://doi.org/10.1016/j.bandc.2009.12.007
Aziz-Zadeh L, Liew SL, Dandekar F (2013) Exploring the neural correlates of visual creativity. Soc Cognitiv Affective Neurosci 8: 475–480. https://doi.org/10.1093/scan/nss021
Hahm J, Kim KK, Park SH, Lee HM (2017) Brain areas subserving torrance tests of creative thinking: an functional magnetic resonance imaging study. Dementia and Neurocognitiv Disord 16(2): 48–53. https://doi.org/10.12779/dnd.2017.16.2.48
Benedek M, Jung RE, Vartanian O (2018) The neural bases of creativity and intelligence: Common ground and differences. Neuropsychologia 118 (Part A): 1–3. https://doi.org/10.1016/j.neuropsychologia.2018.09.006
Frith E, Elbich DB, Christensen AP, Rosenberg MD, Chen Q, Kane MJ, Silvia PJ, Seli P, Beaty RE (2020) Intelligence and creativity share a common cognitive and neural basis. J Exp Psychol Gen. https://doi.org/10.1037/xge0000958
Jung RE, Haier RJ (2007) The Parieto-Frontal Integration Theory (P-FIT) of intelligence: converging neuroimaging evidence. Behav Brain Sci 30(2): 135–154. https://doi.org/10.1017/S0140525X07001185
Lee KH, Choi YY, Gray JR, Cho SH, Chae JH, Lee S, Kim K (2006) Neural correlates of superior intelligence: Stronger recruitment of posterior parietal cortex. NeuroImage 29: 578–586. https://doi.org/10.1016/j.neuroimage.2005.07.036
Pamplona GSP, Neto GSS, Rosset SRE, Rogers BP, Salmon CEG (2015) Analyzing the association between functional connectivity of the brain and intellectual performance. Front Human Neurosci 9: 1–11. https://doi.org/10.3389/fnhum.2015.00061
Kenett YN, Medaglia JD, Beaty RE, Chen Q, Betzel RF, Thompson-Schill SL, Qiu J (2018) Driving the brain towards creativity and intelligence: A network control theory analysis. Neuropsychologia 118 (PtA): 79–90. https://doi.org/10.1016/j.neuropsychologia.2018.01.001
Разумникова ОМ (2009) Связь частотно-пространственных параметров фоновой ЭЭГ с уровнем интеллекта и креативности. Журн высш нервн деятельн 59(6): 686–695. [Razumnikova OM (2009) Svyaz chastotno-prostranstvennyh parametrov fonovoi EEG s urovnem intellekta b kreativnosti. Zurn Vysshei Nervn Deyatel’osti 59(6): 686–695 (In Russ)].
Herrmann CS, Strüber D, Helfrich RF, Engel AK (2016) EEG oscillations: From correlation to causality. Internat J Psychophysiol 103: 12–21. https://doi.org/10.1016/j.ijpsycho.2015.02.003
Stevens CEJ, Zabelina DL (2019) Creativity comes in waves: An EEG-focused exploration of the creative brain. Curr Opinion Behav Sci 27: 154–162. https://doi.org/10.31234/osf.io/ke6wq
Takeshi O, Aihara T, Shimokawa T, Yamashita O (2018) Large-scale brain network associated with creative insight: combined voxel-based morphometry and resting-state functional connectivity analyses. Scient Rep 8: 6477. https://doi.org/10.1038/s41598-018-24981-0
Benedek M, Jauk E, Sommer M, Arendasy M, Neubauer AC (2014) Intelligence, creativity, and cognitive control: The common and differential involvement of executive functions in intelligence and creativity. Intelligence 46: 73–83. https://doi.org/10.1016/j.intell.2014.05.007
Benedek M, Bergner S, Könen T, Fink A, Neubauer AC (2011) EEG alpha synchronization is related to top-down processing in convergent and divergent thinking. Neuropsychologia 49: 3505–3511. https://doi.org/10.1016/ j.neuropsychologia.2011.09.004
Lustenberger C, Boyle MR, Foulser AA, Mellin JM, Fröhlich F (2015) Functional role of frontal alpha oscillations in creativity. Cortex 67: 74-82. https://doi.org/10.1016/j.cortex.2015.03.012
Erickson B, Truelove-Hill M, Oh Y, Anderson J, Zhang FZ, Kounios J (2018) Resting-state brain oscillations predict trait-like cognitive styles. Neuropsychologia 120: 1–8. https://doi.org/10.1016/j.neuropsychologia.2018.09.014
Kounios J, Fleck JI, Green DL, Payne L, Stevenson JL, Bowden EM, Jung-Beeman M (2008) The origins of insight in resting-state brain activity. Neuropsychologia 46: 281–291.https://doi.org/10.1016/j.neuropsychologia.2007.07.013
Briley PM, Liddle EB, Groom MJ, Smith HJF, Morris PG, Colclough GL, Liddle PF (2018) Development of human electrophysiological brain networks. J Neurophysiol 120(6): 3122–3130. https://doi.org/10.1152/jn.00293.2018
Costa V (2014) The EEG as an index of neuromodulator balance in memory and mental illness. Front Neurosci 8: 63. https://doi.org/10.3389/fnins.2014.00063
Solomon EA, Kragel JE, Sperling MR, Sharan A, Worrell G, Kucewicz M, Inman CS, Lega B, Davis KA, Stein JM, Jobst BC, Zaghloul KA, Sheth SA, Rizzuto DS, Kahana MJ (2017) Widespread theta synchrony and high-frequency desynchronization underlies enhanced cognition. Nat Communicat 8: 1704. https://doi.org/10.1038/s41467-017-01763-2
Hacker CD, Snyder AZ, Pahwa M, Corbetta M, Leuthardt EC (2017) Frequency-specific electrophysiologic correlates of resting state fMRI networks. Neuroimage 149: 446–457. https://doi.org/10.1016/j.neuroimage.2017.01.054
Разумникова ОМ (2009) Особенности селекции информации при креативном мышлении. Психология. Журн Высшей школы экономики 6(3): 134-161. [Razumnikova OM (2009) Osobennosti selekcii infoemacii pri kreativnom myshlenii. Zurn vysshei shkoly ekonomiki 6(3): 134–161 (In Russ)].
Razumnikova OM, Krivonogova KD (2019) Electroencephalographic correlates of the activity of the frontoparietal system as predictors of verbal intelligence and non-verbal creativity. Rus Psychol J 16 (2/1): 45–59. https://doi.org/10.21702/rpj.2019.2.1.4
Разумникова ОМ (2002) Способы определения креативности. Новосибирск: НГТУ. [Razumnikova OM (2002) Sposoby opredeleniya kreativnosti. Novosibirsk: NGTU (In Russ)].
Sunavsky A, Poppenk J (2020) Neuroimaging predictors of creativity in healthy adults. NeuroImage 206: 116292. https://doi.org/10.1016/j.neuroimage.2019.116292
Разумникова ОМ, Каган ВА, Панова НВ (2020) Возрастная динамика показателей вербальной и образной креативности школьников. Комплексные исследования детства 2 (2): 72–79. [Razumnikova OM, Kagan VA, Panova NV (2020) Vozrastnaya dinamika pokazatelei verbalnoi I obraznoi kreativnosti. Kompleksnye issledovaniya detstva 2 (2): 72–79 (In Russ)]. https://doi.org/10.33910/2687-0223-2020- 2-2-72-79
Christensen AP, Benedek M, Silvia P, Beaty R (2019) Executive and default network connectivity reflects conceptual interference during creative imagery generation. PsyArXiv Preprints. https://doi.org/10.31234/osf.io/n438d
Boot N, Baas M, Mühlfeld E, de Dreu CKW, van Gaal S (2017) Widespread neural oscillations in the delta band dissociate rule convergence from rule divergence during creative idea generation. Neuropsychologia 104: 8–17. https://doi.org/10.1016/j.neuropsychologia.2017.07.033
Foster PS, Williamson JB, Harrison DW (2005) The ruff figural fluency test: heightened right frontal lobe delta activity as a function of performance. Arch Clin Neuropsychol 20: 427–434. https://doi.org/10.1016/j.acn.2004.09.010
Dunst B, Benedek M, Jauk E, Bergner S, Koschutnig K, Sommer M, Ischebeck A, Spinath B, Arendasy M, Bühner M, Freudenthaler H, Neubauer AC (2014) Neural efficiency as a function of task demands. Intelligence 42 (100): 22–30. https://doi.org/10.1016/j.intell.2013.09.005
Haier RJ, Siegel BV, Nuechterlein KH, Hazlett E, Wu JC, Paek J (1988) Cortical glucose metabolic rate correlates of abstract reasoning and attention studied with positron emission tomography. Intelligence 12: 199–217.
Knyazev GG (2007) Motivation, emotion, and their inhibitory control mirrored in brain oscillations. Neurosci Biobehav Rev 31 (3): 377–395. https://doi.org/10.1016/j.neubiorev.2006.10.004
Kropotov JD (2009) Frontal midline theta rhythm. In: Quantitative EEG, Event-Related Potentials and Neurotherapy 77–95.
Luo Q, Cheng X, Holroyd T, Xu D, Carver F, Blair J (2014) Theta band activity in response to emotional expressions and its relationship with gamma band activity as revealed by MEG and advanced beamformer source imaging. Front Human Neurosci 7: 940. https://doi.org/10.3389/fnhum.2013.00940
Doppelmayr M, Klimesch W, Hödlmoser K, Sauseng P, Gruber W (2005) Intelligence related upper alpha desynchronization in a semantic memory task. Brain Res Bull 66(2): 171–177. https://doi.org/10.1016/j.brainresbull.2005.04.007
Fellinger R, Grube W, Zaune A, Freunberge R, Klimesch W (2012) Evoked traveling alpha waves predict visual-semantic categorization-speed. NeuroImage 59(4): 3379–3388. https://doi.org/10.1016/j.neuroimage.2011.11.010
Drijvers L, Özyürek A, Jensen O (2018) Alpha and beta oscillations index semantic congruency between speech and gestures in clear and degraded speech. J Cognitive Neurosci 30 (8): 1086–1097. https://doi.org/10.1162/jocn_a_01301
Terporten R, Schoffelen J-M, Dai B, Hagoort P, Kosem A (2019) The relation between alpha/beta oscillations and the encoding of sentence induced contextual information. Sci Rep 9: 20255. https://doi.org/10.1038/s41598-019-56600-x
Fox MD, Snyder AZ, Vincent JL, Corbetta M, Van Essen DC, Raichle ME (2005) The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proc Natl Acad Sci U S A 102(27): 9673–9678. https://doi.org/10.1073/pnas.0504136102
Riemer F, Grüner R, Beresniewicz J, Kazimierczak K, Ersland L, Hugdahl K (2020) Dynamic switching between intrinsic and extrinsic mode networks as demands change from passive to active processing. Sci Rep 10(1): 21463. https://doi.org/10.1038/s41598-020-78579-6
Stevens CEJr, Zabelina DL (2020) Classifying creativity: Applying machine learning techniques to divergent thinking EEG data. Neuroimage 219: 116990. https://doi.org/10.1016/j.neuroimage.2020.116990
Khalil R, Karim AA, Kondinska A, Godde B (2020) Effects of transcranial direct current stimulation of left and right inferior frontal gyrus on creative divergent thinking are moderated by changes in inhibition control. Brain Structure & Function 225(6): 1691–1704. https://doi.org/10.1007/s00429-020-02081-y
Hertenstein E, Waibel E, Frase L, Riemann D, Feige B, Nitsche MA, Kaller CP, Nissen C (2019) Modulation of creativity by transcranial direct current stimulation. Brain Stimul 12(5): 1213–1221. https://doi.org/10.1016/j.brs.2019.06.004
Ivancovsky T, Kurman J, Morio H, Shamay-Tsoory S (2019) Transcranial direct current stimulation (tDCS) targeting the left inferior frontal gyrus: Effects on creativity across cultures. Soc Neurosci 14(3): 277-285. https://doi.org/10.1080/17470919.2018.1464505
Lucchiari C, Sala PM, Vanutelli ME (2018) Promoting creativity through transcranial direct current stimulation (tDCS). A critical review. Front Behav Neurosci 2 (12): 167. https://doi.org/10.3389/fnbeh.2018.00167
Дополнительные материалы отсутствуют.
Инструменты
Российский физиологический журнал им. И.М. Сеченова