

Российский физиологический журнал им. И.М. Сеченова, 2023, T. 109, № 2, стр. 241-251
Межполушарная симметрия и асимметрия пик-волновых разрядов абсансного типа, вызванных системным введением пентилентетразола
А. С. Ершова 1, Е. М. Сулейманова 2, А. А. Грищенко 1, *, Л. В. Виноградова 2, И. В. Сысоев 1, 2
1 Саратовский государственный университет им. Н.Г. Чернышевского
Саратов, Россия
2 Институт высшей нервной деятельности и нейрофизиологии РАН
Москва, Россия
* E-mail: vili_von@mail.ru
Поступила в редакцию 21.11.2022
После доработки 27.12.2022
Принята к публикации 28.12.2022
- EDN: NDLJNT
- DOI: 10.31857/S0869813923020048
Аннотация
Абсансная эпилепсия – специфическая генерализованная неконвульсивная форма эпилепсии, чаще всего встречающаяся у детей и подростков. Традиционно считается, что возникающие пик-волновые разряды являются полностью генерализованными в коре обоих полушарий, что наблюдается у людей на поверхностных электроэнцефалограммах и магнитоэнцефалограммах. Однако основное изучение механизмов этой формы эпилепсии проводится на животных, главным образом на крысах, – генетических и фармакологических моделях, поскольку необходимо измерение сигналов не только из коры, но и подкорковых структур мозга (например, таламуса, участвующего в генерации пик-волновых разрядов), что невозможно сделать на людях, у которых нет медицинских показаний для внутричерепных измерений. У животных с абсансной эпилепсией локальные потенциалы почти всегда регистрируются из одного полушария, чтобы охватить максимальное число задействованных структур мозга. При этом степень одновременности появления и прекращения абсансных пик-волновых разрядов в разных полушариях у животных практически не исследована, а сама разметка в подавляющем большинстве случаев делается на основе одного канала моторной или соматосенсорной коры. Цель данной работы – выявить различия и сходство статистических характеристик пик-волновых разрядов в двух полушариях коры на модели фармакологически вызванных судорог абсансного типа. Пик-волновые разряды являются основным энцефалографическим маркером абсансов у крыс, подвергнутых воздействию пентилентетразола. Для детектирования разрядов в работе предложен метод их автоматической разметки, оценены его чувствительность и специфичность на примере записей девяти животных. С помощью данного метода произведена разметка приступов в симметричных отведениях коры из обоих полушарий по отдельности. По результатам анализа длительности приступов оказалось, что для пяти из девяти животных распределение разрядов значимо различается между полушариями, а для других четырех – нет. Следовательно, у крыс-моделей пик-волновая активность может быть генерализована как симметрично, так и несимметрично, что может быть обусловлено индивидуальными особенностями или различными сценариями запуска разрядов.
ВВЕДЕНИЕ
Эпилепсия – хроническое заболевание головного мозга, которое часто сопутствует или провоцирует другие расстройства высшей нервной деятельности, в том числе волну распространяющейся депрессии. Абсансная эпилепсия чаще всего встречается среди детей и подростков, характеризуется потерей сознания на короткое время, иногда она имеет постепенное начало и окончание. У больных абсансной эпилепсией нет показаний для инвазивных исследований и хирургического вмешательства, что затрудняет получение информации о механизмах этого заболевания. Поэтому традиционно экспериментальную работу проводят на животных, у которых возможно производить хирургические вмешательства и записывать сигналы глубоких структур мозга. Самыми популярными в настоящее время являются крысы с генетической предрасположенностью к заболеванию линий WAG/Rij [1] и GAERS [2]. Для этих животных, начиная с некоторого возраста, характерны спонтанные пик-волновые разряды, средняя частота которых составляет 8–9 Гц у крыс WAG/Rij и 5–6 Гц у крыс GAERS [3], а медианная длительность разрядов – около 6 с [4].
Кроме того, исследования проводятся и на фармакологических моделях, которые хороши тем, что позволяют моделировать вызванные судороги абсансного типа у исходно здоровых животных путем однократной интенсивной стимуляции. Наиболее популярными моделями этого типа являются животные, пик-волновые разряды у которых вызваны введением низких доз пентилентетразола (ПТЗ), см. обзор [5]: крысы [6], мыши [7] и морские свинки [8]. При этом более высокие дозы того же вещества вызывают тонико-клонические судороги [9, 10]. Для нас эти модели имеют большое значение в том числе потому, что различные дозы и протоколы введения ПТЗ могут приводить как к эпилептическим приступам, так и к появлению волны распространяющейся депрессии [11–13].
Традиционно принято считать, что у крыс с индуцированными ПТЗ абсансами [12], как и у животных с генетической абсансной эпилепсией [1], а также и у пациентов [14], разряды в коре правого и левого полушарий головного мозга одинаковы. Поэтому, чтобы охватить как можно больше структур мозга, в той или иной степени вовлеченных в пик-волновую активность, все электроды ставят в одно полушарие. Тем не менее, вопрос о том, являются ли такие разряды действительно всегда первично генерализованными в обоих полушариях, систематически не исследовался. Известно, что при других формах эпилепсии, изначально фокальных, волна распространяющейся депрессии возникает только в одном полушарии [15], между тем ПТЗ в больших дозах также вызывает и тонико-клонические приступы, свойственные лимбической и аудиогенной вторично генерализованной эпилепсии, и волну распространяющейся депрессии. Поэтому потенциально можно ожидать, что асимметрия в распространении абсансов, вызванных системным введением ПТЗ, возможна.
Цель данной работы – по временным рядам локальных потенциалов коры двух полушарий проверить симметричность абсансов, вызванных системным введением ПТЗ.
МЕТОДЫ ИССЛЕДОВАНИЯ
Эксперименты проводили на самцах крыс линии Wistar, в возрасте 6–7 мес., полученных из питомника “Столбовая” (Московская область). Электрическая активность неокортекса регистрировалась с помощью электродов (стальных винтов), имплантированных в симметричные области фронтальной коры обоих полушарий по координатам: AP 2; ML ± 2; DV 1 [16]. В качестве индифферентного электрода использовали стальной винт, расположенный над мозжечком. Электроды вживлялись под наркозом (хлоралгидрат, 380 мг/кг) за две недели до начала экспериментов. Электрическую активность коры регистрировали с помощью 4-канального усилителя и АЦП (E14-440, L-Card, Россия) у бодрствующих свободно подвижных животных. Для индукции пик-волновых разрядов использовалось внутрибрюшинное введение конвульсанта ПТЗ в дозе 40 мг/кг. По окончании эксперимента проводили гистологический анализ для определения локализации регистрирующих электродов. В данной работе рассматривались записи 9 животных, длительность всех записей составляла не менее 1 ч, в среднем около 90 мин. Типичный разряд представлен на рис. 1. Из спектрограмм (см. рис. 1b, d) видно, что колебания во время разряда очень нелинейные: можно надежно различить пять гармоник основной частоты.
Рис. 1.
Временные ряды (а, c) и спектрограммы (b, d) пик-волнового разряда, записанного в лобной коре правого (a, b) и левого (с, d) полушарий крысы № 8. Оси X – время в с (t, s), оси Y – напряжение (U, mV) на временных рядах или частота в герцах (f, Hz) на спектрограммах. На всех графиках черные вертикальные линии означают начало и конец разряда.

Непосредственно регистрируемые (необработанные) ряды не подходят для анализа из-за наличия трендов и помех. Поэтому, во-первых, с помощью извлечения скользящего среднего, рассчитанного на временном интервале 2 с, удалялся медленный тренд, обусловленный особенностями процедуры измерения. Затем сигнал дополнительно фильтровался: режекторным фильтром в полосе 49–51 Гц удалялась сетевая 50 Гц наводка, фильтром высоких частот – все частотные компоненты на частотах более 99 Гц: эти компоненты не принимают существенного участия в пик-волновых разрядах. При этом в сигнале сохранялись высокоамплитудные артефакты (движения, компенсации и т.д.), поскольку записи были достаточно длинными и проводились у свободно подвижных животных.
Чтобы быстро и непредвзято проверить гипотезу о симметричном распространении патологической активности, необходимо выделить разряды из фоновой активности и определить длительность разрядов с помощью автоматического метода. Мы использовали очень простой подход, частично основанный на ранее предложенных подходах [17, 18], но без использования спектральных свойств сигнала, так как у рассматриваемой фармакологической модели основная частота не так стабильна, как у крыс WAG/Rij, у которых основной временной масштаб сохраняется не только в течение всего разряда, но и возникает до него [19]. Поскольку мы искали пик-волновые комплексы, предложенный подход основан на обнаружении пиков и состоит в следующем.
Пусть имеется временной ряд значений {xi} – скалярная запись локальных потенциалов поля, где i – номер момента времени. Среднее mi рассчитывается для значений в ряде, предшествующих xi + 1 и расположенных после последнего обнаруженного разряда (либо с начала записи). Затем каждое (xi + 1) сравнивается с (10|mi|), где константа 10 подобрана эмпирически и является параметром метода. Если |xi + 1| > 10|mi|, считается, что xi + 1 является началом пик-волнового комплекса. Если два пика находятся рядом и их разделяет не более τ значений, где τ определяется на основании спектральных свойств сигнала: основной период пик-волновой активности для данной модели абсансов составляет 6 Гц, то все значения между ними считаются частью пик-волнового разряда. Блок-схема алгоритма разряда изображена на рис. 2.

Для проверки точности метода были рассчитаны специфичность и чувствительность. Чувствительность означает способность метода обнаруживать разряды, которые действительно присутствовали, и рассчитывается как Ntf/Nreal, где Ntf – количество реальных пиков-волновых комплексов, найденных алгоритмом, а Nreal – полное количество истинных пиков-волновых комплексов по разметке энцефалографиста. Специфичность – это способность метода избегать ложноположительных результатов. Хорошая специфичность означает относительно небольшое количество ложно выявленных разрядов. Специфичность рассчитывается как 1 – Nff/Nreal, где Nff – число ложноположительных обнаружений. Результаты оценки специфичности приведены в табл. 1 и чувствительности в табл. 2.
Видно, что результаты отличаются для разных животных. В основном, при лучшей чувствительности мы получаем худшую специфичность и наоборот, что очень естественно и типично [18]. Однако среднего значения чувствительности и специфичности вполне достаточно, чтобы использовать метод для автоматического определения, если мы хотим получить общее впечатление и сравнить результаты по левому и правому полушариям.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Поскольку метод показал достаточную чувствительность и специфичность, для анализа использовалась длительность разрядов, полученная с помощью предложенного алгоритма автоматического детектирования. По результатам обнаружения были построены гистограммы длительности разрядов для каждого животного отдельно. Для примера, гистограммы для животных № 8 и № 9 показаны на рис. 3.
Рис. 3.
Гистограммы распределения длительности разрядов для крыс № 8 и № 9. Желтый цвет – оценка распределения для разрядов в правом полушарии, серо-голубой – в левом. По оси X отложена длина разрядов, по оси Y – их количество.

Результаты анализа оказались неоднозначными. Для крыс № 2, 4, 6 и 8 распределения длительности разрядов были очень похожи в обоих полушариях. Для остальных пяти крыс распределения в разных полушариях заметно отличались. Чтобы понять детали, дополнительно подробнее были проанализированы по 30 разрядов у крыс № 8 и № 9, то есть у одного животного, у которого заметных отличий в длине разрядов нет, и у одного, у которого длины разрядов в разных полушариях существенно различаются. Есть три типичные ситуации. Во-первых, в некоторых случаях разряды начинаются одновременно и имеют одинаковую длительность в обоих каналах, как видно из рис. 4. Для крысы № 8 все рассмотренные разряды относятся к этому типу.
Рис. 4.
Фрагмент записи электрической активности коры правого (a) и левого (b) полушарий крысы № 9. По оси X – время в с (t, s), по оси Y – напряжение (U, mV). Черным выделены участки, определенные алгоритмом как разряд.
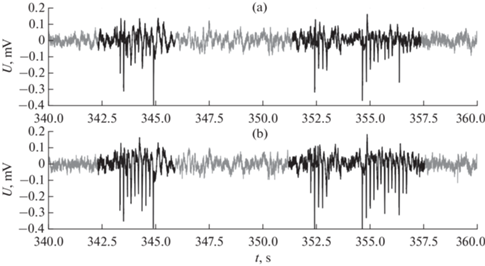
Второй случай, когда в одном отведении разряды существенно менее выражены, чем в другом (в том числе по амплитуде, а также по форме и по наличию высших гармоник в спектре), поэтому они либо не обнаруживаются из-за недостаточно резких пиков, либо обнаруживаются с уменьшенной длиной (см. в качестве типичного примера рис. 5).
Рис. 5.
Фрагмент записи электрической активности коры правого (a) и левого (b) полушарий крысы № 9. По оси X – время в с (t, s), по оси Y – напряжение (U, mV). Черным выделены участки, выделенные алгоритмом, серым – основной сигнал для сравнения.

В некоторых случаях, как это показано на рис. 6, разряд явно развивается только в одном полушарии, в то время как в другом нет эпилептиформной активности, отличной от фона, поэтому нельзя сказать, что алгоритм обнаружения недостаточно чувствителен или что разряд асимметричен и лучше выражен в одном полушарии. Поскольку все три рассмотренных на рис. 4–6 случая были записаны у одного и того же животного в течение одного часа записи в произвольном порядке, гипотезы о том, что эти различия обусловлены положением электродов или какими-то эффектами во время записи, должны быть отвергнуты.
Рис. 6.
Фрагмент записи электрической активности коры правого (a) и левого (b) полушарий крысы № 9. По оси X – время в с (t, s), по оси Y – напряжение (U, mV). Черным выделены участки, выделенные алгоритмом, серым – основной сигнал для сравнения.

Для количественного описания различий в продолжительности разрядов мы использовали два хорошо известных статистических теста: тест Колмогорова–Смирнова и тест Манна–Уитни. Программная реализация обоих тестов была взята из популярного пакета scipy [20]. Оба подхода проверяют гипотезу о том, что две выборки (в нашем случае это выборки длительности приступов в правом и левом полушариях) взяты из одного и того же распределения. Оба метода выдают значение p – вероятность ошибиться, опровергая гипотезу о том, что распределения одинаковы. Как правило, при малых значениях p делается вывод о значимых различиях, в то время как при относительно больших p, например, p > 0.05, исходная гипотеза принимается и выборки считаются принадлежащими одному и тому же распределению.
Для более удобного анализа данных значения p двух тестов были сведены в табл. 3. Для различных очень малых p < 10–5 в таблицу записывалось значение 10–5, поскольку различия между меньшими значениями на наших размерах выборки не имеют статистического смысла. Оба теста показали очень похожие результаты, четко различая две группы животных. Четыре крысы: № 2, 4, 6 и 8 имели симметричные разряды, и для них значения p большие – p > 0.2. Для пяти других животных: № 1, 3, 5, 7, 9 значения p < 10–5 , то есть распределения достоверно различные (даже если мы допустим поправку на множественное тестирование, сделав оценку сверху умножением полученных значений на число животных, все равно оба метода дадут p < 10–4). Как видно из рис. 4, это не означает, что все разряды у этих животных разные, но некоторые или многие из них либо разной длины (например, см. рис. 5), либо развиваются только в одном полушарии (например, см. рис. 6).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В данной работе анализировалась длительность пик-волновых разрядов у крыс с вызванными системным введением ПТЗ абсансами. Анализ производился по отдельности для правого и левого полушарий с целью выявить, в какой степени пик-волновые разряды симметричны. Для непредвзятого определения длины пик-волновых разрядов был предложен и реализован простой алгоритм обнаружения пик-волновых комплексов, для которого были определены специфичность и чувствительность, оказавшиеся достаточно высокими для решения поставленной задачи.
Было показано, что распределение длительности пик-волновых разрядов в разных полушариях отличается у пяти из девяти проанализированных животных и одинаково у остальных четырех. У животных с разным распределением наблюдаются как симметричные билатеральные, так и асимметричные (в одном полушарии короче, чем в другом) разряды и разряды, локализованные только в одном полушарии. Поскольку все животные принадлежат к одной и той же линии Wistar, сходны по массе тела, одного пола и возраста, оперировались одинаково, это означает, что существует индивидуальная предрасположенность к генерации двусторонних или односторонних разрядов. При этом сосуществование как симметричных, так и односторонних разрядов у одних и тех же животных может указывать на наличие как минимум двух различных механизмов запуска эпилептиформной активности абсансного типа, в то время как у четырех животных с симметричными разрядами один из этих механизмов не реализуется.
Выявленные особенности динамики эпилептиформной активности характеризуют именно рассмотренную фармакологическую модель абсансных судорог. По результатам данного исследования нельзя сказать, имеют ли пик-волновые разряды у генетических моделей (крысы WAG/Rij и GAERS) такие же индивидуальные свойства. Но если это так, то это может объяснить, почему эволюция связности во время абсансных приступов сильно различается в популяции, как это было показано в ранее опубликованных работах [21, 22] с использованием различных мер связанности. Кроме того, при рассмотрении распространяющейся депрессии, инициированной инъекцией ПТЗ, односторонний или двусторонний тип волны депрессии может основываться на одном и том же механизме индивидуальной предрасположенности, что и при возникновении симметричных или асимметричных пик-волновых разрядов. Эти вопросы, несомненно, требуют дальнейшего изучения.
Список литературы
Coenen A, van Luijtelaar G (2003) Genetic animal models for absence epilepsy: a review of the WAG/Rij strain of rats. Behav Genet 33(6): 635–655. https://doi.org/10.1023/a:1026179013847
Marescaux C, Vergnes M, Depaulis A (1992) Genetic absence epilepsy in rats from Strasbourg - a review. J Neural Transmis Suppl 35: 37–69. https://doi.org/10.1007/978-3-7091-9206-1_4
Akman O, Demiralp T, Ates N, Onat FY (2010) Electroencephalographic differences between WAG/Rij and GAERS rat models of absence epilepsy. Epilepsy Res 89(2): 185–193. https://doi.org/10.1016/j.eplepsyres.2009.12.005
van Rijn CM, Gaetani S, Santolini I, Badura A, Gabova A, Fu J, Watanabe M, Cuomo V, van Luijtelaar G, Nicoletti F, Ngomba RT (2010) WAG/Rij rats show a reduced expression of CB1 receptors in thalamic nuclei and respond to the CB1 receptor agonist, R(+)WIN55,212-2, with a reduced incidence of spike-wave discharges. Epilepsia 51(8): 1511–1521. https://doi.org/10.1111/j.1528-1167.2009.02510.x
Jafarian M, Esmaeil AM, Karimzadeh F (2020) Experimental Models of Absence Epilepsy. Basic Clin Neurosci 11(6): 715–726. https://doi.org/10.32598/bcn.11.6.731.1
Marescaux C, Micheletti G, Vergnes M, Depaulis A, Rumbach L, Warter JM (1984) A Model of Chronic Spontaneous Petit Mal-like Seizures in the Rat: Comparison with Pentylenetetrazol-Induced Seizures. Epilepsia 25(3): 326–331. https://doi.org/10.1111/j.1528-1157.1984.tb04196.x
Medina AE, Manhaes AC, Schmidt SL (2001) Sex differences in sensitivity to seizures elicited by pentylenetetrazol in mice. Pharmacol Biochem Behav 68(3): 591–596. https://doi.org/10.1016/s0091-3057(01)00466-x
Solmaz I, Gürkanlar D, Gökcil Z, Göksoy C, Özkan M, Erdogan E (2009) Antiepileptic activity of melatonin in guinea pigs with pentylenetetrazol-induced seizures. Neurol Res 31(9): 989–995. https://doi.org/10.1179/174313209X385545
Klioueva IA, van Luijtelaar ELJM, Chepurnova NE, Chepurnov SA (2001) PTZ-induced seizures in rats: effects of age and strain. Physiol Behav 72(3): 421–426. https://doi.org/10.1016/s0031-9384(00)00425-x
Shandra AA, Godlevsky LS (2005) Pentylenetetrazol-Induced Kindling as a Model of Absence and Convulsive forms of Epilepsy. In: Corcoran ME, Mosh’e SL (eds) Kindling 6. Advances in Behavioral Biology, 55. Springer. Boston. MA. https://doi.org/10.1007/0-387-26144-3_6
Vergnes M, Marescaux C (1992) Cortical and thalamic lesions in rats with genetic absence epilepsy. In: Marescaux C, Vergnes M, Bernasconi R (eds) Generalized Non-Convulsive Epilepsy: Focus on GABA-B Receptors. J Neur Transmis 35. Springer. Vienna. https://doi.org/10.1007/978-3-7091-9206-1_5
Vinogradova LV, Koroleva VI (1993) Cortico-caudate spreading depression and seizure activity induced by systemic daily pentylenetetrazole injection in rats. Pavlov J High Nerv Act 43(4): 683–694.
Koroleva VI, Vinogradova LV, Bures J (1993) Reduced incidence of cortical spreading depression in the course of pentylenetetrazol kindling in rats. Brain Res 608(1): 107–114. https://doi.org/10.1016/0006-8993(93)90780-q
Crunelli V, Leresche N (2002) Childhood absence epilepsy: Genes, channels, neurons and networks. Nat Rev Neurosci 3: 371–382. https://doi.org/10.1038/nrn811
Vinogradova LV, Vinogradov VY, Kuznetsova GD (2006) Unilateral cortical spreading depression is an early marker of audiogenic kindling in awake rats. Epilepsy Res 71(1): 64–75. https://doi.org/10.1016/j.eplepsyres.2006.05.014
Paxinos G, Watson C (2005) The rat brain in stereotaxic coordinates. Elsevier. Amsterdam, Boston.
Ovchinnikov A, Luttjohann A, Hramov A, van Luijtelaar G (2010) An algorithm for real-time detection of spike-wave discharges in rodents. J Neurosci Methods 194(1): 172–178. https://doi.org/10.1016/j.jneumeth.2010.09.017
van Luijtelaar G, Luttjohann A, Makarov VV, Maksimenko VA, Koronovskii A, Hramov AE (2016) Methods of automated absence seizure detection, interference by stimulation, and possibilities for prediction in genetic absence models. J Neurosci Methods 260: 144–158. https://doi.org/10.1016/j.jneumeth.2015.07.010
Grishchenko AA, Sysoeva MV, Sysoev IV (2020) Detecting the primary time scale of evolution of information properties for local field potentials in brain at absence epilepsy. Izvestiya VUZ. A-ppl Nonlin Dynam 28(1): 98–110. https://doi.org/10.18500/0869-6632-2020-28-1-98-110
Virtanen P, Gommers R, Oliphant TE, Haberland M, Reddy T, Cournapeau D, Burovski E, Peterson P, Weckesser W, Bright J, van der Walt SJ, Brett M, Wilson J, Jarrod Millman K, Mayorov N, Nelson ARJ, Jones E, Kern R, Larson E, Carey C, Polat I, Feng Y, Moore EW, Vand erPlas J, Laxalde D, Perktold J, Cimrman R, Henriksen I, Quintero EA, Harris CR, Archibald M, Ribeiro AH, Pedregosa F, van Mulbregt P, Contributors S (2020) SciPy 1.0: Fundamental Algorithms for Scientific Computing in Python. Nat Methods 17: 261–272. https://doi.org/10.1038/s41592-019-0686-2
Grishchenko AA, van Rijn CM, Sysoev IV (2017) Comparative Analysis of Methods for Estimation of Undirected Coupling from Time Series of Intracranial EEGs of Cortex of Rats– Genetic Models of Absence Epilepsy. Math Biol Bioinf 12(2): 317–326. https://doi.org/10.17537/2017.12.317
Grishchenko AA, Sysoeva MV, Medvedeva TM, van Rijn CM, Bezruchko BP, Sysoev IV (2020) Comparison of approaches to directed connectivity detection in application to spike-wave discharge study. Cybernet and Physics 9(2): 86–97. https://doi.org/10.35470/2226-4116-2020-9-2-86-97
Дополнительные материалы отсутствуют.
Инструменты
Российский физиологический журнал им. И.М. Сеченова