

Сенсорные системы, 2019, T. 33, № 1, стр. 3-6
Анализ предпочтений полового партнера у Eurydema ornata и E. oleracea в условиях свободного выбора
1 Федеральное государственное бюджетное учреждение науки
Институт проблем передачи информации им. А.А. Харкевича РАН
127051 Москва, Большой Каретный пер., 19, Россия
2 Федеральное государственное автономное образовательное учреждение высшего образования
“Российский Университет Дружбы Народов”, Агробиотехнологический департамент
117198 Москва, ул. Миклухо-Маклая д. 8 к. 2, Россия
* E-mail: zicrona@iitp.ru
Поступила в редакцию 18.09.2018
Аннотация
Протестированы предпочтения самок E. оrnata и E. оleracea в условиях свободного выбора. Гетероспецифическое ухаживание у Eurydema прерывается на более ранних стадиях, чем у Nezara. Одним из механизмов предотвращения межвидовой копуляции могут быть сильные различия в структуре сигналов ухаживания этих видов.
ВВЕДЕНИЕ
Несмотря на большой интерес к изучению коммуникации полужесткокрылых (Cokl, 2008; de Groot et al., 2010; Laumann et al., 2013; Cocroft et al., 2010; McBrien et al., 2003; Panizzi et al., 2000), предпочтения самок и роль вибрационных сигналов в межвидовой изоляции изучалась на небольшом количестве видов (Kon et al., 1994). При этом на внутривидовом уровне на Nezara и нескольких других модельных видах механизмы поиска в распознавании полового партнера изучаются до настоящего времени. Так, на Nezara viridula показано, что изменение отдельных временных элементов сигнала в модельных стимулах (длительности серии пульсов, периода повторения) может существенно влиять на успешность поиска и копуляции (Cokl, 2008; Polajnar, Cokl, 2008; Miklas et al., 2003), однако не предотвращает межвидовые копуляции полностью. В ситуации отсутствия выбора самки Nezara могут копулировать с гетероспецифическими самцами.
У других симпатрических видов, например, у представителей рода Chivania (Blassioli-Moraes et al., 2012; Laumann et al., 2016) гетероспецифических копуляций не происходит даже в эксперименте с отсутствием выбора. Авторы полагают, что ухаживание у этих видов останавливается на более ранней стадии, нежели у видов Nezara, поэтому у них отсутствуют межвидовые скрещивания. Данный вывод кажется логичным, если учесть совместное обитание этих видов на большей части ареала в отличие от видов рода Nezara. Однако это может свидетельствовать о недавнем в эволюционном плане расхождении видов Chinavia. Считается, что недавно разошедшиеся виды используют в основном предкопуляционные барьеры изоляции, так как посткопуляционные могут работать плохо или не работать вовсе Несомненно, акустические и вибрационные сигналы играют важную роль в поиске и распознавании полового партнера у полужесткокрылых (Blassioli-Moraes, 2014). В некоторых работах показано, что сигналы могут кодировать не только видоспецифические, но и индивидуальные параметры особи (Rantala, Kortet, 2003).
Представители рода Eurydema широко распространены в пределах Палеарктики и во многих стациях являются синтопическими, т.е. могут обитать совместно даже на одном кормовом растении. При этом в репертуаре отмечен всего один полифункциональный сигнал, который используется при взаимодействии как самцов, так и самок. Сигналы разных видов сходны по частотным характеристикам, но отличаются по амплитудно-временным параметрам (Shestakov, 2015).
В нашем исследовании мы видеотипировали взаимодействие самцов и самок и измерили 12 различных поведенческих элементов, продемонстрированных обоими полами. Изучили различия в поведении при взаимодействии с самками гетероспецифических и конспецифических самцов. Учитывая, что сигнал ухаживания является мультимодальным, мы попытались оценить, какие сенсорные модальности (химические или акустические) могли бы быть индикатором конспецифичности самца, а какие зависят от индивидуальных характеристик.
МАТЕРИАЛ И МЕТОДИКА
Исходный живой материал для работы собран в полевых условиях. Регионы сбора: Московская, Саратовская области, Краснодарский край. Перевозку насекомых осуществляли в пластиковых контейнерах 10 × 10 × 5 см со слоем фильтровальной бумаги на дне (для контроля влажности и предотвращения поражения плесневыми грибами). В лаборатории насекомых содержали в садках 35 × 35 × 45 см при постоянном наличии свежего кормового растения и 12/12 световом цикле. Температуру поддерживали на уровне 20–25°С. В экспериментах использовались только особи, собранные на предимагинальных стадиях. Самцы и самки до имагинальной линьки содержались раздельно. Такое разделение необходимо, так как опыт предыдущей копуляции может оказывать влияние на поведенческие реакции, поэтому в экспериментах должны использоваться только заведомо девственные самцы и самки.
Для регистрации сигналов в полевых условиях применяли пьезоэлектрический адаптер ГЗК-661. Сигнал с адаптера подавали на микрофонный вход звуковой платы компьютера (ноутбука) или на минидиск-рекордер Sony Hi-MD Walkman MZ-RH910 через согласующий усилитель, сходный по схеме с предварительными усилителями, предназначенными для использования в электрофонах с пьезоэлектрическими звукоснимателями. Для регистрации (проверки наличия) акустической компоненты сигнала запись параллельно велась на микрофон (4191, 1/2 inch; Bruel & € Kjær, Nærum, Denmark), расположенный на расстоянии 1–3 см от насекомых. Частота дискретизации сигналов 44.1 кГц. Измерение амплитудно-временных параметров сигналов проводили при помощи программ CoolEdit (США, Syntrillium) и Turbolab 4.0 (Германия, Bressner Technology). В каждом сигнале использовано не менее 50 измерений для каждого параметра. Статистический анализ сигналов проводили в программах Excel и Statistica.
Мы протестировали предпочтения самок E. оrnata (N = 34) и E. оleracea (N = 37) в условиях свободного выбора. Каждая самка помещалась на растение с одним самцом своего и одним самцом родственного вида. Было проанализировано 12 параметров поведения ухаживания. Учитывались как ответные реакции самки во время ухаживания, так и процент успешных копуляций. Насекомые находились совместно 20–30 мин, при отсутствии ответной реакции пара заменялась на следующую. Для стимуляции активности использована хорошо зарекомендовавшая себя ранее искусственная зимовка. Все особи были маркированы цветными метками на щитке, для идентификации. Каждый час проверяли наличие копулирующих пар. Одновременно вели видеотипирование (Sony DCR-TRV 355E), видеозаписи были проанализированы при помощи программы “Boris” (Friard, Gamba, 2016).
РЕЗУЛЬТАТЫ
Элементы ухаживания. В более чем 60% ссаживаний мы выделили 12 поведенческих элементов процесса ухаживания. Существенных различий между кон- и гетероспецифическими самцами в количестве разных поведенческих элементов мы не обнаружили (Fisher’s Exact Test, two-tailed; p > 0.05). Однако процент экспериментов, в которых были отмечены разные элементы поведения при ухаживании за кон- и гетероспецифическими самками, различались (рис. 1 и 2).
Рис. 1.
Процент экспериментов, в которых был данный поведенческий элемент при ссаживании с самками E. oleracea. Звездочки обозначают статистическую значимость различий между самцами разных видов (Fisher’s Exact Test, two-tailed; *р < 0.05).
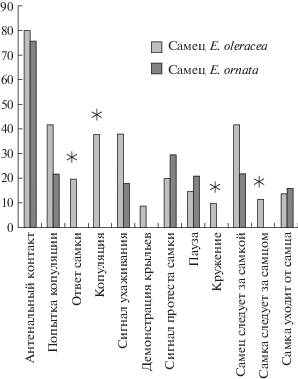
Рис. 2.
Процент экспериментов, в которых был данный поведенческий элемент при ссаживании с самками E. ornata. Звездочки обозначают статистическую значимость различий между самцами разных видов (Fisher’s Exact Test, two-tailed; *р < 0.05).
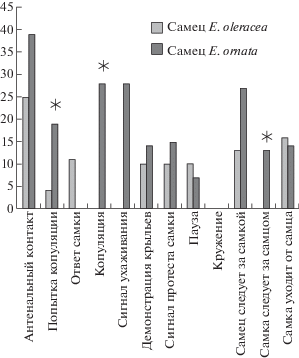
Вибрационные сигналы во время ухаживания. Большинство самцов издавали вибрационные сигналы во время ухаживания за конспецифическими самками. При гетероспецифическом ухаживании сигналы издавали только самцы E. ornata. При этом эмиссия сигнала отмечена у них достоверно реже, чем у самцов E. oleracea (Fisher’s Exact Test, two-tailed; p < 0.05) (рис. 2). Ответы самок зарегистрированы только на сигналы конспецифических самцов. На сигналы самцов другого вида самки E. ornata и E. oleracea не отвечали вовсе. Сигнал протеста издавался самками лишь в нескольких ссаживаниях и в равной степени при контактах с самцами своего и чужого вида. Во всех случаях самки копулировали только с конспецифическими самцами (рис. 1 и 2).
ОБСУЖДЕНИЕ
Наши результаты показали, что самки Euridema, в отличие от Nezara, отвечают на ухаживания и спариваются только с конспецифическими самцами. На наш взгляд, это может говорить о том, что гетероспецифическое ухаживание у них останавливается на более ранней стадии. Одним из важных механизмов, обеспечивающих прекопуляционную изоляцию, могут быть различия в амплитудно-временной структуре сигналов ухаживания. Вибрационные сигналы являются необходимым поведенческим элементом для успешной копуляции у многих видов полужесткокрылых (Cokl et al., 2014). У изученных нами видов лишь в единичных случаях копуляция может происходить без сигнала.
Сигнал ухаживания полужесткокрылых является мультимодальным и помимо акустических включает феромонные, визуальные и тактильные стимулы. Однако феромоны не обеспечивают межвидовую изоляцию, а используются для привлечения самок на растение. После чего для поиска партнера непосредственно на растении применяются вибрационные сигналы, а при непосредственном контакте добавляются визуальные (демонстрация яркой нижней поверхности крыльев) и тактильные, такие как ощупывание. В наших экспериментах успешно завершились только акты ухаживания, сопровождаемые вибрационными сигналами.
Мы отметили, что некоторые самцы E. ornata пытались копулировать за самками E. oleracea, но самцы E. oleracea не ухаживали за гетероспецифическими самками. Данные различия могут быть связаны с относительными размерами особей – относительный размер E. ornatа больше, чем E. oleracea. Кроме того, сигналы самок имеют сходную структуру по амплитудным и частотным характеристикам, а сигналы самцов сильно различаются, но самки E. ornatа издают сигналы реже, чем самки E. oleracea. Количество попыток спаривания с самкой чужого вида соответствует числу ответов самки, т.е. попытки копуляции с гетероспецифической самкой можно объяснить реакцией самца на неспецифический сигнал самки.
В дальнейшем мы планируем изучить роль разных элементов сигналов в распознавании полового партнера в экспериментах без выбора. Самкам будут предъявляться немые самцы и проигрываться стимулы с измененными характеристиками.
Список литературы отсутствует.
Дополнительные материалы отсутствуют.
Инструменты
Сенсорные системы