

Сенсорные системы, 2019, T. 33, № 2, стр. 91-98
Расчет оптической системы глаза китообразных
А. М. Масс 1, *, А. Я. Супин 1, **
1 Федеральное государственное бюджетное учреждение науки
Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский просп., 33, Россия
* E-mail: alla-mass@mail.ru
** E-mail: alex_supin@mail.ru
Поступила в редакцию 04.12.2018
Аннотация
На основании данных о размерах светопреломляющих структур глаза рассчитано положение сфокусированного изображения у нескольких видов китообразных в воздушной и водной средах. В воздухе для световых лучей, проходящих вдоль оси глаза, глаз китообразных миопичен (сфокусированное изображение создается перед сетчаткой), в воде – эмметропичен (сфокусированное изображение создается на сетчатке). Для световых лучей, проходящих через периферийные части роговицы и зрачка, глаз эмметропичен в воде и воздухе.
ВВЕДЕНИЕ
Представители нескольких отрядов класса млекопитающих характеризуются вторичной адаптацией к водной среде обитания. Степень такой адаптации может быть различной. У некоторых насекомоядных (выхухоль Desmana moschata, водяная кутора Neomys fodiens)), грызунов (бобры Castor sp., нутрия Myocastor coypus, капибара Hydrochoerus hydrochaeris, австралийская водяная крыса Hydromys chrysogaster, ондатра Ondatra zibethicus), хищных (выдры Lutra sp., Enhydra lutris) и парнопалых (гиппопотамы Hippopotamus amphibius, Hexaprotodon liberiensis) образ жизни в значительной степени привязан к воде, однако, адаптация к водному образу жизни минимальна и не затронула многие анатомо-физиологические особенности, характерные для наземных млекопитающих. Более глубокая адаптация к водному образу жизни наблюдается у ластоногих – настоящих (Phocidae) и ушатых (Otariidae) тюленей и моржей (Odobenidae). Значительная часть их жизненного цикла проходит в водной среде. Хотя таксономически все они относятся к отряду хищных, их анатомия и физиология существенно перестроены по сравнению с наземными хищными животными.
Только для двух отрядов млекопитающих характерен полностью водный образ жизни, при котором они никогда не покидают водную среду. Это китообразные – киты, дельфины, морские свиньи (Cetacea) и сиреновые (Sirenia). Полностью водный образ жизни у этих животных связан с глубокой анатомо-физиологической перестройкой всех систем организма, в том числе перестройка касается зрительной системы. Изменения, которые претерпела зрительная система водных млекопитающих по сравнению с наземными, в значительной степени обусловлены различными физическими характеристиками воды и воздуха. Самое очевидное из различий касается коэффициентов светопреломления этих сред, что требует соответствующей организации оптической системы глаза. Из других различий можно отметить разную прозрачность воды и воздуха. Прозрачность даже очень чистой воды обеспечивает видимость на расстоянии нескольких десятков метров (в мутной воде видимость может не превышать сантиметры), тогда как в чистом воздухе видимость может составлять километры и выходить за пределы атмосферы.
Различные коэффициенты светопреломления воздуха и воды приводят к тому, что в воздухе и в воде принципиально различна роль роговицы глаза как элемента оптической системы глаза. В воздухе у наземных млекопитающих наружная сферическая поверхность роговицы является эффективным светопреломляющим элементом, поскольку эта поверхность разделяет среды с существенно различающимися коэффициентами светопреломления: воздух с коэффициентом 1.00 и переднюю камеру глаза, содержимое которой имеет практически такой же коэффициент светопреломления, как вода, т.е. 1.34. При этом наружная поверхность роговицы действует как собирающая линза, в значительной степени ответственная за получения сфокусированного изображения на сетчатке. Такая оптическая схема заложена, в частности, в общепринятый “схематический глаз” Гуллстранда для человека (Vojniković, Tamajo, 2013) и в “схематический глаз” кошки и других лабораторных животных (Hartridge, Yamada, 1922; Vakkur et al., 1963; Vakkur, Bishop, 1963; Hughes, 1979).
В воде у водных млекопитающих роль наружной поверхности роговицы как собирающей линзы минимальна или отсутствует, поскольку эта поверхность разделяет среды с практически одинаковыми коэффициентами светопреломления: воду и водянистое содержимое передней камеры глаза. В этом случае основным фокусирующим элементом оптической системы глаза остается хрусталик. Для обеспечения фокусировки без участия роговицы хрусталик приобретает сильновыпуклую, практически сферическую форму. Такая оптическая схема характерна для глаза китообразных (Dawson et al., 1972; Dawson, 1980; Kröger, Kirschfeld, 1992, 1993). Более детальные исследования показали, что у китообразных роговица все же играет некоторую роль в фокусировании изображения, поскольку коэффициент преломления роговичной ткани выше, чем у воды. Роговица действует как рассеивающая линза небольшой силы (Kröger, Kirschfeld, 1994). Однако вклад этой линзы в общую картину светопреломления невелик.
Таким образом, водное и воздушное зрение требуют разных схем оптики глаза. Глаз со слабовыпуклым (чечевицеобразным) хрусталиком может быть эмметропичным в воздухе, поскольку к диоптрийности хрусталика добавляется диоптрийность роговицы, и вместе они обеспечивают фокусирование изображения на сетчатке. В воде диоптрийность роговицы приближается к нулю, диоптрийности одного только хрусталика недостаточно для фокусирования изображения, и глаз сильно гиперметропичен. Напротив, сильновыпуклый (сферический) хрусталик эффективно фокусирует изображение на сетчатке без участия роговицы, поэтому глаз эмметропичен в воде, но в воздухе добавляется диоптрийность роговицы, и глаз сильно миопичен.
Из двух упомянутых групп водных млекопитающих – китообразных и сиреновых – наибольший интерес представляет оптическая система глаза китообразных, а среди них – зубатых китообразных (Odontoceti): зубатых китов, дельфинов и морских свиней. Зубатые китообразные – активные хищники с хорошо развитыми системами экстероцепции, в том числе с хорошо развитым зрением. Их зрительная система интересна тем, что, будучи адаптирована к водной среде, она сохранила способность успешно функционировать в воздухе. Хотя все китообразные ведут полностью водный образ жизни, воздушный тип дыхания привязывает их к приповерхностному слою воды. Они активно пользуются зрением не только под водой, но и в моменты выныривания на поверхность (Madsen, Herman, 1980). Поэтому значительный интерес представлял вопрос: каким образом оптика глаза китообразных обеспечивает им амфибийное (действующее как в воде, так и в воздухе) зрение, несмотря на различные коэффициенты светопреломления этих сред.
Амфибийное зрение китообразных объяснялось специфической конструкцией глаза, включающей согласованные свойства роговицы, зрачка и сетчатки. При слабом освещении (что обычно имеет место под водой) зрачок расширен, как и у всех других наземных и водных млекопитающих. Однако при ярком освещении (что имеет место над поверхностью воды) зрачок сокращается до двух небольших отверстий, расположенных назально и темпорально относительно оптической оси глаза. Эти два отверстия расположены напротив периферических участков роговицы, которые ввиду особенностей прикрепления роговицы к жесткой склере имеют минимальную кривизну (Herman et al., 1975; Dawson et al., 1979). На воздухе эти минимально искривленные участки роговицы действуют как очень слабые линзы, не создающие существенной миопии. Такой способ получения амфибийного зрения потребовал существенного изменения всей конструкции глаза: у китообразных глазной бокал уплощен в аксиальном направлении, имеет вид полусферы, концентричной с хрусталиком, а области наилучшего видения на сетчатке (зоны максимальной концентрации ганглиозных клеток) отодвинуты от оптической оси глаза (Mass, Supin, 2007). Исключение составляет амазонский речной дельфин Inia geoffrensis, имеющий круглый зрачок и шарообразную форму глазного бокала, как у наземных млекопитающих, и одну зону наилучшего видения (Dawson,1980; Dral,1981; Mass, Supin, 1989).
Приведенное выше объяснение амфибийности зрения китообразных может быть окончательно подтверждено, если рассчитать ход световых лучей с учетом всех элементов оптической системы глаза. Такой расчет не производился в предшествующих работах, и он являлся целью настоящего исследования.
МАТЕРИАЛ И МЕТОДЫ
В работе использованы образцы глаз четырех видов китообразных: бутылконосого дельфина (афалины) Tursiops truncatus; белухи Delphinapterus leucas; косатки Orcinus orca и морской свиньи Phocoena phocoena. Все эти виды относятся к подотряду зубатых китообразных, но к разным семействам: Delphinidae (бутылконосый дельфин и косатка), Monodontidae (белуха) и Phocoenidae (морская свинья). Для измерений использовано по одному образцу каждого вида. В тех случаях, когда были доступны несколько образцов, они были использованы для других целей (гистологические исследования сетчатки). Образцы были получены от животных, содержавшихся в коммерческих океанариумах и погибших от болезней, не связанных с поражением зрительной системы. Преднамеренное умерщвление животных для получения материала не производилось ни в одном случае.
Материал фиксировали в 10%-ном забуференном формалине. Перед извлечением глаза из орбиты отмечали ориентацию глазного яблока – помечали его дорсальный, вентральный, назальный и темпоральный полюсы. Извлеченный глаз фотографировали. Затем образец замораживали. На замораживающем микрооме срезали слой за слоем, параллельно оси глаза, до тех пор, пока не доходили до геометрической оси. Полученный продольный срез глаза фотографировали. Последующие измерения выполняли по полученным цифровым фотографиям.
Расчеты основывались на оптической схеме глаза, содержащей четыре светопреломляющих элемента: фронтальную и обратную поверхности роговицы, фронтальную и обратную поверхности хрусталика. Точный расчет хода световых лучей в такой системе – достаточно сложная задача, поскольку для ее решения необходимы точные значения многих параметров, включая отклонения поверхностей от сферической формы, неравномерность коэффициентов светопреломления в различных слоях роговицы и хрусталика, зависимость коэффициентов светопреломления сред глаза от длины световой волны. Многие из этих параметров не были известны для глаз китообразных. Однако задача может быть существенно упрощена, если рассматривать преломление только узкого параксиального (прилежащего к оптической оси системы) пучка световых лучей. В этом случае светопреломляющие поверхности могут быть аппроксимированы сферами, сферической и хроматической аберрациями можно пренебречь, и расчеты можно выполнить, используя простейшие формулы геометрической оптики. Эти формулы включают только радиусы светопреломляющих поверхностей и расстояния между этими поверхностями по оптической оси, а также коэффициенты светопреломления оптических структур.
Измерения и расчеты производили следующим образом. По данным морфологических измерений определяли радиусы фронтальной и обратной поверхностей роговицы, фронтальной и обратной поверхностей хрусталика. Непосредственное измерение коэффициентов светопреломления тканей роговицы и хрусталика было невозможно на имеющемся фиксированном материале, поэтому соответствующие данные брали из литературы. Коэффициенты светопреломления воздуха и воды – общеизвестные физические константы. С использованием этих данных рассчитывали положение сфокусированного изображения бесконечно удаленного объекта. Изображение могло оказаться перед сетчаткой, на ней или за сетчаткой (в последнем случае – мнимое изображение).
Измеряли следующие параметры: расстояния вдоль оси глаза между светопреломляющими поверхностями (фронтальная и обратная поверхности роговицы, фронтальная и обратная поверхности хрусталика); радиусы тех же светопреломляющих поверхностей.
Для измерения радиусов контур поверхности на фотографии продольного среза аппроксимировали дугой. Радиус аппроксимирующей дуги принимали за радиус поверхности.
Для расчета применяли элементарные формулы геометрической оптики. Сначала рассчитывали фокусные расстояния каждой из светопреломляющих поверхностей. Поскольку коэффициенты светопреломления сред перед преломляющей поверхностью и за ней различались, переднее и заднее фокусные расстояния рассчитывались отдельно. Переднее фокусное расстояние равно
и заднее фокусное расстояние равно где f1 и f2 – соответственно переднее и заднее фокусное расстояния, r – радиус поверхности, n1 и n2 – коэффициенты светопреломления сред перед поверхностью и за ней. Для поверхности, у которой выпуклость обращена в сторону прихода светового пучка, радиус r принимали как позитивный, а для поверхности обратной кривизны – негативный. Коэффициент светопреломления для воздуха принимали равным 1.00, для воды, жидкости передней камеры глаза и стекловидного тела 1.34. На основании литературных данных (Kröger, Kirschfeld, 1994), коэффициент светопреломления для ткани роговицы принимали равным 1.45, а для хрусталика 1.53.Положение реального или мнимого изображения, создаваемого преломляющей поверхностью, вычисляли по формуле Ньютона:
где f1 и f2 – переднее и заднее фокусные расстояния, полученные по формулам (1) и (2), a1 и a2 – расстояния соответственно от объекта до передней фокальной точки и от задней фокальной точки до изображения. Поэтому расстояние от объекта до преломляющей поверхности (на оптической оси) равно а расстояние от этой поверхности до изображения равноДля системы, состоящей из нескольких последовательно расположенных элементов (преломляющих поверхностей), расчет производился таким образом, что реальное или мнимое изображение, создаваемое одной из поверхностей, принималось за объект для следующей преломляющей поверхности. В данном случае, в предположении, что система включает четыре преломляющих поверхности, такая процедура последовательно выполнялась 4 раза, причем для первого элемента (наружная поверхность роговицы) объект считался бесконечно удаленным.
Для проверки корректности используемой процедуры она была применена к глазу, анатомические и оптические характеристики которого хорошо известны. В качестве такого образца был использован “схематический глаз” кошки, созданный на основе измерений многих экземпляров (Vakkur et al., 1963; Vakkur, Bishop, 1963). Исходные данные для расчета были взяты из работы (Vakkur, Bishop, 1963). Они приведены на рис. 1 и в таблице 1. Последовательные этапы вычислений дали такие результаты.
Рис. 1.
Схема основных оптических элементов глаза кошки.
Точки пересечения геометрической оси глаза со светопреломляющими поверхностями и сетчаткой: А – фронтальная поверхность роговицы, Б – обратная поверхность роговицы, В – фронтальная поверхность хрусталика, Г – обратная поверхность хрусталика, Д – рецепторный слой сетчатки. Е – расчетное положение сфокусированного изображения. Радиусы светопреломляющих поверхностей и сетчатки: r1 – фронтальная поверхность роговицы, r2 – обратная поверхность роговицы, r3 – фронтальная поверхность хрусталика, r4 – обратная поверхность хрусталика, r5 – сетчатка.
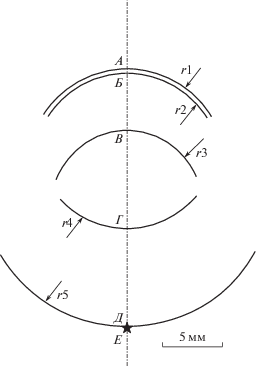
Таблица 1.
Исходные данные и результаты расчетов для оптической системы глаз кошки и китообразных
| Структуры и среда обитания | Кошка | Бутылконосый дельфин | Косатка | Белуха | Морская свинья |
|---|---|---|---|---|---|
| Коэффициенты светопреломления | |||||
| Роговица | 1.38 | 1.45 | 1.45 | 1.45 | 1.45 |
| Хрусталик | 1.55 | 1.53 | 1.53 | 1.53 | 1.53 |
| Радиусы, мм | |||||
| Роговица, фронтальная | 8.75 | 30.0 | 20.0 | 30.0 | 20.0 |
| Роговица, обратная | 7.89 | 15.0 | 18.5 | 8.0 | 15.0 |
| Хрусталик, фронтальная | 7.20 | 4.3 | 4.3 | 4.0 | 4.0 |
| Хрусталик, обратная | –8.05 | 4.3 | 5.7 | 4.0 | 4.0 |
| Расстояния, мм | |||||
| Толщина роговицы | 0.68 | 1.5 | 0.7 | 0.6 | 0.7 |
| Роговица – хрусталик | 4.52 | 1.4 | 1.5 | 1.6 | 0.5 |
| Хрусталик, осевая длина | 8.50 | 8.6 | 12.6 | 8.0 | 8.5 |
| Хрусталик – сетчатка | 8.45 | 13.0 | 15.5 | 12.0 | 11.8 |
| Положение изображения: расстояние от хрусталика, мм | |||||
| Вода, по оси | 13.5 | 13.1 | 12.6 | 12.5 | |
| Вода, наклон | 13.0 | 13.0 | 12.1 | 12.1 | |
| Воздух, по оси | 8.61 | 8.1 | 9.7 | 7.1 | 12.1 |
| Воздух, наклон | 13.0 | 13.0 | 12.1 | 9.0 | |
Преломление на фронтальной поверхности роговицы. Радиус этой поверхности составляет 8.75 мм, а коэффициент светопреломления ткани роговицы равен 1.38. При таких параметрах расчет согласно уравнениям (1) и (2) дает результаты: f1 = = 22.55 мм и f2 = 31.12 мм. Для бесконечно удаленного объекта (1/a1 = 0), a2 = 0, т.е. изображение создается на расстоянии 31.12 мм от фронтальной поверхности роговицы.
Преломление на обратной поверхности роговицы. При толщине роговицы 0.68 мм изображение, созданное ее фронтальной поверхностью, находится на расстоянии 30.44 мм от обратной поверхности. Радиус этой поверхности равен 7.89 мм. При коэффициентах светопреломления n1 = 1.38 и n2 = 1.34, получаем f1 = −272/.2 мм и f2 = −264.3 мм. При таких фокусных расстояниях объект, удаленный от преломляющей поверхности на 30.12 мм, создает изображение на расстоянии 33.28 мм от обратной поверхности роговицы.
Преломление на фронтальной поверхности хрусталика. При расстоянии между роговицей и хрусталиком ( равно 4.52 мм) изображение, созданное обратной поверхностью роговицы, находится на расстоянии 28.76 мм от фронтальной поверхности хрусталика. Радиус фронтальной поверхности хрусталика равен 7.20 мм. При n1 = 1.34 и n2 = 1.55 получаем: a1 = 45.94 мм и a2 = 53.14 мм, т.е. после прохождения светового пучка через фронтальную поверхность хрусталика изображение переместилось на расстояние 20.46 мм от этой поверхности.
Преломление на обратной поверхности хрусталика. При аксиальной толщине хрусталика 8.50 мм изображение, созданное его фронтальной поверхностью, находится на расстоянии –11.96 мм от обратной поверхности. Радиус обратной поверхности хрусталика равен 8.05 мм. При n1 = 1.55 и n2 = 1.34 получаем a1 = 59.42 мм и a2 = 51.37 мм. Таким образом, после прохождения светового луча через обратную поверхность хрусталика изображение переносится на расстояние 8.61 мм от этой поверхности.
Согласно схеме глаза кошки (Vakkur, Bishop, 1963), расстояние от обратной поверхности хрусталика до сетчатки составляет 8.45 мм. Расчет помещает изображение на расстояние 8.61 мм. Расхождение составляет всего 0.16 мм. Принимая во внимание возможные погрешности измерений, а также возможность аккомодации, такое расхождение можно считать пренебрежимым. Таким образом, для известного объекта использованная процедура расчетов дает ожидаемые результаты и поэтому может считаться корректной.
РЕЗУЛЬТАТЫ
Оптическая схема глаза китообразных приведена на рис. 2. Приведен глаз бутылконосого дельфина. Характерными особенностями примененной схемы являются: роговица, выпуклая в центре и менее выпуклая на краях; зрачок, при сужении сводящийся к двум небольшим отверстиям напротив уплощенных участков роговицы; сильновыпуклый, практически сферический хрусталик; глазной бокал в виде полусферы, концентричной с хрусталиком.
Рис. 2.
То же, что рис. 1 для глаза бутылконосого дельфина, кроме
Е – изображение на геометрической оси в воде, Е ' – изображение на геометрической оси в воздухе, Е1 – изображение, создаваемое наклонным пучком световых лучей.
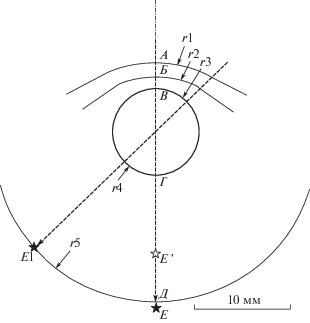
Расчеты выполнены для двух вариантов внешней среды (вода и воздух) и двух направлений хода световых лучей (по оси глаза через расширенный зрачок и наклонно через уплощенные участки роговицы и отверстия суженного зрачка), т.е. всего четыре варианта.
Для светового пучка, проходящего вдоль оси глаза через выпуклую поверхность роговицы, получено значительное различие между водной и воздушной внешней средой. Для водной среды (коэффициент светопреломления 1.34) расчет показал положение сфокусированного изображения (точка Е на рис. 2) на расстоянии 13.5 мм от обратной поверхности хрусталика. Расстояние от обратной поверхности хрусталика до сетчатки (точка Д на рис. 2), по данным измерений, составило 13 мм. Таким образом, расчетное изображение практически совпадало с сетчаткой: расхождение составило всего 0.5 мм. В этой ситуации глаз должен быть охарактеризован как эмметропичный.
Иная ситуация имеет место для воздушной внешней среды (коэффициент светопреломления 1.00). Расчет показал положение сфокусированного изображения (точка Е’ на рис. 2) на расстояние 8.1 мм от обратной поверхности хрусталика, т.е. на значительном расстоянии перед сетчаткой. В этой ситуации глаз характеризуется значительной миопией.
Для наклонного светового пучка, проходящего через уплощенный участок роговицы, положение расчетного изображения одинаково для водной и воздушной сред (точка Е1 на рис. 2): оно находится на расстоянии 13.0 мм от обратной поверхности сферического хрусталика, т.е. практически на сетчатке. В этой ситуации глаз должен быть охарактеризован как эмметропичный.
Результаты измерений и расчетов для всех четырех исследованных видов китообразных приведены в таблице. Поскольку кривизна боковых участков роговицы была очень мала, точно измерить радиус этой кривизны не представлялось возможным. Поэтому при расчетах условно принимали, что кривизна как фронтальной, так и обратной поверхности боковых участков роговицы равна нулю. При этом для наклонных лучей, проходящих через плоские участки роговицы, положение сфокусированного изображения совпадает для воды и воздуха.
Результаты вычислений показали, что у всех четырех исследованных видов в воде сфокусированное изображение оказывается вблизи поверхности сетчатки или на ней. Такое положение сфокусированного изображения характерно как для пучка световых лучей, приходящего вдоль оси глаза, так и для наклонного пучка, хотя для наклонного пучка изображение оказывалось на несколько большем расстоянии, чем для осевого. Это различие обусловлено действием центральной части роговицы. Однако различие положений изображения для осевого и наклонного пучков невелико, т.е. в обоих случаях глаз может считаться эмметропичным.
Иная картина создается в воздухе. Для наклонных световых пучков, проходящих через уплощенные периферийные участки роговицы, сфокусированное изображение также оказывается вблизи сетчатки (эмметропия). Для осевого пучка, проходящего через выпуклую центральную часть роговицы, изображение оказывается на расстоянии нескольких мм перед сетчаткой (миопия).
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Результаты выполненных расчетов следует считать ориентировочными по следующим причинам: форма светопреломляющих структур, измеренная на фиксированных препаратах, может отличаться от формы тех же структур в интактном глазе; доступная точность измерений радиусов и расстояний не превышала 0.1 мм (в таблице все данные измерений приведены с разрешением 0.1 мм); для каждого вида был доступен только один образец, что не позволило учесть индивидуальные вариации; индексы светопреломления не измерялись для каждого из исследованных видов, а принимались одинаковыми на основе литературных данных; светопреломление в живом глазе может изменяться в результате аккомодации, что не могло быть учтено при измерениях на фиксированном материале.
Тем не менее результаты расчетов позволяют оценить справедливость предлагавшихся ранее гипотез о механизмах амфибийного зрения китообразных. Расчеты показали, что зрение не может быть амфибийным для световых лучей, проходящих по оси глаза: в этом случае светопреломление эмметропично только для водной среды, в которой фокусирующая роль выпуклой роговицы минимальна или отсутствует. Сферический или квазисферический хрусталик обеспечивает необходимую фокусировку. В воздухе выпуклая поверхность роговицы действует как дополнительная собирающая линза, и глаз становится миопичным. Поверхность роговицы не действует как собирающая линза для световых лучей, наклонных относительно оси глаза, так как эти лучи проходят через уплощенные участки роговицы.
Обеспечение амфибийности зрения за счет плоской роговицы – принцип, действующий не только у китообразных. Тот же принцип обеспечивает амфибийность зрения у ластоногих (Pinnipedia) – как настоящих тюленей (Phocidae), так и ушатых тюленей или морских львов (Otariidae). Однако в отличие от китообразных, у ластоногих этот принцип реализуется в другом варианте: уплощенный участок имеется в центре роговицы (Dawson et al., 1987; Sivak et al., 1989; Mass,1992; Mass, Supin, 1992, 2003, 2010; Hanke et al., 2006). В комбинации со сферическим или квазисферическим хрусталиком, плоская роговица обеспечивает удовлетворительную фокусировку изображений на сетчатке как в воде, так и в воздухе (Mass, 1992; Mass, Supin, 2003; 2007; 2010; Hanke et al., 2009). В результате ластоногие имеют удовлетворительное зрение как в воздухе (Hanke, Dehnhardt, 2009), так и под водой (Schusterman, Balliet, 1970; Weiffen et al., 2006).
Однако возможны и принципиально иные способы обеспечить амфибийное зрение. У выдр это возможно за счет значительного изменения формы хрусталика при сокращении радужки: увеличение кривизны фронтальной поверхности хрусталика может компенсировать отсутствие роговичной линзы под водой (Murphy et al., 1990; Mass, Supin, 2000). Тем самым обеспечивается удовлетворительное подводное зрение (Gentry, Peterson, 1967; Schusterman, Barrett, 1973).
Таким образом, различные группы водных и полуводных млекопитающих выработали разные способы обеспечения амфибийности зрения. Эти способы достаточно успешно обеспечивают им возможность использования зрения как на воздухе, так и под водой.
Работа выполнена при поддержке РФФИ (грант № 16-04-00-161).
Список литературы отсутствует.
Дополнительные материалы отсутствуют.
Инструменты
Сенсорные системы