

Сенсорные системы, 2019, T. 33, № 2, стр. 99-112
Нейрофизиологические механизмы сравнения двух последовательных ориентаций в задаче рабочей памяти
Е. С. Михайлова 1, *, Н. Ю. Герасименко 1
1 ФГБУН Институт высшей нервной деятельности и нейрофизиологии РАН
117485 Москва, ул. Бутлерова, 5А, Россия
* E-mail: esmikhailova@mail.ru
Поступила в редакцию 09.01.2019
Аннотация
Память на ориентации является важной составляющей зрительно-пространственного поведения человека, позволяя сохранять в актуализированной форме релевантную для этой деятельности информацию. На 33 здоровых испытуемых с нормальным зрением исследовали роль низкоуровневых сенсорных и высокоуровневых префронтальных областей коры в операции сличения текущей и удерживаемой в памяти ориентаций, что критически важно для обнаружения изменений и формирования быстрых адаптационных реакций. Стимулами являлись прямоугольные решетки вертикальной, горизонтальной и наклонной (45°) ориентаций. Установлено, что информативными показателями несовпадения последовательных ориентаций являются значимое повышение амплитуды раннего волнового комплекса Р100/N150 в зрительных областях коры и нарастание амплитуды негативности N240 в префронтальных областях, что, по данным моделирования дипольных источников, сопровождается усилением активности ряда структур префронтальной коры: middle frontal gyrus, frontal pole, pars orbitalis. Таким образом, обнаружение несовпадения текущего сигнала и удерживаемой в памяти информации осуществляется за счет совместного участия зрительной нейросети каудальных областей коры и нейросети исполнительного контроля префронтальных областей, которая включает эту информацию в программу целенаправленного поведения. Полученные данные подчеркивают важность тесного взаимодействия сенсорных и префронтальных областей в зрительной рабочей памяти.
ВВЕДЕНИЕ
Ориентационные свойства являются важными описательными характеристиками объектов зрительного мира, а их сохранение – обязательной составляющей памяти о сложных изображениях и сценах. Сохранение в кратковременной памяти ориентационных свойств обеспечивает механизм поддержания стабильных зрительных репрезентаций при естественных процессах, неожиданно прерывающих сенсорный вход, таких, например, как саккадические движения глаз, фиксации, окклюзии (Luck, Vogel, 1997; Hollingworth et al., 2008). Возможность быстрого обнаружения изменений ориентаций обеспечивает функционирование референтных систем мозга, в координатах которых определяется положение объекта в пространстве, программируются движения глаз и конечностей (Yang et al., 2013; Ma et al., 2011; Christophel et al., 2017). Памяти на ориентационные характеристики уделялось сравнительно небольшое внимание, если сравнивать объем этих работ с объемом всех исследований зрительной рабочей памяти. Норвежским ученым С. Магнуссеном была предложена модель кратковременной памяти на базовые зрительные характеристики (пространственная частота, контраст, ориентация), обозначенная им как “низкоуровневая перцептуальная память” (“low-level perceptual memory”) (Magnussen, 2000; 2009). Согласно этой модели, информация об элементарных признаках стимула сохраняется в сети независимых сенсорных хранилищ, каждое их которых настроено на определенную характеристику стимула. Индивидуальные элементы в континууме измеряемой характеристики встроены в сеть с латеральным торможением, и такое устройство обеспечивает торможение внутри измерений, но не между ними. Низкоуровневая перцептуальная память основана на сохранении таких независимых ресурсов. При ее участии осуществляется имплицитная дискриминация простых зрительных признаков, которая включает поиск и нахождение в памяти низкоуровневых нейронных репрезентаций и их сравнение с характеристиками текущего стимула.
Чем определяется необходимость запоминания и хранения в памяти элементарных признаков зрительного стимула или сцены? Одно из возможных объяснений – это кратковременная операция, сопровождающая и необходимая для формирования высокоточной долговременной памяти на структурно когерентные образы или значимые паттерны (Magnussen, 2000). Согласно этой гипотезе, низкоуровневая перцептуальная память поддерживает perceptual representation system (PRS), расположенную до уровня полного описания объекта (Schacter et al., 1993). Эта поддерживающая система хранения низкоуровневой информации может не ограничиваться диапазоном кратковременной памяти, а участвовать в организации долговременной памяти, занимающей периоды недель и даже месяцев. Вторая возможность – низкоуровневая память – представляет отдельный параллельный имплицитный механизм долговременной памяти, который сохраняет более детальную информацию о зрительном образе и поддерживает иконическую память (pictorial memory) и память зрительных сцен.
Нейрофизиологический анализ памяти на низкоуровневые зрительные характеристики у человека представлен преимущественно данными функциональной МРТ. Полученные результаты убедительно демонстрируют роль ранних зрительных областей в удержании этой информации, хотя неоднозначны в определении корковой локализации процессов. Ряд авторов в задаче на зрительную память описали активацию экстрастриарных областей, расположенных над V1 (Pasternak, Greenlee, 2005). Вместе с тем Оффен и соавт. (Offen et al., 2009) при сравнении активации в задачах на кратковременную память и дискриминацию стимулов, требующей только внимания, не нашли устойчивой активации V1–V3 для процедуры кратковременной зрительной памяти, но обнаружили ее при дискриминации. Напротив, Серенсес и соавт. (Serences et al., 2009), используя повоксельный (voxel-by-voxel) анализ, обнаружили в период удержания повышение уровня BOLD сигнала в ранних зрительных областях, включая V1. Более того, ими было показано, что характер активации зависит от ориентации, удерживаемой в памяти. Эстер и cоавт. (Ester et al., 2009) описали сходный эффект исключительно для активности области V1. В работе Пратте и Тонг (Pratte, Tong, 2014) для рабочей памяти показана характерная для зрительного восприятия пространственная специфичность активации ранних зрительных областей (V1, V2). По их данным, распознавание запомненной ориентации по BOLD-сигналу в период удержания выше в зрительных областях контралатеральных стимулируемому полуполю зрения, т.е. повторяет паттерн, свойственный простому наблюдению. В целом два десятилетия исследований памяти на низкоуровневые зрительные характеристики представили доказательства существования специфического механизма этого вида памяти, тесно связанного с механизмом зрительной дискриминации, локализованного на ранних уровнях пути переработки информации и предположительно организованного по типу набора параллельных независимых домен-специфичных механизмов хранения информации. Подобный механизм описан и для памяти на стимулы других сенсорных модальностей (Magnussen, 2009).
Наряду с представлениями о ведущей роли ранних зрительных областей в организации кратковременной памяти на ориентации и другие низкоуровневые зрительные характеристики, есть целый ряд работ, авторы которых, обсуждая свои данные, значительно расширяют круг участвующих структур. В работе (Solomon, Lennie, 2007) в задаче запоминания цвета или ориентации цветных ориентационных решеток показано, что характер активации зрительных областей соответствовал релевантной характеристике, что, по мнению авторов исследования, указывает на существование нисходящих управляющих влияний. В этом контексте следует особо выделить работу (Harrison, Tong, 2009), в которой авторы обращаются к вопросу о роли нисходящих управляющих влияний и функциональной связи низко- и высокоуровневых областей в рабочей памяти на ориентации. Основная цель их работы – исследование роли ранних зрительных областей в отставленной дискриминации ориентаций. Испытуемый должен был ответить, на сколько угловых градусов повернут тестовый стимул относительно удерживаемого в памяти эталонного (на ±3° или ±6°). Таким образом, сравнивая результаты для двух вариантов разницы “тестовый стимул – эталон”, авторы избегали влияния самой процедуры кратковременной памяти. Используя для анализа функциональной МРТ методы классификации паттернов (“pattern classification methods”), они показали, что по паттерну активации областей V1–V4 с вероятностью 80% можно предсказать, какую из ориентаций испытуемый удерживал в памяти, даже если уровень активации этих областей очень низок. При обсуждении результатов Харрисон и Тонг обращают внимание на роль префронтальной коры, которая интерпретирует подающийся перед эталоном абстрактный ключ, определяющий, какую из ориентаций надо запомнить. Авторы подчеркивают, что задача будущих исследований – это понять, поддерживается ли информация в низкоуровневых областях при участии рекуррентных связей между высоко- и низкоуровневыми областями или связей внутри низкоуровневых областей, или тех и других. Таким образом, зрительная кратковременная память дает возможность оценить связь между перцепцией и высшими когнитивными процессами. Эти результаты согласуются с данным о наличии в период удержания в памяти различных зрительных стимулов устойчивой активации в широкой сети префронтальных, теменных, нижневисочных и латеральных затылочных областях (Courtney et al., 1998; Curtis, D’Esposito, 2003; Xu et al., 2006; Ester et al., 2015).
Важно подчеркнуть, что в подавляющем большинстве работ авторы акцентировали внимание на этапе удержания информации и не затрагивали проблему сопоставления текущего сигнала с хранящейся в памяти информацией. Как пишут авторы обзора (Hyun et al., 2009), зрительная система человека может определять сходство или различия между удерживаемой в памяти репрезентацией предыдущего сигнала с текущей информацией, но механизм этого процесса остается до сих пор неясным.
Цель настоящей работы – используя высокоплотную регистрацию вызванной активности, определить временные характеристики и топографию процессов сличения и дискриминации текущей и удерживаемой в кратковременной памяти ориентаций, а также методом дипольного моделирования определить сеть структур мозга, обеспечивающих эту операцию. Важность поставленной задачи определяется необходимостью понимания механизма, с помощью которого зрительная система может выделить быстрые изменения одной из базовых характеристик зрительной среды.
МЕТОДИКА
В исследовании принимали участие 33 испытуемых (16 мужчин и 17 женщин) c нормальным зрением. Средний возраст – 22.57 ± 0.46 лет. От всех испытуемых было получено письменное согласие на проведение исследования, согласно протоколу, утвержденному этической комиссией ИВНД и НФ РАН. Исследования проводили c 9 до 15 ч.
Стимулы представляли собой черно-белые высококонтрастные прямоугольные решетки с наклоном линий 0° (горизонталь), 90° (вертикаль) и 45°. Размер стимула составил 5.5 угл. град. Для моделирования в эксперименте ситуации сравнения новой и удерживаемой в памяти информации испытуемым в случайном порядке предъявлялись пары решеток разной ориентации. Три пары решеток были совпадающими по ориентации (0 – 0°, 90 – 90° и 45 – 45°), и шесть пар были несовпадающими по ориентации (0 – 90°, 0 – 45°, 90 – 0°, 90 – 45°, и 45 – 0°, 45 – 90°). Первый стимул в паре – эталонный, и по инструкции испытуемый должен был его запомнить. Второй стимул в паре – тестовый.
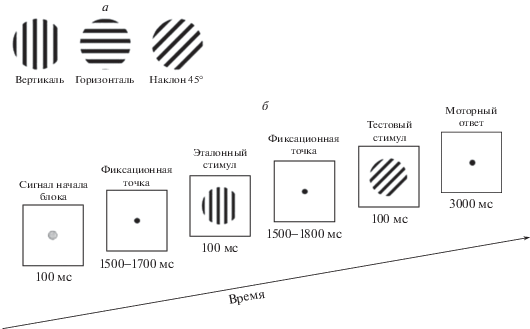
Во время исследования испытуемый сидел в удобном кресле в затемненной и звукозаглушенной комнате на расстоянии 120 см от экрана монитора Dell E1911c (диагональ 17 дюймов). Освещенность на уровне глаз испытуемого – 3 лк. Горизонтальные и вертикальные края корпуса монитора и экрана были закрыты экраном с закругленными краями и круглой апертурой. Стимулы предъявляли в центр экрана монитора при помощи программы E-Prime 2.0 (Psychology Software Tools, Inc., USA). Стимулы и временная схема эксперимента представлены на рис. 1. Каждый блок состоял из следующих событий: зеленая точка длительностью 100 мс, при появлении которой испытуемый должен был сосредоточиться и фиксировать взгляд в центре экрана; черная фиксационная точка, длительность которой в случайном порядке изменялась от 1500 до 1700 мс; эталонный стимул – 100 мс; интервал между эталонным и тестовым стимулами, длительность которого варьировала от 1500 до 1800 мс, в центре экрана – фиксационная точка; тестовый стимул – 100 мс; интервал между блоками 3000 мс, в это время в центре экрана – фиксационная точка. В это же время испытуемый отвечал на вопрос о сходстве эталонной и тестовой ориентаций, нажимая на кнопку. Каждую пару эталонного и тестового стимулов предъявляли по 30 раз, т.е. испытуемому предъявляли 90 пар совпадающих стимулов и 180 пар несовпадающих стимулов, всего 270 пар. Последовательность пар была случайной. Длительность всего эксперимента около 30 мин (всего 270 блоков, каждый длительностью 6200–6700 мс). В середине эксперимента делали перерыв 5–7 мин для отдыха испытуемого и коррекции сопротивления электродов.
Рис. 1.
Условия эксперимента.
а – зрительные стимулы – ориентационные решетки горизонтальной, вертикальной и наклонной ориентации; б – временная последовательность событий в одном блоке эксперимента рабочей памяти – попарного сравнения текущего и удерживаемого в памяти стимулов.
Согласно инструкции, стандартный текст которой зачитывали перед началом эксперимента, испытуемый должен был определить, совпадают или не совпадают по ориентации эталонный и тестовый стимулы и нажать при совпадении на одну, при несовпадении – на другую клавишу выносной клавиатуры. Номера клавиш сообщались в инструкции. Регистрировали правильность ответа и время нажатия на клавишу.
Регистрацию электроэнцефалограммы (ЭЭГ) проводили на 128-канальном энцефалографе фирмы Geodesics (Electrical Geodesics Inc., USA) при помощи шлема HydroCel Geodesic Sensor Net (Electrical Geodesics Inc., USA). Частота квантования сигнала 500 Гц. Сопротивление электродов не превышало 50 кОм. Полосу частот усилителя ограничивали сверху 70, а снизу 0.1 Гц, а также узкополосным заграждающим фильтром для частоты 50 Гц (“notch” фильтр). При записи в качестве индифферентного электрода использовали вертекс (Cz). При последующей “off-line” обработке записи в качестве референтного выбирали усредненный электрод.
Данные ЭЭГ обрабатывали в программе NetStation 4.5.4 (Electrical Geodesics Inc., USA). Предварительно фильтрованные (0.5–45 Гц) записи ЭЭГ каждого испытуемого сегментировали на отрезки длительностью 1300 мс, содержащие интервалы 300 мс до предъявления тестового стимула и 1000 мс после. Далее программно (Net Station EEG Software) исключали реализации с глазными движениями, для которых амплитуда ЭОГ превышала 80 мкВ. В дальнейшем при визуальном анализе исключали фрагменты с двигательными и электромиографическими артефактами. После этого для каждого испытуемого по безартефактным отрезкам ЭЭГ проводили усреднение вызванных потенциалов (ВП) на тестовый стимул для правильных ответов. Коррекцию базовой линии проводили по 300 мс отрезку ЭЭГ до предъявления тестового стимула. Для каждого испытуемого в эксперименте с рабочей памятью на ориентации получено по девять усредненных ВП: три для совпадающих эталона и теста и шесть для несовпадающих эталона и теста.
Амплитуду выбранных для анализа компонентов индивидуальных ВП измеряли в программе NetStation 4.5.4. Замеряли адаптивный минимум или максимум во временном окне компонента, а именно средние значения амплитуды в 4-миллисекундном интервале, соответствующем 2 мс до и после пика компонента. Анализировали амплитуду компонентов ВП, усредненную по кластерам электродов. Усреднение амплитуды проводили в симметричных фронтальных (в левом полушарии сенсоры №№ 21, 22, 25, в правом – №№ 8, 9, 14), теменных (в левом полушарии – №№ 52, 60, 67, в правом – №№ 77, 85, 92), височных (в левом полушарии – №№ 58,59,65, в правом – №№ 90, 91, 96) и затылочных (в левом полушарии – №№ 66, 70, 71, в правом – №№ 76, 83, 84) кластерах электродов. Приведенные номера отведений соответствуют номерам датчиков в 128-канальном шлеме Hydro Cel Geodesic Sensor Net.
Статистический анализ амплитуд компонентов индивидуальных ВП, усредненных по группам электродов, проводили с использованием дисперсионного анализа ANOVA RM (метод повторных измерений). При этом учитывали факторы: “совпадение” (два уровня – совпадение и несовпадение эталона и теста); “полушарие” (два уровня); “ориентация” тестового стимула (три уровня – вертикаль, горизонталь, наклонная ориентация 45°). Значения анализируемых показателей для ситуации несовпадения эталонной и тестовой ориентаций вычисляли как среднее для двух вариантов несовпадений. Например, для тестового стимула – вертикаль – ситуация несовпадения вычислялась как среднее значение показателей в двух парах “эталон горизонталь – тест вертикаль” и “эталон 45° – тест вертикаль”. Результаты ANOVA при необходимости (Mauchly sphericity test) были скорректированы поправкой Гринхауса-Гейссера. При post-hoc сопоставлениях использовали поправку Ньюмена-Кейлса на множественные сравнения (http://medstatistic.ru/articles/kratkiy_kurs.pdf). При анализе точности ответа и ВР использовали дисперсионный анализ ANOVA RM и Т-тест для связанных выборок.
Процедуру моделирования распределенных дипольных источников активности во временном окне компонента N240 (200–270 мс) проводили в пакете Brainstorm 3.2. (Biomedical Imaging Group) программной среды MatLab. Использовался метод расчета wMNE (L2-weightened Minimum Norm Estimates) со стандартной анатомией ICBM 152, реалистичной объемной моделью головы OpenMEEG BEM и стандартными координатами электродов, соответствующими схеме ASA-10-20. Проводили построение объемных распределенных источников (распределение активации в объеме мозга) с их проекцией на поверхность коры — поверхностные распределенные источники. Для моделирования использовали усредненные по группе ВП на каждую из трех ориентаций при правильных ответах в ситуациях совпадения и несовпадения эталонной и тестовой ориентаций. Строили распределение плотности источников (“current sources density”) в период времени, соответствующий развитию компонентов N240 (200–280 мс) в ВП фронтальных отделов. Для соотнесения результатов моделирования с анатомическими структурами мозга поверхностные распределенные источники проецировались на атлас зон интереса Desikan-Killiany (Desikan et al., 2006). Активными считались структуры, плотность тока источников в которых была выше 50% максимальной в рассматриваемый момент времени. Подробное описание методики построения источников приведено в (Крылова и др., 2015).
РЕЗУЛЬТАТЫ
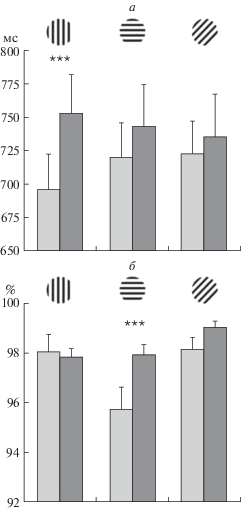
Анализ психофизиологических показателей. Проведен дисперсионный анализ ВР и точности с учетом факторов “совпадение” (совпадение и несовпадение эталонной и тестовой ориентаций) и “ориентация” тестового стимула (горизонталь, вертикаль, наклон 45°). Выявлены значимые эффекты “совпадения” (F1,30 = 7.055; p < 0.01), “ориентации” (F2,60 = 3.44; p < 0.05) и взаимодействие “совпадение” × “ориентация” (F2,60 = 3.81; p < 0.05). На рис. 2, а видно, что в ситуации несовпадения ВР больше, чем при совпадении. При этом уровень значимости различий выше для случаев, когда тестовой ориентацией была вертикаль (p < 0.001, T-test paired). Для тестовых горизонтальной и наклонной ориентаций значимые различия отсутствовали. Вероятность правильного ответа была достаточно высокой – от 95 до 99%. Выявлен значимый эффект “совпадения” (F1,31 = = 4.35; p < 0.05). При несовпадении эталона и теста точность выше, чем при совпадении: для горизонтали 97.95 ± 0.39 и 95.88 ± 0.85 (p < 0.005); для вертикали 97.84 ± 0.35 и 97.65 ± 0.80; для наклонной ориентации 99.02 ± 0.25 и 98.14 ± 0.47, соответственно (рис. 2, б).
Рис. 2.
Психофизические характеристики выполнения задачи сличения двух ориентаций.
а – время двигательной реакции; б – вероятность правильного ответа при совпадении и несовпадении тестовой и эталонной ориентаций в задаче попарного сравнения. Светлые столбики – совпадение, темные – несовпадение. Достоверность статистических различий: *** – р < 0.001 (paired t-test). Приведены средние по группе значения и ошибки среднего.
Анализ компонентов Р100 и N150 в каудальных областях коры. На первом этапе анализировали регионарные различия эффектов “совпадения” эталонной и тестовой ориентаций. ANOVA RM с факторами “область” (три уровня: затылочные, височные, теменные), “полушарие” (два уровня), “совпадение” (два уровня) показало высокозначимый эффект “области” для амплитуды компонентов Р100 (F2,58 = 19.28; p < 0.0005) и N150 (F2,62 = 18.70; p < 0.0005). С учетом полученного мощного влияния “области”, что указывает на выраженные регионарные различия переработки информации, последующий анализ проводили раздельно для затылочных, теменных и височных групп отведений.
Анализ компонента Р100. Проведен дисперсионный анализ амплитуды Р100 коры с учетом факторов “совпадение”, “ориентация” и “полушарие” раздельно для трех областей. Основной эффект “совпадения” во всех анализируемых областях достигает высокого уровня значимости (р < < 0.0001) и не зависит от “ориентации”, о чем говорит отсутствие взаимодействия “совпадение” × × “ориентация”. На рис. 3, а приведены усредненные ВП правой затылочной области при трех вариантах тестовой ориентации (вертикаль, горизонталь и наклон 45°) при совпадении и несовпадении эталона и теста. Видно, что в ситуации несовпадения амплитуда Р100 увеличивалась для всех трех ориентаций тестового стимула. С учетом того, что в затылочной области эффект “ориентации” близок к значимости (F2,62 = 2.93; p = = 0.06), было проведено post-hoc сопоставление амплитуды Р100 при совпадении и несовпадении ориентаций. Показано, что увеличение амплитуды Р100 значимо для всех ориентаций: для вертикали (p < 0.001), для горизонтали (р < 0.01 в правом и p < 0.005 в левом полушарии) и для 45° (p < 0.05).
Рис. 3.
Вызванные потенциалы при совпадении и несовпадении тестовой и эталонной ориентаций.
а – ВП правой затылочной области для решеток трех ориентаций в тестовом стимуле; б – ВП затылочной, височной и теменной областей правого полушария, усредненные для трех ориентаций в тестовом стимуле. На а и б – сплошная линия – совпадение тестовой и эталонной ориентаций, пунктирная линия – несовпадение. Вертикальной линией отмечено начало тестового стимула.
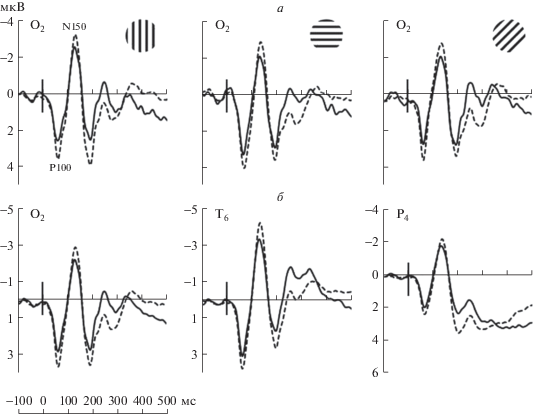
Анализ компонента N150. По данным дисперсионного анализа эффект “совпадения” статистически значим для амплитуды N150 во всех анализируемых областях (0.0005 < p < 0.05). Так же как и для Р100, несовпадение тестовой ориентации и эталона вызывает увеличение амплитуды N150, что отчетливо видно на ВП затылочной, теменной и височной областей правого полушария, усредненным по трем ориентациям (рис. 3, б). Значимость эффекта “совпадения” выше в затылочной и височной областях (F1,32 = 18.43; р < < 0.0001 и F1,32 = 19.2; р < 0.0001, соответственно) по сравнению с теменной (F1,32 = 6.01; р < 0.05). В теменной области выявлено взаимодействие “ориентация” × “полушарие” (F2,62 = 10.53; p < < 0.0005). По данным post-hoc анализа, для вертикальной ориентации амплитуда N150 выше в правом полушарии как при совпадении (p < 0.05), так и при несовпадении (p < 0.01) эталонной и тестовой ориентаций.
Для уточнения регионарной специфики влияния совпадения/несовпадения эталона и теста был проведен дисперсионный анализ амплитуды компонентов Р100 и N150 с учетом факторов “совпадение” и “область”. Для Р100 показано взаимодействие “совпадение” × “область”: в правом полушарии – F2,58 = 3.55; p < 0.05, в левом – F2,58 = 3.74; p < 0.05. Как видно на рис. 4, а, для амплитуды Р100 значимые различия между ситуациями совпадения и несовпадения есть во всех каудальных областях, но они более отчетливы в затылочных.
Рис. 4.
Влияние совпадения и несовпадения тестовой и эталонной ориентаций на амплитуду компонентов Р100 и N150 ВП на тестовый стимул в задаче рабочей памяти.
Приведены средние по группе значения и ошибка среднего для амплитуды компонентов Р100 (а) и N150 (б) в ВП затылочной, теменной и височной областях правого (ПП) и левого (ЛП) полушарий. Светлые столбики – совпадение, темные – несовпадение. Достоверность различий: * – р< 0.05; ** – p < 0.01 (тест Ньюмана-Кейлса).
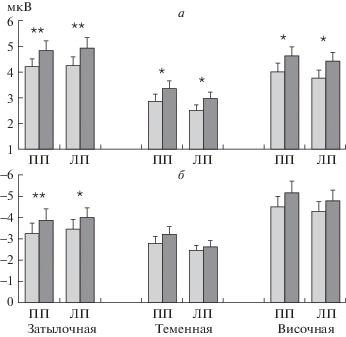
Для амплитуды N150 взаимодействие “совпадение” × “область” значимо в правом полушарии при F2,62 = 5.46; p < 0.01, в левом – при F2,64 = = 7.76; p < 0.001. Как показал последующее post-hoc сопоставление, увеличение амплитуды N150 статистически значимо только в затылочной области (рис. 4, б).
Анализ компонента N240 во фронтальных областях коры. Проведен дисперсионный анализ амплитуды компонента N240 в ВП лобных областей на тестовую ориентацию, усредненной по правому и левому кластерам лобных отведений. Так же, как для амплитуд Р100 и N150 в каудальных областях коры, значение амплитуды N240 для несовпадения эталонной и тестовой ориентаций вычислялась как среднее для двух ориентаций эталонного стимула, несовпадающих с тестовым. В качестве факторов внутрииндивидуальной вариабельности выбраны “совпадение”, “полушарие”, “ориентация”.
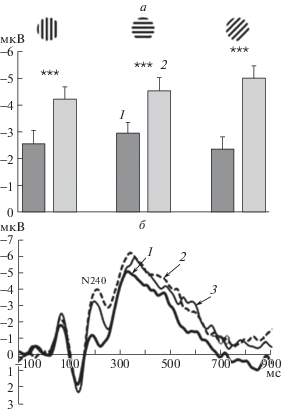
Проведенный дисперсионный анализ показал, что “совпадение” значимо влияет на амплитуду N240. Как видно на представленной на рис. 5 диаграмме амплитуды N240, в правой фронтальной группе электродов (а) и на усредненных ВП правого лобного полюсного отведения Fp2 (б), при несовпадении ориентаций амплитуда N240 выше, чем при их совпадении. Влияние “совпадения” высокозначимо (F1,31 = 54.05; p < 0.0005) и зависит от “ориентации”, о чем говорит взаимодействие “совпадение” × “ориентация” (F2,62 = = 3.85; p < 0.05).
Рис. 5.
Влияние совпадения и несовпадения тестовой и эталонной ориентаций на амплитуду компонента N240 в правой лобной группе отведений.
a – средние по группе значения амплитуды компонента N240 в правой лобной группе отведений. На оси абсцисс – тестовые ориентации. 1 – совпадение эталонной и тестовой ориентаций; 2 – несовпадение ориентаций. Достоверность различий: *** – р < 0.001 (по Ньюману–Кейлсу). Приведены средние по группе значения и ошибка среднего. б – усредненные ВП правого полюсного лобного отведения (Fp2) на тестовую ориентацию при ее совпадении и несовпадении с эталоном. Представлены ВП для тестовой ориентации – вертикаль. Цифрами обозначены три варианта эталонной ориентации: 1 (сплошная толстая линия) – вертикальная (совпадение с эталоном); 2 (пунктирная линия) – наклонная 45° (несовпадение с тестом); 3 (сплошная тонкая линия) – горизонтальная (несовпадение с тестом).
Моделирование распределенных дипольных источников компонента N240. На рис. 6 представлены результаты моделирования распределенных дипольных источников компонента N240 усредненных по группе ВП для тестовой ориентации вертикаль при совпадении (а) и несовпадении (б) эталона и теста. Модель строилась для периода времени, соответствующего развитию компонента N240 (200–280 мс). Представленные на рисунке карты построены для момента времени 254 мс после начала тестового стимула, что соответствует наиболее отчетливой разнице между ситуациями совпадения и несовпадения эталонного и тестового стимулов.
Рис. 6.
Моделирование распределенных источников компонента N240 методом wMNE. Приведены объемные распределенные источники компонента N240 на тестовую ориентацию на аксиальном, саггитальном и коронарном срезах стандартной анатомии головы ICBM 152. Время – 254 мс после начала стимула. а – совпадение тестовой и эталонной ориентаций (эталон-вертикаль, тест-вертикаль); б – несовпадение тестовой и эталонной ориентаций (эталон-вертикаль, тест-45º). На фрагменте б видно, что при несовпадении эталонной и тестовой ориентаций в префронтальной коре возникает дополнительная область активации.
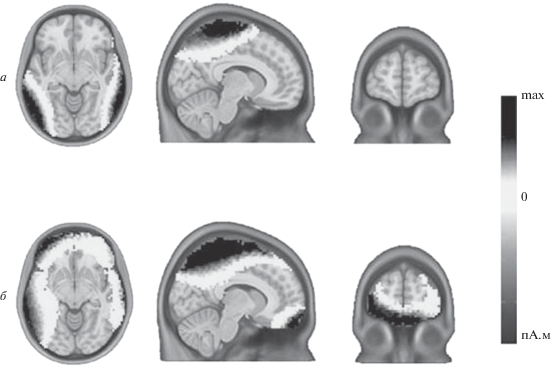
Результаты проекции распределенных дипольных источников N240 на корковые зоны (по атласу Desikan-Killiany) показали, что на этом этапе максимальные значения плотности дипольного тока локализуются в зонах интереса (ROIs) теменной, височной и лобной областей коры. В теменной области это superior parietal, precentral, postcentral gyri, в височной – supperior, middle и inferior temporal gyri. Эти зоны интереса активируются при совпадении и при несовпадении эталонного и тестового стимулов (рис. 6, а и б). При несовпадении эталона и теста в префронтальной коре дополнительно активируются следующие зоны интереса: rostral middle frontal, frontal pole и parsorbitalis gyri, а в височной доле возникает межполушарная асимметрия в виде левостороннего преобладания активации. Эти изменения топографии активации отчетливо видны на рис. 6, б.
ОБСУЖДЕНИЕ
Проведенный в работе анализ точности и ВР при выполнении задачи определения сходства или различия двух последовательных ориентаций в рабочей памяти выявил значительное увеличение латентности двигательной реакции в случае их несовпадения, что соответствует результатам других авторов (Zhang et al., 2003; Wang et al., 2001; Yin et al., 2011).
Анализ амплитуды ВП показал, что информативными показателями несовпадения тестовой и удерживаемой в памяти эталонной ориентаций являются изменения амплитуды как ранних компонентов ВП каудальных областей коры, связанных с сенсорной переработкой этого вида зрительной информации, так и среднелатентных когнитивных компонентов в передних ассоциативных отделах коры.
В проекционных зрительных областях коры несовпадение текущей и удерживаемой в памяти ориентаций сопровождалось увеличением амплитуды ранних компонентов ВП. Для компонентов Р100 и N150 ВП каудальных проекционных областей коры их увеличение не связано с тестовой ориентацией, но зависело от области регистрации и было наибольшим в затылочной коре. Этот факт соответствует роли затылочной области как первого коркового звена детектирования и ранней дискриминации основных ориентаций (Hubel, Wiesel, 1962). Таким образом, раннее обнаружение несовпадения двух ориентаций происходит в проекционной области, нейронные структуры которой настроены на этот признак, и в специфическом для этой операции временном интервале.
Чувствительность к несовпадению ориентаций также была обнаружена нами для амплитуды ранней позитивности Р100 в теменной и височной областях коры, т.е. при сличении текущей и удерживаемой в памяти информации в операции обнаружения несходства ориентаций принимают участие не только ранние зрительные области коры, но и более высокие уровни зрительной иерархии. Вопрос о роли теменной коры в обеспечении различных операций рабочей памяти обсуждался в литературе ранее (Bledowski et al., 2004; Бетелева, Синицын, 2008). В работе (Bledowski et al., 2004) авторы, комбинируя методы ВП и функциональной МРТ, показали, что нижнетеменная область играет важную роль в “хранении репрезентации в буфере” (“storage buffer operations”), необходимую для выполнения последующих операций рабочей памяти. В упомянутой работе особое внимание было обращено на факт временного перекрытия активации нижнетеменной и префронтальной областей, что, по мнению авторов, говорит о параллельности операций рабочей памяти в каудальных и ростральных областях коры. Анализ связанных с событиями потенциалов в работе (Бетелева, Синицын, 2008) показал роль теменной коры при сличении следа стимула с вновь поступившей информацией, хотя в этом исследовании значимые эффекты были обнаружены для сравнительно поздних этапов, после 300 мс, а эффекты совпадения и несовпадения стимулов авторы не анализировали.
Эффекты несовпадения текущей и удерживаемой в памяти ориентаций обнаружены нами и в височной области коры. Ее традиционно связывают с переработкой сложных зрительных стимулов (Allison et al., 1999). Вместе с тем, есть данные о наличии селективных ответов этой области коры на более простые зрительных признаки. Так, например, регистрация сигнала непосредственно с поверхности затылочно-височной коры мозга нейрохирургических пациентов показала существование в ее каудальных отделах ответов на такие простые изображения, как синусоидальные и радиальные решетки (Allison et al., 1999). При моделировании распределенных диполей ранних компонентов ВП в задаче дискриминации ориентаций зоны высокой активации были обнаружены не только в затылочной, но и в височной областях (Крылова и др., 2015). По данным исследования (Angelucci et al., 2002), височная кора активно влияет на переработку информации об ориентационных признаках на уровне V1 через систему нисходящих связей, и, оказывая связанные с контекстом модулирующие влияния, участвует в процессах интеграции информации о локальных и глобальных признаках. Важными являются сведения, представленные в обзоре по рабочей памяти (Christophel et al., 2017), о наличии в период удержания информации устойчивых стимул-специфических ответов нейронов нижневисочной коры (Miller et al., 1993; Chelazzi et al., 2001). Следует подчеркнуть, что в своей работе мы исследовали нейрофизиологические показатели на этапе сличения двух ориентаций, тогда как предметом большинства упоминаемых выше работ был предшествующий сличению этап удержания, для которого характерна высокая степень активности затылочной коры (Pasternak, Greenlee, 2005; Harrison, Tong 2009; Serences et al., 2009; Pratte, Tong, 2014; Ester et al., 2009, 2015). Следовательно, на более динамичном этапе сличения двух репрезентаций система активных мозговых образований, помимо ранних проекционных областей, должна включать более высокие корковые уровни, важные для выполнения последующих когнитивных операций. В этой системе височная кора может выполнять функцию звена, интегрирующего информацию, приходящую снизу от сенсорно-специфических областей и поступающую из расположенных выше передне-ассоциативных корковых зон (Bar et al., 2006).
Увеличение амплитуды ВП при несовпадении текущей и удерживаемой в кратковременной памяти ориентаций показаны также для следующего за Р100 негативного компонента N150 ВП затылочной коры. Согласно представлениям об иерархической организации переработки зрительного сенсорного сигнала (Hillyard, 2009), на этапе ранней селекции (80–140 мс) анализируются элементарные характеристики стимула, такие как ориентация, пространственная частота, контраст. Нейронные механизмы этой операции локализованы на уровне зрительных областей, что в нашей работе соответствует факту повышения амплитуды N150 затылочной области при несовпадении текущей и удерживаемой в памяти ориентаций.
В последние годы в работах по рабочей памяти на низкоуровневые зрительные характеристики, авторы расширяют круг структур, поддерживающих этот вид памяти (Curtis, D’Esposito, 2003; Serences et al., 2009; Christophel et al., 2017). Как пишут авторы обзора (Christophel et al., 2017), в литературе в течение долгого времени доминировало мнение о связи устойчивой активации фронто-париетальной системы с операцией кодирования абстрактных репрезентаций, отражающих такие характеристики объектов, как класс запоминаемого стимула, критерии принятия решения, характер ответа. Считалось, что абстрактные репрезентации, связанные с поведенческой задачей, через систему обратных нисходящих влияний координируют деятельность проекционных областей, где находится детальное описание запоминаемых стимулов (D’Esposito, Postle, 2015). Сходные предположения высказывали и другие авторы (Harrison,Tong, 2009), подчеркивая особую роль префронтальных областей в модуляции активности проекционных зон в соответствии с текущей задачей. Однако в последнее время появились данные о том, что в некоторых субрегионах префронтальной коры может кодироваться информация, связанная с конкретными физическими характеристиками стимула (Stokes et al., 2013; Raposo et al., 2014; Еster et al., 2015). В этом плане наиболее показательно исследование (Еster et al., 2015), в котором успешная реконструкция удерживаемой в памяти ориентации может быть выполнена по сигналам, зарегистрированным не только в первичных зрительных областях (V1-h, V4v/V3a) и заднетеменной коре (IPS0-3), но и в ряде субрегионов дорзолатеральной и вентролатеральной префронтальной коры. Авторы работы (Еster et al., 2015) приходят к выводу, что точные описания удерживаемых в памяти стимулов кодируются в распределенной сети сенсорных и фронто-париетальных областей, что существенно дополняет и обновляет более ранние модели рабочей памяти, постулирующие существование непересекающихся (“disjoint”) фронто-париетальных сетей нисходящего контроля и задних сенсорных сетей, сохраняющих точное описание стимула. При этом во фронто-париетальной системе детализированное описание стимула сочетается с информацией о целях, моторных программах, критериях принятия решения и других внесенсорных факторах, что позволяет гибко контролировать поведение с учетом изменяющихся требований среды.
В этом контексте интересен обнаруженный нами факт, что при несовпадении тестовой и удерживаемой в памяти ориентаций значимо увеличивается амплитуда негативности N240 во фронтальных областях коры. Данные об отражении несовпадения текущей и удерживаемой в памяти информации в параметрах фронтальной негативности можно найти в работах других авторов (Yin et al., 2011). Этот компонент, обозначаемый рядом авторов как N270, характеризуется устойчивым максимумом амплитуды в передних отделах коры, а его источник, по данным дипольного анализа, локализован в структурах префронтальной коры и передней цингулярной зоны (Yin et al., 2011). Он не является стимул-специфическим и регистрируется при конфликте текущей и хранящейся в рабочей памяти информации о форме (Zhang et al., 2001; Wang et al., 2001; Yin et al., 2011), цвете (Tian et al., 2001) или других свойствах объекта, например, характеристиках лицевого паттерна (Zhang et al., 2001). По мнению авторов этих работ, такая негативность отражает несоответствие между сенсорным входом и репрезентацией, хранящейся в памяти. В нашем исследовании роль префронтальных образований в мониторинге несовпадения двух ориентаций подтверждена данными моделирования дипольных источников компонента N240. Несовпадение ориентаций сопровождалось дополнительной активацией латерального, полюсного и орбитального отделов префронтальной коры, т.е. структур, которые участвуют в различных аспектах рабочей памяти – в активном удержании репрезентаций стимулов и дальнейшем манипулировании этой информацией, сличении новой и значимой информации, удержании внимания к релевантной информации и торможении нерелевантной (D’Esposito, 2007; Miller, D’Esposito, 2012; Мачинская, 2015). Следует отметить, что области, связанные с обнаружением несовпадения двух стимулов, активируются также в более простой задаче выделения целевого стимула (D’Esposito et al., 1998; Smith et al., 1995), что говорит об общности механизмов этих двух операций.
ЗАКЛЮЧЕНИЕ
Таким образом, полученные в нашей работе данные демонстрируют, что электрографические проявления несовпадения текущей и хранящейся в рабочей памяти информации об ориентации линий имеют обширную топографию с вовлечением проекционных зрительных, височных, теменных областей и префронтальной коры. При этом в зрительных областях события, связанные с распознаванием несовпадения двух последовательных стимулов, выявляются на более раннем интервале, соответствующем сенсорному этапу переработки сигнала, тогда как в префронтальной коре – позже, на этапе когнитивного анализа, в период развития негативности N240. Следовательно, в обнаружении несовпадения текущего сигнала и удерживаемой в памяти информации принимают участие различные нервные сети. Сети проекционных областей принимают участие в сенсорном анализе сигнала, его первичной сенсорной категоризации и, возможно, сопоставлении двух следующих друг за другом паттернов, также они хранят детализированное описание запоминаемых объектов. Сети передних отделов, или сети исполнительного контроля (“executive control”), обнаруживая несовпадение двух сенсорных сигналов, включают эту информацию в программу целенаправленного зрительного поведения. Полученные данные подчеркивают важность тесного взаимодействия фронтальных и каудальных областей для эффективности рабочей памяти (Kuo et al., 2011; Miller, D’Esposito, 2012). Оценка функционального взаимодействия фронтальных и проекционных областей, анализ нисходящих управляющих и модулирующих влияний, характер обратных связей – все это представляется важным и перспективным направлением будущих исследований.
Работа поддержана грантом РФФИ (Проект № 19-013-00918\19).
Список литературы
Бетелева Т.Г., Синицын С.В. Связанные с событиями потенциалы на разных этапах реализации зрительной рабочей памяти. Физиология человека. 2008. Т. 34. № 3. С. 5–15.
Горбачева И. Методы математической статистики. URL: http://medstatistic.ru/articles/kratkiy_kurs.pdf (дата обращения 01.09.2018).
Крылова М.А., Изъюров И.В., Герасименко Н.Ю., Чаянов Н.В., Михайлова Е.С. Моделирование источников компонентов зрительных вызванных потенциалов человека в задаче определения ориентации отрезков линий. Журн. высш. нервн. деят. 2015. Т. 65. № 6. С. 685–698. https://doi.org/0.7868/S0131164617030146
Мачинская Р.И. Управляющие системы мозга. Журн. высш. нервн. деят. 2015. Т. 65. № 1. С. 33–60. https://doi.org/10.7868/S0044467715010086
Allison T., Puce A., Spencer D.D., McGarthy G. Electrophysiological studies of human face perception. I: Potentials generated in occipitotemporal cortex by face and non-face stimuli. Cereb Cortex. 1999. V. 9. P. 415–430. https://doi.org/.https://doi.org/10.1093/cercor/9.5.415
Angelucci A., Levitt J.B., Walton E.J.S., Hupe J.-M., Bullier J., Lund J.S. Circuits for local and global signal integration in primary visual cortex. J. Neurosci. 2002. V. 22. P. 8633–8646. https://doi.org/.https://doi.org/10.1523/JNEUROSCI.22-19-08633.2002
Bar M., Kassam K.S., Ghuman A.S., Boshyan J., Schmid A.M., Dale A.M., Hamalainen M.S., Marinkovic K., Schacter D.L., Rosen B.R., Halgren E. Top-down facilitation of visual recognition. PNAS. 2006. V. 103. P. 449–454. https://doi.org/10.1073/pnas.0507062103
Bledowski C., Prvulovic C. D., Hoechstetter K., Scherg M., Wibral M., Goebel R., Linden D.E. Localizing P300 generators in visual target and distractor processing: a combined event-related potential and functional magnetic resonance imaging study. J. Neurosci. 2004. V. 24. P. 9353–9560.
Chelazzi L., Miller E.K., Duncan J., Desimone R. Responses of neurons in macaque area V4 during memory-guided visual search. Cereb. Cortex. 2001. V. 11. P. 761–772.
Christophel T.B., Klink P.C., Spitzer B., Roelfsema P.R., Haynes J.D. The distributed nature of working memory. Trends Cogn. Sci. 2017. V. 21. P. 111–124. https://doi.org/10.1016/j.tics.2016.12.007
Courtney S.M., Petit L., Haxby J., Ungerleider L.G. The role of prefrontal cortex in working.memory: examining the contents of consciousness. Phil.Trans. R. Soc. Lond. B. 1998. V. 353. P. 1819–1828.
Curtis C.E., D’Esposito M. Persistent activity in the prefrontal cortex during working memory. Trends. Cogn. Sci. 2003. V. 7. P. 415–423. https://doi.org/10.1016/S1364-6613(03)00197-9
D’Esposito M. From cognitive to neural models of working memory. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2007. V. 362. P. 761–772. https://doi.org/10.1098/rstb.2007.2086
D’Esposito M., Postle B.R. The cognitive neuroscience of working memory. Annu. Rev. Psychol. 2015. V. 66. P. 115–142. https://doi.org/10.1146/annurev-psych-010814-015031
D’Esposito M., Aguirre G.K., Zarahn E., Ballard D., Shin R.K., Lease J. Functional MRI studies of spatial and nonspatial working memory. Cogn. Brain Res. 1998. V. 7. P. 1–13.
Desikan R.S., Segonne F., Fischl B., Quinn B.T., Dickerson B.C., Blacker D., Buckner R.L., Dale A.M., Maguire R.P., Hyman B.T., Albert M.S., Killiany R.J. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. NeuroImage. 2006. V. 31. P. 968–980. http://dx.doi.org/.https://doi.org/10.1016/j.neuroimage.2006.01.021
Ester E.F., Serences J.T., Awh E. Spatially global representations in human primary visual cortex during working memory maintenance. J. Neurosci. 2009. V. 29. P. 15258–15265. https://doi.org/10.1523/JNEUROSCI.4388-09.2009
Ester E.F., Sprague T.C., Serences J.T. Parietal and frontal cortex encode stimulus-specific mnemonic representations during visual working memory. Neuron. 2015. V. 87. P. 1–13. https://doi.org/10.1016/j.neuron.2015.07.013
Harrison S.A., Tong F. Decoding reveals the contents of visual working memory in early visual areas. Nature. 2009. V. 458. P. 632–635. https://doi.org/10.1038/nature07832
Hillyard S.A. Event-related potentials (ERPs) and cognitive processing. Encyclopedia of Neuroscience. 2009. P. 13–18.
Hollingworth A., Richard A.M., Luck S.J. Understanding the function of visual short-term memory in human cognition: transsaccadic memory, object correspondence, and gaze correction. J. Exp. Psychol. Gen. 2008. V. 137. P. 163–181. https://doi.org/10.1037/0096-3445.137.1.163
Hubel D.H., Wiesel T.N. Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex. J. Physiol. 1962. V. 160. P. 106–154.
Hyun J.S., Woodman G.F., Vogel E.K., Hollingworth A., Luck S.J. The comparison of visual working memory representations with perceptual inputs. J. Exp. Psychol. Hum. Percept. Perform. 2009. V. 35. P. 1140–1160. https://doi.org/10.1037/a0015019
Kuo B.C., Yeh Y.Y., Chen A.J.W., D’Esposito M. Functional connectivity during top-down modulation of visual short-term memory representations. Neuropsychologia. 2011. V. 49. P. 1589–1596. https://doi.org/10.1016/j.neuropsychologia.2010.12.043
Luck S.J., Vogel E.K. The capacity of visual working memory for features and conjunctions. Nature. 1997. V. 390. P. 279–281.
Ma Y., Hua X., Wilson F. The egocentric spatial reference frame used in dorsal–lateral prefrontal working memory in primates. Neurosci. and Biobehav. Rev. 2011. V. 36. P. 26–33. https://doi.org/10.1016/j.neubiorev.2011.03.011
Magnussen S. Low-level memory processes in vision. Trends Neurosci. 2000. V. 23. P. 247–251.
Magnussen S. Implicit visual working memory. Scand. J. Psychol. 2009. V. 50. P. 535–542. https://doi.org/.https://doi.org/10.1111/j.1467-9450.2009.00783.x
Miller E.K., Li L., Desimone R. Activity of neurons in anterior inferior temporal cortex during a short- term memory task. J. Neurosci. 1993. V. 13. P. 1460–1478.
Miller B.T., D’Esposito M. Spatial and temporal dynamics of cortical networks engaged in memory encoding and retrieval. Front. Hum. Neurosci. 2012. V. 6 (109). P. 1–11. https://doi.org/10.3389/fnhum.2012.00109
Offen S., Schluppeck D., Heeger D.J. The role of early visual cortex in visual short-term memory and visual attention. Vision Res. 2009. V. 49. P. 1352–1362. https://doi.org/10.1016/j.visres.2007.12.022
Pasternak T., Greenlee M.W. Working memory in primate sensory systems. Nat. Rev. Neurosci. 2005. V. 6. P. 97–107. https://doi.org/10.1038/nrn1603
Pratte M.S., Tong F. Spatial specificity of working memory representations in the early visual cortex. Journal of Vision. 2014. V. 14. P. 1–12. https://doi.org/10.1167/14.3.22
Raposo D., Kaufman M.T., Churchland A.K. A category-free neural population supports evolving demands during decision-making. Nat. Neurosci. 2014. V. 17. P. 1784–1792.
Solomon S.G., Lennie P. The machinery of colour vision. Nat. Rev. Neurosci. 2007. V. 8. P. 276–286. https://doi.org/10.1038/nrn2094
Schacter D.L., Chiu C.-Y. P., Ochsner K.N. Implicit memory: a selective review. Annu. Rev. Neurosci. 1993. V. 16. P. 159–182. https://doi.org/10.1146/annurev.ne.16.030193.001111.
Serences J.T., Ester E.F., Vogel E.K., Awh E. Stimulus-specific delay activity in human primary visual cortex. Psychol. Sci. 2009. V. 20. P. 207–214. https://doi.org/10.1111/j.1467-9280.2009.02276.x
Smith E.E., Jonides J., Koeppe R.A., Awh E., Schu-macher E.H., Minoshima S. Spatial versus object working memory: PET investigations. J. Cog. Neurosci. 1995. V. 7. P. 337–356. https://doi.org/10.1162/jocn.1995.7.3.337
Stokes M.G., Kusunoki M., Sigala N., Nili H., Gaffan D., Duncan J. Dynamic coding for cognitive control in prefrontal cortex. Neuron. 2013. V. 78. P. 364–375. https://doi.org/10.1016/j.neuron.2013.01.039
Tian S., Wang Y., Wang H., Cui L. Interstimulus interval effect on event-related potential N270 in a color matching task. Clin. Electroencephalogr. 2001. V. 32. P. 82–86. https://doi.org/.https://doi.org/10.1177/155005940103200207
Wang H., Wang Y., Kong J., Cui L., Tian S. Enhancement of conflict processing activity in human brain under task relevant condition. Neurosci. Lett. 2001. V. 298. P. 155–158.
Xu X., Collins C.E., Khaytin I., Kaas J.H., Casagrande V.A. Unequal representation of cardinal vs. oblique orientations in the middle temporal visual area. 2006. PNAS. V. 103. P. 17490–17495. https://doi.org/.https://doi.org/10.1073/pnas.0608502103
Yang L.C., Li M.H., Wilson F.A., Hu X.T., Ma Y.Y. Prefrontal attention and multiple reference frames during working memory in primates. Chin Sci Bull. 2013. V. 58. P. 449–455. https://doi.org/10.1007/s11434-012-5462-y
Yin J., Gao Z., Jin X., Ye L., Shen M., Shui R. Tracking the mismatch information in visual short term memory: An event-related potential study. Neurosci. Lett. 2011. V. 491. P. 26–30. https://doi.org/10.1016/j.neulet.2011.01.001
Zhang X., Wang Y., Li S., Wang L. Event-related potential N270, a negative component to identification of conflicting information following memory retrieval. Clin. Neurophysiol. 2003. V. 114. P. 2461–2468.
Дополнительные материалы отсутствуют.
Инструменты
Сенсорные системы