

Сенсорные системы, 2019, T. 33, № 4, стр. 351-354
Оценка метода инактивации церкальных сенсилл сверчка при помощи электроцеркограммы
А. М. Луничкин 1, Ю. А. Баулин 1, 2, М. И. Жуковская 1, А. Н. Князев 1, *
1 Институт эволюционной физиологии и биохимии им. И.М. Сеченова РАН
194223 г. Санкт-Петербург, пр. Тореза, 44, Россия
2 Санкт-Петербургский национальный исследовательский университет информационных технологий,
механики и оптики
197101 г. Санкт-Петербург, Кронверкский пр., 49, Россия
* E-mail: ank50@mail.ru
Поступила в редакцию 20.04.2019
После доработки 28.05.2019
Принята к публикации 14.06.2019
Аннотация
Предложен новый метод регистрации электрической активности изолированного церка – электроцеркограммы. Ответ на стимуляцию толчком воздуха состоит из быстрого негативного on-ответа, затем переходящего в плато, и позитивного off-ответа на прекращение потока. Инактивация церка глицерином, применявшаяся ранее только в поведенческих экспериментах, приводит к уменьшению амплитуды on- и off-ответов на дистантные стимулы в 2–3 раза. Величина плато падала ниже уровня шума для большинства препаратов.
Церки сверчков представляют собой парные конические выросты конца брюшка, на каждом из которых расположено от 50 до нескольких тысяч сенсилл в зависимости от видовой принадлежности и возраста насекомого (Edwards, Palka, 1974; Князев, Попов, 1981). Это щетинковидные волосковые сенсиллы, механорецепторные нитевидные волосковые сенсиллы (трихоботрии), колоколовидные сенсиллы и гравирецепторы – булавовидные сенсиллы (рис. 1, а). Церкальный аппарат детектирует акустические сигналы ближнего поля и направление воздушных потоков, обеспечивая обнаружение хищников, запуск реакции направленного бегства, внутривидовую коммуникацию, сенсорно-моторную регуляцию собственной локомоторной активности через восприятие потоков воздуха, создаваемых насекомым при ходьбе, прыжках или полете (Князев, Попов, 1981; Рожкова, 1993; 1994). Основным условием работы ветрочувствительных сенсилл является возможность свободного колебания кутикулярного волоска под действием потока воздуха. В этологических экспериментах для инактивации их покрывают слоем желеобразного глицерина, препятствующего колебаниям трихоботрий, который впоследствии счищается сверчком в процессе груминга. Изменение поведенческих ответов сверчков Gryllus locorojo Weissman & Gray n. и Phaeophilacris bredoides Kaltenbach, 1986 на акустическую стимуляцию, а также полное возвращение чувствительности после груминга церков свидетельствует об эффективности метода, описанного в работах (Жемчужников, Князев, 2011; Луничкин, Князев, 2017). Прямых физиологических доказательств прекращения или существенного снижения чувствительности периферического сенсорного аппарата до настоящего времени не получено. Общий дискомфорт, вызванный процедурой инактивации, может прерывать поведенческую реакцию на уровне центральной нервной системы, поэтому оценка электрофизиологических периферических ответов церка необходима для дальнейшего использования этого метода в экспериментальной работе.
Рис. 1.
Морфофизиологические характеристики церка сверчка Ph. bredoides.
а – Типы сенсилл, представленные на церке сверчка Ph. Bredoides ( сканирующая электронная микроскопия, микроскоп FEI – Quanta 250, напыление платиной): 1 – булавовидная сенсилла; 2 – волосковидная сенсилла (трихоботрия); 3 – щетинковидная сенсилла; б – схема электроцеркограммы; в – оригинальная запись ЭЦГ в контроле и после инактивации. Пунктирная линия – исходные данные, сплошная – данные, сглаженные по пяти точкам; г – параметры ЭЦГ ответов на толчок воздуха. Звездочками обозначены достоверные различия между группами “контроль” и “инактивация”: ** – p < 0.01; *** – p < 0.001
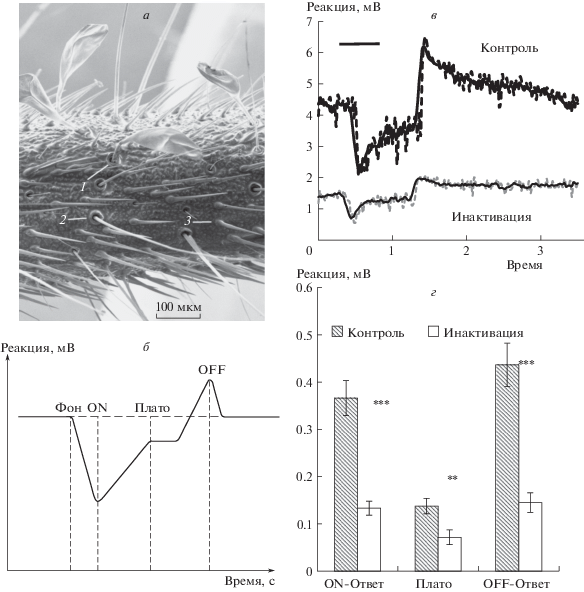
Поскольку общий план строения и сенсорного оснащения церков сходен со строением антенны насекомого – хемо-механосенсорного парного придатка головы, то для оценки метода инактивации церкальных сенсилл был выбран метод отведения суммарного электрического ответа на стимуляцию – электроцеркограммы (ЭЦГ), аналогичный электроантеннограмме (Жуковская и др., 2017). Этот метод отличается простотой приготовления препарата и возможностью исследования ответов на широкий спектр сенсорных стимулов. Основной вклад в генерацию ответа вносит рецепторный потенциал – первичная электрическая реакция сенсорных клеток, при этом величина ответа зависит как от интенсивности стимула, так и от количества реагирующих сенсилл на участке, от которого регистрируется ответ (Kapitskii, Gribakin, 1992). Африканские пещерные сверчки обладают длинными (16 мм) церками, изготовление препарата из них не занимает много времени, что крайне важно для сохранения высокой чувствительности в течение эксперимента. Сверчков содержали при температуре 26°C и влажности воздуха 60%. Световой режим – 12:12 (свет: темнота) создавали при помощи светодиодного осветителя AquaLighter 3 (Collar, Украина). Вода и пища постоянно присутствовали.
Для регистрации ЭЦГ церок отрезали вблизи основания и помещали в держатель. Хлорсеребряные электроды контактировали с дистальным и проксимальным концами церка с использованием смоченных в растворе Рингера полосок фильтровальной бумаги так, чтобы все стимулируемые участки находились между контактами. Места контактов изолировали от центральной части церка при помощи валиков пластилина. Предусилитель с входным сопротивлением 100 МОм и коэффициентом усиления Кус = 10 находился внутри экранированной камеры; его выходной сигнал подавался на вход усилителя Intracellular Electrometer Model 3100 (A-M Systems, США), Кус = 10, низкочастотный фильтр 5 кГц, а затем оцифровывался при помощи осциллографа PPS 10 (Velleman Instruments, США) и сохранялся в виде файлов на компьютере. Стимул представлял собой поток воздуха со скоростью 1.3 м/с, который подавался из трубки диаметром 2 мм под углом около 15˚ к оси церка, по направлению к его основанию, длительностью 1 с. ЭЦГ регистрировали в течение 5 с; интервалы между предъявлениями стимула составляли 30 с. После регистрации пяти ЭЦГ (контроль) церок покрывали глицерином и снова регистрировали пять ЭЦГ (опыт). Использовано 20 церков (10 левых и 10 правых) от 12 взрослых самцов возрастом 14–21 сутки имагинального онтогенеза, взятых из лабораторной культуры. Сопротивление препарата измеряли при помощи цифрового мультимера DT-182 непосредственно перед отведениями; для контрольных и инактивированных церков оно статистически не различалось и составляло 0.8 ± ± 0.2 и 0.9 ± 0.1 МОм соответственно. Данные обрабатывали в программе MS Excel. Каждую запись сглаживали с применением метода скользящего среднего по пяти точкам. “Фон” определяли как среднее от шести последних точек перед подачей стимула. On-ответ электроцеркограммы определяли как разницу между “фоном” и минимальным значением напряжения за время действия стимула. “Плато” находили как разницу между “фоном” и средним от шести последних точек перед выключением стимула. Величину off-ответа считали как разницу между максимальным показателем в течение 0.5 с после выключения стимула и средним от шести последних точек перед выключением стимула (рис. 1, б). Уровень шума рассчитывали как среднеквадратическое отклонение для значений напряжения последних 3 с записи. Абсолютное значение параметра усредняли по пяти повторностям эксперимента и представляли как среднее ± стандартная ошибка. Данные проверяли на нормальность распределения при помощи теста Колмогорова-Смирнова. Достоверность различий (р < 0.05) оценивали с помощью t-критерия Стьюдента.
ЭЦГ на толчок воздуха (рис. 1, б, в) состоял из негативного колебания потенциала – on-ответа, переходящего в плато, и ответа на выключение – положительного off-ответа. Все величины, измеренные для правых и левых церков, статистически не различались, поэтому данные всех 20 церков были объединены. После инактивации все параметры ответа уменьшались в 2–3 раза (рис. 1, в, г). Уровень шума достоверно снижался, однако это снижение составляло около 30%, что связано с уменьшением подвижности кутикулярных отделов сенсилл, оставшиеся 70% включают собственный шум регистрирующей аппаратуры и спонтанную активность церкальных нейронов.
Ответы инактивированного церка можно было визуально выделить из шума только для шести из 20 препаратов. Раздельная статистическая обработка данных для групп, где ответ после нанесения глицерина был виден (1) и не виден (2), показала, что величина on-ответа статистически не различалась для обеих групп до инактивации (0.43 ± 0.08 и 0.34 ± 0.04 мВ, p > 0.05), однако после нанесения глицерина различия оказались достоверными (0.20 ± 0.03 и 0.11 ± 0.01 мВ, р < 0.01). Величина плато после нанесения глицерина падала до уровня шума, измеренного до нанесения глицерина (рис. 1, г). Плато ответов не различалось для групп 1 и 2 как до (0.14 ± 0.03 и 0.14 ± 0.02, p > 0.05), так и после инактивации (0.07 ± 0.03 и 0.07 ± 0.02, p > 0.05). Оff-ответ снижался после инактивации сходно с on-ответом (рис. 1, г). Величина off–ответа статистически не различалась для групп 1 и 2 до инактивации (0.56 ± 0.10 и 0.39 ± 0.05 мВ, p > 0.05), однако после нанесения глицерина различия оказались достоверными (0.24 ± 0.05 и 0.11 ± 0.01 мВ, р = 0.01). Разделенные выборки показали, что исходная чувствительность церков была одинаковой, однако после нанесения глицерина большая их часть снижала свои on– и off–ответы примерно до уровня шума, а меньшая, ответы которых были визуально различимы, снижали ответы, но в меньшей степени. По-видимому, в тех случаях, когда ответ оставался, часть сенсилл не была покрыта глицерином и продолжала отвечать на стимул.
Таким образом, предложенный нами метод отведения ЭЦГ отражает суммарный ответ церкальных ветрочувствительных нейронов в ответ на адекватную стимуляцию. Инактивация церков глицерином достоверно уменьшает регистрируемые ответы на дистантные механические стимулы, делая их неразличимыми на уровне шума в большинстве случаев. Мы считаем, что после минимальной доработки процедуры нанесения глицерина инактивация органа будет практически полной. В дальнейшем метод ЭЦГ можно будет использовать на интактных сверчках, что позволит длительную многократную регистрацию электрической активности от одного животного в ответ на варьирующие по модальности и интенсивности стимулы.
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
Все применимые международные, национальные и/или институциональные принципы ухода и использования животных были соблюдены.
Настоящая статья не содержит результатов каких-либо исследований с участием людей в качестве объектов исследований.
Работа выполнена за счет средств госбюджета, программа АААА-А18-118013090245-6.
Список литературы
Жемчужников М.К., Князев А.Н. Изменение характера направленных двигательных реакций самок сверчков Gryllus argentinus Sauss. на внутривидовые сигналы в условииях сенсорной патологии на разных этапах имагинального онтогенеза. Ж. эвол. биохим. и физиол. 2011. Т. 47. № 6. С. 485–490.
Жуковская М.И., Селицкая О.Г., Фролов А.Н., Щеникова А.В., Грушевая И.В., Малыш Ю.М., Берим М.Н., Трепашко Л.И. Межпопуляционная изменчивость ответов самцов кукурузного мотылька Ostrinia nubilalis Hbn. (Lepidoptera: Crambidae) на феромонные композиции: анализ электроантеннограмм. Ж. эвол. биохим. и физиол. 2017. Т. 53. № 4. С. 308–310.
Князев А.Н., Попов А.В. Функциональная организация церкальной механорецепторной системы сверчка Gryllus bimaculatus. Ж. эвол. биохим. и физиол. 1981. Т. 17. № 5. С. 503–511.
Луничкин А.М., Князев А.Н. Участие структур механосенсорного комплекса сверчка Phaeophilacris bredoides Kaltenbach (Orthoptera, Gryllidae) в запуске двигательных ответов на звук. Ж. эвол. биохим. и физиол. 2017. Т. 53. №. 6. С. 425–436.
Рожкова Г.И. Нейрофизиология церкальной системы насекомых. М.: Наука, 1993. 175 с.
Рожкова Г.И. Переработка информации о направлении в церкальной системе насекомых. Сенсорные системы. 1994. Т. 8. № 3-4. С. 184–199.
Edwards J.S., Palka J. The cerci and abdominal giant fibres of the house cricket, Acheta domesticus. I. Anatomy and physiology of normal adults. Proceedings of the Royal Society of London. Series B. Biological Sciences. 1974. V. 185. № 1078. P. 83–103.
Kapitskii S.V., Gribakin F.G. Electroantennogram of the American cockroach: effect of oxygen and an electrical model. J. Comp. Physiol. A. 1992. V. 170. P. 651–663.
Дополнительные материалы отсутствуют.
Инструменты
Сенсорные системы