

Сенсорные системы, 2020, T. 34, № 1, стр. 12-14
Дизруптивные сигналы – эффективный механизм прерывания нежелательной копуляции у Pentatoma rufipes
Л. С. Шестаков *
Федеральное государственное бюджетное учреждение науки
Институт проблем передачи информации им. А.А. Харкевича РАН
127051 Москва, Большой Каретный пер. 19, Россия
* E-mail: zicrona@yandex.ru
Поступила в редакцию 09.10.2019
После доработки 25.10.2019
Принята к публикации 06.11.2019
Аннотация
Акустический репертуар Pentatoma rufipes, помимо призывных, конкурентных и сигналов ухаживания, включает и особый тип сигнала – дизруптивный. Сигнал служит для предотвращения попыток копуляции и разрушения уже копулирующих пар. Данный тип сигнала издают только самки при попытке самца копулировать. Показано, что сигнал не только препятствует копуляции, но и разрушает уже сформировавшиеся пары.
ВВЕДЕНИЕ
Вибрационную коммуникацию используют представители практически всех групп животных – начиная от нематод и заканчивая такими крупными позвоночными, как слоны. Однако, несмотря на широкий интерес к изучению вибрационной коммуникации насекомых, на данный момент вибрационные сигналы известны всего для 39 из более чем 4000 видов Pentatomidae (Laumann, 2013). Это связано с тем, что подавляющая часть исследований проводилась и проводится на ограниченном числе модельных объектов (Cokl, 2008; de Groot et al., 2010; Cocroft et al., 2010; McBrien, Millar, 2003; Malek et al., 2018).
В основном интерес к вибрационной коммуникации насекомых обусловлен их значимостью как вредителей сельскохозяйственных культур и переносчиков заболеваний растений (Panizzi et al., 2000; Cokl et al., 2014; Malek et al., 2018). Вибрационные сигналы играют важную роль в коммуникации полужесткокрылых и являются одним из необходимых элементов ритуала ухаживания (Cokl et al., 2014). При этом различия в амплитудно-временных и частотных характеристиках сигналов могут быть одним из механизмов эффективной межвидовой изоляции для близких видов, чьи феромоны имеют близкую структуру.
Сигналы протеста или дизруптивные сигналы, служащие для разрушения копулирующих пар, встречаются у насекомых, использующих акустическую коммуникацию. Интересно, что подобные сигналы используют не только нерецептивные самки, но иногда и самцы. Так, у некоторых видов цикадок самец при обнаружении копулирующей пары издает специальный сигнал, который прерывает копуляцию (Nieri, Mazzoni, 2018; Malek et al., 2018). Однако специфический сигнал протеста, препятствующий копуляции, был обнаружен только у одного вида полужесткокрылых – Pentatoma rufipes (рис. 1.), хотя у других видов самки могут совершать похожие движения брюшком, но сигналы при этом не регистрируются (Shestakov, 2015).
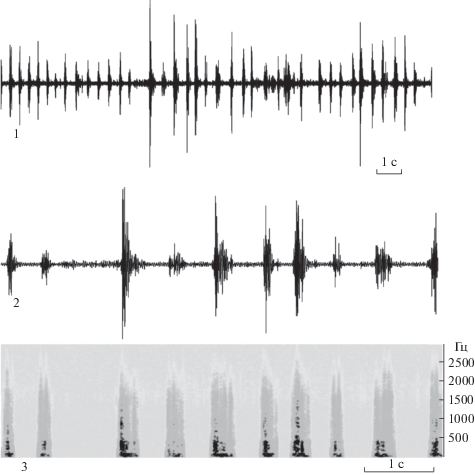
Рис. 1.
Осциллограммы (1–2) и сонограмма (3) дизруптивного сигнала Pentatoma rufipes. Сонограмма дана для осциллограммы 2.
Актуальность изучения коммуникации клопов-щитников обусловлена тем, что многие Pentatomidae являются экономически значимыми вредителями и переносчиками заболеваний растений. Например, карантинный для стран Евразийского экономического союза клоп Halyomorpha halys, который отмечен на южных границах России. В связи с этим исследование их коммуникации представляет значительный интерес не только для расширения представлений об их этологии и выявления новых таксономических признаков, но и для разработки методов контроля численности без использования инсектицидов. Так, на некоторых видах цикадок было показано, что использование искусственных стимулов, созданных на основе дизруптивных сигналов, препятствует эффективной коммуникации и достоверно снижает численность насекомых на растении (Nieri, Mazzoni, 2018; Malek et al., 2018).
МАТЕРИАЛ И МЕТОДИКА
Исходный живой материал Pentatoma rufipes для работы был собран в полевых условиях в Московской, Тульской областях (2013–2015 г., 2019 г.), в Республике Алтай (2015 г.) В лаборатории насекомых содержали в садках 35 × 35 × 45 см при постоянном наличии свежего кормового растения и 12/12 часовом световом цикле. Постоянный световой цикл необходим для исключения влияния изменений фотопериода на активность насекомых. Температура содержания 20–25°С.
Для поведенческих экспериментов с модельными стимулами были использованы только насекомые, выведенные в лаборатории из личинок. Самцы и самки до имагинальной линьки содержались раздельно. Такое разделение необходимо, так как опыт предыдущей копуляции может оказывать влияние на реакцию на тестовый стимул, поэтому в экспериментах должны использоваться только заведомо девственные самцы и самки.
Для записи сигналов в полевых условиях и лаборатории применяли лазерный виброметр PDV 100 (Polytec, Германия) и пьезоэлектрический адаптер ГЗП 311 и ГЗП 661. Частота дискретизации сигнала 44.1 кГц. Такая частота более чем достаточна, так как сигналы полужесткокрылых практически всегда низкочастотные (доминантные частоты около 200 Гц, крайне редко 1–4 кГц). Для генерации стимулов использовали вибростенд 4810, (Bruel & Kjaer). Всего было протестировано 20 пар на стадии начала копуляции и 15 пар в процессе копуляции.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Были записаны дизруптивные сигналы 18 самок. Данный сигнал производится самками, когда самец пытается приступить к копуляции – начинает ощупывать самку антеннами и ставит передние конечности на конец ее брюшка. В этот момент нерецептивная самка скользящими движениями ударяет боками брюшка о субстрат и по самцу. Основные параметры сигнала представлены в табл. 1. Следует заметить, что по нашим наблюдениям длительность сигнала зависит от того, как быстро самец прекратит попытки копуляции. Как только самец отходит от самки эмиссия сигнала сразу же прекращается. В других обстоятельствах, по нашим наблюдениям, этот тип сигнала не издается.
Таблица 1.
Основные параметры дизруптивного сигнала P. rufipes
| Параметр сигнала | Длительность пульса, с | Длительность серии, с | Число пульсов в серии | Период повторения пульсов, мс |
|---|---|---|---|---|
| Среднее значение | 0.138 | 13.7 | 26.8 | 0.212 |
| Стандартное отклонение | 0.15 | 7.3 | 19.2 | 0.12 |
При предъявлении дизруптивного сигнала только формирующимся парам и парам, находящимся в процессе копуляции, во всех случаях происходило разрушение пар. При этом насекомые стремились покинуть растение. Достоверные различия обнаружены лишь в латентных периодах от начала предъявления стимула до момента разрушения пары (двухсторонний точный критерий Фишера; p < 0.05). Латентный период до разрушения пары в начале ухаживания составил 1.687 ± 0.691 с (N = 20). Латентный период у пар, вступивших в копуляцию, 7.012 ± 2.976 с (N = 15).
Таким образом, можно с уверенностью сказать что дизруптивный сигнал самки является эффективным механизмом, препятствующим успешной встрече полов у P.rufipes. Мы предполагаем, что использование дизруптивных сигналов полужесткокрылых может стать новым эффективным механизмом контроля численности. Это подтверждается данными, полученными на некоторых видах цикадок – вредителей винограда. Трансляция дизруптивных сигналов на кормовое растение нарушает нормальный процесс коммуникации, что в свою очередь приводит к невозможности формирования пар и снижению численности популяции.
Работа поддержана РФФИ (грант № 20-04-00553 а).
Список литературы отсутствует.
Дополнительные материалы отсутствуют.
Инструменты
Сенсорные системы