

Сенсорные системы, 2022, T. 36, № 3, стр. 244-251
Интерауральная последовательная маскировка в слуховой системе дельфина
В. В. Попов 1, Д. И. Нечаев 1, А. Я. Супин 1, *, Е. В. Сысуева 1
1 Институт проблем экологии и эволюции РАН
119071 Москва, Ленинский просп., 33, Россия
* E-mail: alex_supin@mail.ru
Поступила в редакцию 17.03.2022
После доработки 06.04.2022
Принята к публикации 29.04.2022
- EDN: QQOUON
- DOI: 10.31857/S0235009222030064
Аннотация
У бутылконосого дельфина Tursiops truncatus измеряли время освобождения коротколатентных слуховых вызванных потенциалов (КСВП) от последовательной маскировки при контактной звуковой стимуляции короткими тональными посылками: при положении источников кондиционирующего и тестирующего сигналов с одной стороны головы (унилатеральная стимуляция) и при положении источников с разных сторон головы (билатеральная стимуляция). В обоих вариантах стимуляции наблюдали значительное подавление тестирующего КСВП при межстимульных задержках менее 1 мс и практически полное восстановление тестирующего ответа (освобождение от маскировки) при задержках около 5 мс. Предполагается, что эффекты последовательной маскировки развиваются преимущественно в бинауральных слуховых структурах, где афферентные потоки от обеих улиток взаимодействуют.
ВВЕДЕНИЕ
Для зубатых китообразных (зубатых китов, дельфинов и морских свиней) характерны уникальные возможности слуховой системы. Обзоры свойств слуха китообразных приведены в нескольких монографиях (Au, 1993; Supin et al., 2001; Au, Hastings, 2008). В этих обзорах отмечаются высокая чувствительность, широкий частотный диапазон, высокое быстродействие, высокая пространственная избирательность слуховой системы. Однако некоторые аспекты работы слуховой системы китообразных по-прежнему представляют интерес для более детального изучения, поскольку касаются фундаментальных механизмов слухового анализа. К таким аспектам, в частности, относится взаимодействие между временным и пространственным анализами.
Значительная часть сведений о пространственной избирательности слуховой системы дельфинов получена методом регистрации коротколатентных слуховых вызванных потенциалов (КСВП). Ввиду сильного развития слуховой системы у дельфинов форма слуховых вызванных потенциалов может быть зарегистрирована неинвазивно, путем наложения регистрирующих электродов на поверхность головы животного. Это свойство делает КСВП приемлемым для исследования широкого круга слуховых функций у китов и дельфинов. В том числе КСВП широко применялись для исследования пространственной избирательности и временной разрешающей способности.
Методом КСВП для дельфинов была показана значительная острота приемного луча, как бинаурального, создаваемого совместной работой обоих ушей (Popov, Supin, 1988, 2009; Popov et al., 1992; Kastelein et al., 2005), так и монаурального для каждого уха в отдельности (Popov, Supin, 1991; Popov et al., 2006). Помимо высокой остроты луча приема показана значительная интерауральная разница интенсивности принимаемых сигналов, если их источник не находится на продольной оси головы (Popov, Supin, 1991; Popov et al., 2006; Mulsow et al., 2014). Данные о высокой пространственной избирательности, полученные методом КСВП, дополняются поведенческими данными: показаны острая направленность луча приема (Zaitseva et al., 1975; Au, Moore, 1984) и низкие пороги минимально различаемого угла между источниками звука (minimum audible angle) (Renaud, Popper, 1975; Dubrovskiy, 1990).
Уникальное быстродействие слуховой системы дельфинов также демонстрировалось разными методами. Демонстративные данные, касающиеся быстродействия слуховой системы, были получены при регистрации КСВП, вызванных короткими парными звуковыми импульсами (Popov, Supin, 1990; Supin, Popov, 1995, 2015; Popov et l., 2001, 2020; Supin et al., 2007). Измерение быстродействия этим методом основано на том, что при коротких межимпульсных интервалах ответ на второй стимул угнетен (вариант прямой последовательной маскировки), однако восстанавливается при удлинении интервала (эффект освобождения от маскировки). Чем выше быстродействие слуховой системы, тем быстрее происходит освобождение от маскировки. Регистрация КСВП показала, что при одинаковом уровне двух импульсов в паре и одинаковых спектральных характеристиках этих импульсов восстановление второго ответа происходит очень быстро: при межимпульсных интервалах от 1 до 5 мс, в зависимости от уровня, одинакового для обоих импульсов. Если уровень первого импульса выше, чем уровень второго, то восстановление может потребовать больше времени (Popov, Supin, 1990; Supin, Popov, 2015); если спектральные характеристики двух импульсов различаются, то восстановление второго ответа происходит еще быстрее (Popov et al., 2001).
Однако раздельное исследование пространственной избирательности и быстродействия слуховой системы не дает полного представления о ее работе в естественных условиях. Сложные акустические сцены, как правило, характеризуются разными временными соотношениями между сигналами, исходящими от разных источников, занимающих в пространстве разное положение относительно животного. Поэтому способность выделять определенный звуковой сигнал из всего многообразия звуков, характеризующего естественную звуковую сцену, зависит не только от пространственной избирательности или быстродействия, но и от взаимодействия между этими двумя характеристиками слуховой системы, т.е. от способности раздельно воспринимать сигналы, приходящие в разное время и с разных направлений. Учитывая уникальные способности слуховой системы дельфинов, такая способность и механизмы ее реализации значимы для понимания фундаментальных механизмов слуха.
К настоящему времени анализ пространственно-временных взаимоотношений слуховой системой дельфинов исследован в минимальной степени. Предметом исследований были либо временное разрешение при одинаковом (чаще всего, на продольной оси головы) положении источников последовательных сигналов, либо пространственная избирательность слуха при неизменном временном соотношении сигналов. В данном исследовании предпринята попытка проанализировать временное и пространственное разрешение слуха дельфина не порознь, а во взаимодействии. Для этого методом парных стимулов в сочетании с регистрацией КСВП измеряли время восстановления ответа на второй стимул при различном положении источников двух стимулов. Для получения возможно более контрастной картины использовали источники стимулов, расположенные по разные стороны от медиальной плоскости головы: один – справа, другой – слева. Такая схема стимуляции предполагала исследование взаимодействия между сигналами, один из которых поступает преимущественно в правое, а другой – в левое ухо.
Осуществление монауральной стимуляции у дельфинов в свободном поле затруднено из-за специфической организации их звукопроводящих путей. У дельфинов основной путь проведения звуков из внешней среды к слуховой улитке обеспечивается жировым тяжем, расположенным в нижней челюсти (мандибулярная гипотеза, Norris, 1968, 1969; Norris, Harvey, 1974). Звук попадает в этот звукопроводящий канал через “звуковое окно” – тонкую костную стенку на наружной поверхности нижней челюсти (мандибулы). Мандибулярная гипотеза была подтверждена измерением порогов потенциалов действия слухового нерва (McCormick et al., 1970; 1980) и слуховых вызванных потенциалов ствола мозга (Bullock et al., 1968). Однако более поздние исследования показали, что местом вхождения звука в мандибулярный проводящий канал может быть не только латеральная, но и медиальная поверхность нижней челюсти (Cranford et al., 2008; Popov et al., 2016). Поэтому при латеральном положении источника звук может передаваться к обеим (правой и левой) слуховым улиткам; например, при правостороннем расположении источника справа звук может достигать улитки как через латеральную поверхность правой половины нижней челюсти, так и через медиальную поверхность левой половины; при левостороннем расположении – наоборот.
Чтобы исключить эту неопределенность, в данном исследовании мы использовали контактную звуковую стимуляцию, при которой источник звука непосредственно контактировал с звуковым окном на латеральной поверхности нижней челюсти, в зоне максимальной чувствительности к звуковым сигналам. Возможность прохождения звука через звуковое окно контралатеральной стороны при этом полностью не исключалась, но обеспечивался намного больший уровень звука на стороне расположения излучателя, чем на контралатеральной стороне.
МЕТОДИКА
Объект и условия эксперимента. Объектом исследования была взрослая (28 лет) самка бутылконосого дельфина (афалины) Tursiops truncatus. До использования в данном исследовании животное в течение 25 лет находилось в коммерческом дельфинарии. Во время исследования животное содержали на Утришской морской станции Российской академии наук (Краснодарский край, Новороссийский район) в бассейне, наполненном морской водой (диаметр 6 м, глубина 1.7 м). По окончании серии экспериментов животное было возвращено в коммерческий дельфинарий.
На время эксперимента уровень воды в бассейне понижали до 35 см. Животное укладывали на носилки, выполненные из звукопрозрачного материала (рыболовная сеть). Носилки с животным устанавливали на опоры; при низком уровне воды в бассейне спина животного и дорзальная поверхность головы с дыхалом оставались над поверхностью воды. Тем самым обеспечивались беспрепятственное дыхание и надводное положение участков для установки регистрирующих электродов.
Эксперименты проводили ежедневно, длительность эксперимента составляла 2–3 ч, после чего животное освобождали из носилок и уровень воды в бассейне восстанавливали до 1.7 м.
Регистрация КСВП. Для регистрации КСВП на поверхности тела устанавливали два электрода: активный и референтный. Электроды удерживались присосками. Активный электрод устанавливали на дорзальной поверхности головы, по средней линии, 7–8 см каудальнее дыхала; референтный электрод – на спинном плавнике; оба электрода над поверхностью воды. Заземляющий электрод находился в воде, в которую было погружено тело животного.
Электроды для регистрации биопотенциалов были соединены экранированными кабелями со входом усилителя биопотенциалов LP511 (Grass Technologies, Warwick, США). Усилитель обеспечивал усиление 80 дБ в диапазоне частот 100–3000 Гц. Усиленные биопотенциалы подавали на вход 16-битного аналого-цифрового преобразователя, который входил в состав платы сбора данных NI DAQ-6251 (National Instruments, Austin, США). Частота стробирования для оцифровки составляла 40 кГц. Дальнейшую обработку сигналов, переведенных в цифровую форму, выполняли при помощи стандартного персонального компьютера.
Звуковая стимуляция. Стимулами служили тональные посылки с несущей частотой 64 кГц. Как показали многочисленные предшествующие исследования (Supin et al., 2001), эта частота лежит в пределах диапазона высокой слуховой чувствительности у бутылконосых дельфинов. Несущая частота была модулирована огибающей в форме одного периода смещенной косинусоиды (рис. 1, а). Период огибающей частоты был таким, что посылка содержала шесть циклов несущей. Спектр такой посылки имеет пик на несущей частоте 64 кГц и эквивалентную (по мощности) прямоугольную ширину ±8 кГц относительно центральной частоты (б).
Рис. 1.
Характеристики звуковых стимулов.
а – форма сигнала, б – спектр сигнала; вертикальные пунктирные линии обозначают эквивалентную прямоугольную ширину.

Стимулы синтезировали в цифровой форме с использованием стандартного персонального компьютера и программы собственной разработки, составленной в программной оболочке LabVIEW (National Instruments). Частота стробирования составляла 512 кГц (восемь точек на один цикл несущей частоты), что не только соответствовало пределу Найквиста, но и позволяло с хорошей точностью генерировать сигналы с несущей частотой 64 кГц. Программа обеспечивала синтез двух стимулов (тональных посылок) с независимыми характеристиками и с регулируемой задержкой один (тестирующий) относительно другого (кондиционирующего). Тестирующий и кондиционирующий стимулы могли подаваться либо через один и тот же излучатель (унилатеральная стимуляция), либо через разные излучатели, один из которых контактировал с правой, а другой – с левой стороной головы (билатеральная стимуляция).
Стимулы, синтезированные в цифровой форме, преобразовывали в аналоговую форму при помощи двухканального 16-битного цифроаналогового преобразователя, входящего в состав той же платы сбора данных, которая выполняла регистрацию биопотенциалов.
Стимулы, преобразованные в аналоговую форму, усиливали по мощности усилителями собственной разработки (полоса воспроизводимых частот 0.1–200 кГц) и воспроизводили через пьезокерамические преобразователи B&K 8104 (Bruel & Kjaer, Naerum, Дания). Два преобразователя, воспроизводящие стимулы в двух каналах, имели идентичные характеристики. Для контактной звуковой стимуляции ассистент экспериментатора, защищенный гидроизолирующим костюмом, находился в экспериментальном бассейне рядом с животным и удерживал излучатели звука таким образом, чтобы боковая цилиндрическая (излучающая) поверхность каждого из них касалась, но не прижималась к зоне акустического окна на латеральной поверхности нижней челюсти, под водой: один – справа, другой – слева.
При использованном способе стимуляции животное воспринимало звуковое излучение в ближнем поле преобразователя. В этих условиях интенсивность воспринимаемого звука не могла быть охарактеризована способом, принятым для дальнего поля, как уровень звукового давления в точке приема (дБ относительно 1 мкПа звукового давления). Поэтому интенсивность звука представляли в относительных единицах, как уровень источника звука: дБ относительно 1 мкПа на расстоянии 1 м от источника. Уровень источника звука рассчитывали по данным калибровки преобразователя в режиме излучения, представленной производителем.
Программа эксперимента и обработка данных. Основной методический прием данной работы состоял в применении парных звуковых стимулов от двух разных источников и определении времени освобождения ответа на второй стимул от последовательной маскировки. Одна из проблем, возникающих при такой схеме эксперимента, состоит в том, что при коротких межстимульных интервалах ответы на первый (кондиционирующий) и на второй (тестирующий) стимулы частично накладываются друг на друга, что затрудняет измерение характеристик второго ответа. Известное решение этой проблемы состоит в использовании процедуры вычитания: из ответов на пару стимулов вычитается ответ, вызванный только одним кондиционирующим стимулом. Предполагается, что таким образом можно выделить второй ответ в чистом виде.
Этот методический прием вполне эффективен, но он удовлетворительно работает только в том случае, если ответ, вызванный кондиционирующим стимулом в паре, и ответ, вызванный одним кондиционирующим стимулом, абсолютно идентичны. Это условие не всегда удается соблюсти по ряду причин. Одна из них состоит в том, что для выделения КСВП из шума приходится усреднять от нескольких сотен до нескольких тысяч реализаций. Хотя слуховая система дельфинов реагирует на стимулы, предъявляемые в темпе 10−20 или более имп/с, накопление необходимого количества реализаций требует времени, измеряемого минутами. За это время может измениться положение животного в носилках, положение излучателя относительно головы, или произойти другие изменения, хотя бы и незначительные. В результате в разностной кривой будет присутствовать не полностью скомпенсированный первый ответ, что не позволит точно измерить амплитуду второго ответа. Еще одна причина неполной компенсации первого ответа состоит в разной степени адаптации слуха. Серия из пар стимулов имеет больший средний уровень, чем серия из только кондиционирующих стимулов, что приведет к разной степени адаптации и также к неполной компенсации кондиционирующего ответа.
Чтобы исключить или хотя бы минимизировать эти эффекты, в данной работе было применено чередующееся предъявление пар стимулов и одиночных кондиционирующих стимулов (рис. 2).
Рис. 2.
Временная диаграмма предъявления кондиционирующего и тестирующего стимулов (только моменты предъявления стимулов, без точного воспроизведения их формы). Приведен фрагмент последовательности импульсов длительностью 400 мс (полная длина последовательности 125 с). Конд – кондиционирующий стимул, Тест – тестирующий стимул, Окно 1 – регистрация ответа на пару стимулов, Окно 2 – регистрация ответа на кондиционирующий стимул. Задержка тестирующего стимула относительно кондиционирующего не демонстрируется на сжатой шкале времени.

Ожидали, что при такой схеме эксперимента любые долговременные изменения чувствительности, как случайные, так и связанные с адаптацией, будут одинаково влиять на амплитуду кондиционирующих ответов при действии как парных, так и только кондиционирующих стимулов, что обеспечит необходимый результат процедуры вычитания.
Для выделения сигнала из шума методом усреднения последовательность, в которой чередовались парные и кондиционирующие стимулы, повторялась 1000 раз. Полученная запись содержала чередующиеся парные и одиночные ответы. Программа собственной разработки, созданная в программной оболочке LabVIEW, выделяла из этой записи 20-мс эпохи и сортировала их таким образом, чтобы получить усредненную запись ответов на пару стимулов и ответа на кондиционирующий стимул (рис. 3).
Рис. 3.
Выделение ответа на тестирующий стимул процедурой вычитания (билатеральная стимуляция). Негативность активного электрода соответствует отклонению вверх.
1 – ответ на пару стимулов с межстимульным интервалом 1 мс; 2 – ответ на кондиционирующий стимул; 3 – результат поточечного вычитания кривой 2 из кривой 1; выявляется ответ, задержанный на 1 мс относительно ответа на кондиционирующий стимул. Коротколатентные компоненты, характерные для ответа на кондиционирующий стимул, отсутствуют в разностной кривой.

Вычитанием одной формы потенциалов из другой программа также выделяла ответы на тестирующий стимул.
Для накопления данных, необходимых для выделения КСВП из фонового шума, чередующиеся стимулы (пара кондиционирующий-тестирующий и только кондиционирующий) предъявляли с частотой 16 имп/с, всего по 1000 стимулов каждого вида. Таким образом, накопление данных для получения одного комплекта КСВП занимало 125 с.
РЕЗУЛЬТАТЫ
Эффективность выделения ответа на тестирующий стимул. В соответствии с ожиданиями примененная методика чередующегося предъявления пар стимулов и кондиционирующего стимула дала хорошие результаты. Эффективность метода проверяли путем регистрации ответов при таких задержках между кондиционирующим и тестирующим стимулами, при которых перекрытие ответов на оба стимула было незначительным, так что можно было идентифицировать каждый из ответов. Пример такого эксперимента показан на рис. 3. При межстимульной задержке, равной 1 мс, суммарный ответ на пару стимулов имеет сложную форму и образован несколькими колебаниями; латентный период регистрируемого комплекса (включая акустическую задержку) 1.2 мс (1). Ответ только на кондиционирующий стимул имеет тот же латентный период, но меньшую длительность из-за отсутствия в комплексе ответа на задержанный стимул (2). Процедура вычитания дает кривую, в которой отсутствуют коротколатентные компоненты, т.е. отсутствует незадержанный ответ (3). Поэтому колебания потенциала, выделенного процедурой вычитания, являются ответом на задержанный стимул.
Приведенный пример показывает, что при равной интенсивности двух стимулов амплитуда задержанного (тестирующего) ответа меньше, чем амплитуда незадержанного (кондиционирующего). Этот эффект показывает, что тестирующие ответы, выделенные методом вычитания, могут быть использованы для измерения зависимости амплитуды тестирующего ответа от межстимульного интервала.
Зависимость амплитуды тестирующего ответа от межстимульного интервала при унилатеральной и билатеральной стимуляции. Используя методику разностных кривых для выявления ответа на тестирующий стимул, исследовали зависимость амплитуды тестирующего ответа от межстимульного интервала. Сравнивали результаты, полученные при унилатеральной и билатеральной стимуляции. Для компенсации возможной асимметрии слуховой чувствительности усредняли данные, полученные при симметричных вариантах расположения источников звука.
– При унилатеральной стимуляции регистрировали КСВП, когда источники обоих стимулов находились сначала справа, потом слева. Результаты обеих серий усредняли.
– При билатеральной стимуляции регистрировали КСВП, когда источник кондиционирующего стимула находился справа, а тестирующего – слева. Затем положения двух источников меняли на противоположные. Результаты обеих серий усредняли.
При всех вариантах стимуляции стимулы с приведенными выше параметрами вызывали четкие КСВП. Амплитуда тестирующего ответа зависела от межстимульного интервала.
На рис. 4 представлены КСВП, возникающие в ответ на задержанный (тестирующий) стимул с задержками от 0.3 до 7 мс, полученные при унилатеральном расположении источников обоих стимулов (усреднение данных, полученных при право- и левостороннем расположении).
Рис. 4.
КСВП на тестирующий стимул при различных межстимульных задержках при унилатеральной контактной стимуляции. Усреднение ответов, полученных при право- и левостороннем положении источников звука. Негативность активного электрода соответствует отклонению вверх. Представлены только тестирующие ответы, выделенные из оригинальных записей; кондиционирующие ответы удалены процедурой поточечного вычитания. Момент кондиционирующего стимула соответствует началу записи, момент тестирующего стимула отмечен стрелкой. Межстимульные задержки (от 0.3 до 7 мс) указаны около записей.
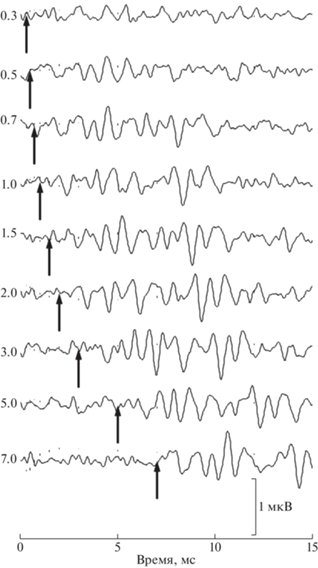
Ответы на кондиционирующий стимул в приведенных записях удалены применением процедуры вычитания. На рис. 5 представлены аналогичные КСВП, полученные при билатеральном расположении источников двух стимулов (усреднение данных, полученных при право- и левостороннем расположении источника кондиционирующего стимула и, соответственно, лево- и правостороннем расположении источника тестирующего стимула).
Рис. 5.
То же, что рис. 4, при билатеральной стимуляции. Усреднение ответов, полученных при право- и левостороннем положении источника кондиционирующих стимулов и, соответственно, лево- и правостороннем положении источника тестирующих стимулов.

Обе серии демонстрируют качественно сходную динамику зависимости ответа от межстимульного интервала: значительное угнетение ответа при коротких интервалах и восстановление при удлинении интервала. Количественно зависимость амплитуды КСВП от межстимульного интервала при унилатеральной и билатеральной стимуляции представлена графиками на рис. 6.
Рис. 6.
Зависимость амплитуды тестирующего КСВП (в процентах, от амплитуды кондиционирующего ответа на стимул такой же интенсивности) от межстимульного интервала при уни- и билатеральной стимуляции (как указано в легенде).

Амплитуду КСВП измеряли как разность между позитивным и негативным максимами ответа. Оба графика демонстрируют значительное угнетение тестирующего ответа при коротких интервалах (начиная с 0.3 мс) и восстановление при удлинении интервалов до 5 мс и более. Графики для унилатеральной и билатеральной стимуляции не совпадают, но расхождения между ними имеют характер скорее случайных отклонений, чем закономерного различия. Различия между двумя графиками не достигают критерия статистической достоверности (P = 0.22).
ОБСУЖДЕНИЕ
Основной результат проведенных исследований состоял в том, что унилатеральная (оба стимула приходят с одной стороны) и билатеральная (кондиционирующий и тестирующий стимулы приходят с разных сторон головы) cтимуляции вызывают у дельфина практически одинаковую последовательную маскировку тестирующего стимула: значительное подавление тестирующего ответа при межстимульных интервалах короче 1 мс и освобождение от маскировки при интервалах от нескольких мс.
Электрод для регистрации КСВП, установленный на средней линии головы, одинаково регистрирует электрические потенциалы, генерируемые право- и левосторонними структурами мозга. КСВП, регистрируемый в таких условиях, является суммой электрической активности право- и левосторонних структур.
Использованный в данной работе метод контактной стимуляции позволяет определить, на каком уровне слуховой системы развивается последовательная маскировка, проявляющаяся в снижении амплитуды КСВП при парных стимулах с короткими интервалами. Метод контактной стимуляции не может полностью устранить “перекрестную” стимуляцию, т.е. стимуляцию правой слуховой улитки при расположении источника звука слева, или наоборот, поскольку не исключено распространение звука к слуховой улитке противоположной стороны. При контактной стимуляции следует, по меньшей мере, ожидать, что стимуляция ипсилатеральной слуховой улитки будет намного более интенсивной, чем контралатеральной. Поэтому представленные выше данные позволяют оценить, в какой части слуховой системы – монауральной или бинауральной – возникают эффекты последовательной маскировки. Если бы эффекты последовательной маскировки возникали в монауральных слуховых структурах, бинауральная стимуляция (кондиционирующий стимул подается в одну слуховую улитку, тестирующий – в другую) не могла бы привести к значительному угнетению тестирующего ответа. Одинаковые эффекты последовательной маскировки для унилатеральной и билатеральной стимуляции означают, что эффекты последовательной маскировки развиваются преимущественно в бинауральных слуховых структурах, где афферентные потоки от обеих улиток взаимодействуют.
Применительно к анализу звуковых сцен в естественных ситуациях этот вывод означает, что анализ временной динамики звуковых сигналов от “целевого” источника зависит от наличия источников звука в других секторах сцены – не только в ипсилатеральном, но и в контралатеральном секторе, и звуковой шум от этих источников может маскировать сигнал. Вместе с тем, благодаря высокому быстродействию процессов в слуховой системе дельфина, в любом случае последовательная маскировка кратковременная, и освобождение от маскировки происходит в течение нескольких мс после прекращения действия маскера.
Работа поддержана Российским научным фондом (грант 22-25-00025).
Список литературы отсутствует.
Дополнительные материалы отсутствуют.
Инструменты
Сенсорные системы