

Сенсорные системы, 2023, T. 37, № 1, стр. 35-48
Магнитная навигация животных, контрастная чувствительность зрения и закон Вебера–Фехнера
В. Н. Бинги *
Федеральный исследовательский центр Институт общей физики им. А.М. Прохорова РАН
119991 Москва, ул. Вавилова, 38, Россия
* E-mail: vnbin@mail.ru
Поступила в редакцию 13.10.2022
После доработки 28.10.2022
Принята к публикации 02.11.2022
- EDN: ATRZSG
- DOI: 10.31857/S0235009223010031
Аннотация
Известно, что некоторые животные могут реагировать на очень малые изменения магнитного поля (в тысячу раз меньше геомагнитного поля) и используют это для навигации по магнитному рельефу Земли. Природа молекулярного сенсора магнитного поля остается, однако, неопределенной, хотя установлено, что магнитное чувство связано со зрением. Полагают, что магнетохимическая реакция лежит в основе функционирования магнитного сенсора. Криптохромы фоторецепторов, выстилающих сетчатку глаза, содержат фотоиндуцированные спин-коррелированные пары радикалов, участвующие в формировании нервного импульса и чувствительные к магнитному полю. Поэтому животное могло бы ощущать магнитное поле как изменение яркости больших полей зрения и ориентироваться по их контрасту. Однако чувствительность отдельных сенсоров – радикальных пар – очень низка. Ранее предполагали, что это затруднение преодолевается статистическим усилением контрастной чувствительности за счет параллельной обработки мозгом первичных сигналов миллионов фоторецепторов. В настоящей работе эта гипотеза проверена. Найдено, что пороговое ощущение яркостного контраста почти линейно зависит от логарифма углового размера контрастирующего стимула, что является характерным для физиологии ощущений, подчиняющихся закону Вебера–Фехнера. Контрастная чувствительность увеличивается с ростом числа фоторецепторов, участвующих в распознавании стимула, однако, количественно этого увеличения недостаточно для надежного объяснения магнитной навигации животных.
ВВЕДЕНИЕ
Известно, что некоторые виды сезонно мигрирующих животных обладают способностью ощущать малейшие изменения магнитного поля (МП) Земли и использовать эту способность для ориентирования и навигации – нахождения мест сезонного обитания в условиях отсутствия других ориентиров. Эту способность называют магнитным чувством или магниторецепцией. Благодаря специализированным магнитным рецепторам животные могут осуществлять миграции даже ночью в условиях облачности. Специальным рецептором обладают, например, некоторые птицы (Hore, Mouritsen, 2016), морские черепахи (Lohmann et al., 2004) и насекомые (Wan et al., 2021). Установлено, что этот рецептор совмещен со зрительной системой и обладает чувствительностью к изменениям МП порядка нескольких десятков нТл. Полагают, что животные “видят” изменения МП как крупные области их поля зрения, отличающиеся по яркости, – как области затемнения или осветления. Заметим, что имеются экспериментальные данные о том, что организм человека также реагирует на столь малые изменения МП, хотя, субъективно, и не “видит” их (Pishchalnikov et al., 2019).
Предполагаемый молекулярный механизм этого явления основан на магнетохимической реакции – зависимости переноса фотоэлектрона, образующего спин-коррелированную радикальную пару с другим электроном внутри белка, от спинового состояния пары, которое в свою очередь зависит от МП (Schulten et al., 1978; Hore, Mouritsen, 2016; Buchachenko, 2014). Проблема, наряду с другими нефизическими трудностями этого предположения (Астахова и др., 2019), состоит в низкой чувствительности данного механизма к МП, теоретически около 0.1% на 50 мкТл – уровень геомагнитного поля. В экспериментах in vitro чувствительность еще на порядки меньше (Binhi, Prato, 2018). В работе (Binhi, Prato, 2017) авторы предположили, что необходимое усиление сигнала МП могло бы быть достигнуто за счет параллельной обработки мозгом сигналов, одновременно поступающих от миллионов фоторецепторов. Так, сетчатка глаза человека содержит около 90 млн палочек и около 5 млн колбочек (Curcio et al., 1990). Параллельная обработка сигналов могла бы заключаться в чем-то близком к их суммированию или осреднению. По закону больших чисел результат очень мало отличался бы от истинного среднего. В соответствии с центральной предельной теоремой математической статистики относительное отклонение от среднего уменьшалось бы, а отношение сигнал–шум (чувствительность к МП), наоборот, возрастало пропорционально корню из числа рецепторов, задействованных в детектировании МП. Известно, например, что некоторые насекомые улучшают их контрастную чувствительность на два порядка за счет суммирования сигналов многих нейронов (Stöckl et al., 2016).
Цель настоящей работы состояла в экспериментальной проверке этой гипотезы. Животное, как предполагается, реагирует на изменение яркости части поля зрения вследствие зависимости сигналов фоторецепторов от направления и/или величины МП. При действии относительно слабого МП, порядка геомагнитного, отклик отдельного фоторецептора на сигнал МП по механизму радикальных пар невелик. Относительное изменение скорости ракции с участием такой пары не превышает, как сказано выше, 0.001 по порядку величины. Правдоподобно, что синхронные магнитные сигналы множества упорядоченных на сетчатке фоторецепторов приводят к изменению ощущения яркости, как если бы яркость объекта восприятия, изображение которого на сетчатке попадает на указанную группу фоторецепторов, изменилась на ту же относительную величину.
Поэтому действие МП ниже было смоделировано в виде контролируемого изменения яркости визуального стимула серого цвета. Если бы оказалось, что человек способен различать два поля визуального стимула с мало отличающейся яркостью, и предельно обнаружимая разность яркости уменьшалась (а чувствительность, соответственно, росла) вместе с ростом углового размера сравниваемых полей – с ростом числа задействованных фоторецепторов, то действие слабого МП на зрительный аппарат животных было бы менее парадоксальным.
Принципы обработки визуальной информации мозгом человека и, например, птицей, осуществляющей магнитную ориентацию и навигацию, конечно, сильно отличаются. Отличия определяются многими факторами: бинокулярность зрения против латеральности; отличия в микродвижении глаз; иные множества распознаваемых объектов; цели обработки информации; объемы зрительного восприятия; глубины резкости и величины аккомодации; времени сохранения ощущений и многое другое. Здесь подобные вопросы психофизиологии зрения не обсуждаются, поскольку задачей является только установление типа зависимости минимально обнаружимой разности яркости от размера воспринимаемого объекта – только то, что имеет прямое отношение к проверяемой гипотезе.
Способность зрения обнаруживать различия в яркости называется контрастным восприятием. В литературе имеется несколько количественных определений ахроматического яркостного контраста (Pelli, Bex, 2013) в соответствии со спецификой задач исследования. Ниже использован так называемый контраст Майкельсона, описывающий относительную разницу равномерных яркостей ${{b}_{1}} > {{b}_{2}}$ различных участков изображения следующим образом:
(1)
$k \equiv \frac{{{{b}_{1}} - {{b}_{2}}}}{{{{b}_{1}} + {{b}_{2}}}} = \frac{{{{\Delta }}b}}{{2b}},$Зрение способно различать небольшие по величине контрасты. Для описания такой способности используют понятие порогового контраста $k{\kern 1pt} '$, т.е. минимального обнаружимого контраста. Известно, что пороговый контраст зависит от многих факторов. Детально изучены зависимости от геометрии изображений, сравниваемых по яркости, от базового уровня яркости, неоднородности яркости и от временных характеристик сравнения (Bertalmio, 2020). Эти закономерности широко используют на практике для эффективного кодирования изображений, в фото и киноиндустрии, в психологии, нейрологии, офтальмологии. Настоящая работа сфокусирована на изучении зависимости порогового контраста от телесного угла стимула или его площади, если угол не слишком велик.
Различение контрастов подчиняется основному психофизиологическому закону – закону Вебера–Фехнера. Закон Вебера утверждает, что минимально обнаружимое изменение ${\text{d}}\xi $ интенсивности, или силы какого-либо раздражителя, пропорционально силе ξ раздражителя: ${\text{d}}\xi \propto \xi $. Фехнер ввел количественное определение ощущения ψ и постулировал, что равные изменения ${\text{d}}\psi $ ощущения вызываются равными относительными изменениями силы раздражителя ${\text{d}}\xi {\text{/}}\xi $, т.е. ${\text{d}}\psi = \gamma ~{\text{d}}\xi {\text{/}}\xi $. Отсюда следует закон Вебера–Фехнера
где ${{\xi }_{0}}$ – константа. Другими словами, интенсивность ощущения приблизительно есть линейная функция логарифма силы раздражителя.Поскольку ψ – это квалиа, относится к области индивидуальных ощущений, доступных лишь “первому лицу”, величина $\psi $ и коэффициент γ по отдельности не могут быть определены операционально. Поэтому далее под численной оценкой ощущений понимается доступное математическим абстракциям безразмерное отношение $y \equiv \psi {\text{/}}\gamma = {\text{ln}}\left( {\xi {\text{/}}{{\xi }_{0}}} \right)$.
Как видно, ${{\xi }_{0}}$ есть максимальное значение силы раздражителя ξ, еще не вызывающее появление ощущения. Существует, однако, фоновый уровень ощущений, связанный с наличием внутренних собственных шумов (Красильников, Шелепин, 1997). Шум присутствует в информационных каналах передачи при последовательном преобразовании информации от внешних сигналов к сигналам рецепторов и далее к нейронным возбуждениям, ответственным за возникновение ощущений. Поэтому более корректно записать закон Вебера–Фехнера в виде
где ${{y}_{0}}$ – ощущение, вызываемое собственным шумом сенсорного канала, а ${{\xi }_{0}}$ – максимальная сила раздражителя, еще не вызывающая превышения силы ощущения над ${{y}_{0}}$, или наоборот, минимальная сила, только начинающая вызывать такое превышение. Величина ${{\xi }_{0}}$ может быть приблизительно определена в каждом конкретном случае по субъективным ощущениям испытуемого при постепенном увеличении ξ от нуля или снижении от больших значений.Поскольку ${{y}_{0}}$ – это ощущение собственного шума, идеализируют, что ${{y}_{0}}$ не зависит от силы раздражителя и проявляется в виде неопределенности ощущения y при любом значении раздражителя в физиологически нормальном интервале. Другими словами, ${{y}_{0}}$ – это минимально возможная разница в ощущениях, воспринимаемая именно как разница. Математически это обстоятельство можно записать в следующем виде:
где ${{\Delta }}\xi $ – минимальное изменение силы раздражителя, обнаружимое по разнице ощущений. Отсюда и из (2) следует закон Вебера где c – численный коэффициент, точное значение которого несущественно, поскольку c фактически представляет выбранный масштаб для ощущений y.Чувствительность E к раздражителю ξ определяют как величину, обратную к ${{\Delta }}\xi $,
чем сильнее раздражитель, тем ниже чувствительность. Таковы основные соотношения, следующие из закона Вебера–Фехнера.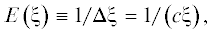
Далее речь идет о зрительных ощущениях, индуцируемых световым потоком и зависимых таким образом от яркости стимула $b$. Закон Вебера тогда имеет вид
Так как $c \ll 1$ и, следовательно, ${{\Delta }}\xi \ll \xi $, то ${{\Delta }}\xi = {{\Delta }}b{\text{\;d}}\xi {\text{/d}}b$, откуда после подстановки в (3) получаем соотношение, определяющее минимально ощутимую разность яркостей,
(4)
${{\Delta }}b = c{{\left[ {\frac{{\text{d}}}{{{\text{d}}b}}{\text{ln}}\left[ {\xi \left( b \right)} \right]} \right]}^{{ - 1}}}.$Заметим, что физическим раздражителем является не яркость сама по себе, а скорее поток света, зависящий еще и от телесного угла, под которым виден стимул или объект. Минимальная разница силы раздражителя, обнаружимая при каком-то значении b, как и сама сила раздражителя ξ, будет тогда зависеть от размера стимула.
Поток света, попадающий на сетчатку от объекта, может быть одинаков для яркого объекта малого размера и тусклого объекта большого размера. Значит, надо ожидать, что минимальное различимое приращение яркости будет меньше для больших фигур и больше для малых. Другими словами, создавая одинаковые приращения силы раздражителя изменение яркости будет большим для малого и малым для большого объекта. Рассматривается, конечно, идеализированная ситуация, где стимул – это простая плоская фигура с относительной площадью a и c равномерной яркостью b. Даже и в этом случае, вряд ли можно ожидать, что сила раздражителя пропорциональна, например, произведению $ab$, поскольку сила раздражителя может зависеть не только от физического стимула, но и от его психического коррелята. Например, в согласии с законом Вебера, минимальная распознаваемая разница длин двух отрезков линий, изображенных на бумаге, пропорциональна их длине, в то время как раздражителем является не поток света, а мысленный образ отрезков. Поэтому вид зависимости $\xi \left( {a,b} \right)$ остается неопределенным. Соответственно, получим из (4)
(5)
${{\Delta }}b = c{{\left[ {\frac{{\text{d}}}{{{\text{d}}b}}{\text{ln}}\left[ {\xi \left( {a,b} \right)} \right]} \right]}^{{ - 1}}}.$Есть существенное отличие в интерпретациях соотношений (4) и (5). Согласно (4) значению b соответствует минимальная ощутимая разница яркостей ${{\Delta }}b$. Соответствие взаимное: заданная, или фиксированная, минимально ощутимая разница определяет яркость b, при которой она имеет место. Согласно соотношению (5) заданная минимально ощутимая разница ${{\Delta }}b$ определяет взаимозависимость между a и b, при которой обеспечивается заданный уровень ${{\Delta }}b$. Если яркость фиксирована каким-либо значением $b{\kern 1pt} *$, как будет в нижеописываемом эксперименте, то заданный уровень ${{\Delta }}b$ будет определять площадь стимула a, при которой ${{\Delta }}b$ приобретает заданное значение. Учитывая это обстоятельство и разделив (5) на $2b$, получим выражение для порогового контраста в следующем виде
(6)
$k{\kern 1pt} ' = c{{\left[ {2b{\kern 1pt} *\frac{{\text{d}}}{{{\text{d}}b}}{\text{ln}}\left[ {\xi \left( {a,b{\kern 1pt} *} \right)} \right]} \right]}^{{ - 1}}},$(7)
$E\left( a \right) \propto b{\kern 1pt} *\frac{{\text{d}}}{{{\text{d}}b}}{\text{ln}}\left[ {\xi \left( {a,b{\kern 1pt} *} \right)} \right],$Закономерные связи между пороговым контрастом и размером контрастных полей изучены мало. С одной стороны, тот факт, что предмет большего размера виден при меньших контрастах, известен давно (Blackwell, 1946; Муравьева и др., 2010). Пороговый контраст уменьшается, а контрастная чувствительность $1{\text{/}}k{\kern 1pt} '$ растет при увеличении углового размера изображений. С другой стороны, редкие утверждения в литературе относительно типа такой зависимости противоречивы. В работе (Riccó, 1877) автор приводит закономерность $k{\kern 1pt} '{{\Omega }} = {\text{Const}}$, где Ω – телесный угол объекта, но она применима для объектов малого углового размера, звезд, например, когда объект выглядит единым целым (Crumey, 2014); напомним, что разрешающая способность глаза человека около $1'$ (угл. мин). Контрастная чувствительность человека (контраст Вебера) в большинстве случаев составляет приблизительно 1% (Bertalmio, 2020: Nadler et al., 1990). Иногда утверждают, что контрастная чувствительность не зависит в широких пределах от размера стимула (Pelli, Bex, 2013). В работах (Кучерявый, 2004; Домбругов, 1979), напротив, имеются данные о такой зависимости, но тип ее неясен.
Наиболее полно связь контрастной чувствительности и размера объекта изучена в работе (Blackwell, 1946), где измеряли контрастный порог у нескольких испытуемых при различных яркостях фона и размерах диска с отличающейся от фона яркостью. Зависимость от размера носила сложный характер. Диапазон угловых размеров диска составлял 0.01–6°, что существенно меньше предполагаемых угловых размеров полей зрения животных, осуществляющих магнитную навигацию по яркостному контрасту.
В настоящей работе исследована зависимость $k{\text{'}}$ от размера стимула в большем диапазоне угловых размеров сравниваемых полей зрения – до 16° на одно поле и 32° на целый контрастирующий объект, или стимул. Кроме того, изучался пороговый контраст двух в остальном одинаковых полей изображения, а не контраст диска на окружающем фоне, использована современная статистическая обработка данных и дан комментарий относительно типа зависимости контрастной чувствительности от размера стимула.
ОПИСАНИЕ ТЕСТА
Компьютерные тесты давно используются для исследования закономерностей контрастной чувствительности, поскольку позволяют легко и в широких пределах менять контраст и пространственную структуру изображений (Pelli, Bex, 2013).
В настоящей работе для определения зависимости контрастной чувствительности зрения человека от телесного угла предъявляемого стимула был разработан специализированный компьютерный тест, учитывающий особенности восприятия контрастных изображений. Хорошо известно, что контрастная чувствительность зависит от плавности перехода между полями разной яркости (Watson, Ahumada, 2005) – эффект “полос Маха”. Для изучения этого эффекта используют изображения в виде решеток с различной пространственной частотой контрастирующих полос и различной глубиной контраста. Оказывается, зрение человека наиболее чувствительно в среднем пространственно-частотном диапазоне. Контраст как мелких деталей, так и крупных областей, различается хуже. Предметом настоящей работы была контрастная чувствительность в пространственно-низкочастотной области, и присутствие резких переходов между контрастирующими полями, содержащих хорошо распознаваемые пространственные спектральные компоненты, было нежелательным. Поэтому в настоящей работе изучался пороговый контраст между двумя полями, разделенными полосой фоновой яркости, что исключало сравнение полей по границе их непосредственного примыкания. Это лучше соответствует поставленной задаче моделирования визуального поля зрения с двумя большими мало отличающимися по яркости областями. Специальный интерес представляла зависимость чувствительности от размера предъявляемого стимула.
В серии из нескольких сот испытаний испытуемому предъявлялся визуальный стимул – избражение на экране монитора, рис. 1.
Рис. 1.
Общий вид предъявляемого изображения, около 60 см по диагонали, для обнаружения более темного поля из двух смежных полей на изображении.
Контраст между смежными полями на рисунке $g = 0.038$ (10 бит); в тестировании использовались контрасты 0, 1 и 2 бита.

Изображение представляло случайно расположенные два смежных пятна, или поля прямоугольной формы заданной площади, приблизительно пропорциональной телесному углу, серого цвета с немного разной яркостью. Всего имелось пять значений площади – 60; 600; 6000; 60 000 и 600 000 пикселей. Для обозначения площади далее используется относительная величина a, т.е. отношение заданной площади поля к минимальному значению площади ($a = 1$, 10; 100; 1000; 10 000) либо индекс площади ${{x}_{i}} \equiv {\text{lg}}\left( {{{a}_{i}}} \right)$. Угловой размер объекта $u \equiv {\text{arctg}}\left( {l{\text{/}}d} \right)$, где $l \propto \sqrt a $ – линейный размер объекта и d – расстояние до него, в описываемом эксперименте около 60 см. Угловые размеры по ширине минимального и максимального изображений полей составили приблизительно 0.16° и 15.8°, или 9.7' и 950' соответственно. Область угловых размеров полей за пределами этого диапазона нами не исследовалась ввиду ограниченных характеристик монитора.
Под яркостью b далее понимается не фотометрическая яркость,т.е. отношение силы света, испускаемой в заданном направлении, к площади светящегося объекта, а связанная с ней относительная величина средней яркости поверхности монитора, когда 0 соответствует минимуму, а 1 – максимуму светимости. Испытуемый должен был ответить на вопрос, какое поле темнее – левое или правое? Ответ был либо верным, либо ошибочным, что позволяло легко оценивать вероятность распознавания. Один из уровней яркости был 0.5, а другой доходил до 0.49 (заметно темнее при большой площади поля). Контраст Майкельсона k (1) в этом случае, следовательно, связан с разностью яркости соотношением $k \approx {{\Delta }}b$, что было удобно.
Тестирование проводилось с использованием монитора Apple Led Cinema 27. Максимальная яркость 330 кд/м2, разрешение 2560 × 1440 (109 п/дюйм), контрастность 1000: 1, однородность распределения яркости в районе 11 кд/м2. В мониторе используется LCD матрица LM270WQ1 от LG Electronics, которая построена на базе технологии H-IPS. Цветовое разрешение 16.7 млн цветов RGB, или 8 бит/цвет. Это означает, что имеется ${{2}^{8}} = 256$ градаций яркости на один цветовой канал. Если весь диапазон изменения яркости и разности яркостей, т.е. в отсутствие и при полной засветке экрана монитора, помещается в интервал (0, 1), то минимальный шаг яркости серого при равной светимости субпикселей RGB есть один бит или 1/256 ≈ 0.0039.
Фотометрические измерения светимости экрана не проводились, поскольку цель состояла в выяснении типа функциональной зависимости контрастной чувствительности фотопического зрения от размера стимула. Яркость при полной засветке экрана в эксперименте составила около 200 кд/м2. Контраст в 1 бит при линейной зависимости яркости от задаваемого относительного уровня соответствовал изменению яркости на величину порядка 0.8 кд/м2.
В предварительном тесте, направленном на установление предельного контраста при максимальном размере стимула, использовали 10 значений задаваемого контраста g от 0.001 до 0.01 с шагом 0.001. В основном тесте использовалось шесть значений задаваемого контраста – 0.001; 0.002; 0.004; 0.006; 0.008 и 0.01, по два значения на каждый отображаемый монитором контраст κ (каппа). Повторим, из-за дискретности светимости монитора отображаемый монитором контраст κ не совпадает с задаваемым в программе контрастом g.
Основная часть тестирования была направлена на то, чтобы найти индивидуальный пороговый контраст в зависимости от размера полей стимула. Во всех испытаниях поля обязательно различались по программно заданному контрасту. Более темное поле появлялось слева или справа случайно, с вероятностью 0.5. Если бы испытуемый все время нажимал кнопку СЛЕВА или СПРАВА, то число правильных ответов было бы равно в среднем половине числа попыток, с известным стандартным отклонением. Если на кнопки нажимать случайно, то правильных ответов также будет около половины. Ожидалось, что если испытуемый правильно распознает разницу яркости полей и нажимает соответствующую кнопку, то доля правильных ответов будет выше случайного уровня.
Тестирование проводилось сериями по 200–400 отдельных испытаний, или попыток, для каждого значения площади пятна. Длительность одной серии составляла от 10 до 30 мин и зависела, главным образом, от уровня усилий испытуемого дать правильный ответ. На каждое отдельное испытание испытуемый тратил от долей до нескольких секунд; время на ответ не ограничивалось. Для повышения заинтересованности в верном распознавании испытуемые были заранее извещены о материальной стимуляции за верное распознавание контрастов выше уровня случайности.
Если бы тестирование в серии проводилось при единственном значении заданного контраста, то было бы возможно субъективное влияние испытуемого на результат. Так, если бы испытуемый не ощущал контраста в ходе нескольких начальных попыток, то он мог посчитать, что установленное значение контраста слишком мало для различения и перестать совершать усилия для различения. Для того чтобы устранить такой субъективизм, одна серия тестирования осуществлялась для двух значений контраста, выбираемых программой случайным образом так, что испытуемый заранее не знал о контрасте демонстрируемого стимула. Контрасты g шли достаточно близкими парами 0.001 и 0.004; 0.002 и 0.006; 0.008 и 0.01, так что распознать разницу между ними можно было только статистически. Серии испытаний повторялись несколько раз на каждую пару значений контрастов и площади стимула.
По завершении серии число попыток N и число верных ответов n записывалось в файл для каждого значения из пары контрастов. Затем значения площади или контрастов менялись, и начиналась новая серия испытаний. По завершении каждой серии испытаний осуществлялся перерыв. Это было необходимо, поскольку чаще всего испытуемому приходилось принимать определенное решение, когда он не ощущал контраста – при этом усиливался дискомфорт и накапливалась усталость.
Тестирование прошли двое испытуемых 60– 70-летнего возраста в здоровом состоянии, обладающие умеренной миопией и пресбиопией, с использованием корректирующих очков. Пробными тестированиями не удалось установить, что уровень естественного фонового освещения в месте эксперимента закономерно влиял на результаты, скорее всего потому, что фоновое освещение “засвечивало” лишь боковое поле зрения, а основную часть поля зрения занимало изображение монитора с темным фоном. Поэтому специальных мероприятий для обеспечения строго одинаковых условий фонового освещения не предпринималось. Эксперимент проводился в условиях как естественного дневного, так и искусственного вечернего освещения, с перерывами, и был закончен за четыре недели.
Всего было проведено более 18 тысяч отдельных испытаний, все из которых использованы в обработке. Еще около трех тысяч начальных испытаний не вошли в обработку, но были необходимы для установления релевантных пределов изменения контрастов и размеров стимулов и для уточнения дизайна и программного кода эксперимента.
ОБРАБОТКА ИЗМЕРЕНИЙ
В нашем эксперименте число повторов серий испытаний с одинаковыми x и κ было от четырех до восьми. При малом числе повторов наблюдаемая дисперсия могла оказаться малой случайно, из-за эффекта выборки. Это, конечно, не означает, что истинная дисперсия столь же мала. Поэтому в оценке вариабельности измерений использованы ожидаемые дисперсии, вычисленные на основе теоретического распределения числа верных ответов (см. Приложение). Параметры распределения определялись из имеющихся экспериментальных значений. Ожидаемые дисперсии больше наблюдаемых и точнее отражают разброс данных, который бы имел место при большом числе повторов испытаний.
Ожидаемые дисперсии вычислялись путем симуляции возможных экспериментальных данных на основе имеющихся (см. Приложение). Было проведено 20 симуляций, построены соответствующие им зависимости контрастной чувствительности от размера стимула и найденная усредненная зависимость.
РЕЗУЛЬТАТЫ
В предварительном тестировании определяли зависимость вероятности распознавания q от программно задаваемого контраста g. На рис. 2 приведена зависимость $q\left( g \right)$ в интервале $g \in \left( {0.001,{\text{\;}}0.01} \right)$ для максимального размера стимула $x = 4$.
Рис. 2.
Средняя по испытуемым оценка вероятности распознавания программно заданных контрастов – синие кружки, вероятность распознавания заданных контрастов 0.0038 и 0.004 – красные квадраты.
Размер стимула $x = 4$. Показаны стандартные отклонения.
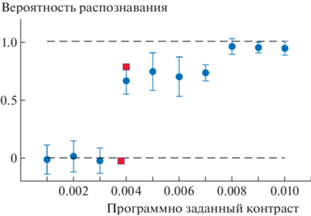
Видна дискретность отображения контрастов дисплеем. Например, при изменении контраста g от 0.004 до 0.007 ощущаемый контраст меняется мало, поскольку соответствующие вероятности распознавания, по-видимому, равны. Другими словами, имеем равные отображаемые контрасты κ при задании контрастов g в интервалах 0.001–0.003; 0.004–0.007 и 0.008–0.01.
Для подтверждения того, что монитор демонстрирует изменение контраста лишь при переходе g через границу в 1 бит, проведен отдельный тест с программно заданным контрастом 0.0038 (<1 бит), либо случайно 0.004 (>1 бит), при максимальном в эксперименте размере стимула $x = 4$. Результат испытуемого А составил 92 из 189 при 0.0038 и 188 из 211 при 0.004. Эти точки, вероятности q, представлены на рис. 2 квадратами и показывают скачкообразное изменение q при переходе устанавливаемого контраста через 1 бит.
Нетрудно заключить, что отображаемые на мониторе контрасты определяются целыми значениями бит – 0, 1, 2 и так далее. Исследуемый диапазон контрастов в настоящей работе ограничен значениями κ = 0, 1 и 2 бит, которым, как сказано выше, соответствуют программно задаваемые контрасты от 0 до 0.0039 (0 бит), от 0.004 до 0.0078 (1 бит) и от 0.008 до 0.01 (2 бита). Поэтому исследование зависимости распознавания от контрастов имеет пределы: можно сказать лишь, что в интервале контрастов от 0 до 2 бит вероятность распознавания меняется от 0 до приблизительно 1. Это, однако, не мешает установить зависимость вероятности распознавания от размера стимула.
Тестирование в условиях отсутствия отображаемого контраста при задаваемых значениях 0.001; 0.002 и 0.003 нуждается в комментарии. Во всех этих случаях контраст κ полей стимула на мониторе отсутствовал, но испытуемому приходилось отвечать на вопрос “справа или слева находится более темное поле?”. Вопрос в данных условиях становился некорректным. Испытуемый не знал об отсутствии контраста. В этом случае любой ответ не был интерпретируемым в терминах вопроса, а измеренное отношение числа “правых”, как и “левых”, ответов к числу попыток отражало лишь оценку вероятности случайного выбора из двух априори равновероятных возможностей. Эта оценка не может быть приписана “вероятности правильного ответа” p или “вероятности распознавания” $q \equiv 2p - 1$ контраста. Очевидно, вероятность распознавания отсутствующего контраста должна быть положена равной нулю, что и было сделано в последующих расчетах q и в определении зависимости порогового контраста от размера стимула.
В основной части тестирования определялись вероятности q при различных значениях размера стимула x и при задаваемых контрастах g = 0.001; 0.002; 0.004; 0.006; 0.008; 0.1. В силу вышеуказанной дискретности отображаемых контрастов κ результаты, полученные для контрастов 0.001 и 0.002, объединялись и приписывались контрасту $\kappa = 0$ бит. Соответственно, результаты для контрастов 0.004 и 0.006 приписывались контрасту $\kappa = 1$ бит, а 0.008 и 0.01 – контрасту $\kappa = 2$ бита.
Результаты обработки накопленных данных представлены в виде таблицы оценок вероятностей распознавания q и их стандартных отклонений, усредненные по двум испытуемым, где в первом столбце приведены значения индекса площади ${{x}_{i}} \equiv {\text{lg}}\left( {{{a}_{i}}} \right)$, в верхнем ряду – контрасты ${{\kappa }_{k}}$, а в остальных содержатся вероятности $q\left( {{{x}_{i}},{{\kappa }_{k}}} \right)$, т.е. матрица ${{q}_{{i,k}}} \pm \sigma _{{i,k}}^{q}~$ (табл. 1).
Таблица 1.
Усредненные вероятности распознавания
| x | $\kappa = 0$ | $\kappa = 1$ | $\kappa = 2$ |
|---|---|---|---|
| 0 | 0.0243 ± 0.13 | 0.104 ± 0.113 | 0.368 ± 0.297 |
| 1 | 0.0185 ± 0.142 | 0.301 ± 0.22 | 0.54 ± 0.28 |
| 2 | −0.0397 ± 0.124 | 0.447 ± 0.195 | 0.828 ± 0.0739 |
| 3 | −0.0264 ± 0.125 | 0.636 ± 0.243 | 0.945 ± 0.0326 |
| 4 | −0.004 ± 0.121 | 0.673 ± 0.13 | 0.949 ± 0.063 |
Построенные по этим данным зависимости вероятности распознавания q от индекса размера стимула x показаны на рис. 3 для трех значений контраста κ в битах.
Рис. 3.
Усредненные вероятности распознавания q в зависимости от размера стимула x для разных значений контраста κ.
Показаны ошибки средних.
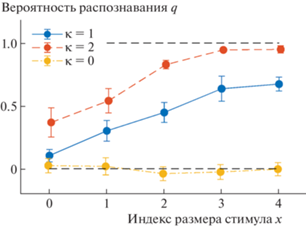
Видно, что с ростом контраста вероятность распознавания увеличивается. При ненулевом контрасте вероятность q увеличивается и с ростом x. При нулевом контрасте распознавание отсутствует.
Для того чтобы связать контрастную чувствительность с размером стимула, надо аппроксимировать полученные значения $q\left( {x,\kappa } \right)$ подходящей гладкой функцией. Для этого использована регрессия q на x и κ. Из вида зависимостей рис. 3 следует, что подходящей является регрессия в виде квадратичной функции двух переменных. Она представляет собой поверхность второго порядка $W\left( {x,\kappa } \right)$ переменных x и κ, принимающих уже непрерывные значения в интервалах [0, 4] и [0, 2] соответственно.
Результат регрессии показан на рис. 4, а, где изображены экспериментальные точки ${{q}_{{i,k}}}$ и функция, построенная по коэффициентам регрессии, поверхность $W\left( {x,\kappa } \right)$, аппроксимирующая точки.
Рис. 4.
а – результат регрессии q на x и κ квадратичной функцией $W\left( {x,\kappa } \right)$, б – способ определения зависимости $\kappa {\kern 1pt} ' = \kappa {\kern 1pt} '\left( x \right)$ минимально обнаружимого контраста от индекса размера стимула.
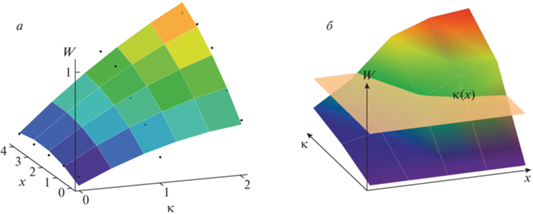
Далее надо ввести определение того, что называется пороговым контрастом $\kappa {\kern 1pt} '$. Обычно его определяют как контраст, соответствующий вероятности распознавания, равной 1/2. Другими словами, если вероятность q распознавания превышает 0.5, то можно считать, что контраст распознан, или обнаружен.
Уравнение $W\left( {x,\kappa } \right) = 0.5$ неявно задает функцию $\kappa {\kern 1pt} ' = \kappa {\kern 1pt} '\left( x \right)$, которая определяет минимальное значение контраста, обнаружимого при площади стимула x с вероятностью 0.5. Эта функция дается линией пересечения поверхности $W\left( {x,\kappa } \right)$ плоскостью $W = 0.5$, рис. 4, б. Контрастная чувствительность E тогда есть функция, обратная к $\kappa {\kern 1pt} '\left( x \right)$, т.е. $E\left( x \right) \equiv 1{\text{/}}\kappa {\kern 1pt} '\left( x \right)$.
Заметим, что если бы тестирования были повторены, то значения матрицы ${{q}_{{i,k}}}$, где $i = 0,{\text{\;}}...,{\text{\;}}4$ соответствует индексу площади и $k = 0,{\text{\;}}1,{\text{\;}}2$ контрасту (табл. 1), были бы другими, поскольку имеются как эмпирические, так и теоретические отклонения доли правильных ответов от их ожидаемых значений. Как было сказано выше, экспериментальные данные в случае гетерогенного массива измерений могут не отражать истинного размера разброса данных (их неопределенности) из-за эффекта выборки. Для оценки реальных значений неопределенности использована симуляция возможных экспериментальных данных на основе имеющихся. Симуляция значений ${{q}_{{i,k}}}$ проведена с учетом случайных отклонений (см. Приложение). Результат расчета функций чувствительности $E\left( x \right)$ по 20 последовательным симуляциям показан на рис. 5, а.
Рис. 5.
а – симулированные зависимости контрастной чувствительности $E\left( x \right) \equiv 1{\text{/}}\kappa {\kern 1pt} '\left( x \right)$ от индекса $x = {\text{lg}}\left( a \right)$ площади стимула; б– аппроксимации симуляций различными функциями.
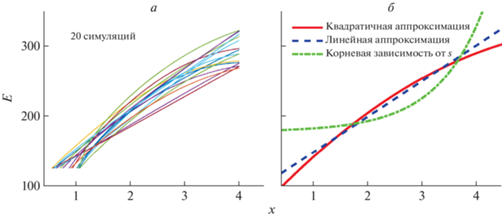
Каждая функция построена из зависимости $\kappa {\kern 1pt} '\left( x \right)$ – численного решения уравнения $W\left( {x,\kappa } \right) = 0.5$, где W найдено из решений уравнений регрессии для симулированных значений ${{q}_{{i,k}}}$. Симулированные функции $E\left( x \right)$ демонстрируют устойчивый характер, близкий к линейному по x.
Видно, что при малой площади стимула контрастная чувствительность падает, а зависимость $E\left( x \right)$ в целом близка к линейной с небольшим отрицательным вкладом квадратичности. На рис. 5, б показаны результаты аппроксимации симуляционных данных различными функциями. Использованы: квадратичная по x зависимость ${{c}_{1}} + {{c}_{2}}x + {{c}_{3}}{{x}^{2}};$ линейная по x функция ${{c}_{1}} + {{c}_{2}}x$; зависимость ${{c}_{1}} + {{c}_{2}}\sqrt {{{{10}}^{x}}} $, где ${{c}_{{1 - 3}}}$ – это константы, различные в каждом случае и найденные численно в процессе аппроксимации кривых на рис. 5, а методом Левенберга–Марквардта. Из рисунка следует, что линейная по x – логарифмическая по площади функция аппроксимирует экспериментальные данные существенно лучше, чем корневая зависимость от площади $s = 60 \times {{10}^{x}}$ стимула.
ОБСУЖДЕНИЕ
Тестирование показало (рис. 2), что пороговый контраст, различимый фотопическим зрением человека, составляет 0.004 или менее. Это пороговый контраст при яркости, близкой к 0.5, или 100 кд/м2. В процентном отношении пороговый контраст меньше 1%, что соответствует литературным данным.
В своей работе (Blackwell, 1946) автор обработал результаты 220 тысяч отдельных попыток распознавания контраста стимула девятью испытуемыми в течение четырех месяцев, что потребовало построения специализированного здания. Распознавались объекты семи размеров на фоне экрана при порядка 10 значениях его яркости.
В нашем эксперименте накоплено около 20 тысяч измерений двумя испытуемыми в течение четырех недель; использован стандартный десктоп компьютер и оригинальная программа к нему для обработки измерений. В эксперименте приняли участие всего двое испытуемых, однако для установления основного мотива связи контрастной чувствительности с размером стимула этого оказалось достаточно. Измерения проведены при одном значении средней яркости при шести задаваемых и трех действительных значениях контраста. Распознавался контраст между двумя объектами равного углового размера при пяти значениях размера, с максимальным телесным углом контрастирующего объекта приблизительно на порядок большим, чем в работе (Blackwell, 1946). Неопределенность здесь связана с невозможностью корректного сравнения: неясно, что следует считать размером контрастирующего объекта в нашем случае – размер одной прямоугольной области из двух смежных или размер обеих областей.
Различие результатов вызвано методологической разницей определения того, что есть пороговый контраст (рис. 6).
Рис. 6.
Данные по контрастной чувствительности в линейном и логарифмическом масштабах по вертикальной оси
1 – (Blackwell, 1946); 2 – линейная зависимость; 3 – результаты настоящей работы при определении контрастирующего объекта как размера одной прямоугольной области. При определении размера объекта как размера двух смежных областей кривые 2 и 3 немного смещаются вправо, на ${\text{lg}}2 = 0.3$ порядка.
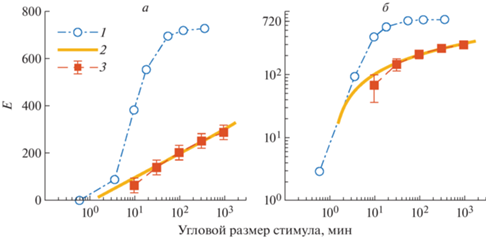
Блэквел определял пороговый контраст как разницу яркостей круга и фона при переменной яркости фона, а в нашем исследовании пороговый контраст определялся разницей яркостей двух прямоугольных полей при практически постоянном уровне средней яркости. Кроме того, в работе Блэквела не был устранен эффект лучшего распознавания контраста на резкой границе, что привело к завышению контрастной чувствительности.
В работе (Blackwell, 1946) найдено, что в области угловых размеров 1–6° (60–360') и фоновой яркости 3.4–340 кд/м2 контрастный порог и, соответственно, контрастная чувствительность не зависели от яркости фона и почти не зависели от размера объекта. В нашем эксперименте при средней яркости около 100 кд/м2 угловой размер стимула менялся до 16° (960') или 32° (1920'); при этом зависимость от размера была обнаружена. Она соответствует линейной по логарифму углового размера зависимости.
Зависимость контрастной чувствительности, приблизительно линейная по логарифму площади стимула, – главный результат настоящей работы. При увеличении площади на четыре порядка, а размера на два порядка, чувствительность возрастает всего в несколько раз: в 4 раза на рис. 6, кривые 3. Логарифмическая зависимость контрастной чувствительности E от площади стимула напоминает закон Вебера–Фехнера, согласно которому ощущение приблизительно пропорционально логарифму силы раздражителя. Отличие состоит в том, что в нашем случае это похоже на зависимость между “обратными” переменными: чувствительность тем больше, чем меньше пороговое ощущение, а рост размера стимула эквивалентен уменьшению его яркости.
В физиологии зрения закон Вебера–Фехнера означает, что в некотором диапазоне яркостей сила зрительного ощущения яркости пропорциональна логарифму яркости (Муравьева и др., 2010). В нашем случае раздражителем является контраст, но сила раздражителя зависит не только от величины контраста, но и параметрически от площади стимула. Ощущение контраста пропорционально чувствительности к контрасту – чем больше чувствительность к контрасту, тем сильнее его ощущение. Поэтому закон Вебера–Фехнера проявляется здесь косвенно, в зависимости контрастной чувствительности от логарифма углового размера стимула как параметра силы раздражителя
рис. 6, кривые 2. Видно, что при изменении углового размера стимула в $m = 100$ раз, или на два порядка, контрастная чувствительность изменяется на $\lambda ~{\text{lg}}\left( m \right) \approx 200$ единиц, или на $\lambda \approx 100$ единиц на порядок.Ранее было предположено, что число фоторецепторов сетчатки, участвующих в распознавании контраста, приблизительно пропорционально телесному углу, под которым виден стимул. Если это предположение верно, то полученная логарифмическая зависимость далеко не соответствует той проверяемой гипотезе, что чувствительность возрастает линейно с ростом числа задействованных фоторецепторов сетчатки, т.е. в зависимости от $\sqrt s $, рис. 5, б. Аппроксимации реальных данных квадратичной и корневой зависимостями имеют даже разный знак выпуклости. Линейная аппроксимация существенно ближе к измерениям, чем корневая.
Интересно оценить возможный коэффициент усиления относительного магнитного сигнала единичной радикальной пары вследствие логарифмического роста контрастной чувствительности с увеличением размера стимула. Можно ли объяснить этим эффектом наблюдаемое обнаружение некоторыми видами организмов изменений магнитного поля порядка 20–30 нТл? Как было отмечено во Введении, величина относительного магнитного сигнала радикальной пары составляет около 10–3 на 50 мкТл, или порядка 10–6 на несколько десятков нТл. Реален ли рост контрастной чувствительности (8) хотя бы на пять порядков, необходимых для правдоподобного объяснения?
На сегодня нет надежных данных о том, в каком типе фоторецепторов – палочках или колбочках – локализованы магниточувствительные криптохромы. Полагают, однако, что молекулы зрительного пигмента расположены в наружном сегменте фоторецептора из множества плотно уложенных мембранных дисков (Астахова и др., 2019), это могло бы обеспечить необходимое для магнитной чувствительности упорядоченное расположение криптохромов. Примем для грубых оценок, что на один фоторецептор приходится десять молекул криптохрома, т.е. десять радикальных пар, функционирующих синхронно, совместно “запускающих” нервный импульс. При этом достигается приблизительно троекратное ($\sqrt {10} $) начальное статистическое усиление сигнала одной радикальной пары.
При фокусном расстоянии глаза R 17 мм и плотности фоторецепторов колбочек ρ около 105 мм–2 (Curcio et al., 1990), число фоторецепторов под изображением стимула на сетчатке есть $u = \rho {{R}^{2}}~{\text{ta}}{{{\text{n}}}^{2}}\left( {0.0003{\text{\;}}\varepsilon } \right)$, где ε – угловой размер стимула в мин, или $u \approx {{\varepsilon }^{2}}$. Из рис. 6 видно, что абсолютный порог чувствительности достигается при угловом размере стимула около 1 мин и несколько меньше, согласно работе (Blackwell, 1946). Изображение такого стимула, следовательно, накрывает около 1 фоторецептора, что естественно, и распознается с чувствительностью ${{E}_{0}}$ около 10.
Изображение размером на три порядка большее, 103 мин (16º), накрывающее порядка миллиона рецепторов и соответствующее предполагаемому размеру областей измененной яркости в поле зрения, распознавалось бы с чувствительностью, увеличенной, согласно вышеприведенному соотношению приблизительно на $3{{\lambda }} = 300$ единиц, т.е. с чувствительностью $E \approx 310$, если допустить, что найденная закономерность (8) допускает экстраполяцию до малых значений размера стимула в 1 мин. Другими словами, чувствительность возрастает приблизительно в $E{\text{/}}{{E}_{0}} \approx 30$ раз. Вместе с троекратным начальным статистическим усилением общее усиление магнитного сигнала составляет около 100, т.е. всего два порядка, в то время как для надежного обнаружения магнитных сигналов требуется около пяти. Усиление могло бы оказаться несколько больше, так как оно обратно пропорционально чувствительности ${{E}_{0}}$, с которой обнаруживаются контрасты стимулов минимального размера. Однако скомпенсировать три порядка разницы за счет этого вряд ли возможно.
Наблюдаемое в настоящей работе усиление чувствительности, или снижение минимально обнаружимого контраста с ростом площади стимула, не решает проблему магнитной навигации животных, хотя и способствует решению. Для того чтобы рассмотренный механизм усиления мог рассматриваться в качестве рабочей гипотезы для объяснения магнитной чувствительности на уровне 30 нТл, надо допустить, что отдельная радикальная пара в криптохроме глаза птицы, в силу каких-то особенных условий, способна менять вероятность реакции при 50-мкТл изменении магнитного поля на 10%, а не на 0.1%. Такие реакции в магнетохимии пока не известны.
С одной стороны, пока не подтверждается, что радикально парный механизм мог бы объяснить магнитную навигацию птиц с учетом усиления контрастной чувствительности при параллельной обработке мозгом сигналов от миллионов фоторецепторов. С другой стороны, не исключено, что эволюционное совершенствование магниторецептора птицы, т.е. ее зрительного аппарата, привело не только к появлению магнитной чувствительности, но и к появлению особых алгоритмов обработки параллельных сигналов, более полно соответствующих статистической модели повышения отношения сигнал/шум в корневой зависимости от числа сигналов. Интересно было бы получить данные о способности магнитно навигирующих птиц различать контрасты стимулов при разной их площади.
ЗАКЛЮЧЕНИЕ
В настоящей работе по данным измерений установлено, что минимальная ощущаемая разница яркостей существенно зависит от размера объекта – параметра, описывающего интенсивность раздражителя. Контрастная чувствительность логарифмически зависит от углового размера контрастирующего стимула. Это соответствует общему психофизиологическому мотиву – закону Вебера–Фехнера, что не было очевидно заранее. Симуляция измерений с учетом наблюдаемого разброса значений вероятностей распознавания контрастов демонстрирует устойчивость вывода о логарифмическом характере зависимости контрастной чувствительности от размера стимула.
Результаты настоящей работы не подтверждают гипотезу, что чувствительность зрительного аппарата к яркостному контрасту может возрастать пропорционально корню из числа задействованных фоторецепторов. В то же время усиление контрастной чувствительности логарифмически с ростом размера стимула далеко не достаточно для объяснения известной чувствительности некоторых видов организмов к магнитным полям, в 1000 раз меньшим геомагнитного поля. Поэтому вопрос о том, каким образом подобная чувствительность совместима с концепцией магнитного биологического сенсора на основе механизма радикальных пар остается открытым.
Список литературы
Астахова Л.А., Ротов А.Ю., Кавокин К.В., Чернецов Н.С., Фирсов М.Л. Связь магнитного компаса и фоторецепции у птиц: гипотезы и нерешенные вопросы. Журнал общей биологии. 2019. Т. 80. № 2. С. 83–94. https://doi.org/10.1134/S0044459619020040
Муравьева С.В., Пронин С.В., Шелепин Ю.Е. Контрастная чувствительность зрительной системы человека. Экспериментальная психология. 2010. Т. 3. № 3. С. 5–20.
Домбругов Р.М. Телевидение. Киев. Вища Школа. 1979.
Красильников Н.Н., Шелепин Ю.Е. Функциональная модель зрения. Оптический журнал. 1997. Т. 64. № 2. С. 72–82.
Кучерявый А.А. Бортовые информационные системы. Ульяновск. УлГТУ. 2004.
Bertalmio M. Vision мodels for high dynamic range and wide colour gamut imaging. London: Acad. Press, 2020.
Binhi V.N., Prato F.S. Biological effects of the hypomagnetic field: An analytical review of experiments and theories. PLoS ONE. 2017. V. 12. № 6. P. e0179340. https://doi.org/10.1371/journal.pone.0179340
Binhi V.N., Prato F.S. Rotations of macromolecules affect nonspecific biological responses to magnetic fields. Scientific Reports. 2018. V. 8. № 1. P. 13495. https://doi.org/10.1038/s41598-018-31847-y
Blackwell H.R. Contrast thresholds of the human eye. J. Opt. Soc. America. 1946. V. 36. № 11. P. 624–643.
Buchachenko A. Magneto-biology and medicine. New York. Nova Science. 2014.
Crumey A. Human contrast threshold and astronomical visibility. Monthly Notices of the Royal Astronomical Society. 2014. V. 442. № 3. P. 2600–2619. https://doi.org/10.1093/mnras/stu992
Curcio C.A., Sloan K.R., Kalina R.E., Hendrickson A.E. Human photoreceptor topography. The Journal of Comparative Neurology. 1990. V. 292. № 4. P. 497–523.
Hore P.J., Mouritsen H. The radical-pair mechanism of magnetoreception. Annual Review of Biophysics. 2016. V. 45. № 1. P. 299–344. https://doi.org/10.1146/annurev-biophys-032116-094545
Lohmann K.J., Lohmann C.M., Ehrhart L.M., Bagley D.A., Swing T. Animal behaviour: geomagnetic map used in sea-turtle navigation. Nature. 2004. V. 428. № 6986. P. 909–910. https://doi.org/10.1038/428909a
Nadler M.P., Miller D., Nadler D.J. (Eds). Glare and contrast sensitivity for clinicians. New York: Springer-Verlag, 1990.
Pelli D.G., Bex P. Measuring contrast sensitivity. Vision Research. 2013. V. 90. P. 10–14. https://doi.org/10.1016/j.visres.2013.04.015
Pishchalnikov R.Y., Gurfinkel Y.I., Sarimov R.M., Vasin A.L., Sasonko M.L., Matveeva T.A., Binhi V.N., Baranov M.V. Cardiovascular response as a marker of environmental stress caused by variations in geomagnetic field and local weather. Biomedical Signal Processing and Control. 2019. V. 51. P. 401–410. https://doi.org/10.1016/j.bspc.2019.03.005
Riccó, A. Relazione fra il minimo angolo visuale e l’intensitГ luminosa. Memorie della Societa Degli Spettroscopisti Italiani. 1877. V. 6. P. B29–B58.
Schulten K., Swenberg C., Weller A. A biomagnetic sensory mechanism based on magnetic field modulated coherent electron spin motion. Zeitschrift fur Physikalische Chemie. 1978. V. 111. № 1. P. 1–5.
Stöckl A.L., O’Carroll D.C., Warrant E.J. Neural summation in the hawkmoth visual system extends the limits of vision in dim light. Current Biology. 2016. V. 26. № 6. P. 821–826. https://doi.org/10.1016/j.cub.2016.01.030
Wan G., Hayden A.N., Iiams S.E., Merlin C. Cryptochrome 1 mediates light-dependent inclination magnetosensing in monarch butterflies. Nature Communications. 2021. V. 12. № 1. P. 771. https://doi.org/10.1038/s41467-021-21002-z
Watson A.B., Ahumada A.J. A standard model for foveal detection of spatial contrast. Journal of Vision. 2005. V. 5. № 9. P. 717–740. https://doi.org/10.1167/5.9.6
Weisstein E.W. Beta Binomial Distribution. From MathWorld – A Wolfram Web Resource. URL: https://mathworld.wolfram.com/BetaBinomialDistribution.html (accessed 2022.07.18).
Дополнительные материалы отсутствуют.
Инструменты
Сенсорные системы