

Сенсорные системы, 2023, T. 37, № 1, стр. 60-64
“Акустическое окно” на нижней челюсти дельфина: оценка толщины кости
Е. В. Сысуева 1, *, В. В. Попов 1
1 Институт проблем экологии и эволюции им. А.Н. Северцова РАН
119071 Москва, Ленинский проспект, д. 33, Россия
* E-mail: evgeniasysueva@gmail.com
Поступила в редакцию 03.10.2022
После доработки 21.10.2022
Принята к публикации 04.11.2022
- EDN: AUDIQM
- DOI: 10.31857/S0235009223010079
Аннотация
Нижнечелюстная гипотеза звукопроведения у дельфинов предполагает, что произошло функциональное замещение слуховой раковины “акустическим окном” на нижней челюсти. При помощи промышленной томографии были построены 3D модели нижних челюстей шести видов зубатых китов (Odontoceti). Анализ толщины кости в области предполагаемого “акустического окна” показал, костная стенка связана с размером животного. Область наибольшего истончения внешней стенки обнаружилась в верхней части проксимального участка нижней челюсти или рядом с ее нижним углом, а не по центру, как предполагалось ранее.
ВВЕДЕНИЕ
Нижняя челюсть зубатых китов (Odontoceti) выполняет не только функцию захвата и удержания добычи, но и участвует в процессах периферического слуха. Челюсти разных видов имеют схожее строение. Различия наблюдаются в ростральной части и связаны, главным образом, с особенностями добычи пропитания. В отличие от аналогичных структур наземных млекопитающих, нижняя челюсть зубатых китов полая внутри и имеет нижнечелюстное отверстие с внутренней стороны (рис. 1).
Рис. 1.
Нижние челюсти наземного и водного млекопитающих.
а – пятнистый олень, Cervus nippon; б – амазонский дельфин, Inia geoffrensis. (По материалам сайта зоомузея МГУ).

В полости заключен жировой тяж, для которого нижнечелюстное отверстие служит выходом в мягкие ткани (Nummela et al., 2007). Жировой тяж разрастается дистально вплоть до соединения со средним ухом. Он играет роль звукопроводящего канала.
Зубатые киты лишены ушной раковины, и до сих пор нет единого мнения о том, где находится место звукоприема на поверхности головы этих животных (акустическое окно).
Современные технологии исследований позволяют подвергнуть критической оценке имеющиеся гипотезы о механизмах звукопроведения у зубатых китов. Основная и наиболее популярная гипотеза – нижнечелюстная, обоснованная в серии работ конца 70-х–начала 80-х годов прошлого века (Norris, 1968, 1969; Norris, Harvey, 1974). Согласно гипотезе, участок (“акустическое окно”), расположенный на внешней стороне нижней челюсти в области ее расширения и максимального истончения кости (“panbone”), участвует в передаче звуковых колебаний из внешней водной среды в жировой тяж внутри нижней челюсти и далее в структуры среднего и внутреннего уха.
Предметом настоящего исследования была область максимального истончения костной стенки нижней челюсти (panbone). Гипотеза не указывает ее точного расположения, но предполагает, что она свойственна для всего подотряда зубатых китов. В гипотезе не уточняется, как различается толщина panbone у разных видов, хотя в подотряд входят и такие крупные животные, как косатки (до 10 м) и относительно миниатюрные морские свиньи (1.5–2 м). Также открытым остается вопрос, почему возможное утолщение panbone с увеличением размера тела не ухудшает передачу звука в жировой тяж. Следует отметить, что диапазон воспринимаемых частот у всех зубатых китов, независимо от размеров тела, примерно одинаков.
Все эти факты побудили нас дополнить имеющиеся данные по морфологии нижней челюсти зубатых китов и определили следующие задачи исследования: определить минимальную толщину кости в предполагаемой области panbone у разных видов; определить расположение наиболее “тонких” участков у разных видов; показать зависимость между толщиной panbone и размером тела зубатого кита.
МЕТОДИКА
Экземпляры нижних челюстей зубатых китов были отобраны из морфологических коллекций Института проблем экологии и эволюции им. А.Н. Северцова и Утришской морской станции (рис. 2).
Рис. 2.
Нижние челюсти зубатых китов, отобранные для исследования.
Вверху – ветви нижних челюстей морской свиньи Phocoena phocoena; внизу – то же для белухи Delphinapterus leucas.
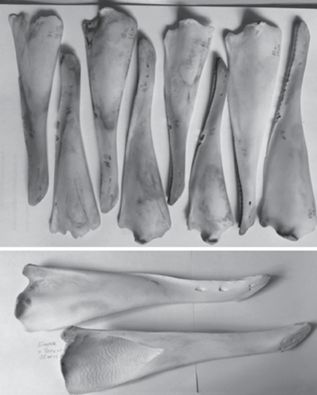
Для большинства образцов, к сожалению, не известны пол и возраст. Всего для исследования удалось получить 17 образцов нижней челюсти от 11 особей шести видов зубатых китов: морской свиньи Phocoena phocoena; иравадийского дельфина Orcaella brevirostris; обыкновенного дельфина Delphinus delphis; бутылконосого дельфина Tursiops truncatus; белухи Delphinapterus leucas и косатки Orcinus orca.
Все отобранные образцы сканированы на 3D промышленном томографе с разрешением 100 мкм. Полученные 3D реконструкции обработаны в программе VGStudio Max 3.5.1. Для характеристики толщины стенок был применен сферический анализ (sphere analysis), принцип которого состоял в том, что между границами материала вписывается сфера и определяется ее диаметр.
РЕЗУЛЬТАТЫ
Получены 3D реконструкции 17 челюстей от 11 особей шести видов зубатых китов (табл. 1): морской свиньи (шесть образцов от четырех особей); иравадийского дельфина (два образца от одной особи); обыкновенного дельфина (один образец от одной особи); бутылконосого дельфина (три образца от двух особей); белухи (три образца от двух особей) и косатки (два образца от одной особи).
Таблица 1.
Сводная таблица по полученным данным
| Вид | Средний размер тела (м) (по Carwardine, 2000) | № образца | Минимальная толщина, мм | Расположение: 1– верхние части проксимального участка нижней челюсти; 2 – угол нижней челюсти |
|---|---|---|---|---|
| Orcinus orca | 5.5–9.8 | 1 (левая ветвь) | 1.37 | 2 |
| 1 (правая ветвь) | 1.70 | 2 | ||
| Delphinapterus leucas | 3–5 | 1 (левая ветвь) | 0.56 | 1 |
| 1 (правая ветвь) | 0.58 | 2 | ||
| 2 (левая ветвь) | 0.45 | 1 | ||
| Tursiops truncatus | 1.9–3.9 | 1 (левая ветвь) | 0.76 | 2 |
| 2 (левая ветвь) | 0.46 | 2 | ||
| 2 (правая ветвь) | 0.46 | 2 | ||
| Orcaella brevirostris | 2.1–2.6 | 1 (левая ветвь) | 0.32 | 2 |
| 1 (правая ветвь) | 0.35 | 2 | ||
| Delphinus delphis | 1.7–2.4 | 1 (правая ветвь) | 0.46 | 2 |
| Phocoena phcoena | 1.4–1.9 | 1 (левая ветвь) | 0.29 | 1 |
| 1 (правая ветвь) | 0.29 | 1 | ||
| 2 (левая ветвь) | 0.25 | 2 | ||
| 2 (правая ветвь) | 0.32 | 1 | ||
| 3 (левая ветвь) | 0.36 | 2 | ||
| 4 (правая ветвь) | 0.34 | 2 |
Из полученных данных (рис. 3) видно, что для большинства образцов толщина костной стенки увеличивается с увеличением размера тела.
Рис. 3.
Примеры результатов сферического анализа для 3D моделей нижних челюстей и выборочные значения толщины костной стенки.

У самых маленьких представителей подотряда зубатых китов – морских свиней – минимальная толщина внешней стенки составила от 0.25 до 0.36 мм, у представителей видов средних размеров – иравадийского дельфина, обыкновенного дельфина и бутылконосого дельфина – от 0.32 до 0.46 мм; крупные виды, такие как белуха и косатка, имели толщину кости от 0.45 до 1.7 мм. Исключением был образец челюсти бутылконосого дельфина, у которого толщина костной стенки превышала таковую у других образцов того же вида, что могло быть связано с возрастом, полом и размером конкретной особи (табл. 1).
Определено расположение истонченных участков в области panbone. Наибольшего истончения внешняя стенка достигала в верхней части проксимального участка нижней челюсти или рядом с нижним углом нижней челюсти.
ОБСУЖДЕНИЕ
В ходе проведенного исследования были получены дополнительные данные по морфологии нижней челюсти шести видов зубатых китов. Данные по толщине латеральной стенки нижней челюсти китообразных можно найти только в работах одного коллектива (Nummela et al., 2004; 2007). Методами классической морфометрии ими были получены данные для 30 видов зубатых китов, в том числе и для пяти из тех шести видов, что изучали мы: для косатки – 5.48 мм; белухи – 1.84 мм; бутылконосого дельфина – 2.03 мм; обыкновенного дельфина – 1.14 мм; морской свиньи – 0.57 мм. Эти значения больше, чем полученные нами, но также коррелируют с размером тела. Однако эти данные трудно напрямую сравнить с нашими результатами. В работе (Nummela et al., 2007) минимальная толщина предполагаемой области panbone определялась для всех видов лишь в одном месте сечения. Сечение проходило вертикально через 3/4 от размера нижнечелюстного отверстия, отложенных от каудального конца нижней челюсти. Сечение с большой вероятностью не захватывало все тонкие участки области panbone, но соответствовало акустическому окну. Кроме того, существенным для сравнения является факт неопределенности возраста образцов и связанных с ним размеров и толщины челюсти.
Нами был использован метод, позволяющий увидеть все перепады толщин на латеральной поверхности нижней челюсти, что позволило составить полное представление о местонахождении истонченных участков. Так, области наибольшего истончения внешней стенки нижней челюсти у исследованных образцов располагались преимущественно в верхней части проксимального участка нижней челюсти или рядом с нижним углом нижней челюсти, а не по центру нижнечелюстного отверстия, как предполагалось ранее.
В своей работе (Norris, 1968) автор предполагал, что несмотря на то, что по своим акустическим свойствам кость отличается от тканей тела и воды и поэтому может быть препятствием при прохождении звука через костную пластину нижней челюсти в жировой тяж, тонкая кость (panbone) путем вибрации служит проведению звука из внешней среды. В данной работе выявлена тенденция утолщения тонких участков латеральной стенки нижней челюсти с увеличением размера тела: чем больше размер тела у данного вида, тем больше толщина кости. Это означает, что увеличение толщины костной стенки в области “акустического окна” может привести к ухудшению проникновения звука в жировой тяж: крупные представители подотряда должны слышать хуже, но этого не происходит. Например, слух у косатки по чувствительности не уступает слуху мелких зубатых китов (Szymanski et al., 1999). Возможно, это один из нюансов, который говорит нам о том, что нет одной единственной рабочей гипотезы звукопроведения и звук способен проникать к акустическим структурам в голове дельфина разными путями.
Какое-то время назад нижнечелюстная гипотеза была доработана. По современным представлениям, местом первичного вхождения звука являются мягкие ткани области глотки дельфина (Cranford et al., 2008). Звук, не встречая никаких препятствий на пути, проникает в жировой тяж и далее через медиальную сторону нижней челюсти. Возможность работы “акустического окна” допускается для проведения высоких частот при определенном угле падения.
ЗАКЛЮЧЕНИЕ
Данные настоящего исследования получены на единичных образцах; тем не менее, они имеют значение. Факты о расположении тонких участков на нижней челюсти нужно принять во внимание при проведении исследований по методу контактной акустической стимуляции для корректной установки излучателя звука на челюсти зубатых китов, а также при интерпретации полученных данных.
Работа поддержана РНФ (грант № 22-25-00025).
Список литературы отсутствует.
Дополнительные материалы отсутствуют.
Инструменты
Сенсорные системы