

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 1, стр. 41-64
Биофациальный анализ нижне- и среднеюрских отложений Сибири по палиноморфам
1 Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН
Новосибирск, Россия
2 Новосибирский национальный исследовательский государственный университет
Новосибирск, Россия
* E-mail: GoryachevaAA@ipgg.sbras.ru
Поступила в редакцию 26.02.2018
После доработки 22.03.2018
Принята к публикации 25.10.2018
Аннотация
Представлены результаты биофациального анализа нижне-среднеюрских отложений Западной и Восточной Сибири. Проведено комплексное изучение различных групп палиноморф (спор и пыльцы наземных растений, цист динофлагеллат, акритарх, празинофитов, зигнемовых водорослей и др.) и на основании детального анализа распределения отдельных таксонов и количественных соотношений различных групп микрофитофоссилий в разнофациальных разрезах Западной и Восточной Сибири установлены определенные комбинации палиноморф, отражающие различные условия обитания. Полученные данные позволили выделить девять типов эколого-фациальных ассоциаций, которые являются показателями разных обстановок – от континентальных, прибрежно-морских, мелководных до типично морских глубоководных.
ВВЕДЕНИЕ
Настоящая работа является продолжением исследований, посвященных разностороннему изучению нижне-среднеюрских отложений Сибири (Горячева, 2008, 2011, 2017; Горячева, Никитенко, 2016; Никитенко и др., 2013, 2017, 2018 и др.). В ней представлены результаты изучения юрских палиноморф с точки зрения их потенциала для палеоэкологических реконструкций. Для нижней и средней юры подобная работа выполнена впервые, однако методическая основа была разработана ранее Н.К. Лебедевой на примере верхнемеловых отложений Сибири (Лебедева, 2008, 2010). Существуют разные точки зрения на степень корректности палеоэкологических и палеогеографических реконструкций, выполненных на основе анализа распределения только палиноморф. В первую очередь это обусловлено большим количеством разнообразных факторов, которые могли на них повлиять как в море, так и на континенте. Безусловно, совместные палеонтологические, седиментологические, геохимические и др. исследования дают наиболее исчерпывающую информацию об обстановках, которые существовали в тот или иной отрезок времени. Однако часто при изучении кернового материала из разнофациальных толщ с закрытых территорий оказываются доступны сведения только по микрофитофоссилиям. В таких случаях палинологические данные предоставляют единственную возможность для восстановления условий осадконакопления вмещающих пород, информация о которых необходима в том числе для уточнения и детализации схем фациального районирования нижней и средней юры Сибири. Все вышеперечисленное определяет актуальность проведенных исследований и полученных данных.
МАТЕРИАЛ И МЕТОДЫ
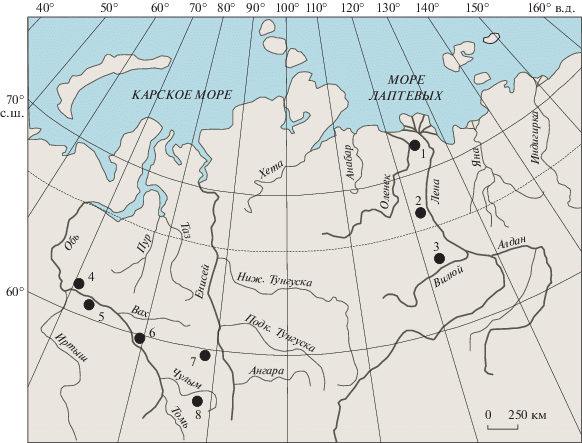
Материалом для статьи послужила коллекция из 514 образцов из естественных обнажений и скважин, собранная автором и предоставленная в разные годы Б.Л. Никитенко, В.П. Девятовым, Л.Г. Вакуленко и В.И. Ильиной по Западной Сибири (обнажения 14 и 15 на р. Золотой Китат и скважины Лульяхская 5, Северо-Правдинская 521, Западно-Тымская 1, Восток 4) и Восточной Сибири (обнажения 5, 6, 7, 14, 16 на р. Келимяр и скважины Средне-Накынская 360, Ыгыатто-Тюнгская 1) (рис. 1).
Рис. 1.
Схема местонахождения изученных разрезов. 1 – р. Келимяр, обн. 5, 6, 7, 14, 16; 2 – скв. Ыгыатто-Тюнгская 1; 3 – скв. Средне-Накынская 360; 4 – скв. Северо-Правдинская 521; 5 – скв. Лульяхская 5; 6 – скв. Западно-Тымская 1; 7 – скв. Восток 4; 8 – р. Золотой Китат, обн. 14 и 15.
Биостратиграфия и описание вышеуказанных разрезов опубликованы ранее (Вдовин, Ильина, 1967; Князев и др., 2003; Ян и др., 2006; Никитенко, 2009; Девятов и др., 2010; Вакуленко и др., 2010; Горячева, 2006, 2008, 2011, 2017; Горячева, Никитенко, 2016). Все существующие геологические данные (палеонтологические, литологические и др.) по исследованным разрезам были привлечены в настоящей статье при выполнении биофациального анализа.
Обработку образцов после предварительного очищения и опробования соляной кислотой на карбонатность выполняли сначала плавиковой кислотой, а затем пирофосфатом калия, с последующим разделением органической и минеральной частей осадка в тяжелой кадмиевой жидкости с удельным весом 2.25 путем центрифугирования.
Палинологический анализ включал изучение как спор и пыльцы наземных растений, так и водных одноклеточных микрофитофоссилий: цист динофлагеллат (отдел Dinoflagellata, подотдел Dinokaryota, класс Dinophyceae), акритарх (группа Acritarcha неясной систематической принадлежности), празинофитов (отдел Chlorophyta, класс Prasinophyceae), зеленых водорослей, близких к зигнемовым (отдел Chlorophyta, класс Zygnematophyceae) и колониальных водорослей (отдел Chlorophyta, класс Chlorophyceae).
Для статистической обработки палиноспектров и их интерпретации подсчитывали не менее 200–300 зерен, за 100% принимали всю совокупность микрофитофоссилий. Процентное содержание каждого таксона рассчитывали от общего количества.
Для всех изученных разрезов были построены диаграммы количественного содержания спор, пыльцы и разных групп микрофитопланктона. На основе анализа их распределения по разрезу и соотношениям морских (цисты динофлагеллат, акритархи, празинофиты) и наземных (споры и пыльца, пресноводные водоросли) микрофоссилий было выявлено несколько типов ассоциаций с достаточно устойчивым сочетанием определенных групп палиноморф, отражающих обстановки от континентальных до типично морских.
Методика выделения и описание типов эколого-фациальных ассоциаций палиноморф
Пространственное распределение микрофитопланктона в отложениях морских бассейнов древних геологических эпох во многом сходно с тем, что происходит в современных условиях. Зависимость распределения ископаемого микрофитопланктона от многих факторов, в том числе таких, как соленость, удаленность от берега и др., дает возможность использовать его как показатель условий обитания. Таксономический состав и количественное содержание микрофитопланктона в одновозрастных разнофациальных осадках, проявляющих прямую зависимость от условий седиментации, могут быть привлечены как один из критериев для палеогеографических реконструкций.
Эколого-фациальная приуроченность микрофитопланктона была выявлена, во-первых, на основании анализа распределения палиноморф в тех разрезах, в которых условия формирования осадков уже восстановлены по палеонтологическим, литологическим и другим данным, и во-вторых, на основании сведений по экологии современных одноклеточных водорослей (Hulburt, 1963; Яшнов, 1965; Boalch, Parke, 1971; Harrison et al., 1977; Wall et al., 1977; Tappan, 1980; Bujak, Devies, 1983; Goodman, 1987; Sarjeant et al., 1987; Taylor, 1987; Harland, 1988; Водоросли…, 1989; Prauss, Rigel, 1989; Yoo, 1991; Hinga, 1992; Fensome et al., 1993; Martin, 1993; Hoek et al., 1995; Batten, Grenfell, 1996; Dale, 1996; Geel, Grenfell, 1996; Guy-Ohlson, 1996; Mudie, Harland, 1996; Strother, 1996; Zonneveld et al., 2001; Marret, Zonneveld, 2003 и др.).
Использование существующих сведений по экологии современных диноцист; зигнемовых водорослей, к которым близки ископаемые формы Schizosporis и Ovoidites; празинофитов сем. Tasmanitaceae и колониальных водорослей Botryococcus помогло восстановить возможные условия обитания растений, продуцирующих ископаемые палиноморфы, а соотношение преимущественно морских микрофоссилий (цисты динофлагеллат, акритархи, празинофиты) и палиноморф наземного происхождения (споры и пыльца) позволило реконструировать относительное положение береговой линии. Экологическая приуроченность таксонов, которые известны только как ископаемые, устанавливалась с привлечением литературных данных и данных по другим совместно встречающимся группам фоссилий (фораминиферы, аммониты, двустворки, белемниты).
В результате детального анализа распределения отдельных таксонов и количественных соотношений различных групп микрофитофоссилий в разрезах Западной и Восточной Сибири автором были установлены определенные комбинации палиноморф, которые отражают разные условия обитания. Полученные данные позволили выделить девять типов эколого-фациальных ассоциаций (рис. 2). Под ассоциацией понимается совокупность палиноспектров с достаточно устойчивым сочетанием определенных групп палиноморф. Количественные соотношения этих групп могут меняться в некоторых пределах.
Рис. 2.
Структура типов эколого-фациальных ассоциаций палиноморф и условная схема их распределения на модельном профиле от континентальных до типично морских обстановок. 1 – споры и пыльца наземных растений; 2 – диноцисты; 3 – акритархи; 4 – празинофиты; 5 – зигнемовые водоросли; 6 – колониальные зеленые водоросли; 7 – формы неясной систематической принадлежности.
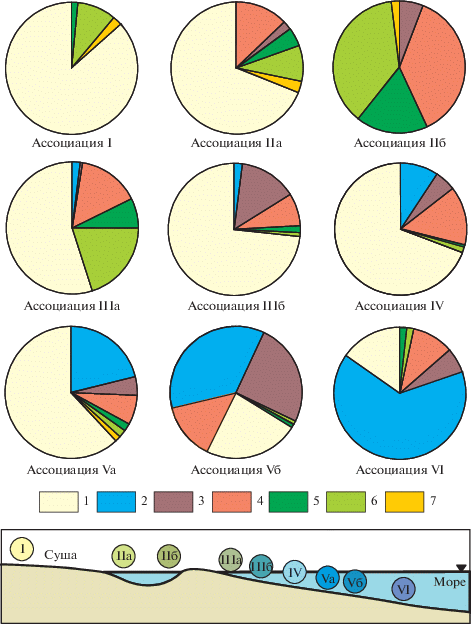
В результате, с учетом всей существующей информации по палеонтологии, геологии и палеогеографии Сибири, выделенные типы ассоциаций были выстроены на идеализированном модельном профиле по увеличению степени мористости, от континентальных до типично морских условий (рис. 2).
Ниже дана характеристика выделенных типов эколого-фациальных ассоциаций палиноморф:
I. Споры и пыльца наземных растений доминируют (часто до 100%). Микрофитопланктон представлен зигнемовыми водорослями (2–3%), колониальными зелеными водорослями (иногда до 65%), формами неясного систематического положения – Inaperturopollenites и Aletes striatus (до 15%).
Породы: песчаники с углефицированным растительным детритом (УРД), алевролиты с отпечатками макрофлоры, редко глины.
IIa. Споры и пыльца доминируют (55–85%). Микрофитопланктон: акритархи – единично, празинофиты – 5–40%, зигнемовые водоросли – 3–15%, колониальные водоросли – 5–20%, формы неясной систематической принадлежности – до 10%.
Породы: песчаники с УРД, алевролиты, переслаивание алевролитов и аргиллитов, редко глины.
IIб. Отсутствуют споры и пыльца наземных растений. Микрофитопланктон: акритархи – 7.5%, празинофиты – 36%, зигнемовые водоросли – 17%, колониальные зеленые водоросли – 35%, формы неясной систематической принадлежности – 4.5%.
Порода: известняки.
IIIa. Споры и пыльца, как правило, доминируют (от 30 до 90%). Микрофитопланктон: диноцисты – до 1–3%, акритархи – 0.5–3%, празинофиты – до 15%, зигнемовые водоросли – 0.5–9%, колониальные водоросли – до 40% (редко больше), формы неясной систематической принадлежности – до 1%.
Породы: алевролиты, глинистые алевролиты с прослоями песчаника, редко песчаники с УРД; фауна: двустворки, фораминиферы.
IIIб. Споры и пыльца преобладают (60–97%), диноцисты – 2–10%, акритархи – до 30%, празинофиты – до 18%, зигнемовые водоросли – 0.5–1%, колониальные зеленые водоросли – до 1%.
Породы: аргиллиты, алевролиты, алевритистые глины; фауна: двустворки, фораминиферы.
IV. Споры и пыльца – от 40 до 90%. Микрофитопланктон: диноцисты – до 14%, акритархи – до 10% (редко до 50%), празинофиты – до 30%, зигнемовые водоросли – до 3%, колониальные зеленые водоросли – до 4%, формы неясной систематической принадлежности – до 1%.
Породы: переслаивание аргиллитов и алевролитов с редкими прослоями глинистых песчаников; фауна: фораминиферы.
Va. Споры и пыльца – 40–65%. Микрофитопланктон: диноцисты – до 50%, акритархи – до 9%, празинофиты – до 50%, зигнемовые – до 3%, колониальные зеленые водоросли – до 5%, формы неясного систематического положения – до 1.5%.
Породы: аргиллиты, в том числе слабо сидеритизированные, алевритистые глины, алевриты; фауна: аммониты, двустворки, белемниты, фораминиферы.
Vб. Споры и пыльца наземных растений до 50%. Микрофитопланктон: диноцисты – до 65%, акритархи – до 47%, празинофиты – до 80%, колониальные зеленые водоросли – 0.5–1.5%.
Породы: глины, алевритистые глины; фауна: аммониты, двустворки, фораминиферы.
VI. Споры и пыльца – до 50%. Микрофитопланктон: диноцисты – до 80–90%, акритархи – до 30%, празинофиты от 3 до 20%. Возможно присутствие единичных зигнемовых и колониальных зеленых водорослей.
Породы: аргиллиты, глины, алевритистые глины; фауна: белемниты, аммониты, двустворки, гастроподы.
Далее приведен биофациальный анализ разрезов Западной и Восточной Сибири на основе вышеописанных типов эколого-фациальных ассоциаций палиноморф и закономерностей распределения различных групп микрофитофоссилий.
ЭКОЛОГО-ФАЦИАЛЬНЫЕ АССОЦИАЦИИ ПАЛИНОМОРФ В ИЗУЧЕННЫХ РАЗРЕЗАХ ВОСТОЧНОЙ СИБИРИ
Обнажение 16, р. Келимяр
В отложениях нижней юры, представленных глинистыми породами кыринской и келимярской свит, выделяются три ассоциации – Va, Vб и VI (рис. 3).
Рис. 3.
Типы эколого-фациальных ассоциаций палиноморф в обн. 16 на р. Келимяр. 1 – глины; 2 – глины битуминозные; 3 – глины алевритистые; 4 – глины алевритовые; 5а – песок, песчаник, 5б – гравелит; 6а – галька, 6б – валуны; 7 – конкреционные прослои; 8 – включения, конкреции: а – сидеритовые известняки, б – фосфориты, в – пирит.
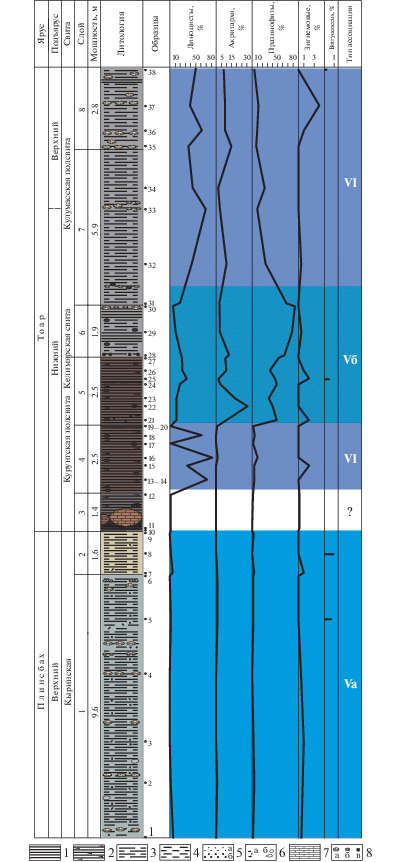
Ассоциация Va (слои 1–2, обр. 1–9, верхний плинсбах) установлена в алевритистых глинах. Фауна: фораминиферы, остракоды, остатки аммонитов, двустворок, белемнитов, гастропод, серпул и ракообразных.
В общем составе микрофоссилий преобладают споры и пыльца наземных растений (до 95%).
В составе микрофитопланктона диноцисты представлены одним родом Nannoceratopsis (0.3–5.7%). Для ассоциации характерно присутствие нанноцератопсисов как с двумя антапикальными рогами (Nannoceratopsis deflandrei Evitt), развитие которых происходило в умеренно мелководной зоне моря с довольно активным гидродинамическим режимом (Evitt, 1961), так и c одним антапикальным рогом (Nannoceratopsis senex van Helden), которые приурочены к умеренно глубоководным, но достаточно удаленным от берега обстановкам (Evitt, 1961; Ильина и др., 1994; Никитенко, Поспелова, 1996 и др.). Диноцисты с одним удлиненным антапикальным рогом Nannoceratopsis deflandrei anabarensis Iljina характерны для фотической зоны бассейна с усиленной гидродинамикой (Ильина и др., 1994). Количество акритарх варьирует от 0.6 до 1.2%, празинофитов – от 0.3 до 4%. Встречены в очень малых количествах зигнемовые (0.4–1%) и редкие сегменты колониальных водорослей Botryococcus (0.4–1%).
Таким образом, для ассоциации характерно преобладание спор и пыльцы наземной растительности и присутствие в небольших количествах диноцист, акритарх, празинофитов и единичных зигнемовых. В целом фациальную обстановку можно определить как относительно мелководную зону моря с нестабильным солевым режимом и активной гидродинамикой.
По фораминиферам в самой нижней части слоя 1 палеообстановка охарактеризована как внутренняя часть средней сублиторали, в остальной части слоя 1 – как внешняя часть верхней сублиторали, а в слое 2 – как внутренняя часть верхней сублиторали (Никитенко и др., 2011).
Ассоциация VI (слой 4, обр. 13–20, курунгская подсвита, келимярская свита, нижний тоар) установлена в сланцеватых глинах. Фауна: фораминиферы.
В общем составе палиноморф преобладает микрофитопланктон. Доля наземной составляющей от 5 до 40%. Значительно увеличилась роль диноцист (12–80%) с доминированием Nannoceratopsis senex, который, имея овальную форму с одним коротким антапикальным рогом, свободно парил в фотической зоне моря, достигая максимального развития в хорошо прогретых приповерхностных водах морского бассейна с небольшими глубинами на удалении от зоны прибоя (Ильина и др., 1994; Никитенко, Поспелова, 1996). Акритархи представлены редкими экземплярами, на фоне постоянного присутствия в небольшом количестве празинофитов (3–10%) и единичных зигнемовых (до 1%). Можно сделать вывод о формировании осадков в достаточно удаленной от берега части моря.
По фораминиферам палеообстановка охарактеризована как средняя сублитораль (Никитенко и др., 2011).
Ассоциация Vб (слои 5, 6 и самый низ сл. 7, обр. 21–31, келимярская свита, нижний тоар) установлена в алевритистых глинах. Фауна: аммониты, белемниты, фораминиферы.
Споры и пыльца наземных растений составляют от 7 до 40% от общего количества микрофоссилий. Преобладает микрофитопланктон (60–93%). Для этой ассоциации характерно разнообразие диноцист, представленных восемью родами. В количественном отношении диноцисты составляют от 12 до 32%.
Количество празинофитов Tasmanites и Leiosphaeridia от 35 до 83%. Акритархи составляют от 2 до 30%. Редко встречаются зигнемовые (до 2%) и единичные колониальные ботриококкусы (до 1%).
Таким образом, уменьшение количества, но увеличение разнообразия диноцист, присутствие акритарх, большое количество празинофитов и при этом присутствие единичных зигнемовых и ботриококкусов могут свидетельствовать об обстановках, характерных для неглубоководного и, возможно, слабоаэрируемого морского бассейна.
По фораминиферам установлено чередование обстановок внешней части верхней сублиторали (верхняя часть слоя 4–нижняя часть слоя 5), внутренней части средней сублиторали (верхняя часть слоя 5), внешней части верхней сублиторали (самый верх слоя 5–самый низ слоя 6), внутренней части средней сублиторали (средняя часть слоя 6) и внешней части верхней сублиторали (верхняя часть слоя 6–нижняя часть слоя 7) (Никитенко и др., 2011).
Ассоциация VI (слои 7–8, обр. 32–38, келимярская свита, нижний–верхний тоар) установлена в алевритистых глинах. Характерно преобладание микрофитопланктона (до 90%) над наземной растительностью. Количество празинофитов сокращается вверх по разрезу от 25–26 до 5%. Постоянно присутствуют в небольших количествах акритархи (2–15%). В составе микрофитопланктона отмечаются единичные зигнемовые (1–4%) и ботриококкусы (до 1%).
В ассоциации происходит смена доминирования бореально-атлантических диноцист Nannoceratopsis на бореально-арктическую перидиниоидную группу, представленную Phallocysta и Valvaeodinium, что, вероятно, объясняется начавшимся в конце раннего тоара и в позднем тоаре похолоданием климата и приуроченностью этих форм к богатым питательными веществами участкам относительного глубоководья (Prauss, 1989; Ильина и др., 1994; Никитенко, Поспелова, 1996).
В целом количественная представительность диноцист, уменьшение роли празинофитов и наличие акритарх могут свидетельствовать об умеренно глубоководной зоне моря с нормальным солевым и газовым режимом.
По фораминиферам установлено чередование двух обстановок: внешней части верхней сублиторали (нижняя часть слоя 7 и самый верх слоя 8) и внутренней части средней сублиторали (верхняя часть слоя 7 и большая часть слоя 8) (Никитенко и др., 2011).
Обнажение 5 (канавы 1, 2), р. Келимяр
В отложениях нижней юры, представленных глинистыми породами кыринской и келимярской свит, выделяются три ассоциации – Va, Vб и VI (рис. 4, 5).
Рис. 4.
Типы эколого-фациальных ассоциаций палиноморф в обн. 5 (канава 1) на р. Келимяр. Условные обозначения см. на рис. 3.
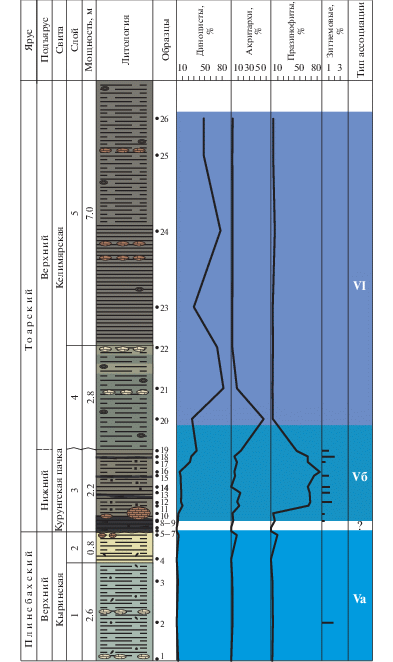
Рис. 5.
Типы эколого-фациальных ассоциаций палиноморф в обн. 5 (канава 2) на р. Келимяр. Условные обозначения см. на рис. 3.
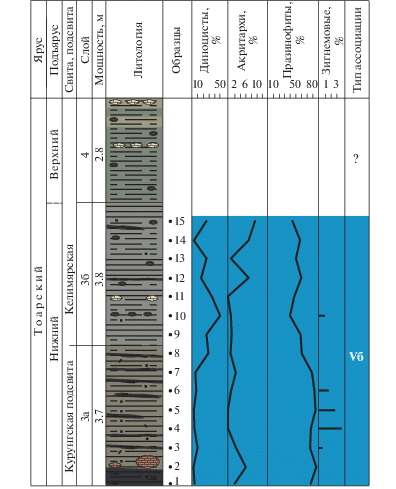
Ассоциация Va (слои 1, 2, обр. 1–5, кыринская свита, верхний плинсбах) установлена в алевритистых глинах и алевритах. Фауна: фораминиферы, остракоды.
В общем составе микрофоссилий преобладают споры и пыльца наземных растений.
Диноцисты представлены Nannoceratopsis (N. deflandrei, N. senex) и Mendicodinium. Количество акритарх не более 8%, празинофитов до 9%. Единичные зигнемовые присутствуют только в нижней части слоя 1.
Таким образом, для ассоциации характерно преобладание спор и пыльцы наземных растений и присутствие в небольших количествах диноцист, акритарх, празинофитов и зигнемовых. Подобный состав микрофитопланктона характеризует фациальную обстановку как приближенную к берегу довольно мелководную зону моря.
По фораминиферам палеообстановка охарактеризована как внутренняя часть верхней сублиторали (слой 2) (Комплексное…, 2010).
Ассоциация Vб (канава 1, слой 3, обр. 8–19 и канава 2, обр. 1–15, келимярская свита, нижний тоар) установлена в глинах. Фауна: фораминиферы.
В общем составе преобладает микрофитопланктон, а споры и пыльца составляют от 2 до 28%. Для ассоциации характерно разнообразие диноцист, представленных несколькими родами. В количественном отношении диноцисты составляют от 5 до 50% с тенденцией к увеличению их содержания вверх по разрезу. В целом среди микрофитопланктона доминируют празинофиты (44–88%). Акритархи составляют от 3.5 до 15%. Единично встречаются зигнемовые (до 3%).
Таким образом, небольшое количество диноцист, присутствие акритарх и очень большое количество празинофитов, при участии единичных зигнемовых, могут свидетельствовать об обстановках, характерных для достаточно мелководного морского бассейна, возможно плохо аэрируемого.
По фораминиферам палеообстановка определена как внешняя часть верхней сублиторали (слой 3) (Комплексное…, 2010).
Ассоциация VI (слои 4–5, обр. 20–26, келимярская свита, верхний тоар) установлена в алевритистых глинах. Фауна: фораминиферы, аммониты, белемниты и двустворки.
В целом для ассоциации характерно преобладание морского микрофитопланктона (от 33 до 88%). Диноцисты составляют от 26 до 82%. Резко сокращается количество празинофитов (до 6%). Постоянно присутствуют акритархи (54% в обр. 20). Зигнемовые и колониальные водоросли отсутствуют.
Среди диноцист происходит смена доминирования Nannoceratopsis на Phallocysta и Valvaeodinium, параллельно с сокращением количества празинофитов. Аналогичное событие было отмечено и в обн. 16. Вероятно, перидиниоидные диноцисты были менее толерантны к застойным условиям, чем нанноцератопсисы.
По составу микрофитопланктона фациальную обстановку можно охарактеризовать как удаленную от берега, умеренно глубоководную зону моря с нормальным солевым и газовым режимом.
По фораминиферам палеообстановка определена как внешняя часть верхней сублиторали (слои 4, 5) (Комплексное…, 2010).
Обнажение 7, р. Келимяр
В отложениях нижней юры, представленных глинистыми породами келимярской свиты, выделяются две сменяющие друг друга ассоциации – VI и Vб (рис. 6).
Рис. 6.
Типы эколого-фациальных ассоциаций палиноморф в обн. 7 на р. Келимяр. Условные обозначения см. на рис. 3.
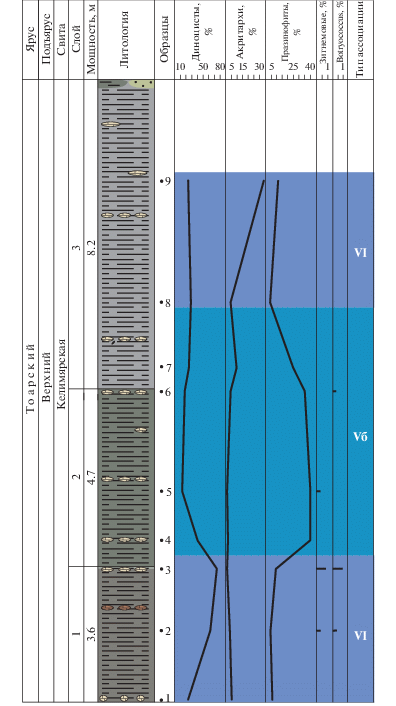
Ассоциация VI (слой 1, обр. 1–3, келимярская свита, верхний тоар) установлена в темно-серых глинах. Фауна: аммониты, фораминиферы.
В общем составе микрофоссилий, как правило, преобладает микрофитопланктон, а споры и пыльца наземных растений составляют от 17 до 66%. Для ассоциации характерно доминирование перидиниоидных диноцист Valvaeodinium. Диноцисты составляют от 23 до 73% с тенденцией к увеличению содержания вверх по разрезу.
Празинофиты (4.5–9%) и акритархи (1–5%) присутствуют постоянно, но в небольших количествах. Зигнемовые и колониальные водоросли представлены единичными таксонами (до 1%).
Учитывая доминирование диноцист, наличие акритарх и празинофитов и присутствие единичных зигнемовых и колониальных водорослей, фациальную обстановку можно охарактеризовать как достаточно удаленную от берега зону моря со стабильным солевым режимом.
Ассоциация Vб (слои 2–3, обр. 4–7, келимярская свита, верхний тоар) установлена в плотных глинах. Фауна: аммониты, двустворки, фораминиферы, белемниты, редкие гастроподы.
В общем составе микрофоссилий микрофитопланктон часто немного преобладает над спорами и пыльцой (17–49%). В целом для ассоциации характерно сокращение роли диноцист и резкое увеличение и преобладание празинофитов. Диноцисты составляют 12–40%, празинофиты – 25–45%, акритархи – 1–9%. По одному таксону зигнемовых и ботриококкусов встречено в обр. 5 и 6 соответственно.
Сокращение количества диноцист, присутствие акритарх и большое количество празинофитов, при участии редких единичных зигнемовых и колониальных водорослей, может свидетельствовать об обстановках, характерных для плохо аэрируемого морского бассейна.
Ассоциация VI (слой 3, обр. 8–9, келимярская свита, поздний тоар) установлена в темно-серых глинах. Фауна: фораминиферы, двустворки, белемниты, редкие гастроподы.
Споры и пыльца составляют от 30 до 64%. В группе микрофитопланктона доминируют диноцисты (22–27%) и акритархи (4–36%). Количество празинофитов значительно сокращается. Зигнемовые и колониальные водоросли отсутствуют.
Фациальную обстановку можно охарактеризовать как достаточно удаленную от берега зону моря со стабильным солевым и газовым режимом.
Обнажение 6, р. Келимяр
В нижнеюрских глинах обн. 6 установлена смена двух типов ассоциаций: IIIб и Vб (рис. 7).
Рис. 7.
Типы эколого-фациальных ассоциаций палиноморф на р. Келимяр в обн. 6. Условные обозначения см. на рис. 3. Кур. псв. – курунгская подсвита келимярской свиты.
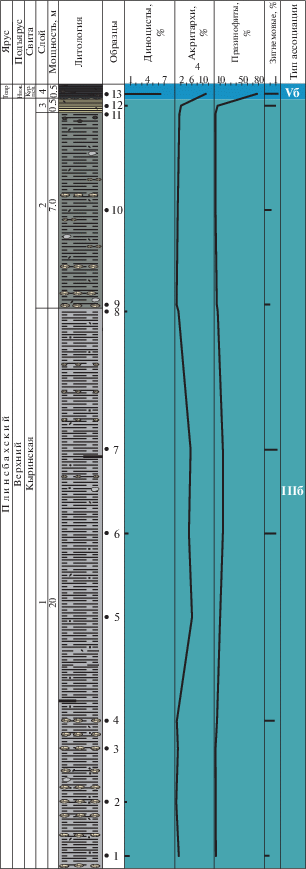
Ассоциация IIIб (слои 1–3, обр. 1–12, кыринская свита, верхний плинсбах) установлена в алевритистых глинах. Фауна: фораминиферы, двустворки. В верхней части слоя 2 встречены остатки обугленной древесины.
Споры и пыльца наземных растений значительно преобладают и составляют от 79 до 98%. Микрофитопланктон представлен единичными диноцистами Fromea и Nannoceratopsis senex, которые встречаются не во всех образцах. Акритархи (0.3–6%) и празинофиты (0.5–15%) присутствуют примерно в равных количествах. Стабильно в составе микрофитопланктона отмечены единичные зигнемовые (до 1%).
Фациальную обстановку можно охарактеризовать как прибрежно-морскую с активной гидродинамикой и опреснением.
Ассоциация Vб (слой 4, обр. 13, курунгская подсвита, келимярская свита, нижний тоар) установлена в черных листоватых сапропелевых глинах. Фауна: фораминиферы.
В общем составе преобладает микрофитопланктон, а споры и пыльца составляют 3.5%. Для ассоциации характерно присутствие небольшого количества (6.5%) перидиниоидных диноцист Phallocysta и Valvaeodinium. Акритархи присутствуют в количестве 11%, при значительном доминировании празинофитов Leiosphaeridia и Tasmanites (78%). Зигнемовые редки (до 1.5%).
Небольшое количество диноцист, присутствие акритарх и доминирование празинофитов, при участии редких зигнемовых, может свидетельствовать об обстановках, характерных для стагнационного этапа развития морского бассейна.
Обнажение 14, р. Келимяр
Все образцы (обр. 1–22) содержали недостаточное для статистической обработки количество микрофоссилий, что не позволило выявить соотношение разных групп палиноморф, восстановить эколого-фациальные обстановки и определить соответствующий тип ассоциации. Однако присутствие единичных диноцист, акритарх, празинофитов и зигнемовых позволяет предположить прибрежно-морские или морские обстановки в период формирования плинсбах-тоарских глин обн. 14.
Фауна: фораминиферы, двустворки, серпулы, брахиоподы в сл. 1–3 и аммониты, фораминиферы, белемниты, двустворки в сл. 5–7.
По фораминиферам палеообстановки охарактеризованы как внешняя часть верхней сублиторали (слой 1), чередование условий внешней и внутренней частей верхней сублиторали (слой 2), внутренняя часть верхней сублиторали (слой 3), верхняя сублитораль (слой 4), внешняя часть верхней сублиторали (слои 5, 6, 7) (Комплексное…, 2010).
Скв. Средне-Накынская 360
В отложениях нижней и средней юры, которые вскрываются скважиной Средне-Накынской 360, выделяются три типа ассоциаций – IIIa, Va и Vб (рис. 8).
Рис. 8.
Типы эколого-фациальных ассоциаций палиноморф в скв. Средне-Накынская 360. 1 – песчаник; 2 – алевролит глинистый; 3 – алевролит; 4 – аргиллит; 5 – галька.
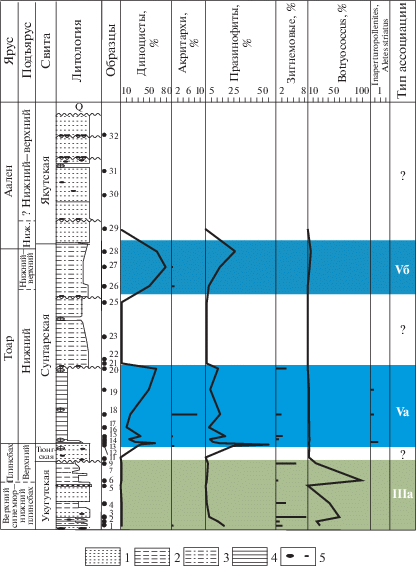
Ассоциация IIIa (обр. 1–7, укугутская свита, верхний синемюр–плинсбах) установлена в глинистых алевролитах с прослоями песчаника. Фауна: фораминиферы (Горячева, Никитенко, 2016).
Споры и пыльца преобладают над группой микрофитопланктона, который представлен диноцистами одного рода Mendicodinium (0.4–1.9%). Акритархи Leiofusa jurassica Cookson et Eisenack присутствуют в количестве до 0.5%. Содержание празинофитов, с преобладанием Leiosphaeridia, составляет от 0.8 до 16%. Зигнемовые водоросли (Schizosporis) составляют от 0.4 до 9%. Для ассоциации характерно большое количество колониальных зеленых водорослей Botryococcus (18–98%). Таким образом, с учетом редких диноцист, постоянного присутствия празинофитов и зигнемовых на фоне большого количества Botryococcus фациальные условия можно определить как прибрежно-морские, с опреснением вод.
Ассоциация Vа (обр. 11–20, тюнгская и сунтарская свиты, верхний плинсбах–нижний тоар) установлена в аргиллитах, переслаивающихся с песчаниками. Фауна: фораминиферы.
Роль наземных палиноморф довольно значительна. В группе микрофитопланктона преобладают диноцисты (10–62%) с доминированием Nannoceratopsis senex. Акритархи встречаются нестабильно и в небольших количествах (0.3–9%). Увеличилась роль празинофитов (до 55%). Регулярно присутствуют зигнемовые (1.2–3.5%). Резко уменьшилось количество ботриококкусов (до 4%). В ассоциации присутствуют единичные формы неясной систематической принадлежности – Inaperturopollenites и Aletes striatus Sachanova et Iljina.
Увеличение количества диноцист, акритарх и празинофитов при сокращении содержания Botryococcus может свидетельствовать о трансгрессивной направленности развития бассейна и последовательной смене опресненных прибрежных условий на более мористые. Однако довольно большое количество празинофитов дает основание предположить слабую аэрацию водной толщи.
Ассоциация Vб (обр. 26–28, сунтарская свита, верхняя часть нижнего тоара–верхний тоар) установлена в алевролитах. Фауна: фораминиферы.
Споры и пыльца имеют подчиненное значение (5–42%). Среди диноцист наблюдается доминирование бореально-арктических перидиниоидных Phallocysta eumekes Dӧrhӧfer et Davies (50–79%). Содержание акритарх не превышает 1%. Среди празинофитов (3–27%) доминирует Leiosphaeridia. Колониальные зеленые водоросли встречаются нестабильно и составляют от 0 до 6%. Зигнемовые отсутствуют.
По доминированию диноцист, наличию акритарх и празинофитов, отсутствию пресноводных водорослей и низкому содержанию колониальных зеленых водорослей можно сделать вывод о морских обстановках в период формирования этих толщ.
Скв. Ыгыатто-Тюнгская 1 (бассейн р. Вилюй)
Скважина Ыгыатто-Тюнгская 1 вскрывает нижнеюрские отложения, в которых выделено три ассоциации – IIIa, IIIб, IV (рис. 9).
Рис. 9.
Типы эколого-фациальных ассоциаций палиноморф в скв. Ыгыатто-Тюнгская 1. 1 – алевролит глинистый; 2 – алевролит песчаный; 3 – аргиллит алевритистый; 4 – алеврит, алевролит; 5 – песчаник; 6 – галька; 7 – конкреционные прослои; 8а – остатки древесины, 8б – углефицированный растительный детрит.
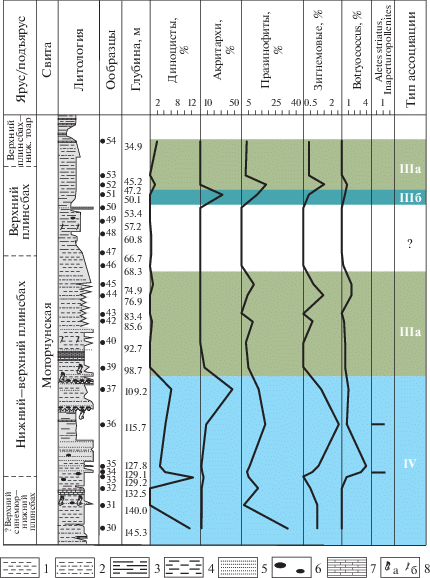
Ассоциация IV (обр. 30–37, моторчунская свита, верхний синемюр–нижний плинсбах) выделена в переслаивающихся аргиллитах и алевролитах с редкими прослоями глинистых песчаников. Фауна: фораминиферы.
Споры и пыльца преобладают. Диноцисты содержатся в количестве от 4 до 12%. Акритархи составляют до 46%. Стабильно встречаются празинофиты. В очень небольших количествах определены зигнемовые (1–2.6%) и колониальные водоросли Botryococcus (0.7–3.9%). Единичные формы неясной систематической принадлежности Inaperturopollenites и Aletes striatus найдены только в двух образцах.
Таким образом, по присутствию диноцист, празинофитов и акритарх, при участии небольшого количества празинофитов и зигнемовых водорослей, можно предположить обстановку, характерную для мелководной части шельфа с нестабильным солевым режимом.
Ассоциация IIIa установлена на двух уровнях (обр. 39–45 и обр. 52–54, моторчунская свита, плинсбах–нижний тоар) в переслаивающихся серых и темно-серых алевролитах, песчаниках и аргиллитах.
Характерно доминирование спор и пыльцы наземных растений (78–97%). Микрофитопланктон представлен в незначительном количестве. Диноцисты встречаются нестабильно и не превышают 2%. Празинофиты содержатся в количестве от 3 до 17%, зигнемовые водоросли – 0.4–1.5%, колониальные зеленые водоросли – 0.3–1.4%.
Незначительное количество диноцист, отсутствие акритарх, небольшое количество празинофитов, зигнемовых водорослей и колониальных водорослей свидетельствуют о мелководных прибрежно-морских (с опреснением) условиях обитания.
Ассоциация IIIб (обр. 51, моторчунская свита, верхний плинсбах) выделена в переслаивающихся серых аргиллитах и алевритистых аргиллитах. Фауна: фораминиферы.
Наземные палиноморфы (56.5%) незначительно преобладают над микрофитопланктоном. Диноцисты отсутствуют. Количество акритарх составляет 31%, празинофитов – 11.8%, зигнемовых водорослей – 0.4%, колониальных зеленых водорослей – 0.4%.
Состав и количественные соотношения палиноморф свидетельствуют о прибрежно-морских обстановках.
ЭКОЛОГО-ФАЦИАЛЬНЫЕ АССОЦИАЦИИ ПАЛИНОМОРФ В ИЗУЧЕННЫХ РАЗРЕЗАХ ЗАПАДНОЙ СИБИРИ
Скв. Северо-Правдинская 521 (Широтное Приобье)
В нижнеюрских отложениях, вскрытых скв. Северо-Правдинская 521, выделено три типа ассоциаций: IIIа, IIа и I (рис. 10).
Рис. 10.
Типы эколого-фациальных ассоциаций палиноморф в скв. Северо-Правдинская 521. 1 – песчаник; 2 – алевролит; 3 – аргиллит; 4 – уголь; 5 – галька, конгломерат; 6 – интракласты: а – глинистые, б – углистые; 7 – гравий, гравелит; 8 – УРД, 9 – алевропесчаник.
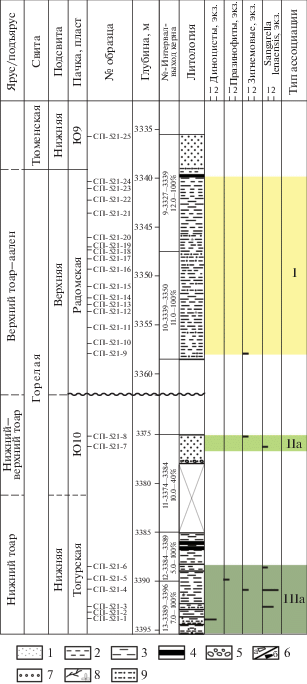
Ассоциация IIIа (обр. 1–6, горелая свита, тогурская пачка, нижний тоар) определена в переслаивающихся серых аргиллитах, алевролитах и песчаниках. Доминируют споры и пыльца наземных растений. Микрофитопланктон представлен единичными экземплярами диноцист, празинофитов и зигнемовых. Также встречены редкие формы неясной систематической принадлежности Sangarella lenaensis, которые приурочены “к осадкам озерно-пойменного и озерно-болотного генезиса, формирующихся в нестабильных обстановках приморских равнин, периодически заливаемых морем” (Пещевицкая, Фрадкина, 2001). В целом подобный состав солоновато-водного микрофитопланктона может свидетельствовать о влиянии моря на формирование вмещающих отложений. Согласно палеогеографическим картам китербютского горизонта (Гурари и др., 2005; Девятов и др., 2011), породы исследованного интервала формировались в прибрежной мелководной части шельфа.
Ассоциация IIа (обр. 7–8, горелая свита, тоар) определена в серых песчаниках. Доминируют споры и пыльца (не менее 90%). Микрофитопланктон представлен единичными экземплярами зигнемовых и Sangarella lenaensis. Можно предположить, что отложения формировались в условиях заболоченных низких прибрежных равнин, вероятно периодически заливаемых морем.
Ассоциация I (обр. 9–24, радомская пачка, верхняя подсвита горелой свиты, верхний тоар–аален) выделена в серых переслаивающихся алевролитах и песчаниках. Для ассоциации характерно полное отсутствие микрофитопланктона и наличие только спор и пыльцы наземных растений, что указывает на формирование вмещающих пород в сугубо континентальных обстановках.
Скв. Луль-Яхская 5П (Широтное Приобье)
В бат-(?)келловейских аргиллитах и глинистых алевролитах, вскрытых скважиной Луль-Яхской 5П, установлены две ассоциации – I и Va (рис. 11).
Рис. 11.
Типы эколого-фациальных ассоциаций палиноморф в скв. Луль-Яхская-5П. 1 – алевролит; 2 – песчаник; 3 – аргиллит; 4а – сидерит; 4б – пирит; 5а – отпечатки флоры; 5б – УРД; 6а – белемниты; 6б – двустворки; 6в – аммониты; к.к. – от конца керна.
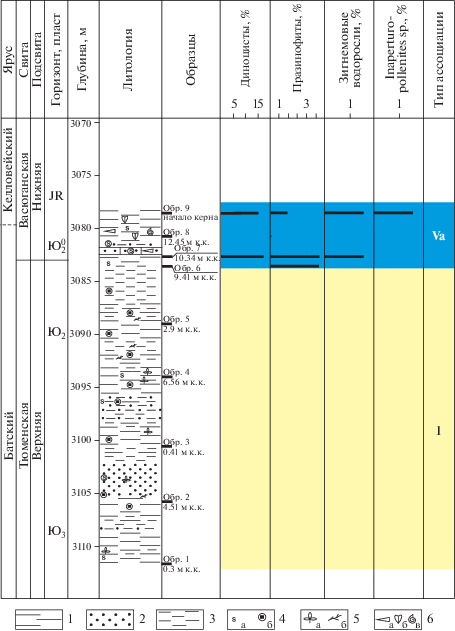
Ассоциация I (обр. 1–5, тюменская свита, бат) выделена в переслаивающихся алеврито-глинистых и песчано-алевролитовых породах.
Для ассоциации характерно отсутствие микрофитопланктона и наличие только спор и пыльцы наземных растений, в связи с чем можно предположить континентальный генезис вмещающих пород.
По седиментологическим данным палеообстановки характеризуются как меандровая коса (обр. 2), заболоченная пойменная равнина (обр. 3), пойменное озеро (обр. 4) и межгрядово-ложбинная прибрежная равнина (обр. 5) (Оценка…, 2004).
Ассоциация Va (обр. 6–9, тюменская и васюганская свиты, верхний бат–нижний келловей) выявлена в темно-серых аргиллитах и алевролитах. Фауна: аммониты, белемниты, двустворки.
Споры и пыльца доминируют. Микрофитопланктон представлен диноцистами нескольких родов (до 17%), празинофитами Leiosphaeridia и Tasmanites (1–5.2%), зигнемовыми водорослями Ovoidites (до 1.5%).
Наблюдается резкий переход от континентальных условий к морским обстановкам, что, вероятно, отражает позднебатскую–раннекелловейскую трансгрессию. Наличие диноцист свидетельствует об установлении морских обстановок с нормальным солевым режимом, что подтверждается присутствием в этих же слоях аммонитов, белемнитов и двустворок.
Скв. Западно-Тымская 1 (юго-восток Западной Сибири)
Ассоциация IIа (обр. 189–162, тогурская свита, нижний тоар) установлена в переслаивающихся алевролитах и аргиллитах (рис. 12).
Рис. 12.
Тип эколого-фациальной ассоциации палиноморф в скв. Западно-Тымская 1. 1 – аргиллит; 2 – алевролит; 3 – алевропесчаник.

Во всех палиноспектрах преобладают наземные палиноморфы. Микрофитопланктон встречается нестабильно в виде единичных таксонов акритарх Micrhystridium, празинофитов Tasmanites и Leiosphaeridia, зигнемовых Schizosporis, колониальных зеленых водорослей Botryococcus и форм неясной систематической принадлежности Aletes striatus.
Подобный состав микрофитопланктона позволяет предположить солоноватоводные обстановки, существующие в условиях прибрежных равнин, периодически заливаемых морем, что способствовало образованию озер.
Находки в средней части тогурской свиты представителей из отряда Phyllopoda (подотряд Conchostraca, род Pseudostheria?; определения А.Е. Игольникова (ИНГГ СО РАН)) свидетельствуют об озерном или лагунном с опреснением генезисе осадков (Чунихин, 2009).
Седиментологические данные указывают на формирование вмещающих пород в пределах крупного бассейна озерного типа (Ян и др., 2006).
Скважина Восток 4 (юго-восток Западной Сибири)
В нижне-среднеюрских отложениях, вскрытых скважиной Восток 4, установлена смена трех типов ассоциаций IIIа, IIа и I (рис. 13).
Рис. 13.
Типы эколого-фациальных ассоциаций палиноморф в разрезе скв. Восток 4. Условные обозначения см. на рис. 10.

Рис. 13.
Окончание
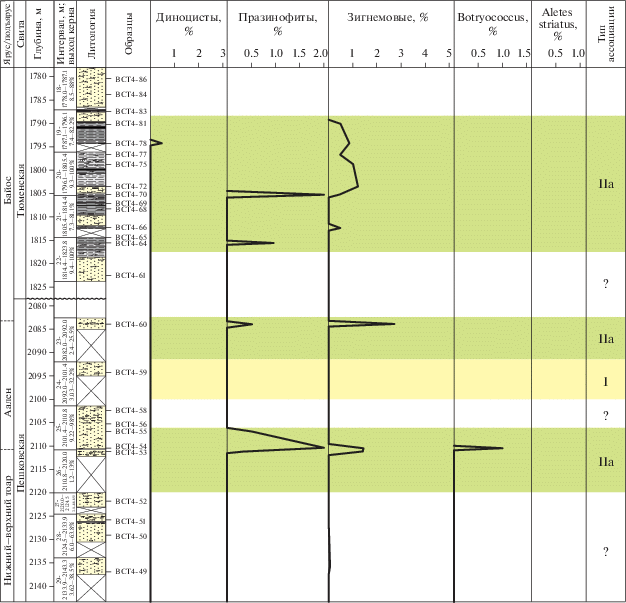
Ассоциация IIIа (обр. 5–9, урманская свита, верхний плинсбах) выделена в серых и буровато-серых песчаниках. Фауна: двустворки.
Преобладают споры и пыльца (до 98%).
Микрофитопланктон: единичные цисты динофлагеллат родов Mendicodinium и Sentusidinium, празинофиты Pterospermella, зеленые водоросли Schizosporis, колониальные водоросли Botryococcus и формы неясной систематической принадлежности Aletes striatus и Inaperturopollenites.
Наличие диноцист и празинофитов, которые являются обитателями морских обстановок, позволяет говорить о мелководных прибрежно-морских условиях.
На основе литологического анализа Л.Г. Вакуленко с коллегами (Вакуленко и др., 2010) предполагают, что формирование этой части урманской свиты “происходило вблизи морского побережья с периодическими кратковременными ингрессиями моря, во время которых формировались прибрежно-континентальные осадки с морской фауной, впоследствии размывающиеся речными системами при отступлении моря”.
Ассоциация IIIа (обр. 25–29, иланская свита, нижний тоар) выделена в толще переслаивающихся песчаников, алевролитов светло-серых и аргиллитов темно-серых. Фауна: многочисленные Phyllopoda.
Преобладают споры и пыльца наземных растений (89–98%).
Микрофитопланктон встречен в небольших количествах. В его состав входят: диноцисты (1.5–3.8%), акритархи (0.5%), празинофиты (до 1%), зигнемовые (до 5%), ботриококкусы (до 1%) и формы неясной систематической принадлежности Aletes striatus и Sangarella lenaensis.
Присутствие вышеперечисленного микрофитопланктона может свидетельствовать об условиях прибрежно-морских, с опреснением. Это могли быть обстановки эстуариевого или лагунного опресненного типа, также подобные фации могут быть характерны для прибрежного осолоненного озера.
По данным литологического анализа (Вакуленко и др., 2010), формирование иланской свиты “связано с существовавшим в это время довольно крупным озером или серией озер”.
Ассоциация IIа (обр. 17–24 и обр. 30–39, иланская свита, нижний тоар; обр. 46, пешковская свита, тоар; обр. 53–55, пешковская свита, тоар–а-ален; обр. 60, пешковская свита, аален; обр. 64–81, тюменская свита, байос) установлена в переслаивающихся серых, темно-серых песчаниках, алевролитах и аргиллитах.
Наземные палиноморфы доминируют (до 99%). Микрофитопланктон (до 3%) представлен празинофитами, зигнемовыми, колониальными Botryococcus и формами неясной систематической принадлежности. Такой состав может свидетельствовать о слабо солоноватоводных условиях обитания. Возможно, обстановки представляли собой заболоченные прибрежные участки суши.
Ассоциация I (обр. 59, пешковская свита, аален) определена в светло-серых песчаниках.
Для ассоциации характерно полное отсутствие микрофитопланктона и наличие только спор и пыльцы наземных растений, в связи с чем можно предположить континентальные условия, в которых формировались вмещающие породы.
По результатам литолого-седиментологических исследований накопление пешковской и тюменской свит происходило в обстановках обширной озерно-аллювиальной равнины (Вакуленко и др., 2010).
Обнажение 14, р. Золотой Китат
В разрезе обнажения 14 выделены два типа ассоциаций – IIa и IIб (рис. 14).
Рис. 14.
Типы эколого-фациальных ассоциаций палиноморф на р. Золотой Китат в обн. 14. 1 – уголь; 2 – глина; 3 – мергель; 4 – известняк; 5 – глина с включениями известняка; 6 – глина с включениями пирита.
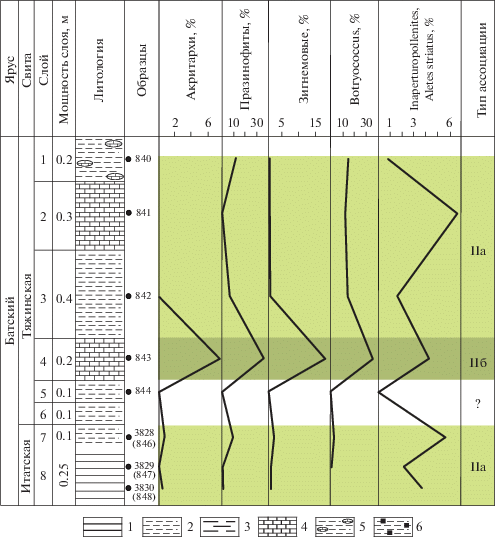
Ассоциация IIa (обр. 3838 (846)–3830 (848), слои 7–8, итатская свита–низы тяжинской свиты, бат; обр. 840–842, слои 1–3, тяжинская свита, бат) установлена в углях, глинах голубовато-серых и известняке голубовато-сером. Для нее характерно доминирование наземных палиноморф (80–95%). Общее содержание микрофитопланктона не превышает 20%. Акритархи (Veryhachium) составляют 0.4–1.2%, но встречаются нестабильно и в верхней части тяжинской свиты не обнаружены, празинофиты (Leiosphaeridia) – 1–12%, зигнемовые (Ovoidites, Sсhizocysta laevigata) – 0.2–2%, колониальные зеленые водоросли (Botryococcus) – 1.3–14%. Содержание форм неясной систематической принадлежности (Inaperturopollenites и Aletes striatus) составляет 2–5.5%.
По присутствию редких акритарх и празинофитов и довольно большому количеству колониальных зеленых водорослей и зигнемовых можно предположить солоноватоводную обстановку. Возможно, отложения формировались в условиях прибрежных равнин, периодически заливаемых морем, что способствовало образованию болот или озер.
Ассоциация IIб (обр. 843, слой 4, тяжинская свита, бат) выявлена в голубовато-сером известняке. Она примечательна полным отсутствием спор и пыльцы наземных растений и наличием только микрофитопланктона. Акритархи содержатся в количестве 7.7%. Содержание празинофитов (Leiosphaeridia) доходит до 36%, зигнемовых водорослей (Ovoidites) до 18%. Колониальные зеленые водоросли (Botryococcus) составляют 35%, формы неясного систематического положения – до 4.5%.
Присутствие среди микрофитопланктона акритарх, большого количества празинофитов и зигнемовых водорослей говорит о прибрежной опресненной обстановке. Возможно, данная фация отражает обстановки эстуариевого или лагунного типа.
Обнажение 15, р. Золотой Китат
Тяжинская свита в обнажении 15 у пос. Новый Свет характеризуется наличием двух ассоциаций – I и IIa (рис. 15).
Рис. 15.
Типы эколого-фациальных ассоциаций палиноморф на р. Золотой Китат в обн. 15. Условные обозначения см. на рис. 14.

Ассоциация IIa (обр. 864, слой 6, тяжинская свита, бат; обр. 859–858, слой 5, тяжинская свита, бат) выделена в глинах голубовато-серых и глинах голубовато-серых, известковистых. Для нее характерно содержание наземных палиноморф до 79%. Среди микрофитопланктона встречаются акритархи (Leiofusa) (0.5%), празинофиты Leiosphaeridia (4%), зигнемовые водоросли (Ovoidites) (3.5%), колониальные зеленые водоросли (Botryococcus) (7%). Количество форм неясного систематического положения (Inaperturopollenites) не превышает 6.5%.
Подобный состав микрофитопланктона отвечает обстановке солоноватоводного водоема недалеко от морского бассейна или, возможно, залива лагунного типа. Близость морского бассейна определяется наличием редких акритарх и небольшого количества празинофитов.
Ассоциация I (обр. 857–855, слои 5–4, тяжинская свита, бат; обр. 852–850, слой 2, тяжинская свита, бат) выделена в глинах голубовато-серых известковистых, глинах черных сажистых. В общем составе доминируют споры и пыльца наземных растений (до 89%). Из представителей микрофитопланктона присутствуют зигнемовые (1–2%), ботриококкусы 4–63% и формы неясного систематического положения.
Подобный состав палиноморф характерен для ассоциации I континентального типа седиментогенеза.
Таким образом, изучение разных групп микрофитопланктона, динамики его распределения и фациальной приуроченности позволило сделать выводы о генезисе вмещающих отложений в естественных обнажениях или скважинах и восстановить эколого-фациальные обстановки (от континентальных до морских относительно глубоководных) в период их формирования.
ЗАКЛЮЧЕНИЕ
Впервые для нижней и средней юры Западной и Восточной Сибири проведено комплексное изучение всех групп палиноморф (спор и пыльцы наземных растений, диноцист, акритарх, празинофитов, зигнемовых и колониальных водорослей) из разнофациальных разрезов. Выявлена эколого-фациальная приуроченность микрофитопланктона на основе анализа распределения палиноморф в тех разрезах, где условия осадконакопления были восстановлены по палеонтологическим, литологическим или другим данным, а также с привлечением существующих сведений по экологии современных и ископаемых одноклеточных водорослей. По количественным соотношениям различных групп микрофитофоссилий в разрезах Западной и Восточной Сибири выделено девять типов ассоциаций палиноморф, которые являются показателями разных обстановок – от континентальных до типично морских. Полученные результаты могут быть использованы для уточнения и детализации региональных стратиграфических схем и схем фациального районирования нижней и средней юры Сибири, являющихся инструментом при проведении поисково-разведочных и геолого-съемочных работ.
Благодарности. Автор выражает искреннюю признательность Б.Л. Никитенко, В.Г. Князеву, В.П. Девятову, Л.Г. Вакуленко и $\boxed{{\text{В}}{\text{.И}}{\text{. Ильиной}}}$ за предоставленные геологические материалы и научные консультации.
Источники финансирования. Работа выполнена при поддержке проекта ФНИ № 0331-2019-0004 и гранта РФФИ (проект № 19-05-00130).
Список литературы
Вакуленко Л.Г., Аксенова Т.П., Ельцов И.С. и др. Литолого-фациальная характеристика юрских отложений южной части Предъенисейской нефтегазоносной субпровинции // Геология и геофизика. 2010. № 4. С. 128–139.
Вдовин В.В., Ильина В.И. К вопросу о нижней границе тяжинской свиты // Стратиграфия мезозоя и кайнозоя Средней Сибири. Новосибирск: Наука, 1967. С. 94–97.
Водоросли. Справочник. Киев: Наукова думка, 1989.
Горячева А.А. Палиностратиграфия среднеюрских отложений в разрезах бассейна р. Золотой Китат (юго-восток Западной Сибири) // Новости палеонтологии и стратиграфии. Приложение к журналу “Геология и геофизика”. 2006. Т. 47. Вып. 8. С. 125–137.
Горячева А.А. Палинологическая характеристика тоарских отложений в разрезах скважин Западно-Тымская 1 и Северо-Правдинская 521 (Западная Сибирь) // Новости палеонтологии и стратиграфии. Приложение к журналу “Геология и геофизика”. 2008. Т. 49. Вып. 10–11. С. 205–211.
Горячева А.А. Палиностратиграфия нижне-среднеюрских отложений в разрезе скважины Восток 4 (юго-восток Западной Сибири) // Стратиграфия. Геол. корреляция. 2011. Т. 19. № 3. С. 27–47.
Горячева А.А. Биостратиграфия нижнеюрских отложений Восточной Сибири по данным палинологии // Стратиграфия. Геол. корреляция. 2017. Т. 25. № 3. С. 29–60.
Горячева А.А., Никитенко Б.Л. Биостратиграфия нижнеюрских отложений в разрезе скв. Средне-Накынская 360 (Восточная Сибирь) // Водоросли в эволюции биосферы. Материалы II Палеоальгологической конференции (г. Новосибирск, 10–16 октября 2016 г.). Новосибирск: ИНГГ СО РАН, 2016. С. 33–35.
Гурари Ф.Г., Девятов В.П., Демин В.И. и др. Геологическое строение и нефтегазоносность нижней–средней юры Западно-Сибирской провинции. Новосибирск: Наука, 2005. 156 с.
Девятов В.П., Князев В.Г., Никитенко Б.Л. и др. Граница плинсбаха и тоара севера Восточной Сибири и стратиграфическое положение курунгской пачки келимярской свиты (р. Келимяр, бассейн р. Оленек) // Отечественная геология. 2010. № 5. С. 105–112.
Девятов В.П., Никитенко Б.Л., Шурыгин Б.Н. Палеогеография Сибири в юрском периоде на этапах основных перестроек // Новости палеонтологии и стратиграфии. Приложение к журналу “Геология и геофизика”. 2011. Т. 52. Вып. 8. С. 87–103.
Ильина В.И., Кулькова И.А., Лебедева Н.К. Микрофитофоссилии и детальная стратиграфия морского мезозоя и кайнозоя Сибири. Новосибирск: ОИГГиМ СО РАН, 1994.
Князев В.Г., Девятов В.П., Кутыгин Р.В. и др. Зональный стандарт тоарского яруса Северо-Востока Азии. Якутск: Изд-во СО РАН, 2003.
Комплексное полевое геологическое изучение естественных выходов мезозоя нижнего течения р. Оленек, побережья Оленекского залива моря Лаптевых и стратиграфический, биофациальный и седиментационный анализ образцов из разрезов мезозоя бассейна р. Оленек и острова Котельный. Отчет по договору (заключительный) 2-10 в 2-х томах. Рук. Никитенко Б.Л. Новосибирск: Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН, 2010. 511 с.
Лебедева Н.К. Биофациальный анализ верхнемеловых отложений Усть-Енисейского района по палиноморфам // Стратиграфия. Геол. корреляция. 2008. Т. 16. № 2. С. 81–97.
Лебедева Н.К. Палинофации верхнемеловых отложений севера Сибири // Стратиграфия. Геол. корреляция. 2010. Т. 18. № 5. С. 70–87.
Никитенко Б.Л. Стратиграфия, палеобиогеография и биофации юры Сибири по микрофауне (фораминиферы и остракоды). Новосибирск: Параллель, 2009.
Никитенко Б.Л., Поспелова В.Ю. Микробиота (бентос и фитопланктон) и биофации в ранне-среднеюрских морях на севере Сибири // Геодинамика и эволюция Земли. Материал Научн. конф. РФФИ. Новосибирск: Изд-во СО РАН НИЦ ОИГГМ, 1996. С. 177–180.
Никитенко Б.Л., Суан Г., Девятов В.П. и др. Тоарское аноксидное океаническое событие (T-OAE) в Арктике (седиментологические, микропалеонтологические и геохимические свидетельства) // Палеонтология, стратиграфия и палеогеография мезозоя и кайнозоя бореальных районов. Материалы научной сессии в 2 т. Новосибирск: ИНГГ СО РАН, 2011. С. 202–209.
Никитенко Б.Л., Шурыгин Б.Н., Князев В.Г. и др. Стратиграфия юры и мела Анабарского района (Арктическая Сибирь, побережье моря Лаптевых) и бореальный зональный стандарт // Геология и геофизика. 2013. Т. 54. № 8. С. 1047–1082.
Никитенко Б.Л., Девятов В.П., Лебедева Н.К. и др. Стратиграфия юры и мела архипелага Новосибирские острова (море Лаптевых и Восточно-Сибирское море), фациальное районирование и литостратиграфия // Геология и геофизика. 2017. Т. 58. № 12. С. 1867–1885.
Никитенко Б.Л., Девятов В.П., Лебедева Н.К. и др. Биостратиграфия и особенности геохимии органического вещества юры и мела архипелага Новосибирские острова (Российская Арктика) // Геология и геофизика. 2018. Т. 59. № 2. С. 209–228.
Оценка перспектив и выявление благоприятных зон распространения высокодебитных коллекторов в горизонте Ю2 на территории Юганского Приобья. Отчет по договору (заключительный) 06-03. Рук. Казаненков В.А. Новосибирск: Институт нефтегазовой геологии и геофизики им. А.А. Трофимука СО РАН, 2004. 250 с.
Пещевицкая Е.Б., Фрадкина А.Ф. Ревизия мезозойского вида Laevigatisporites lenaensis Fradkina 1964 (морфология, систематика, значение для стратиграфии и палеофациальных реконструкций) // Новости палеонтологии и стратиграфии. Приложение к журналу “Геология и геофизика”. 2001. Вып. 4. Т. 42. С. 109–123.
Чунихин С.А. Конхостраки перми и триаса Западной Сибири. Автореф. дисc. … канд. геол.-мин. наук. Томск, 2009. 23 с.
Ян П.А., Вакуленко Л.Г., Горячева А.А. и др. Строение, состав и условия формирования тогурской свиты по результатам бурения Западно-Тымской скважины № 1 (нижний тоар, Западная Сибирь) // Палеонтология, биостратиграфия и палеогеография бореального мезозоя. Материалы научной сессии, Новосибирск, 26–28 апреля, 2006 г. Новосибирск: Гео, 2006. С. 213–216.
Яшнов В.А. Водные массы и планктон Calanusglacialis и Calanuspacificus как индикаторы определенных водных масс морей Тихого океана // Зоологический журнал. 1965. Т. 42. С. 1005–1021.
Batten D.J., Grenfell H.R. Green and blue-green algae. Botryococcus // Palynology: Principles and Applications. Eds. Jansonius J., McGregor D.C. Salt Lake City: American Association of Stratigraphic Palynologists Foundation, 1996. V. 1. P. 205–214.
Boalch G.T., Parke M. The prasinophycean genera (Chlorophyta) possibly related to fossil genera, in particular the genus Tasmanites // Proc. II Planktonic Conf. Roma, 1971. Pl. 1. P. 99–105.
Bujak J.P., Davies E.H. Modern and fossil Peridiniineae // AASP Contrib. Ser. 1983. № 13. 203 p.
Dale B. Dinoflagellate cyst ecology: modeling and geological applications // Palynology: Principles and Applications. Eds. Jansonius J., McGregor D.C. Salt Lake City: American Association of Stratigraphic Palynologists Foundation, 1996. V. 3. P. 1249–1275.
Evitt W.R. The dinoflagellate Nannoceratopsis Deflandre: morphology, affinities and intraspecific variability // Micropalaeontology. 1961. V. 7. P. 305–316.
Fensome R.A., Taylor F.J.R., Norris G., Sarjeant W.A.S., Wharton D.I., Williams G.L. A classification of living and fossil dinoflagellates // Micropalaeontology Spec. Publ. 1993. № 7. 351 p.
Geel B., Grenfell H.R. Spores of Zygnemataceae // Palynology: Principles and Applications. Eds. Jansonius J., McGregor D.C. Salt Lake City: American Association of Stratigraphic Palynologists Foundation, 1996. V. 2. P. 173–181.
Goodman D.K. Chapter 15. Dinoflagellate cysts in ancient and modern sediments // The biology of dinoflagellates. Botanical Monographs. V. 21. Ed. Taylor F.G.R. Oxford: Blackwell Scientific, 1987. P. 649–722.
Guy-Ohlson D. Prasinophycean algae // Palynology: Principles and Applications. Eds. Jansonius J., McGregor D.C. Salt Lake City: American Association of Stratigraphic Palynologists Foundation, 1996. V. 2. P. 181–191.
Harland R. Quaternary dinoflagellate cyst biostratigraphy of the North Sea // Palaeontology. 1988. V. 33. № 3. P. 877–903.
Harrison P.J., Conway H.L., Holmes R.W., Davies C.O. Marine diatoms growth in chemostat under silicate of ammonium limitation. III. Cellular chemical composition and morphology of Chaetoceros debilis, Skeletomea costatum, and Thalassiosira gravida // Mar. Biol. 1977. V. 43. P. 19–31.
Hinga K.R. Co-occurrence of dinoflagellate blooms and high pH in marine enclosures // Marine Ecology Progress Ser. 1992. V. 86. P. 181–187.
Hoek C., Mann D.G., Jahns H.M. Algae. An Introduction to Phycology. Cambridge: Cambridge Univ. Press, 1995.
Hulburt E.M. The diversity of phytoplanktonic population of oceanic, coastal and estuarine regions // Marine Res. 1963. V. 21. P. 81–93.
Marret F., Zonneveld K.A.F. Atlas of modern organic-walled dinoflagellate cyst distribution // Rev. Palaeobot. Palynol. 2003. V. 125. P. 1–200.
Martin F. Acritarchs: a review // Biol. Rev. Cabr. Phil. Soc. 1993. V. 68. № 4. P. 475–538.
Mudie P.J., Harland R. Chapter 21. Aquatic Quaternary // Palynology: Principles and Applications. Eds. Jansonius J., McGregor D.C. Salt Lake City: American Association of Stratigraphic Palynologists Foundation, 1996. V. 2. P. 843–877.
Prauss M. Dinozysten – Stratigraphie und Palynofazies im Oberen Lias und Dogger von NW-Deutschland // Palaeontographica. 1989. Bd. 214. P. 1–124.
Prauss M., Rigel W. Evidence from phytoplankton associations for causes of black shale formation in epicontinental seas // NJ Geol. Palaontol. Monatsh. 1989. H. 11. P. 671–682.
Sarjeant W.A.S., Lacalli T., Gaines G. The cysts and skeletal elements of dinoflagellates: speculations on the ecological causes for their morphology and development // Micropalaeontology. 1987. V. 33. № 1. P. 1–36.
Strother P.K. Acritarchs // Palynology: Principles and Applications. Eds. Jansonius J., McGregor D.C. Salt Lake City: American Association of Stratigraphic Palynologists Foundation, 1996. V. 1. P. 81–106.
Tappan H. The Paleobiology of plant protists. San Francisco, 1980. 1028 p.
Taylor F.G.R. Chapter 11: Ecology of dinoflagellates // The biology of dinoflagellates. Botanical Monographs. V. 21. Ed. Taylor F.G.R. Oxford: Blackwell Scientific, 1987. P. 399–529.
Wall D., Dale B., Lohmann G., Smith W.K. The environmental and climatic distribution of dinoflagellate cysts in modern marine sediments from regions in the north and south Atlantic oceans and adjacent seas // Marine Micropalaeontology. 1977. V. 2. P. 121–200.
Yoo K.L. Population dynamics of dinoflagellate community in Masan Bay with a note on the impact of environmental parameters // Marine Pollution Bull. 1991. V. 23. P. 185–188.
Zonneveld K.A.F., Hoek R.P., Brinkhuis H., Willems H. Geographical distribution of organic-walled dinoflagellate cysts in surfical sediments of the Benguella upwelling region and their relationship to upper oceanic conditions // Rev. Palaeobot. Palynol. 2001. V. 48. P. 25–72.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция