

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 1, стр. 65-72
О стратиграфическом положении меловых отложений с гребенкинской флорой, Северо-Восток России
С. В. Щепетов *
Ботанический институт им. В.Л. Комарова РАН
Санкт-Петербург, Россия
* E-mail: shchepetov@mail.ru
Поступила в редакцию 16.04.2018
После доработки 13.07.2018
Принята к публикации 28.03.2019
Аннотация
Гребенкинская флора позднеальбского–раннетуронского возраста является одной из самых представительных среди флор середины мелового периода Северо-Востока России. Однако стратиграфическое положение вмещающих ее слоев относительно флороносных отложений близкого возраста было неясным. Показано, что наиболее вероятным стратиграфическим аналогом терригенной кривореченской свиты является вулканогенная саламихинская толща, содержащая растительные остатки буор-кемюсской флоры. Отложения с гребенкинской флорой залегают стратиграфически ниже вулканогенных образований, включающих чаунскую флору.
ВВЕДЕНИЕ
На Северо-Востоке Азии широко представлены континентальные отложения мелового возраста. Они заполняют несколько осадочных бассейнов, а также слагают огромную геологическую структуру – Охотско-Чукотский вулканогенный пояс (ОЧВП) (рис. 1а). Интенсивное изучение этих образований началось в середине ХХ века, и сразу же встал вопрос об обосновании возраста стратифицированных геологических тел. Остатков морских моллюсков и фораминифер континентальные образования не содержат, ископаемые споры и пыльца в вулканогенных толщах сохраняются чрезвычайно редко, а изотопные методы в то время были мало пригодны для целей локальных стратиграфических корреляций. Однако в терригенных отложениях и в туфовых прослоях вулканогенных толщ нечасто, но регулярно встречаются остатки ископаемых растений. К началу 1970-х годов был накоплен значительный палеофлористический материал, который обобщила В.А. Самылина (1974). Она предложила базовую схему развития ископаемой флоры в меловом периоде для региона в целом. Правда, позже было установлено, что на территории Северо-Востока России меловая флора развивалась по-разному в зависимости от палеоландшафтных обстановок (Герман, 1993; Щепетов, 1995). Однако, при всех недостатках, схема Самылиной впервые дала возможность определять возраст флороносных отложений в относительно узких пределах и коррелировать их друг с другом.
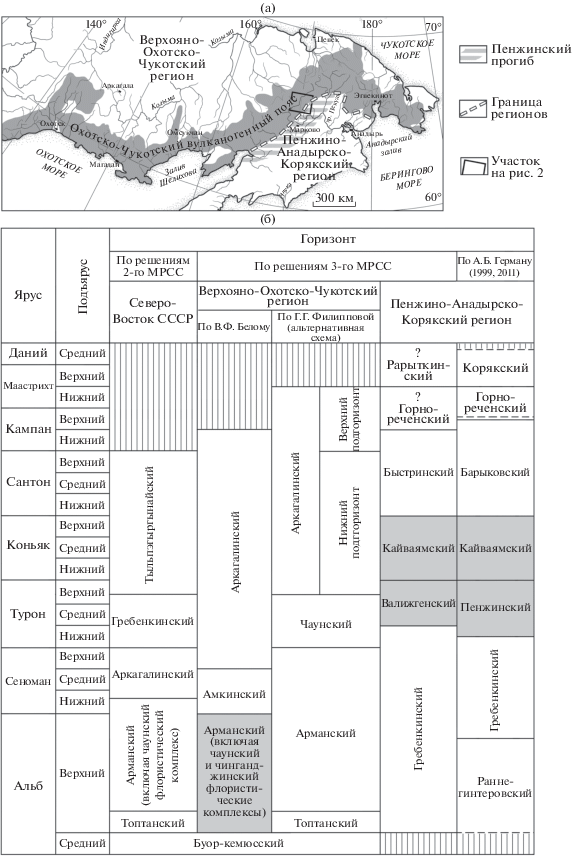
Рис. 1.
(а) Элементы районирования Северо-Востока России для мелового периода и (б) стратиграфическое расчленение неморского мела (по Решения…, 2009, с изменением; серым цветом выделены одновозрастные образования).
В данной статье использованы термины “палеофлористический комплекс” или “тафофлора”. Это элементарные палеофлористические единицы, представляющие собой совокупность ископаемых растений из одного или нескольких территориально и стратиграфически близких местонахождений. Тафофлоры или палеофлористические комплексы, обладающие существенно сходными чертами, то есть характерным сочетанием таксонов, качественным и количественным соотношением групп растений и т.д., автор, вслед за А.Б. Германом (2011), рассматривает как ископаемую флору или палеофлору, характеризующую определенный этап развития растительного мира на значительной части территории (например, в Северном Приохотье). Слоями с собственными названиями (например, “гребенкинские слои”) обозначены вспомогательные биостратиграфические подразделения, представляющие собой отложения с остатками одноименной палеофлоры (Стратиграфический…, 2006, Статья VII.10.)
В северо-восточной части ОЧВП вулканогенные образования граничат с терригенными морскими и континентальными отложениями Пенжинского прогиба (рис. 1а). На протяжении мелового периода здесь, как и на территории современного Корякского нагорья, неоднократно происходили трансгрессии и регрессии моря. Чередование в разрезах морских и континентальных флороносных отложений в ряде случаев дает возможность датировать комплексы ископаемых растений по остаткам морских моллюсков (аммонитов и иноцерамов) и фораминифер. Возраст флористических комплексов из образований ОЧВП и внутриматериковых бассейнов определяется путем сравнения их систематического состава с составом фаунистически датированных “приморских” палеофлор – гребенкинской, пенжинской, кайваямской и барыковской (Герман, 2011).
Гребенкинская палеофлора из отложений кривореченской свиты Пенжинского прогиба является наиболее представительной среди приморских палеофлор мелового возраста Северо-Востока России. По сопутствующим остаткам морской фауны, а также палинологическим, изотопным и палеомагнитным методами установлено, что она существовала в конце альба, сеномане и начале турона (Щепетов и др., 1992; Филиппова, Абрамова, 1993; Spicer et al., 2002). Довольно многочисленные захоронения остатков растений гребенкинской палеофлоры установлены на протяжении примерно 200 км вдоль северо-западного борта Пенжинского прогиба и, соответственно, юго-восточной границы ОЧВП в бассейне р. Анадырь. Гребенкинская флора могла бы стать главным фитостратиграфическим репером неморского мела Северной Пацифики, однако с ней связан ряд не решенных пока вопросов.
Первоначально считалось, что гребенкинская флора существовала после арманского и аркагалинского этапов развития, что она моложе всех флористических комплексов ОЧВП (Самылина, 1974; Белый, 1977; Решения…, 1978) (рис. 1б). Однако со временем удалось доказать, что в последовательности этапов развития меловой флоры Северо-Востока России гребенкинский этап был значительно более ранним, чем принято было считать (Герман, 1993, 2011; Филиппова, Абрамова, 1993; Щепетов, 1995; Решения…, 2009).
На Третьем Межведомственном стратиграфическом совещании (МРСС) было признано, что арманская и чинганджинская флоры являются одновозрастными (Решения…, 2009) (рис. 1б). В.А. Самылина еще в 80-х годах ХХ в. сопоставила чинганджинскую флору с приморскими пенжинской и кайваямской флорами и датировала ее туроном–коньяком (Самылина, 1988). Эту датировку, подтвержденную более поздними исследованиями (Герман, 2011; Головнева и др., 2011), никто никогда не оспаривал, однако арманская и чинганджинская флоры на Третьем МРСС были датированы поздним альбом (Решения…, 2009).
По представлениям автора, арманская и чинганджинская флоры действительно являются одновозрастными в геологическом масштабе времени, но датировать их, конечно, следует туроном–коньяком (Herman et al., 2016). Предполагавшийся ранее предшественник арманской флоры – топтанский флористический комплекс – решениями Третьего МРСС включен в состав буор-кемюсской флоры (Решения…, 2009). В итоге получается, что буор-кемюсская флора закончила свое существование в начале позднего альба, а сменившие ее арманская и чинганджинская флоры появились только в туроне. Перерыв между временами существования этих флор как раз и приходится на время существования гребенкинской флоры. Однако в стратиграфических разрезах меловых отложений ОЧВП нет решительно никаких признаков такого перерыва. Остается предположить, что либо аналоги гребенкинской палеофлоры в ОЧВП пока не найдены (что маловероятно), либо флористические комплексы, одновозрастные гребенкинской палеофлоре, вдали от приморских низменностей имели иной систематический состав, и мы просто не узнаем их, не признаем “гребенкинскими” и датируем как-то иначе.
Решить эту проблему могли бы данные о стратиграфических соотношениях отложений кривореченской свиты с вулканогенными образованиями ОЧВП. Однако в научной литературе такие соотношения никогда не обсуждались. Вряд ли стоит рассчитывать на новые полевые исследования стратотипического района кривореченской свиты, поскольку больше не осталось специалистов соответствующей квалификации, способных совершать дальние маршруты по анадырской тундре. Поэтому для решения поставленного вопроса автор попытался проанализировать доступные картографические и литературные данные.
Стратотипической местностью кривореченской свиты является бассейн р. Кривая, правого притока р. Убиенка. Стратотипическим районом свиты автор называет более обширную территорию – бассейн верховьев р. Убиенка и часть правобережья р. Чинейвеем (рис. 2а).
Рис. 2.
(а) Схема распространения альбских–позднемеловых накоплений в бассейне среднего течения р. Анадырь и верховьев р. Большой Анюй по материалам государственных геологических карт масштаба 1 : 200 000 и (б) корреляция местных стратиграфических подразделений этого района.
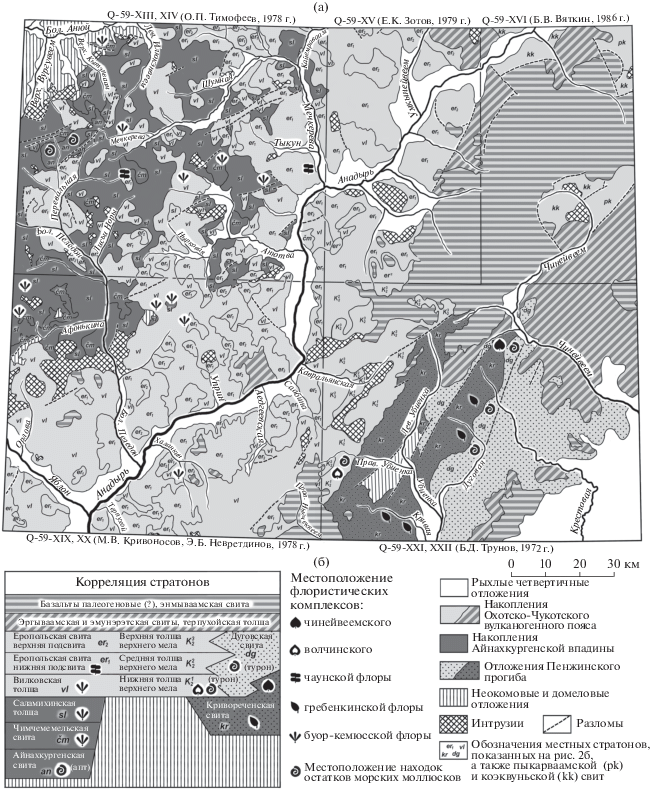
Меловые отложения в бассейне среднего течения р. Анадырь в разные годы изучали В.Ф. Белый, И.П. Васецкий, В.Г. Гаврилов, В.М. Гринфельд, А.Д. Девятилова, Б.Н. Елисеев, В.Н. Завражнов, В.Ф. Зуев, Г.Г. Кайгородцев, И.Н. Котляр, Л.Л. Красный, Э.Б. Невретдинов, Г.П. Терехова, Б.Н. Трунов. Позже на правобережье р. Анадырь проводили полевые работы А.Б. Герман, С.В. Щепетов, Р.Э. Спайсер, А.Т. Альберг, П.М. Рис и М.И. Райкевич. Кривореченскую свиту с опорным разрезом по р. Кривая в 1962 г. выделил Г.Г. Кайгородцев.
СТРАТИГРАФИЧЕСКОЕ ПОЛОЖЕНИЕ ГРЕБЕНКИНСКИХ СЛОЕВ В СТРАТОТИПИЧЕСКОМ РАЙОНЕ КРИВОРЕЧЕНСКОЙ СВИТЫ
По данным Б.Н. Трунова (Государственная…, 1977), кривореченская свита в своем стратотипическом районе и по левобережью р. Левая Убиенка представляет собой мощную (1200–2200 м) толщу туфоконгломератов, туфопесчаников и алевролитов. В бассейне р. Правая Убиенка и в верховьях р. Левая Убиенка в ее составе появляются лавы и лавобрекчии, туфы и туфобрекчии среднего состава. Свита с угловым несогласием залегает на различных горизонтах юры, валанжинского и готеривского (?) ярусов нижнего мела (Государственная…, 1977).
На левобережье р. Левая Убиенка, в верховьях р. Дуговая и на правобережье р. Чинейвеем кривореченскую свиту без признаков стратиграфического перерыва c постепенным переходом перекрывают морские отложения дуговской свиты. Они содержат остатки морских моллюсков туронского возраста (Государственная…, 1977; Терехова, 1988). В бассейнах рек Убиенка и Кавральянская кривореченскую свиту согласно перекрывает безымянная вулканогенная нижняя толща верхнего мела. В нижнем слое стратотипического разреза этой толщи по руч. Каменистый, описанному Труновым, помимо остатков растений, обнаружены остатки морских моллюсков. В этих же местонахождениях в 1974–1976 гг. Невретдиновым и Девятиловой проведены новые палеонтологические сборы. По остаткам двустворок и аммонитов они уверенно датировали вмещающие отложения туроном (Девятилова и др., 1980; Невретдинов, 1980). Собранные здесь же остатки растений рассматриваются как волчинский флористический комплекс, также датируемый туроном (Девятилова и др., 1980; Филиппова, Абрамова, 1993). По представлениям Невретдинова и Девятиловой, данные слои с остатками фауны и флоры располагаются в средней части разреза описанной ими волчинской свиты (Девятилова и др., 1980; Невретдинов, 1980).
В поле выходов кривореченской свиты на левобережье р. Чинейвеем между ручьями Болотный и Обильный А.И. Дворянкин, Е.Л. Лебедев и Н.И. Филатова (Дворянкин и др., 1993) собрали комплекс остатков растений, названный “чинейвеемским”. По особенностям систематического состава он “несомненно, моложе флор пенжинского (валижгенского) типа (поздний турон–коньяк), которые, в свою очередь, наследуют гребенкинские флоры (сеноман)” (Дворянкин и др., 1993, с. 475). Так как в чинейвеемском комплексе присутствуют формы, сходные с представителями семейства Fagacea, его отнесли к сенону. Поскольку новый флористический комплекс оказался моложе гребенкинской флоры и фауны дуговской свиты, Дворянкин с соавторами сочли, что вмещающие отложения не относятся к кривореченской свите, а слагают “тектонический блок шириной около 4 км, ограниченный дугообразной формы разломами” (Дворянкин и др., 1993, с. 473). А.Б. Герман проанализировал флористические определения Лебедева, ознакомился с коллекцией растительных остатков и пришел к выводу, что чинейвеемская тафофлора “безусловно более молодая, чем гребенкинская флора правобережья р. Анадырь, и наиболее близка пенжинской тафофлоре Северо-Западной Камчатки и п-ова Елистратова, возраст которой надежно установлен как турон, исключая, вероятно, начало турона” (Герман, 2011, с. 54).
Вскоре после выхода статьи Дворянкина и его соавторов (1993) С.В. Щепетов ознакомился со всеми опубликованными и фондовыми материалами по данному району, провел дешифрирование аэрофотоснимков. Никаких структурных элементов, которые можно было бы истолковать как тектонические разломы, отделяющие отложения с чинейвеемской тафофлорой от кривореченской и дуговской свит, обнаружить не удалось. Как и показано на карте Трунова (Государственная…, 1977), местонахождение чинейвеемского комплекса расположено в самой верхней части разреза кривореченской свиты близ подошвы дуговской свиты.
Таким образом, в стратотипическом районе кривореченской свиты гребенкинские слои, датируемые концом альба–началом турона, согласно перекрываются туронскими слоями с волчинской тафофлорой и слоями с чинейвеемским комплексом, датируемым туроном, исключая начало. Волчинские слои входят в состав нижней толщи верхнего мела, а чинейвеемские слои представляют собой верхнюю часть кривореченской свиты, которую согласно перекрывает дуговская свита.
СТРАТИГРАФИЧЕСКОЕ ПОЛОЖЕНИЕ ГРЕБЕНКИНСКИХ СЛОЕВ ОТНОСИТЕЛЬНО ВУЛКАНОГЕННЫХ ОБРАЗОВАНИЙ В ВЕРХОВЬЯХ РЕК АНАДЫРЬ И БОЛЬШОЙ АНЮЙ
Как уже отмечалось, стратотипическая местность кривореченской свиты показана на листах геологических карт Q-59-XXI, XXII (Государственная…, 1977). С запада к ним примыкают листы карты Q-59-XIX, XX, составленные М.В. Кривоносовым и Э.Б. Невретдиновым в 1978 г. (Государственная…, 1981). На границе этих листов безымянная нижняя толща верхнего мела получает название “вилковская”. Средняя толща верхнего мела становится нижней подсвитой еропольской свиты, а верхняя толща верхнего мела – верхней подсвитой еропольской свиты.
Такая корреляция почти не оставляет простора для дискуссии о соотношении вилковской толщи и нижней толщи верхнего мела – это разные названия одного и того же стратифицированного геологического тела. Лишь в единственном случае – на левобережье р. Кавральянская – небольшой изолированный выход вулканитов вилковской толщи показан примыкающим к полю выходов верхней толщи верхнего мела, сложенной вулканитами такого же состава. На листах геологических карт Q-59-XIII, XIV, XIX, XX, XXI, XXII (рис. 2) показано, что стратифицированное геологическое тело, на большей части территории называемое “вилковская толща”, в верховьях р. Убиенка перекрывает кривореченскую свиту и содержит остатки туронских аммонитов, а в верховьях рек Большой Пеледон и Большой Анюй залегает на базальтовой саламихинской толще.
Саламихинская толща с угловым несогласием перекрывает отложения готерива и чимчемемельской свиты (Государственная…, 1981). По более ранним данным В.Ф. Белого (1977), чимчемемельская и подстилающая ее айнахкургенская свиты перекрыты мощными вулканическими накоплениями яблонской серии. В междуречье Яблон–Чимчемемель серия разделяется на саламихинскую, нкулилэкинскую и вилковскую толщи. Саламихинская толща содержит Birisia onychioides (Vassil. et K.-M.) Samyl., Onychiopsis cf. elongate (Geyler) Yok., Heilungia tschuktschorum Samyl. et Philipp., Taeniopteris sp., Sphenobaiera longifolia (Pom.) Florin, Phoenicopsis sp. indet., Podozamites cf. lanceolatus (Lindl. et Hutt.) F. Braun, Pityophyllum nordenskioldii (Heer) Nath. (определения А.Ф. Ефимовой, В.А. Самылиной, Г.Г. Филипповой). Нкулилэкинская толща содержит Birisia onychioides (Vassil. et K.-M.) Samyl., Coniopteris sp., Ginkgo ex gr. adiantoides (Ung.) Heer, Phoenicopsis sp., Podozamites sp. indet., Pityophyllum nordenskioldii (Heer) Nath. (определения А.Ф. Ефимовой). В вилковской толще встречены Baiera cf. polimorpha Samyl., Sphenobaiera longifolia (Pom.) Florin, S. angustiloba (Heer) Florin, S. aff. uninervis Samyl., Pityophyllum sp. indet. (определения А.Ф. Ефимовой).
Далее Белый (1977, с. 15) отмечает: “Из нерасчлененных вулканогенных образований яблонской серии на междуречье Олой–Яблон… К.Б. Куликовым были собраны обильные остатки растений: Equisetites aff. burejensis Heer, Coniopteris compressa Vassil., Tchaunia sp., Cladophlebis sp., Heilungia tschuktschorum Samyl. et Philipp., Heilungia sp., Taeniopteris sp., Pterophyllum sp., Sphenobaiera longifolia (Pon.) Florin, Phoenicopsis ex gr. angustifolia Heer, Ph. ex gr. speciosa Heer, Araucarites sp., Pagiophyllum sp., Cephalotaxopsis cf. magnifolia Font. var successiva Holl., C. cf. micropylla laxa Holl., C. cf. anadyrensis Krysht., Cephalotaxopsis sp., Parataxodium cf. wigginsii Arnold et Lowth., Leptostrobus sp. (определения А.Ф. Ефимовой, В.А. Самылиной и Г.Г. Филипповой)”. На составленных позднее среднемасштабных геологических картах этой территории (листы Q-58-XXI, XXII, XXIII, XXIV) данные вулканогенные образования разделены на саламихинскую и вилковскую толщи.
Яблонскую серию в низовьях рек Пеледон и Яблон (рис. 2) Белый (1977) также рассматривает как нерасчлененную. По его данным, в нижней части ее разреза здесь обнаружены отпечатки Heilungia aff. udensis Koschman, Baiera cf. polymorfa Samyl., Pityophyllum sp. indet., а в средней части собраны Podozamites sp. indet., Pityopyllum sp. indet. (определения А.Ф. Ефимовой). В самой верхней части яблонской серии обнаружены отпечатки Podozamites sp. indet., Cephalotaxopsis cf. sangarensis Vassil., Cephalotaxopsis sp. indet., Pityophyllun nordenskioldii (Heer) Nath. (определения А.Ф. Ефимовой). На современных государственных геологических картах масштаба 1 : 200 000 (листы Q-59-XIX, XX) вулканогенные образования яблонской серии разделены на саламихинскую и вилковскую толщи (Государственная…, 1981).
В верхней части айнахкургенской свиты и в чимчемемельской свите содержатся представительные комплексы остатков растений буор-кемюсской флоры (Самылина, 1974, 1976; Белый, 1977; Решения…, 1978, 2009). “Сходные флористические комплексы с Arctopteris, Birisia onychioides, Nilssoniopteris prynadae, Pagiophyllum rtiangulare, Cephalotaxopsis borealis и Desmyophyllum magnum известны и из вышележащих саламихинской (базальтовой) и вилковской (андезитовой) толщ. По имеющимся сейчас в нашем распоряжении данным, можно думать, что флористические комплексы названных выше стратиграфических единиц… соответствуют одному этапу развития флоры и могут быть включены в буор-кемюсскую стратофлору” (Самылина, 1976, с. 109).
Из приведенной цитаты следует, что Самылина изучила не только остатки растений, указанные в работе Белого (1977), но и какой-то дополнительный материал. К сожалению, списков в своей работе она не приводит, а коллекции, скорее всего, утрачены. Заключение Самылиной о том, что флористические комплексы саламихинской и вилковской толщ относятся к буор-кемюсской флоре, конечно, является предположением. Однако представляется крайне маловероятным, что эти небогатые комплексы могут относиться к какой-то более молодой флоре, поскольку ни в одном из местонахождений не обнаружено остатков покрытосеменных растений.
Таким образом, верхняя часть слоев c буор-кемюсской флорой оказывается на одном стратиграфическом уровне с туронскими аммонитами и двустворками, а также волчинским флористическим комплексом совсем иного систематического состава.
Слои с остатками буор-кемюсской флоры широко распространены как на Северо-Востоке России, так и на Северной Аляске (Самылина, 1976; Герман, 2011). Возраст их в решениях Третьего МРСС определен как ранний–средний альб, то есть более древний, чем возраст гребенкинских слоев (Решения…, 2009) (рис. 1б). Если это действительно так, то кривореченская свита должна залегать выше толщи вулканитов среднего состава, которую называют “вилковской” или “нижней толщей верхнего мела”. Если же допустить, что В.А. Самылина (1976) ошиблась в диагнозе тафофлор вилковской толщи в верховьях р. Большой Анюй и Яблон, что на самом деле тафофлоры эти более молодые, то придется признать наличие между саламихинской и вилковской толщами перерыва с верхов альба до низов турона. Последнее представляется автору еще менее вероятным, чем ошибка палеофитолога.
Саламихинскую толщу подстилают чимчемемельская и айнахкургенская свиты. В нижней части последней содержатся остатки фауны ауцелл аптского или, скорее всего, позднеаптского возраста (Паракецов, Паракецова, 1989; Решения…, 2009). В верхней части айнахкургенской свиты (за пределами района, показанного на рис. 2а) и в отложениях чимчимемельской свиты содержатся представительные комплексы остатков растений буор-кемюсской флоры (Самылина, 1976; Решения…, 1978, 2009). Эти образования, конечно, также являются буор-кемюсскими слоями, точнее, их нижней частью.
Из Решений Третьего МРСС (2009) следует, что слои с чаунской флорой располагаются на уровне нижней части гребенкинских слоев – в верхнем альбе. Однако на листе Q-59-IX геологической карты – к северу от рассматриваемой территории – нижняя подсвита еропольской свиты переименована в пыкарваамскую свиту, а верхнееропольская подсвита названа коэквуньской свитой. Из отложений данных свит в пределах Пегтымельского прогиба известны многочисленные находки растений чаунской флоры (Белый, 1977; Щепетов, 1991; Филиппова, Абрамова, 1993). Эти данные свидетельствуют о том, что гребенкинские слои никак не могут располагаться на уровне чаунских и тем более выше их по разрезу.
ЗАКЛЮЧЕНИЕ
Проведенный анализ картографических и литературных данных позволяет сделать вывод о том, что наиболее вероятным стратиграфическим аналогом гребенкинских слоев (кривореченской свиты, исключая верхи) является вулканогенная саламихинская толща, включающая комплекс остатков растений буор-кемюсской флоры. Из этого следует, что на территории Северо-Востока России с конца альба по начало турона одновременно существовали две флоры – буор-кемюсская и гребенкинская. Причем первая, возможно, продолжала существовать и в середине туронского века, когда гребенкинская палеофлора уже сменилась другой флорой, которую представляют волчинский и чинейвеемский комплексы.
Рассмотренные здесь материалы являются общедоступными, и автор будет только рад, если на их основе кто-то сможет сделать иной вывод. Если же возражений не последует, то придется признать возможность сосуществования мезофитной буор-кемюсской флоры и кайнофитной гребенкинской флоры, в которой уже доминировали покрытосеменные растения.
Ранние покрытосеменные растения, безусловно, были эксплерентами (Герман, 2011). Можно предположить, к примеру, что их семена в большинстве своем содержали значительный запас питательных веществ. Это делало их мало мобильными – на значительные расстояния они разносились только водой. Морская вода и принесла откуда-то с востока семенной материал к северо-западному берегу бассейна Пенжинского прогиба. В конце альба здесь началась стремительная “отсыпка” приморских низменностей. Покрытосеменные растения вместе с сопутствующими им таксонами голосеменных немедленно заселили ювенильные поверхности. Не имея конкурентов, они дали всплеск численности и разнообразия – сформировалась гребенкинская флора. Продвинуться вглубь материка это сообщество не смогло: все реки здесь текли в море, а дальше на сотни километров простирались безводные поля лав и базальтовой тефры, подобные Толбачинскому Долу на современной Камчатке. За этими полями вокруг внутренних бассейнов Айнахкургенской и Чимчемемельской впадин продолжала существовать буор-кемюсская флора. В туроне покрытосеменные растения приобрели большую “мобильность” (может быть, им стали помогать насекомые и животные?), однако в полной мере освоить вулканические поля так и не смогли.
Благодарности. За помощь в работе над статьей автор выражает глубокую признательность Т.Б. Русаковой (СВКНИИ ДВО РАН) и А.Б. Герману (Геологический институт РАН).
Источники финансирования. Работа выполнена в рамках темы госзадания Ботанического института РАН “Ископаемые растения России и сопредельных территорий: систематика, филогения, палеофлористика и палеофитогеография” (АААА-А19-119021190031-8), при поддержке Российского фонда фундаментальных исследований (проект № 19-05-00121).
Список литературы
Белый В.Ф. Стратиграфия и структуры Охотско-Чукотского вулканогенного пояса. М.: Наука, 1977. 171 с.
Герман А.Б. Этапность и цикличность развития позднемеловой флоры Анадырско-Корякского субрегиона // Стратиграфия. Геол. корреляция. 1993. Т. 1. № 1. С. 87–96.
Герман А.Б. Меловая флора Анадырско-Корякского субрегиона (Северо-Восток России): систематический состав, возраст, стратиграфическое и флористическое значение. М.: ГЕОС, 1999. 122 с.
Герман А.Б. Альбская–палеоценовая флора Северной Пацифики. М.: ГЕОС, 2011. 280 с. (Тр. ГИН РАН. Вып. 592).
Головнева Л.Б., Щепетов С.В., Алексеев П.И. Чинганджинская флора (поздний мел, Северо-Восток России): систематический состав, палеоэкологические особенности и стратиграфическое значение // Чтения памяти А.Н. Криштофовича. 2011. Вып. 7. С. 37–61.
Государственная геологическая карта СССР масштаба 1 : 200 000. Серия Анадырская. Лист Q-59-XXI, XXII. Объяснительная записка. Составитель Трунов Б.Д. Магадан, 1977. 69 с.
Государственная геологическая карта СССР масштаба 1 : 200 000. Серия Еропольская. Лист Q-59-XIX, XX. Объяснительная записка. Составители Кривоносов В.М., Невретдинов Э.Б. Москва: Союзгеолфонд, 1981. 89 с.
Дворянкин А.И., Лебедев Е.Л., Филатова Н.И. Чинейвеемский флористический комплекс на Северо-Востоке России (правобережье р. Анадырь) и его геологическое значение // Докл. АН. 1993. Т. 333. № 4. С. 472–476.
Девятилова А.Д., Невретдинов Э.Б., Филиппова Г.Г. Стратиграфия верхнемеловых отложений бассейна среднего течения р. Анадырь // Геология и геофизика. 1980. № 12. С. 62–70.
Невретдинов Э.Б. Новые данные о возрасте верхнемеловых вулканогенных образований левобережья верховьев р. Анадырь // Материалы по геологии и полезным ископаемым Северо-Востока СССР. Вып. 25. Магадан: Севвостокгеология, 1980. С. 226–228.
Паракецов К.В., Паракецова Г.И. Стратиграфия и фауна верхнеюрских и нижнемеловых отложений Северо-Востока СССР. М.: Недра, 1989. 298 с.
Решения 2-го Межведомственного регионального стратиграфического совещания по докембрию и фанерозою Северо-Востока СССР (Магадан, 1974–1975 гг.). Магадан: ГКП СВТГУ, 1978. 192 с.
Решения Третьего Межведомственного регионального стратиграфического совещания по докембрию, палеозою и мезозою Северо-Востока России (Санкт-Петербург, 2002). СПб.: Изд-во ВСЕГЕИ, 2009. 268 с.
Самылина В.А. Раннемеловые флоры Северо-Востока СССР (К проблеме становления флор кайнофита) // XXVII Комаровские чтения. Л.: Наука, 1974. 56 с.
Самылина В.А. Меловая флора Омсукчана. Л.: Наука, 1976. 207 с.
Самылина В.А. Аркагалинская стратофлора Северо-Востока Азии. Л.: Наука, 1988. 131 с.
Стратиграфический кодекс России. 3-е издание. Отв. ред. Жамойда А.И. СПб.: ВСЕГЕИ, 2006. 96 с.
Терехова Г.П. О возрасте кривореченской свиты и гребенкинского флористического комплекса // Стратиграфия и палеонтология фанерозоя Северо-Востока СССР. Магадан: СВКНИИ ДВО АН СССР, 1988. С. 100–117.
Филиппова Г.Г., Абрамова Л.Н. Позднемеловая флора Северо-Востока России. М.: Недра, 1993. 348 с.
Щепетов С.В. Среднемеловая флора чаунской серии (Центральная Чукотка). Магадан: СВКНИИ ДВО АН СССР, 1991. 145 с.
Щепетов С.В. Стратиграфия континентального мела Северо-Востока России. Магадан: СВКНИИ ДВО РАН, 1995. 122 с.
Щепетов С.В., Герман А.Б., Белая Б.В. Среднемеловая флора правобережья р. Анадырь. Магадан: СВКНИИ ДВО РАН, 1992. 165 с.
Herman A.B., Golovneva L.B., Shczepetov S.V., Grabovsky A.A. The Late Cretaceous Arman Flora of Magadan Oblast, Northeastern Russia // Stratigraphy and Geol. Correlation. 2016. V. 24. № 7. P. 651–760. https://doi.org/10.1134/S0869593816070029
Spicer R.A., Herman A.B., Ahlberg A., Valdes P.J., Hoffman Ch.-Ch. Plants and climate: Recent developments with CLAMP and its applications in Earth system science // Сборник памяти члена-корреспондента АН СССР, профессора В.А. Вахрамеева (к 90-летию со дня рождения). Ред. Ахметьев М.А., Герман А.Б., Долуденко М.П., И-гнатьев И.А. М.: ГЕОС, 2002. С. 126–127.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция