

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 2, стр. 77-102
Род Arcticoceras Spath, 1924 (Ammonoidea) и инфразональная биостратиграфия нижнего бата Бореальной области
Д. Н. Киселев *
Педагогический университет им. К.Д. Ушинского
Ярославль, Россия
* E-mail: dnkiselev@mail.ru
Поступила в редакцию 11.11.2018
После доработки 30.11.2018
Принята к публикации 28.03.2019
Аннотация
Анализ изменчивости по таксономически значимым признакам рода Arcticoceras (Cardioceratide, Arctocephalitinae) позволяет свести в синонимику A. ishmae большую часть ранее выделенных видов этого рода (в том числе и A. harlandi), характерных для раннего бата. Приводится обоснование невалидности рода Greencephalites, виды которого рассматриваются как кадиконические морфы различных видов Arctocephalites и Arcticoceras. Результаты ревизии рода Arcticoceras позволяют упразднить зону Harlandi, которая присутствует в биостратиграфических шкалах различных регионов Арктики. Инфразональный объем зоны Ishmae Бореального (Арктического) стандарта включает как минимум три биогоризонта, индексами которых могут быть подвиды A. ishmae.
ВВЕДЕНИЕ
Род Arcticoceras Spath, 1924 имеет большое значение для стратиграфии морского бореального нижнего и среднего бата Панбореальной надобласти, включая центральную и северную части Европейской России, Новую Землю, Северную Сибирь (бассейн р. Лена), Северо-Восточную Сибирь (побережье Охотского моря), Землю Франца-Иосифа, Шпицберген, Арктическую Канаду (Канадский Арктический архипелаг, Юкон), Северную Аляску и Восточную Гренландию. Этот род включает следующие виды и подвиды с пригодными названиями:
тип рода Arcticoceras ishmae (Keyserling) (лектотип: Сакс, 1976, табл. IX , фиг. 1а, 1б; Репин и др., 2006, табл. 13 , фиг. 1а–1г);
A. ishmae tenuicostatum Repin (голотип: Репин и др., 2006, табл. 8 , фиг. 5);
A. kochi Spath (голотип: Spath, 1932, pl. XV, fig. 1);
A. michaelis Spath (голотип: Spath, 1932, pl. XIII, figs. 3a, 3b);
A. pseudolamberti Spath (лектотип: Spath, 1932, pl. XIV, fig. 2);
A. stepankovi Tuchkov (голотип: Тучков, 1954, табл. III, фиг. 5);
A. excentricum Voronetz (лектотип: Воронец, 1962, табл. XIV , фиг. 1);
A. harlandi Rawson (голотип: Rawson, 1982, pl. II, figs. 3, 4);
A. cranocephaloide Callomon (голотип: Callomon, 1985, pl. I, fig. 1).
К непригодным видам и подроду, чьи описания содержатся только в неопубликованном отчете, относятся A. lenaense Voronetz (Воронец, Ершова, 1963, табл. V , фиг. 1, табл. VI , фиг. 1–3), A. (Incostarcticoceras) tenuicostatum Voronetz (Воронец, Ершова, 1963, табл. III, фиг. 1–5) и A. (I.) intercostatum Voronetz (Воронец, Ершова, 1963, табл. I , фиг. 1–3). К такой же категории относится вид A. crassiplicatum Callomon, а также вариации A. ishmae α и β, упоминаемые в публикациях (Callomon, 1993; Митта и др., 2011; Mitta et al., 2014, 2015), но не описанные по правилам Международного кодекса зоологической номенклатуры (МКЗН).
Среди пригодных видов наиболее часто используются A. ishmae, A. cranocephaloide, A. harlandi и A. kochi. Они являются индексами зон или слоев с фауной нижнего и среднего бата. Зона Ishmae ранее выделялась в базальной части нижнего келловея Русской платформы (Решения…, 1955) или Печорской плиты (Бодылевский, 1960; Меледина, 1987), а в настоящее время установлена в нижнем бате Европейской России (Митта, 2009 и др.). Также она определена в среднем или верхнем бате различных регионов Арктической Канады (Frebold, 1961; Poulton, 1987 и др.), Восточной Гренландии (Callomon, 1984, 1993 и др.), Северной Сибири (Меледина, 1994). Зона Kochi первоначально выделена в Восточной Гренландии (Donovan, 1953; Callomon, 1959), затем в части Арктической Канады (Frebold, 1964) и Северной Сибири (Меледина, 1973, 1987). По виду-индексу A. harlandi изначально был определен фаунистический горизонт в Арктической Канаде (Callomon, 1984), а затем и зона в том же регионе (Poulton, 1987) и Северной Сибири (Меледина, 1991, 1994). Фаунистический горизонт harlandi позднее был установлен в Восточной Гренландии (Callomon, 1993) и Европейской России (Митта, Сельцер, 2002 и др.). Зона Cranocephaloide уверенно выделяется только в Восточной Гренландии (Callomon, 1979 и др.) и Северной Сибири (Меледина, 1991).
Другие виды Arcticoceras использовались в качестве индексов биостратиграфических подразделений в единичных случаях. Так, в Северо-Восточной Сибири выделены слои с A. stepankovi (Меледина, 1973) и зона Stepankovi (Репин, 2002), а в Европейской России биогоризонт A. excentricum (Гуляев, 2013).
Среди вышеперечисленных биостратонов наибольшей стабильностью характеризуется лишь зона Cranocephaloide. Остальные зоны и инфразональные подразделения, выделенные по арктикоцерасам, неоднократно меняли свои названия в различных региональных стратиграфических шкалах. Чаще всего эта смена связана с изменением точки зрения на диагностику видов-индексов тем или иным автором. При этом одни виды перестают рассматриваться как валидные, в частности A. kochi (Callomon, 1984, 1993; Меледина, 1991, 1994) и A. harlandi (Гуляев, 2013), а диагноз других существенно сужается (например, A. ishmae).
Неоднозначная трактовка столь важных для стратиграфии видов может привести к существенным стратиграфическим ошибкам или изменениям в стратиграфической шкале. Например, для севера Европейской России С.В. Мелединой (1994) вместо зоны Ishmae выделено подразделение в ранге “слои с фауной” – слои с A. ishmae и A. harlandi, при этом делается вывод о конденсированности слоев в стратотипическом разрезе на р. Дрещанка. Это решение основано исключительно на факте совместной встречаемости в одних и тех же слоях разреза диахронных видов, которые являются индексами зон Ishmae и Harlandi в Восточной Гренландии и Арктической Канаде. Изучение этих слоев, проведенное автором данной статьи (результаты пока не опубликованы), показало, что они не содержат следов какой бы то ни было конденсации по литологическим признакам (неоднородности напластования, конкреционных горизонтов, специфической сохранности окаменелостей и пр.). В этой связи напрашивается вывод, что вышеуказанные аммониты или определены не совсем корректно, или не имеют диахронного распространения.
Позднее в тех же слоях В.В. Митта (Mitta et al., 2015) найдена последовательность экземпляров, одни из которых, происходящие из нижней части слоя, определены как A. harlandi, а другие, из средней и верхней части слоя, как A. ishmae. Результаты этого исследования противоречат выводу Мелединой о совместной встречаемости этих видов в конденсированном горизонте, но отвечают распределению арктикоцерасов в разрезах Восточной Гренландии и Арктической Канады.
Данный пример позволяет предполагать существование методологических недостатков в диагностике видов рода Arcticoceras, что требует обстоятельного изучения комплекса признаков, которые в той или иной степени рассматриваются как видоспецифичные. Эти признаки характеризуют межвидовые отличия и лучше всего отражены в рубрике “Cравнение” при описании таксонов. Рассмотрим эти отличия в трактовке различных авторов для видов-индексов, диагностика которых наиболее дискуссионна (A. kochi, A. excentricum и A. harlandi).
A. kochi. При первом описании вида A. kochi указывается три отличительных признака (Spath, 1932, p. 53): “Этот вид лучше всего описать как наиболее близкий к A. ishmae, отличающийся главным образом большей шириной раковины, более грубой скульптурой и более резким наклоном ребер.” Той же трактовки отличий придерживался и Х. Фребольд (Frebold, 1961). С.В. Меледина (1973, с. 82) приводит близкий комплекс отличий по скульптуре: “Arcticoceras kochi отличается от A. ishmae (Keys) и A. pseudolamberti (Spath, 1932, p. 55, pl. XIV, fig. 2) характером скульптуры раковины – толстыми, расширяющимися к периферии ребрами на средних оборотах”. Этот же вид от “A. excentricum Voronetz… отличается менее грубыми и широкими ребрами, наклоном ребер вперед и выгибом их к устью на наружной стороне” (Меледина, 1973, с. 82). В более поздней работе тот же автор упоминает и отличия в форме оборотов (Меледина, 1987, с. 88): “A. ishmae (Keys.) отличается от A. kochi Spath меньшей уплощенностью раковины (В/Т = 1.2–1.4 против 1.45 и более у A. kochi), высоким субтреугольным сечением; тонкими, резкими, частыми, более сильно выгнутыми ребрами на боковых сторонах и более сильным выгибом их на вентральной стороне”.
Таким образом, понимание различий между обоими видами у Спэта и Мелединой диаметрально противоположное: по Спэту более вздутой раковиной обладает A. kochi, в то время как у Мелединой эта особенность скорее свойственна A. ishmae. Также у этих авторов не совпадают отличия по величине изгиба ребер на боковой стороне раковины: по Спэту этим признаком в большей мере характеризуется A. kochi, а у Мелединой – A. ishmae.
В настоящее время практически общепринято, что оба вида являются синонимами (Callomon, 1975; Poulton, 1987; Митта, Сельцер, 2002 и др.)
A. excentricum. В авторском описании (Воронец, 1962, с. 46, 47) этот вид “несколько напоминает Arcticoceras ishmae Keyserling (1846), но отличается широким пупком и почти прямым направлением ребер… От A. kochi Spath… отличается широким и эксцентричным пупком, иным направлением ребер и отсутствием восстановленных ребер близ устья”.
В данном описании отличия A. excentricum от A. ishmae и A. kochi практически одинаковые, и, следовательно, исходя из формальной логики, два последних вида таксономических отличий, по сути, не имеют. Этот вывод справедлив, если не учитывать при сравнении признаки эксцентричности умбилика.
Под эксцентричностью пупка Воронец понимала форму умбилика, испытывающего расширение на конечной жилой камере, как это показано на примере Cranocephalites pompeckji var. laevis Spath (Воронец, 1962, с. 34). Этот признак трактуется автором как видоспецифичный ошибочно, поскольку раскручивание умбиликальной спирали – это признак конечной жилой камеры (КЖК) большинства аммонитов, а не таксона. Он проявляется на терминальных раковинах хорошей сохранности у любых видов Arcticoceras.
Видоспецифичность “отсутствия восстановленных ребер” вблизи устья у A. excentricum также сомнительна: эта особенность в одинаковой мере присутствует у большинства арктикоцерасов.
Попытку более точного описания отличий A. excentricum можно увидеть в работе Мелединой (1987, с. 89): “A. excentricum Voron. отличается от A. kochi Spath вздутой раковиной, широкой вентральной стороной, более грубыми и дольше сохраняющимися ребрами”. Первый признак (пропорции поперечного сечения) в качестве отличительного признака для данного вида использовать трудно, поскольку образцы типовой серии A. excentricum сильно деформированы и для измерений ширины оборота совершенно непригодны. Остальные признаки будут разобраны ниже.
A. harlandi. Автором вида его отличия охарактеризованы следующим образом (Rawson, 1982, p. 98): “A. harlandi отличается от других Arcticoceras менее узким вентером, на котором ребра слабо наклонены вперед”. Столь узкий диапазон отличий в той же мере характеризует и другие виды, поэтому пользоваться им для различения таксонов крайне затруднительно. Этот набор отличительных признаков очень близок к таковым A. excentricum, что позволило Пултону предположить, что оба вида могут быть синонимами (Poulton, 1987, p. 50). В качестве дополнительного отличительного признака Пултон рассматривает конечный диаметр раковины, который у лектотипа A. excentricum превышает таковой юконских образцов. Одним из важнейших отличительных признаков этого вида, по Пултону, является низкая частота весьма рельефных (грубых) ребер.
По Мелединой (1987, с. 91), вид “A. harlandi Rawson отличается от A. ishmae (Keys.) вздутой раковиной, близкими значениями высоты и толщины оборотов, более широкими ребрами и более поздним их сглаживанием”, т.е. в той же мере, в какой A. excentricum отличается от A. kochi (см. выше). Такое сходство отличительных признаков не является случайным, если рассматривать как синонимы A. harlandi и A. excentricum, с одной стороны, и A. ishmae и A. kochi, с другой.
Таким образом, большинство трактовок границ вида выглядят как весьма неопределенные. Более детально охарактеризовали отличия A. harlandi В.В. Митта и В.Б. Сельцер (Митта, Сельцер, 2002, с. 21), при этом они пришли к выводу, что такие виды, как A. excentricum и A. stepankovi, настолько близки, что могут рассматриваться как старшие синонимы этого вида (но в работе они помещены в синонимику A. harlandi со знаком вопроса). В качестве отличительных признаков были использованы только параметры формы поперечного сечения на разных стадиях морфогенеза. Также эти авторы предположили, что A. harlandi из бассейна р. Печора, изображенные С.В. Мелединой, “могут происходить из одного слоя с A. ishmae (Keyserling) и, возможно, являются лишь вздутыми морфами последнего вида” (Митта, Сельцер, 2002, с. 21). Несмотря на это, в более поздних работах В.В. Митта (Mitta et al., 2015) выделил в разрезах на р. Дрещанка горизонт A. harlandi, самостоятельный по отношению к вышележащему горизонту A. ishmae.
Общий набор видоспецифических признаков, упомянутых различными исследователями, приведен в табл. 1.
Таблица 1.
Частота использования (упоминания) видоспецифических признаков при сравнении видов Arcticoceras
| Видоспецифический (отличительный) признак или параметр | A. ishmae (Keys.) | A. ishmae tenuicostatum Repin | A. kochi Spath | A. stepankovi Tuchkov | A. excentricum Voronetz | A. harlandi Rawson | A. lenaense Voronetz (MS) | A. intercostatum Voronetz (MS) | A. tenuicostatum Voronetz (MS) | Всего | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| для всех видов | для пригодных видов | ||||||||||
| 1. Выраженность (рельефность) скульптуры и межреберное расстояние | 7 | 1 | 7 | 1 | 3 | 3 | 1 | 1 | 2 | 26 | 19 |
| 2. Форма и направление изгиба ребер на вентральной и боковых сторонах | 4 | 6 | 2 | 5 | 1 | 1 | 1 | 1 | 21 | 16 | |
| 3. Пропорции поперечного сечения раковины или оборота | 7 | 4 | 2 | 3 | 4 | 20 | 20 | ||||
| 4. Сглаживание ребристости на конечной жилой камере | 2 | 1 | 2 | 3 | 8 | 8 | |||||
| 5. Относительный диаметр умбилика | 1 | 2 | 1 | 4 | 4 | ||||||
| 6. Эксцентричность пупка | 1 | 2 | 1 | 4 | 4 | ||||||
| 8. Конечный диаметр раковины | 1 | 1 | 1 | 3 | 3 | ||||||
| 7. Восстановление ребер вблизи конечного устья | 1 | 1 | 2 | 2 | |||||||
| 9. Характер рассеченности лопастной линии | 1 | 1 | 2 | 2 | |||||||
Примечание. Подсчеты основаны на следующих работах: Spath, 1932; Тучков, 1954; Frebold, 1961; Воронец, 1962; Ефимова и др., 1968; Меледина, 1973, 1987; Rawson, 1982; Poulton, 1987; Митта, Сельцер, 2002; Репин и др., 2006. Признаки расположены в порядке возрастания суммарной частоты. Для подсчета использовались лишь публикации, в которых присутствует раздел с таксономическими описаниями. Каждое упоминание признака фиксировалось в таблице одновременно для обоих сравниваемых видов.
Как видно, большинство специалистов важнейшим видоспецифическим признаком у Arcticoceras признают особенности ребристости и, в меньшей степени, относительную высоту или ширину раковины (оборота). Если же учитывать только пригодные виды, то важнейший признак – относительная высота оборота. В этой связи цель настоящей работы состоит в том, чтобы выяснить, какое значение имеют эти признаки для диагностики. В литературе неоднократно указывалось (Callomon, 1993; Митта, Сельцер, 2002) на их высокую вариабельность, препятствующую надежному распознаванию видов арктоцефалитин. Поэтому можно предполагать, что принципы демаркации видов, рассмотренные выше, в целом не способны выявить реальные таксономические различия. Этому способствует и то обстоятельство, что в изучении Arcticoceras ряд признаков не используется вообще, например коэффициент ветвления ребер. Также упомянутые виды практически не изучены и не сравнивались по указанным признакам на разных стадиях морфогенеза, обсуждались лишь особенности строения их взрослых оборотов.
Изучение изменчивости видоспецифических признаков позволит разработать критерии сходства и отличия видов и оценить реальное морфологическое разнообразие видов внутри рода. Для оценки изменчивости составлена база данных, которая включает измерения 90 экземпляров Arcticoceras по стандартным морфологическим признакам. Эта база данных включает 31 экземпляр из личных сборов и музейных коллекций из разрезов Тимано-Печорского района и 59 экземпляров, изображенных в различных публикациях, в том числе все номенклатурные типы. Дополнительно измерены некоторые Arctocephalites Spath (24 экз.) и Greencephalites Repin (7 экз.), в основном номенклатурные типы и топотипы.
ОТНОСИТЕЛЬНАЯ ВЫСОТА ОБОРОТА (ОВО)
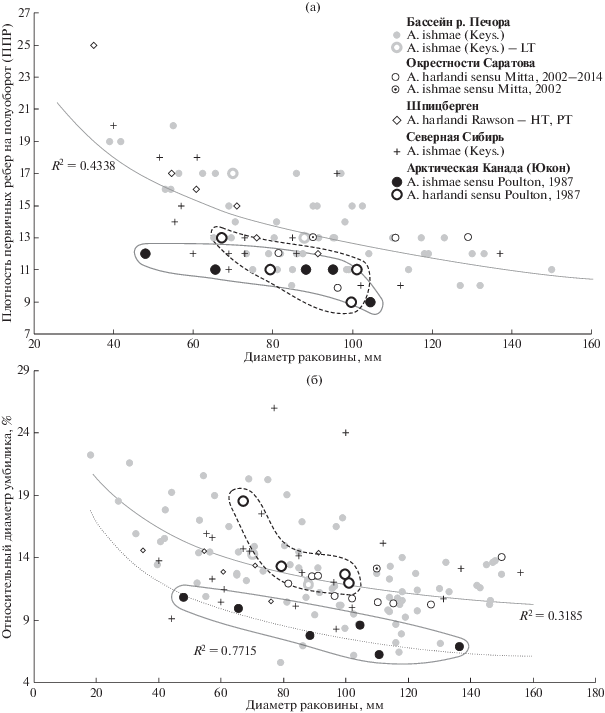
Этот признак, вычисляемый как отношение высоты оборота к его ширине, является одним из наиболее используемых параметров, отражающих пропорции поперечного сечения раковины. Для анализа распределения было получено 95 измерений по 47 экземплярам Arcticoceras из Европейской России (бассейн р. Печора и окрестности Саратова) и 67 измерений по 48 экземплярам Arcticoceras, Greencephalites и типовых Arctocephalites из арктических разрезов Восточной Гренландии, Шпицбергена, Арктической Канады, Северной и Восточной Сибири. Часть измерений сделана по фотографиям, поэтому их значения могут незначительно отличаться от оригинальных.
Наиболее представительная выборка измерений (73) характеризует арктикоцерасов бассейна р. Печора (разрезы по рекам Печора, Уса, Адзьва, Ижма и Дрещанка), откуда происходят номенклатурные типы A. ishmae – типового вида рода Arcticoceras. Поэтому картина изменчивости ОВО для печорской выборки является наиболее показательной среди других видов данного рода, происходящих из зоны Ishmae. Выборки из других регионов незначительны по объему и фрагментарны, поэтому по ним можно судить об изменчивости лишь условно. В этой связи оценка изменчивости и таксономического значения рассматриваемого признака будет осуществляться по выборке печорской популяции.
Арктикоцерасы из бассейна р. Печора характеризуются широким диапазоном изменчивости ОВО (табл. I , фиг. 1–8 ; табл. II, фиг. 1–4): от сильно уплощенных в боковом направлении дискоконических оборотов (ОВО = 1.6) до сильно вздутых сфероконических оборотов (ОВО около 0.52). Такая амплитуда значений показывает весьма высокую изменчивость (коэффициент вариации 13.2%). Она наблюдается при различном диаметре раковины: от ранних оборотов (около 20 мм) до конечной жилой камеры (от 110 до 160 мм). Это свидетельствует о том, что никакой тенденции в изменении пропорций раковины в морфогенезе у арктикоцерасов не происходит. В целом у большинства экземпляров (если не у всех) ОВО с возрастом практически не меняется в морфогенезе, поэтому те или иные пропорции раковины закладываются на ранних стадиях развития и затем сохраняются до терминальной стадии.
Таблица I . Ряды изменчивости Arcticoceras ishmae (бассейн р. Печора). а – вид сбоку, б – вид с вентральной стороны. Длина масштабной линейки 10 мм. 1а, 1б – Arcticoceras ishmae (Keys.) morpha ishmae (Keys.), экз. ЯрГПУ № АДЗ-9, Республика Коми, р. Адзьва, разрез Никифорова Щелья, нижний бат, зона Ishmae; 2а, 2б – Arcticoceras ishmae (Keys.) morpha stepankovi Tuchkov, часторебристая разновидность, экз. ЯрГПУ № АДЗ-13, Республика Коми, р. Адзьва, разрез Никифорова Щелья, нижний бат, зона Ishmae; 3а, 3б – Arcticoceras ishmae (Keys.) morpha stepankovi Tuchkov, редкоребристая разновидность, экз. ЦНИГР Музей № 1/1370, Республика Коми, р. Ижма, разрез у порога Съелом, нижний бат, зона Ishmae, экземпляр, описанный Д.Н. Соколовым (1912, с. 15, рис. 1) под названием Macrocephalites krylowi Milach.; 4а, 4б – Arcticoceras ishmae (Keys.) morpha pseudishmae (Spath), экз. ЯрГПУ № АДЗ-10, Республика Коми, р. Адзьва, разрез Никифорова Щелья, нижний бат, зона Ishmae; 5а, 5б – Arcticoceras ishmae (Keys.) morpha ishmae (Keys.), экз. ЦНИГР Музей № 12/274, колл. А.Н. Замятина, Республика Коми, левый берег р. Ижма, порог Разливной, нижний бат, зона Ishmae; 6а, 6б – Arcticoceras ishmae (Keys.) morpha ishmae (Keys.), разновидность с низким поперечным сечением, экз. ЦНИГР Музей № 21/274, колл. А.Н. Замятина, Республика Коми, левый берег р. Ижма, порог Разливной, нижний бат, зона Ishmae; 7а, 7б – Arcticoceras ishmae (Keys.) morpha stepankovi Tuchkov, экз. ЦНИГР Музей № 18/274, колл. А.Н. Замятина, Республика Коми, левый берег р. Ижма, порог Разливной, нижний бат, зона Ishmae; 8а, 8б – Arcticoceras ishmae (Keys.) morpha pseudishmae (Spath), экз. ЦНИГР Музей № 26/274, колл. А.Н. Замятина, Республика Коми, левый берег р. Ижма, порог Разливной, нижний бат, зона Ishmae.
Здесь и в табл. II, III: ЯрГПУ – Геологический музей им. А.Н. Иванова (Ярославский государственный педагогический университет); ЦНИГР Музей – Центральный научно-исследовательский геологоразведочный музей им. академика Ф.Н. Чернышева (Санкт-Петербург).

Таблица II. Arcticoceras ishmae из разрезов бассейна р. Печора и Северной Сибири. Все фигуры даны в натуральную величину. Длина масштабной линейки 10 мм. Звездочкой обозначено начало жилой камеры. 1 – Arcticoceras ishmae (Keys.) morpha pseudishmae (Spath), экз. ЯрГПУ № АДЗ-10, Республика Коми, р. Адзьва, разрез Никифорова Щелья, нижний бат, зона Ishmae, вид сбоку и с вентральной стороны см. в табл. I , фиг. 4; 2 – Arcticoceras ishmae (Keys.) morpha stepankovi Tuchkov, экз. ЦНИГР Музей № 1/1370, Республика Коми, р. Ижма, разрез у порога Съелом, нижний бат, зона Ishmae, вид сбоку и с вентральной стороны см. в табл. I , фиг. 3; 3 – Arcticoceras ishmae (Keys.) morpha ishmae (Keys.), разновидность с низким поперечным сечением, экз. ЯрГПУ № АДЗ-337, Республика Коми, р. Адзьва, разрез Никифорова Щелья, нижний бат, зона Ishmae; 4 – Arcticoceras ishmae (Keys.) morpha ishmae (Keys.), разновидность с высоким поперечным сечением, экз. ЯрГПУ № АДЗ-12, Республика Коми, р. Адзьва, разрез Никифорова Щелья, нижний бат, зона Ishmae; 5 – Arcticoceras ishmae (Keys.) morpha stepankovi Tuchkov, конечная жилая камера с апертурой, лектотип Arcticoceras excentricum Voronetz, экз. ЦНИГР Музей № 38/9209, изображен в работе Н.С. Воронец (1962, табл. XIV , фиг. 1) с противоположной стороны, р. Хатыстах – левый приток р. Лена.

Таблица III. Arcticoceras ishmae из разрезов низовьев р. Лена. Длина масштабной линейки 10 мм. 1 – Arcticoceras ishmae (Keys.) morpha ishmae (Keys.), оригинал Arcticoceras lenaense Voronetz (MS), экз. № 13и, изображен в неопубликованной рукописи Н.С. Воронец и Е.С. Ершовой (1963, табл. 7 , фиг. 1), обнажение 13, р. Лена, приток Согуру-Тигие; 2 – Arcticoceras ishmae (Keys.) morpha ishmae (Keys.), оригинал Arcticoceras (Incostarcticoceras) tenuicostatum Voronetz (MS), экз. № 106р, изображен в неопубликованной рукописи Н.С. Воронец и Е.С. Ершовой (1963, табл. 3 , фиг. 1), обнажение 106, р. Лена, приток Атыркан; 3 – Arcticoceras ishmae (Keys.) morpha ishmae (Keys.), оригинал Arcticoceras (Incostarcticoceras) tenuicostatum Voronetz (MS), экз. № 106и, изображен в неопубликованной рукописи Н.С. Воронец и Е.С. Ершовой (1963, табл. 3 , фиг. 2), обнажение 106, р. Лена, приток Атыркан; 4 – Arcticoceras ishmae (Keys.) morpha stepankovi Tuchkov, лектотип Arcticoceras excentricum Voronetz, экз. ЦНИГР Музей № 38/9209, изображен в работе Н.С. Воронец (1962, табл. XIV , фиг. 1) с противоположной стороны без части оборота, тот же вид с полным оборотом см. в табл. II, фиг. 5, р. Хатыстах – левый приток р. Лена; 5 – Arcticoceras ishmae (Keys.) morpha stepankovi Tuchkov, оригинал Arcticoceras (Incostarcticoceras) intercostatum Voronetz (MS), экз. № 63-9a, изображен в неопубликованной рукописи Н.С. Воронец и Е.С. Ершовой (1963, табл. 1, фиг. 2), обнажение 63, р. Хатыстах – левый приток р. Лена.

Распределение вариантов ОВО по экземплярам арктикоцерасов из бассейна р. Печора соответствует нормальному закону и, следовательно, является однородным (рис. 1а, 2). Между образцами с полярными значениями пропорций поперечного сечения оборота наблюдаются плавные ряды переходных форм для каждой стадии морфогенеза. Эти ряды получены из выборок популяций разных районов бассейна р. Печора – из разрезов на р. Ижма (табл. 1) и р. Адзьва (табл. 2). Поэтому, несмотря на столь высокую амплитуду значений данного признака, все экземпляры из данной выборки, даже с маргинальными значениями параметра, следует относить к одному виду и рассматривать не более чем морфы или вариации.
Рис. 1.
Распределение экземпляров Arcticoceras ishmae (Keys.) из разрезов бассейна р. Печора (р. Ижма и р. Адзьва) по относительной высоте оборотов (а) и относительному диаметру умбилика (б). Эмпирическое распределение показано гистограммой, теоретическое – гауссовой кривой.
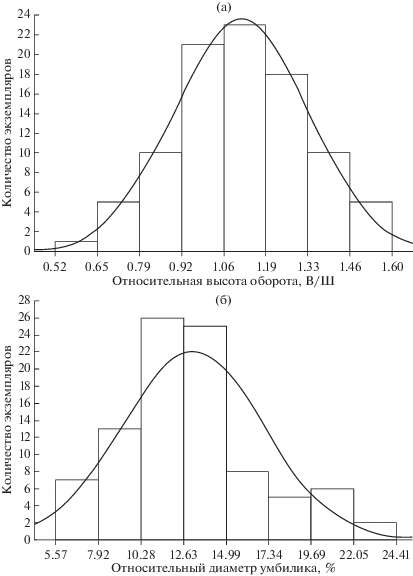
Таблица 2.
Размер выборки и распределение Arcticoceras и других арктоцефалитин по группам плотности скульптуры по номинальному числу измерений (N) и в процентах
| Таксоны арктоцефалитин | Плотность первичных ребер | |||||
|---|---|---|---|---|---|---|
| низкая <12 ребер |
средняя –НОРМА 12–15 ребер |
высокая >15 ребер |
||||
| N | % | N | % | N | % | |
| A. ishmae (Keys.) Европейской России | 11 | 21.5 | 29 | 56.9 | 11 | 21.5 |
| A. ishmae (Keys.) бассейна р. Печора | 10 | 21.3 | 25 | 53.2 | 12 | 25.5 |
| A. ishmae (Keys.) окрестностей Саратова | 1 | 20 | 4 | 80 | 0 | 0 |
| A. ishmae (Keys.) sensu Mitta1 (бассейн р. Печора, Саратов) | 1 | 16.7 | 2 | 33.3 | 3 | 50 |
| A. harlandi Rawson sensu Mitta1 (бассейн р. Печора, Саратов) | 3 | 30 | 7 | 70 | 0 | 0 |
| A. kochi Spath, типовая серия, В. Гренландия | 1 | 50 | 1 | 50 | 0 | 0 |
| Arcticoceras Северной и Восточной Сибири | 3 | 18.75 | 9 | 56.2 | 4 | 25 |
| A. harlandi Rawson, типовая серия, Шпицберген | 0 | 0 | 3 | 50 | 3 | 50 |
| Arcticoceras Арктической Канады | 6 | 75 | 2 | 25 | 0 | 0 |
| A. ishmae (Keys.) sensu Poulton, 1987 | 2 | 100 | 0 | 0 | 0 | 0 |
| Arcticoceras harlandi Rawson sensu Poulton, 1987 | 3 | 75 | 1 | 25 | 0 | 0 |
| A. cranocephaloide Call., типовая серия, В. Гренландия | 1 | 25 | 2 | 50 | 1 | 25 |
| Greencephalites верхнего байоса Арктики | 2 | 28.6 | 5 | 71.4 | 0 | 0 |
Экземпляры с высокими оборотами (ОВО > 1) в целом соответствуют лектотипу A. ishmae, поэтому могут относиться к типичным разновидностям данного вида – A. ishmae morpha ishmae (табл. I , фиг. 1, 5, 6; табл. II, фиг. 3, 4). Экземпляры с примерно изометричными обротами (ОВО = 1) неоднократно описывались в литературе под различными названиями, но чаще всего не относились к A. ishmae. В частности, подобная форма из порога Съелом р. Ижма была описана в работе Д.Н. Соколова (1912, с. 14, 15) под названием Macrocephalites krylowi Milach. (табл. I , фиг. 3; табл. II, фиг. 2). Позднее Мелединой (1987) было показано, что данный экземпляр относится к роду Arcticoceras, а по видоспецифическим признакам он наиболее близок к A. harlandi. По мнению В.В. Митта (Митта, Сельцер, 2002, с. 20, 21), он в полной мере может быть определен как A. harlandi.
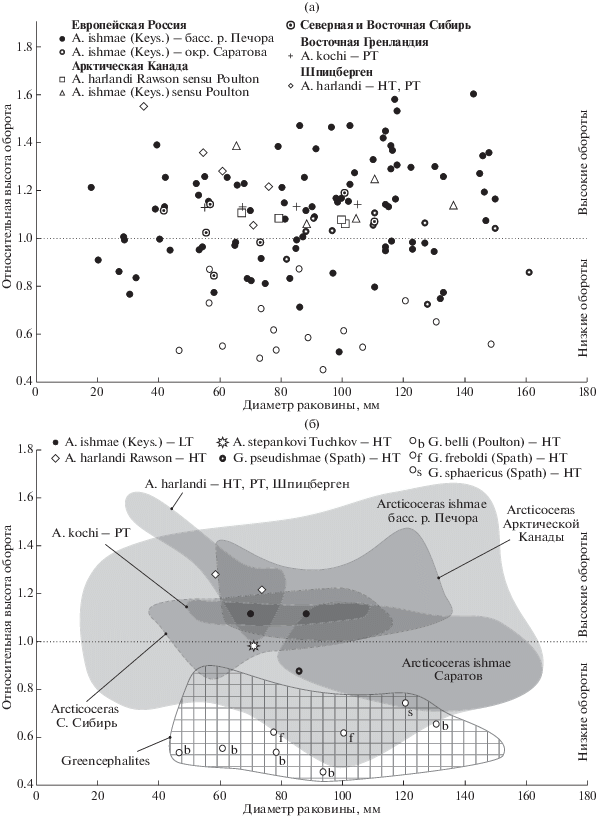
Между тем ни один из типовых экземпляров A. harlandi, изображенных в статье Раусона (Rawson, 1982, pl. 2, figs. 1–5), не характеризуется вздутыми или изометрическими средними или взрослыми оборотами (рис. 2). Более того, голотип этого вида имеет более высокие обороты, чем лектотип A. ishmae, а другие образцы из типовой серии отличаются еще более высокими значениями ОВО, чем голотип. В этой связи представление о вздутом морфотипе A. harlandi, который рассматривается как наиболее типичный для данного вида некоторыми авторами (но не автором вида), следует считать ошибочным.
Рис. 2.
Распределение экземпляров арктоцефалитин (Arcticoceras и “Greencephalites”) верхнего байоса, нижнего и среднего бата по относительной высоте оборотов и диаметру раковины. (а) – все экземпляры; (б) – номенклатурные типы и морфологические ареалы. Сплошной заливкой показаны морфологические ареалы Arcticoceras, сетчатой заливкой – Greencephalites. HT – голотип, LT – лектотип, PT – паратип.
Наиболее близок к экземпляру Соколова голотип вида A. stepankovi, как по форме оборотов, так и по скульптуре. Поэтому вариации A. ishmae с таким морфотипом будут обозначены в данной работе как A. ishmae morpha stepankovi (табл. I , фиг. 2, 3, 7).
Экземпляры с очень низкими, часто кадиконическими оборотами встречаются в выборках арктоцефалитин зоны Ishmae весьма редко. У Спэта (Spath, 1932) подобные формы отнесены к роду Cadoceras, а в современных работах они чаще всего определяются как представители родов Arctocephalites (Митта, Сельцер, 2002, табл. 4 , фиг. 1; Митта, 2009, табл. 2, фиг. 2) и Greencephalites (Репин и др., 2007, с. 134; Митта, Альсен, 2013, фототаблица, фиг. 2; Mitta et al., 2015, fig. 7). Только Дж. Калломон (Callomon, 1993, p. 100) такие экземпляры рассматривал как экстремально вздутую морфу A. ishmae. К данному морфотипу он отнес ранее выделенный гренландский вид A. pseudishmae (Spath) (голотип: Spath, 1932, pl. VIII, fig. 6), происходящий из арктикоцерасовых слоев (зона Ishmae, фауна 16, по Callomon, 1993). В бассейне р. Печора такие экземпляры найдены в разрезах по рекам Адзьва (табл. I , фиг. 4; табл. II, фиг. 1) и Ижма (табл. I , фиг. 5) и обозначаются здесь как A. ishmae morpha pseudishmae. Вероятно, к этой же морфе следует относить все экземпляры со вздутыми оборотами из зоны Ishmae окрестностей Саратова (Митта, Сельцер, 2002, табл. 4 , фиг. 1; Mitta et al., 2015, fig. 7).
Остальные арктоцефалитины из саратовских разрезов (Сокурский карьер), определяемые как A. ishmae и A. harlandi в работах В.В. Митта (Митта, Сельцер, 2002; Митта, 2009; Mitta et al., 2014), здесь рассматриваются как вышеупомянутые вариации A. ishmae. Поле изменчивости саратовских арктикоцерасов (по 10 измерениям) не распространяется за пределы поля A. ishmae бассейна р. Печора и занимает в основном правую его часть. Семь из десяти измерений характеризуют экземпляры с высокими оборотами, свойственными A. ishmae morpha ishmae. К той же морфе по ОВО могут быть отнесены и все типовые экземпляры A. kochi и, в меньшей степени, Arcticoceras из Арктической Канады (Юкон), описанные Пултоном (Poulton, 1987) как A. ishmae и A. harlandi. Однако отличия по другим признакам (см. ниже) позволяют рассматривать юконские экземпляры как самостоятельные хроноподвиды A. ishmae.
В целом можно констатировать, что признак, характеризующийся ОВО, не имеет большого значения для диагностики видов арктикоцерасов. Незначительные отличия среднестатистического параметра у отдельных популяций в различных регионах Панбореальной надобласти, вероятно, носят случайный характер, обусловленный малым размером выборки. Соответственно, несколько выделенных видов (A. kochi, A. harlandi, A. stepankovi) следует считать младшими синонимами A. ishmae.
ОТНОСИТЕЛЬНЫЙ ДИАМЕТР УМБИЛИКА (ОДУ)
Этот параметр, выражающий процентное отношение диаметра умбилика к диаметру раковины, редко рассматривается как видоспецифический признак в роде Arcticoceras, в первую очередь для раннебатских видов (табл. 1). Между тем арктоцефалитины позднего байоса (Greencephalites) и Arcticoceras среднего бата отличаются от раннебатских видов вполне отчетливо весьма широким умбиликом на всех стадиях морфогенеза (рис. 3). В особенности это характерно для среднебатского A. cranocephaloide, у которого не наблюдается перекрытия морфологического поля ни с одним экземпляром более древних арктикоцерасов.
Рис. 3.
Распределение экземпляров арктоцефалитин (Arcticoceras и Greencephalites) верхнего байоса, нижнего и среднего бата по относительному диаметру умбилика и диаметру раковины. (а) – все экземпляры; (б) – номенклатурные типы и морфологические ареалы. Сплошной заливкой показаны морфологические ареалы Arcticoceras, сетчатой заливкой – Greencephalites. HT – голотип, LT – лектотип, PT – паратип.
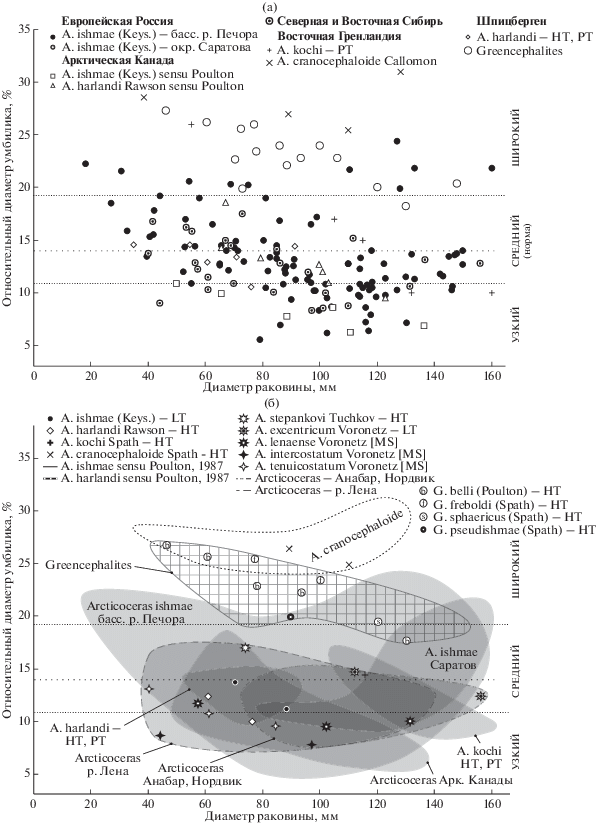
Раннебатские Arcticoceras бассейна р. Печора, к которым относится, в первую очередь, типовой материал A. ishmae, характеризуются наибольшей выборкой (82 измерения по 44 экземплярам) по сравнению с выборками из других регионов (рис. 3а) и, как следствие, более высокими параметрами изменчивости ОДУ (коэффициент вариации 23.7%). Несмотря на высокую амплитуду изменчивости (ОДУ от 5.5 до 24.4%, мода 13.13%), в популяции трудно выделить какие-либо дискретные группировки, которые можно было бы рассматривать как разные таксоны. В этой связи все экземпляры этой выборки следует относить к A. ishmae. Как и в случае ОВО, распределение является нормальным (рис. 1б), а в морфогенезе размер умбилика не изменяется в рамках какого-либо тренда. У отдельных экземпляров прослеживается незначительное уменьшение диаметра умбилика на взрослых оборотах, однако на конечной жилой камере большинство раковин испытывает разворачивание умбиликальной спирали и, соответственно, расширение умбилика.
Остальные виды раннебатских арктикоцерасов практически не выходят за пределы поля изменчивости A. ishmae (рис. 3б). Максимальным диаметром умбилика обладают наиболее вздутые формы типа A. ishmae morpha pseudishmae и A. ishmae morpha stepankovi, в то время как экземпляры с более высокими оборотами максимально инволютны. Типовые экземпляры A. ishmae, A. kochi и A. harlandi почти в одинаковой степени характеризуются значениями, близкими к средним, по обоим рассмотренным параметрам формы раковины. Можно утверждать, что эти признаки составляют пару ковариации, что обусловливает их близкое распределение.
ПЛОТНОСТЬ ПЕРВИЧНЫХ РЕБЕР (ППР)
Этот признак считается большинством исследователей важнейшим для диагностики видов Arcticoceras. Он коррелятивно связан с рельефностью скульптуры, поэтому большинство редкоребристых экземпляров имеют более груборебристый морфотип, чем часторебристые.
Измерения плотности ребер проводились для полуоборота, поэтому некоторые экземпляры характеризуются двумя измерениями. Тем не менее количество измерений по этому признаку существенно меньше, чем по признакам формы раковины. Так, выборка печорской популяции представлена 51 измерением по 38 экземплярам, а суммарное число измерений всех арктикоцерасов составляет 97. Уменьшение числа измерений связано с тем, что часть экземпляров на терминальной стадии развития лишены скульптуры. Также частоту ребер не всегда удается измерить вследствие плохой сохранности образца.
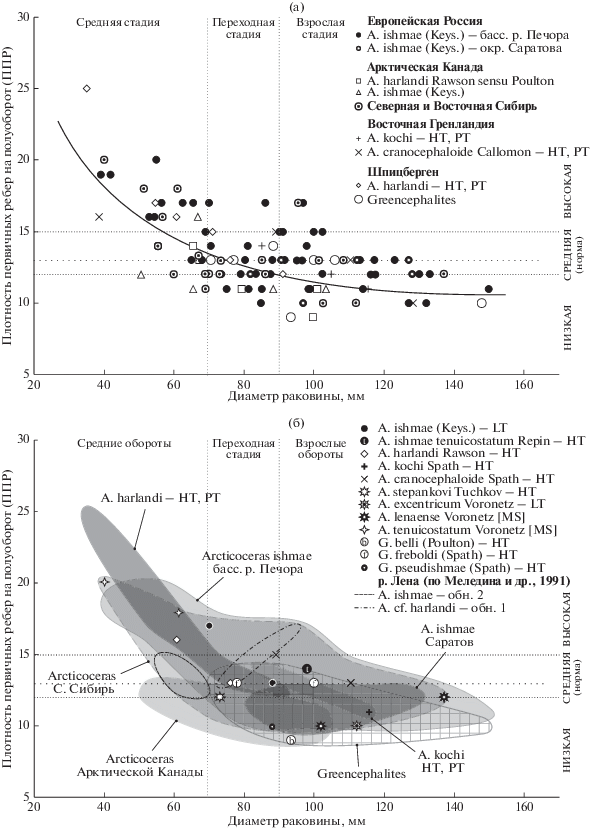
В отличие от ранее рассмотренных признаков, плотность ребер изменяется в морфогенезе закономерно, образуя морфогенетический тренд: при меньшем диаметре раковины плотность ребер максимальна у большинства изученных групп, а на взрослых оборотах число ребер существенно снижается (рис. 4). Наблюдаемый тренд характеризуется хорошей корреляцией (r = 0.64) и степенной зависимостью (R2 = 0.47), что позволяет провести периодизацию морфогенеза по данному признаку – определить пограничную область между средними и взрослыми оборотами. Она отвечает диаметру раковины с диапазоном значений 70–90 мм. В этом интервале находятся номенклатурные типы Arcticoceras ishmae, A. harlandi, A. pseudishmae и A. stepankovi, что облегчает сравнение и диагностику этих видов при переходном диаметре раковины.
Рис. 4.
Распределение экземпляров арктоцефалитин (Arcticoceras и Greencephalites) верхнего байоса, нижнего и среднего бата по плотности первичных ребер и диаметру раковины. (а) – все экземпляры; (б) – номенклатурные типы и морфологические ареалы. Сплошной заливкой показаны морфологические ареалы Arcticoceras, сетчатой заливкой – Greencephalites. HT – голотип, LT – лектотип, PT – паратип.
Уменьшение плотности первичных ребер с возрастом наблюдается у большинства изученных экземпляров, причем это хорошо заметно даже в пределах одного оборота. Изменение ребристости на всех стадиях развития не исследовалось, поскольку имеющийся материал не пригоден для полного разворачивания раковины. Поэтому рассмотренная картина развития плотности скульптуры в основном является статистической. Тем не менее она хорошо показывает, что (а) использование малочисленной выборки без учета морфогенетического тренда и изменчивости может привести к ошибкам в определении; (б) изучение распределений по ППР следует производить отдельно как минимум для взрослых и средних оборотов, в противном случае распределение обязательно будет дискретным даже в пределах одного вида. К сожалению, в нашем распоряжении материала недостаточно для такого исследования, поэтому тип распределения по отдельным стадиям в настоящий момент установить невозможно.
Частота ребристости у всех изучаемых арктоцефалитин находится в интервале от 9 до 25 ребер на пол-оборота, что свидетельствует о весьма высокой изменчивости (коэффициент вариации 15.7%). Это значение во многом отражает онтогенетическую изменчивость (по De Baets et al., 2015), поскольку ППР изменяется в морфогенезе. Для отдельных стадий развития показатель изменчивости будет существенно ниже. Так, в интервале Д = 60–80 мм коэффициент вариации составляет примерно 12.15%, а при Д = 100–150 мм оно уже около 9.3%.
Нормальные (средние) значения плотности ребер находятся в интервале от 12 до 15 ребер; соответственно, низкая плотность составляет менее 12 ребер, а высокая – более 15.
Большую часть морфологического пространства плотности скульптуры занимает выборка арктикоцерасов бассейна р. Печора, у которых данный признак изменяется от 10 до 20 ребер на пол-оборота (коэффициент вариации 15%). Около 50% этой выборки составляют экземпляры с нормальной (средней) плотностью ребер (табл. 2).
Материал из других регионов гораздо меньше печорского по числу экземпляров, поэтому дает несколько отличающееся распределение, но в целом морфологические ареалы различных популяций Arcticoceras практически не выходят за пределы морфологического поля печорской выборки. Ближе всего к печорскому находится сибирский материал, что сказывается на общих показателях изменчивости (табл. 2, рис. 4). Морфоареалы популяций остальных регионов находятся или в области взрослых оборотов (арктикоцерасы окрестностей Саратова), или преимущественно в переходной области (оригиналы из Арктической Канады), либо в области средних оборотов и переходной области (типовая серия A. harlandi Rawson). Такое распределение, скорее всего, случайное и обусловлено незначительным объемом выборок. В этой связи наблюдаемые небольшие отличия в ППР различных выборок объясняются не таксономическими отличиями, а изменчивостью в пределах единственного вида A. ishmae. Можно констатировать, что плотность скульптуры, по крайней мере, для раннебатских арктикоцерасов не имеет таксономического значения. Об этом дополнительно свидетельствует и факт совпадения морфоареалов этой группы с позднебайосскими “Greencephalites” и среднебатскими A. cranocephaloide, которые хорошо разделяются по другим признакам – по пропорциям поперечного сечения и относительному диаметру умбилика.
Ошибочность диагностики видов по ППР объясняется и чисто субъективными причинами. Разберем это на примере вида A. harlandi. Типовой материал с Земли Короля Карла характеризуется средними (голотип) и высокими (два паратипа) значениями плотности ребер (табл. 2). Голотип вида почти не отличим по ППР от лектотипа A. ishmae (рис. 4б), а тонкоребристые паратипы обладают наибольшей плотностью из всех измеренных арктикоцерасов. Тем не менее у многих авторов сложилось устойчивое убеждение, что A. harlandi обладает непременно редко- и груборебристым морфотипом в сочетании с низким поперечным сечением, по сравнению с A. ishmae. Это представление было одновременно и независимо сформулировано Пултоном (Poulton, 1987) и Мелединой (1987), а не автором вида, после чего поддержано другими специалистами. Возможно, такое мнение сложилось из-за особого внимания исследователей к одному из паратипов (Rawson, 1982, pl. 2. fig. 7), который действительно обладает более грубыми ребрами. Тем не менее плотность ребер у этого экземпляра находится в средней (нормальной) области, как и у типичных A. ishmae. Характерно, что сам Пултон в выборку A. ishmae поместил экземпляры только с низкой ППР, в то время как у A. harlandi оставлял экземпляры и со средней плотностью (табл. 2).
Низкая плотность ребер наблюдается у экземпляров с различным сочетанием модальности других морфологических признаков, например относительной высоты оборотов. В области низкой плотности находятся типы A. kochi (высокие обороты), A. stepankovi (изометричные обороты) и A. pseudishmae (вздутые обороты) (рис. 4б). Также к этой категории принадлежат типы A. excentricum и A. lenaense, однако пропорции поперечного сечения у них неизвестны в силу плохой сохранности типового материала. Тем не менее можно утверждать, что редкоребристые формы не обязательно обладают низким поперечным сечением, а изменчивость плотности ребер не когерентна с изменчивостью параметров формы раковины. Это также заметно по распределению типовых экземпляров видов Greencephalites, обладающих вздутой раковиной: одни из них характеризуются низкой плотностью ребер (G. belli), другие находятся в области нормы рода Arcticoceras (G. freboldi).
Другим примером недооценки изменчивости видов по ППР может служить диагностика арктикоцерасов в разрезе Атыркан на р. Лена (северная часть Чекуровской антиклинали, по Меледина, 1991), использованная в неопубликованной работе Н.С. Воронец и Е.С. Ершовой (1963). Арктикоцерасы собраны из одного горизонта, подошвы слоя 1, при этом редкоребристые и часторебристые экземпляры отнесены к разным видам, которым даны новые названия – A. intercostatum (редкоребристый вид) и A. tenuicostatum (часторебристый вид) (табл. III, фиг. 2, 3). Обе формы ничем не отличаются от различных морф печорских арктикоцерасов и, по нашему мнению, должны быть отнесены к A. ishmae.
ХАРАКТЕР НАКЛОНА РЕБЕР
Этот признак рассматривается многими специалистами как один из важнейших для диагностики видов рода Arcticoceras (табл. 1). Он может быть выражен как минимум с помощью двух параметров: угла наклона ребер по направлению к устьевой части раковины и кривизной изгиба ребер в привентральной области. У всех арктикоцерасов ребра прорзирадиатные, характеризующиеся небольшим углом наклона, который может меняться от 6° до 30°. При этом величина наклона ребер прямо пропорциональна их кривизне: более наклоненные ребра серповидно изогнуты, как это выражено у типичных разновидностей A. ishmae или A. kochi, в то время как ребра с более радиальным направлением обычно спрямленные, как у форм, часто определяемых как A. harlandi или A. excentricum. Таким образом, эти признаки составляют ковариационную пару.
Изучение наиболее массового материала из нижнебатских арктикоцерасовых слоев из бассейна р. Печора показало, что параметры изгиба ребер незначительно коррелируют с формой поперечного сечения оборотов (r = 0.53): раковины с высокими оборотами покрыты более наклоненными и серповидно изогнутыми ребрами, в то время как экземпляры с низким поперечным сечением характеризуются спрямленными и более радиальными ребрами. Таким образом, эти признаки также образуют пару ковариации. Как было показано выше, относительная высота оборота сильно варьирует в пределах вида и не имеет значения для диагностики арктикоцерасов. В этой связи любые вариации формы изгиба ребер можно рассматривать исключительно как проявление изменчивости, варианты которой позволяют характеризовать отдельные морфы A. ishmae. Наибольшим изгибом ребер обладает A. ishmae morpha ishmae (табл. I , фиг. 5), средняя и минимальная степень изгиба свойственна A. ishmae morpha stepankovi (табл. I , фиг. 3, 7) и A. ishmae morpha pseudishmae (табл. I , фиг. 8 ). В пределах каждой морфы также наблюдается небольшая изменчивость изгиба ребер. Так, для тонкоребристых экземпляров в целом характерен наиболее сильный изгиб ребер, однако нередки варианты со спрямленными ребрами. С другой стороны, у экземпляров с очень низкими оборотами (ОВО = 0.5–0.6) наклон ребер может быть несколько выше (табл. I , фиг. 4), чем у отдельных экземпляров с изометричными оборотами. Такую же изменчивость можно наблюдать и у байосских арктоцефалитин, характеризующихся вздутым морфотипом оборотов. Так, у Arctocephalites ex gr. sphaericus Spath наклон ребер может сильно меняться независимо от высоты оборотов: у паратипа вида (Spath, 1932, pl. XVI, fig. 5) ребра прямые и субрадиальные, а у форм, обозначенных как A. cf. sphaericus Spath (Callomon, 1979, pl. 1, figs. 2, 3), они имеют сильный наклон, но без характерного выгиба в сторону устья.
Таким образом, изгиб ребер не имеет большого значения для диагностики видов арктикоцерасов. В такой же мере это может относиться и к другим арктоцефалитинам.
СГЛАЖИВАНИЕ РЕБРИСТОСТИ НА КОНЕЧНОЙ ЖИЛОЙ КАМЕРЕ
У Arcticoceras, как и у большинства арктоцефалитин, на терминальном обороте и конечной жилой камере происходит редукция скульптуры вплоть до полного ее исчезновения. При этом область терминального оборота, покрытая скульптурой, может по размеру отличаться у разных видов, поэтому угловая длина этой области может иметь диагностическое значение. В частности, С.В. Меледина предполагала, что A. harlandi отличается от A. ishmae, помимо комплекса других отличительных признаков, более поздним сглаживанием ребер. Вероятно, этот признак действительно может быть видоспецифичным при условии стабильности его проявления в морфогенезе, обусловленной низкой изменчивостью.
Для проверки этой гипотезы изучена выборка Arcticoceras на стадии терминальной раковины, у которых измерена длина гладкой и скульптированной частей в угловых величинах. Наибольшую часть выборки составляют экземпляры из бассейна р. Печора, на ее основе можно судить о масштабах и диапазоне изменчивости этого признака. Среди арктикоцерасов печорской выборки встречаются экземпляры совершенно гладкие (Lr = 0°) и частично ребристые (360° > Lr > 0°), при этом отсутствуют экземпляры, полностью покрытые скульптурой (Lr = 360°). Длина скульптированной части терминального оборота изменяется от 0° до 169°, среднее значение составляет около 77.7°. Изменчивость этого признака очень высока (коэффициент вариации 64%).
Таким образом, положение точки сглаживания скульптуры у печорских A. ishmae изменяется в таких широких пределах, что вряд ли может рассматриваться как таксономический признак. У Arcticoceras различных регионов Арктики этот признак также не постоянен. Так, у груборебристых экземпляров, которые часто определяются как A. harlandi или A. excentricum, стадия гладкой раковины может занимать большую часть оборота, а не половину оборота, что хорошо заметно у лектотипа A. excentricum (табл. II, фиг. 5). Тем не менее не исключено, что в хорошей выборке у стратиграфически или географически различных форм могут быть найдены статистические отличия по этому признаку. Поскольку редукция скульптуры напрямую связана с терминальной стадией развития, она характеризует также и изменчивость скорости морфогенеза. Более подробно этот вопрос будет рассмотрен в отдельной статье.
ОБСУЖДЕНИЕ
Результаты изучения изменчивости арктикоцерасов, рассмотренные выше, позволяют уточнить таксономический состав и филогенетические взаимоотношения поздних арктоцефалитин, а также критически оценить некоторые модели систематики этой группы аммонитов. Эти уточнения позволяют внести определенные изменения в стратиграфические схемы нижнего и среднего бата Бореальной области.
О видовом составе рода Arcticoceras
Среди перечня признаков, которые принято рассматривать как видоспецифические для Arcticoceras, диагностическое значение имеют лишь некоторые из них, в первую очередь относительный диаметр умбилика и размеры ребристой зоны на терминальном обороте. По этим признакам среднебатские A. cranocephaloide надежно отделяются от раннебатских арктикоцерасов. Среди последних явно выраженных различий, которые превышали бы порог изменчивости по каждому признаку, не наблюдается. Ни один из признаков, перечисленных в табл. 1, не пригоден для выделения более чем одного вида в пределах этой группы. Незначительные отличия между популяциями из различных регионов Арктики существенно меньше общего диапазона изменчивости A. ishmae бассейна р. Печора и носят в основном случайный характер. В этой связи арктикоцерасы из зон Harlandi и Ishmae, выделяемых в нижнем бате на большей части Панбореальной надобласти, следует рассматривать в объеме единственного вида A. ishmae. Различные вариации этого вида рассматриваются здесь как морфы, обозначенные пригодными, но невалидными названиями. Они являются не более чем формальными обозначениями, отражающими формы внутривидовой изменчивости. Названия морф являются условными, поскольку не регламентируются правилами МКЗН. Тем не менее для обозначения каждой из них выбраны приоритетные наименования. К таким морфам относятся следующие:
A. ishmae (Keys.) morpha ishmae (Keys.). Соответствует лектотипу вида и лучше всего различается на средних и взрослых оборотах, не достигших стадии гладкой раковины. Пропорции поперечного сечения на этих стадиях высокие, ребристость плотная или умеренно плотная, ребра серповидно изогнутые в сторону устья и характеризуются большим наклоном. Конечный оборот сильно инволютный с незначительным разворотом умбилика и высоким поперечным сечением. Этой морфе соответствует большая часть типового материала A. harlandi, в том числе и голотип, оригинал A. ishmae tenuicostatum Repin и типовой материал A. kochi. Сюда же относятся экземпляры из бассейна р. Лена, обозначенные Н.С. Воронец как A. lenaense Voronetz (MS) и A. tenuicostatum Voronetz (MS). Эта морфа численно преобладает в имеющихся выборках и поэтому может рассматриваться как норма вида.
A. ishmae (Keys.) morpha stepankovi Tuchkov. Редкоребристая морфа с изометричным поперечным сечением оборотов. Ребра спрямленные, характеризуются небольшими кривизной и углом наклона. К этой морфе относится голотип A. stepankovi, типовые оригиналы A. excentricum и экземпляры из различных регионов, часто определяемые как A. harlandi. К ним принадлежат A. harlandi sensu Meledina (Меледина, 1987, табл. 31 , фиг. 1), A. harlandi sensu Mitta (Митта, Сельцер, 2002, табл. 1, фиг. 1, табл. 2; Митта, 2009, табл. 2, фиг. 3; Митта и др., 2011, табл. 1, фиг. 5; Mitta et al., 2014, fig. 6-1; Mitta et al., 2015, fig. 6-3). Из видов с непригодными названиями к этой же морфе, вероятно, следует относить экземпляры A. intercostatum Voronetz (MS) из бассейна р. Лена.
A. ishmae (Keys.) morpha pseudishmae (Spath). Это разновидность с низкими и часто вздутыми, кадиконическими, оборотами, весьма широким умбиликом, относительно редкой ребристостью, состоящей из спрямленных, слабо наклоненных ребер. К этой морфе Ю.С. Репин и В.В. Митта отнесли несколько груборебристых, с низким поперечным сечением экземпляров из нижнего бата разрезов р. Ижма и окрестностей Саратова, которые, как они считают, принадлежат к видам родов Arctocephalites и Greencephalites (Митта, Сельцер, 2002, табл. 4 , фиг. 1; Mitta et al., 2015; figs. 6–7).
Две последние морфы занимают подчиненное положение по отношению к численности первой морфы, которая рассматривается как норма вида. Не исключено, что численное соотношение этих морф может меняться во времени или пространстве. Если такое смещение нормы вида будет зафиксировано статистически, то это может послужить достаточным основанием для выделения хроноподвидов или географических подвидов, однако на имеющемся материале такого рода процедуру осуществить в настоящий момент невозможно.
Род Greencephalites и происхождение кадоцератин
В род Greencephalites Repin, 2007 (типовой вид Cadoceras freboldi Spath, 1932) его автор поместил несколько видов поздних арктоцефалитин из пограничных слоев бореального верхнего байоса и нижнего бата, характеризующихся в основном кадиконическим типом раковины на всех стадиях морфогенеза. Позднее В.В. Митта (Митта, Альсен, 2013; Mitta et al., 2015) и Д.Б. Гуляев (2013) включили в этот род несколько форм из разрезов нижнего бата Европейской России (см. выше). Основной признак данного рода, отличающий его от изохронных ему Arctocephalites и Arcticoceras, характеризующихся более высоким поперечным сечением, – пропорции раковины или относительная высота оборота. Все остальные признаки в той или иной степени присутствуют у других подродов. При таком подходе основным принципом родовой систематики поздних арктоцефалитин следует считать, по сути, их разделение на таксоны, отличающиеся широкими раковинами, и таксоны с высокими раковинами. При этом роды с более высокими раковинами (Arctocephalites и Arcticoceras) диахронны, поскольку сменяют друг друга в нескольких зонах верхнего байоса–нижнего бата, а род с широкими раковинами (Greencephalites) имеет совместное распространение с вышеперечисленными родами. Таким образом, относительная высота оборота в данном случае рассматривается как важнейший родоспецифический признак.
В общих чертах этот подход к систематике лег в основу новой версии происхождения кадоцератин, предложенной В.В. Митта (2016). Согласно этой гипотезе, подсемейство Cadoceratinae Hyatt, представители которого характеризуются кадиконическим морфотипом, происходит от рода Greencephalites, а род Paracadoceras Crickmay, ранее многими исследователями относившийся к кадоцератинам, происходит от рода Arcticoceras.
Изучение вариабельности относительной высоты оборота у арктикоцерасов (см. выше) показало, что кадиконические экземпляры в имеющейся выборке представляют весьма редкую морфу изменчивости, а не самостоятельные виды или, тем более, род. Поэтому как минимум все кадиконические арктоцефалитины из нижнего бата следует относить к роду Arcticoceras. Такого же подхода придерживался Дж. Калломон (Callomon, 1993). Позднебайосские кадиконические арктоцефалитины, такие как Arctocephalites crassum (Madsen), A. sphaericus Spath, A. freboldi (Spath) и A. belli Poulton, вероятно, также следует рассматривать как проявление изменчивости разных видов рода Arctocephalites. Таким образом, для расщепления основного ствола арктоцефалитин на две ветви, от которых происходят разные таксоны, не существует достаточных оснований. Скорее всего, происхождение кадоцератин следует вести от единственного рода Arcticoceras, как это неоднократно постулировалось в различных работах (Callomon, 1985; Callomon, Wright, 1989; Меледина, 1994).
Зона Arcticoceras ishmae и ее распространение в Панбореальной надобласти
Результаты анализа изменчивости Arcticoceras позволяют большинство раннебатских видов свести в синонимику A. ishmae. Среди этих видов присутствуют виды-индексы зон, выделенных, помимо зоны Ishmae, в нижнем бате различных регионов Панбореальной надобласти (рис. 5). В первую очередь это относится к виду A. harlandi. В настоящее время зона Harlandi входит (или предлагается к установлению) в стратиграфические шкалы Арктической Канады (Poulton, 1987), Северной Сибири (Меледина, 1991, 1994), Земли Франца-Иосифа (Репин, 2005, 2007), бассейна р. Печора (Репин, 2005, 2007; Репин и др., 2006, 2007), а также в различные версии Бореального или Арктического стандарта (Захаров и др., 1997, 2005; Репин, 2005, 2007; Репин и др., 2007; Меледина и др., 2011; Шурыгин и др., 2011; Никитенко и др., 2013). Одновременно разрабатываются другие варианты этой шкалы, где зона Harlandi отсутствует, а в нижнем бате сохраняется лишь одна арктикоцерасовая зона Ishmae (Zakharov, Rogov, 2014).
Рис. 5.
Географическое положение разрезов зоны Arcticoceras ishmae в Панбореальной надобласти. 1 – окрестности Саратова; 2а–2в – разрезы, где Arcticoceras известны только по упоминаниям в литературе, но без изображений: 2а – бассейн р. Лена (Пермский край) (Аркадьев, 2018); 2б – Самарская Лука (Сазонов, 1957); 2в – Елатьма (Сазонов, 1957); 3 – бассейн р. Печора (разрезы на реках Ижма, Дрещанка, Адзьва); 4 – Новая Земля; 5 – Земля Франца-Иосифа; 6 – Шпицберген (земля Короля Карла, Западный Шпицберген); 7 – Баренцево море (Бассейн Тромсё); 8 – Северное море (скважина 211/21-1, по Callomon, 1975); 9 – Восточная Гренландия; 10 – Анабарский залив: 11 – окрестности низовьев р. Лена (Оленекский залив; р. Лена, Оленекская протока); 12 – побережье Охотского моря (р. Вилига); 13 – Северная Аляска (разрезы Canning River, по Imlay, 1976); 14 – Юкон (разрез Salmon Cache Canyon); 15 – Канадский Арктический архипелаг (о. Принца Патрика, по Frebold, 1961).
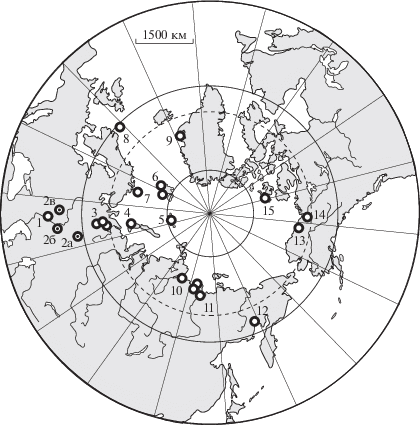
В этой связи представляет несомненный интерес ответ на вопрос, насколько обосновано выделение или установление зоны Harlandi в различных регионах.
Шпицберген. На Земле Короля Карла (разрез D827 острова Свенскёйа), откуда происходит типовой материал A. harlandi, все находки данного вида (пять экземпляров) приурочены к подошве пачки Дюнефелля (Rawson, 1982). В данной пачке не выявлено какой-либо последовательности арктикоцерасов. Известны и другие разрезы с арктикоцерасами на Земле Короля Карла, например на острове Короля Чарльза (Kongsøya) в основании пачки Ретциюсфелля (Smith et al., 1976). Арктикоцерасы в этом разрезе представлены крупными раковинами хорошей сохранности (Smith et al., 1976, table 3 ), однако они так и не были описаны, а их последовательность также не выявлена. В этой связи выделение зоны Harlandi в типовом местонахождении как минимум преждевременно. Также этому препятствует морфологическая близость типов Раусона экземплярам вида A. ishmae из типовой местности (бассейн р. Печора). Как было показано выше, типовой материал A. harlandi в основном относится к A. ishmae morpha ishmae, что автоматически делает ненужным выделение новой арктикоцерасовой зоны.
Бассейн р. Печора. В бассейне р. Ижма, откуда происходит тип A. ishmae, наиболее представительные разрезы нижнего бата находятся на р. Дрещанка (правый приток р. Ижма). Несмотря на то, что из этих разрезов С.В. Мелединой и В.В. Митта упоминались и изображались находки как A. ishmae, так и A. harlandi, смена этих видов не доказана ни для каких разрезов. Согласно Мелединой (1987, 1994), оба вида встречаются совместно в одних и тех же слоях, что препятствует выделению не только зоны Harlandi, но и зоны Ishmae в разрезах. Позднее Митта (Mitta et al., 2015, fig. 3) показал, что некая смена этих видов в пачке III существует. При этом A. harlandi отмечается в подошве пачки, а A. ishmae – в верхней ее части. Тем не менее автором не приводится ни одного изображения арктикоцерасов, найденных in situ в лучшем разрезе (№ 5), где такая последовательность могла бы быть зафиксирована. Все изображения принадлежат экземплярам из разных разрезов (№ 3, 5, 6, 9) либо найденным в осыпи или в слое, но без указания на привязку относительно кровли или подошвы. В той же работе (Mitta et al., 2015) отмечается, что в разрезах на самой р. Ижма (порог Бычье Горло) оба вида встречаются в одном слое и горизонте, что опять же препятствует установлению зоны Harlandi или каких-либо подразделений внутри зоны Ishmae.
Окрестности Саратова. Здесь, в первую очередь в Сокурском разрезе, слои с арктикоцерасами располагаются, по представлениям В.В. Митта, выше зоны Besnosovi (хотя вместе с арктикоцерасами были встречены ораницерасы). В основании толщи глин с арктикоцерасами, ниже “белемнитового уровня”, отмечаются находки A. harlandi, а в средней и верхней части этой толщи, на “белемнитовом уровне” и выше – A. ishmae (Митта, Сельцер, 2002; Митта и др., 2011; Mitta et al., 2014). Эти слои отнесены к зоне Ishmae, в которой установлены биогоризонты harlandi, ishmae α и ishmae β. Большинство изображенных арктикоцерасов, происходящих из Сокурского разреза, определено как A. harlandi, при этом всего из этого разреза было изображено 7 экземпляров данного вида (Митта, Сельцер, 2002, табл. 1, фиг. 1, табл. 3 , фиг. 3, табл. 5 , фиг. 3, табл. 6 , фиг. 1; Митта и др., 2011, табл. 1, фиг. 5; Mitta et al., 2014, fig. 5 (6), fig. 6 (1)). Из них только 2 образца имеют точную, посантиметровую, привязку к слою. Вид A. ishmae изображен два раза (Митта, Сельцер, 2002, табл. 3 , фиг. 1; табл. 2, фиг. 1). Сравнение изображений экземпляров обоих видов и анализ измерений показывает следующее:
(1) Указанные экземпляры A. harlandi sensu Mitta и A. ishmae sensu Mitta характеризуются настолько мелкими различиями, что практически неотличимы друг от друга. Обе формы располагаются в средней и верхней частях морфологического поля изменчивости печорской выборки A. ishmae по ведущим морфологическим признакам (рис. 6), т.е. преимущественно в области нормы этого вида.
Рис. 6.
Распределение экземпляров арктоцефалитин Arcticoceras нижнего бата по плотности первичных ребер (а), относительному диаметру умбилика (б) и диаметру раковины. Морфологические ареалы показаны только для выборок из разрезов Salmon Cache Canyon (Юкон).
(2) Часть определений A. harlandi sensu Mitta больше соответствуют общепринятому диагнозу A. ishmae, т.е. имеют противоречивый характер (например, Митта и др., 2011, табл. 1, фиг. 5). То же самое относится к определению вида A. ishmae, изображенный экземпляр которого больше соответствует устоявшемуся диагнозу A. harlandi (Митта, Сельцер, 2002, табл. 3 , фиг. 1). Это свидетельствует о том, что данные определения весьма произвольны. Не исключено, что упомянутые экземпляры могли быть идентифицированы противоположным образом, если были бы найдены в другом регионе или располагались бы в иной стратиграфической последовательности.
В настоящее время по имеющейся выборке арктикоцерасов в Сокурском разрезе крайне затруднительно проводить какое бы то ни было инфразональное деление зоны Ishmae. На этом основании любые варианты такого деления, имеющиеся в литературе, следует рассматривать как недостаточно обоснованные.
Северная и Восточная Сибирь. Слои нижнего бата, охарактеризованные арктикоцерасами, развиты в низовьях рек Лена и Оленек, а также в пределах Анабарского и Нордвикского структурно-фациальных районов. Зона Harlandi наиболее уверенно устанавливается в низовьях Лены, где она занимает нижнюю часть чекуровской свиты, а в остальных районах зона представлено фрагментарно (Меледина и др., 1991; Меледина, 1991, 1994). Выше зоны Harlandi в чекуровской свите выделяется зона Ishmae. Палеонтологическая характеристика обеих зон представлена арктикоцерасами плохой сохранности, в основном раздавленными ядрами, поэтому об их видовой принадлежности можно судить в основном лишь по особенностям ребристости. Большинство определений аммонитов из обеих зон чекуровской свиты даны в открытой номенклатуре, причем в зоне Harlandi к такому типу относятся все определения – Arcticoceras cf. harlandi (Меледина и др., 1991, табл. 4 , фиг. 1; табл. 4 , фиг. 1) и A. cf. excentricum (Меледина и др., 1991, табл. 2, фиг. 1; табл. 7 , фиг. 1; табл. 8 , фиг. 4).
Анализ признаков изображенных аммонитов позволяет считать разделение арктикоцерасов чекуровской свиты на виды весьма проблематичным. Указанные аммониты характеризуются близкими модальностями видоспецифических признаков, в первую очередь плотности ребер, поэтому среди них можно выделять лишь варианты изменчивости. Более того, нередко арктикоцерасы из зоны Ishmae, определенные как A. ishmae (Меледина и др., 1991, табл. 10 , фиг. 3) или A. cf. ishmae (Меледина и др., 1991, табл. 4 , фиг. 2–5), лучше соответствуют привычному диагнозу A. harlandi Raws. sensu Poulton, 1987 как редкоребристой форме арктикоцерасов. С другой стороны, экземпляры из верхов чекуровской свиты, определенные как A. cf. harlandi, больше соответствуют диагнозу A. ishmae как часторебристой форме (рис. 4б.). По сути, обе формы не имеют принципиальных видовых отличий друг от друга, поэтому их следует рассматривать как один вид. Этот вывод подтверждается данными Н.С. Воронец и Е.С. Ершовой (1963), которые в разрезе Атыркан (северное крыло Чекуровской антиклинали) в подошве чекуровской свиты нашли комплекс арктикоцерасов, которые обнаруживают полный ряд изменчивости по признакам плотности ребер и коэффициенту ветвления. Среди них хорошо различимы A. ishmae morpha ishmae (табл. III, фиг. 1–3) и A. ishmae morpha stepankovi. Обе морфы известны также в Анабарском и Нордвикском районах, откуда они обычно определялись как A. excentricum (табл. II, фиг. 5; табл. III, фиг. 4, 5).
Восточная Гренландия. Среднеюрские разрезы этого региона охарактеризованы бореальными аммонитами с наибольшей детальностью по сравнению с другими регионами Арктики. Дж. Калломон (Callomon, 1993) отнес к зоне Ishmae четыре фаунистических горизонта, выделенных по арктикоцерасам: (14) A. harlandi, (15, 16) A. ishmae (α и β) и (17) A. sp. nov. (crassiplicatum). Таким образом, намечена последовательность, в которой A. harlandi занимает базальное положение в арктикоцерасовых слоях. Этот вид, по Калломону, представлен крупными грубо- и редкоребристыми экземплярами. К сожалению, собранные Калломоном аммониты, к которым относятся индексы этих подразделений, не были изображены или описаны, поэтому предложенное деление до сих пор является декларативным. Переопределения оригиналов Спэта, для которых указаны название морфы и фаунистический горизонт (Callomon, 1993, table 1), не вносят существенной ясности в проблему распознавания хроновариаций гренландских арктикоцерасов. Так, среди макроконховых Arcticoceras, которых Спэт относит к A. kochi, одна часть типовой серии отнесена к A. ishmae α (Spath, 1932, pl. 12, fig. 1; pl. 13, figs. 4, 5; pl. 14, fig. 1), а другая к A. ishmae β (Spath, 1932, pl. 15, fig. 1, голотип). Такому разделению также противоречит сравнительная характеристика этих морф, данная автором (Callomon, 1993, p. 100), где указывается, что морфа A. ishmae β отличается от A. ishmae α более изящной ребристостью, сильным выгибом ребер на вентере и ранним сглаживанием ребер на КЖК. Между тем указанные оригиналы различных морф характеризуются противоположным сочетанием признаков. В частности, наиболее сглаженный по скульптуре вариант отнесен к морфе A. ishmae α (Spath, 1932, pl. 14, fig. 1), а груборебристый к морфе A. ishmae β (Spath, 1932, pl. 15, fig. 1). Возможно, опубликование материала, собранного Калломоном и хранящегося в Геологическом музее университета Копенгагена, поможет подтвердить его стратиграфическую концепцию, но в настоящий момент это осуществить невозможно.
Арктическая Канада. Зона Harlandi выделена впервые Т.П. Пултоном (Poulton, 1987) в разрезе каньона Салмон Кэш, расположенном в долине р. Пьёркупайн (территория Юкон). Аммониты этой зоны найдены в слое 53, выше которого располагается зона Ishmae (слои 55–60). Таким образом, в разрезе наблюдается последовательность арктикоцерасов, в которой базальное положение занимает A. harlandi.
Сравнительно-морфометрическое изучение арктикоцерасов из этих слоев показывает, что, с одной стороны, они в основном не выходят за границы поля изменчивости A. ishmae, а с другой – имеют статистические отличия по двум признакам – относительному диаметру умбилика и частоте ребер (рис. 6). Умбилик у канадских экземпляров в целом у́же (в среднем 10.7%), чем у арктикоцерасов из других регионов (13.2% у печорской выборки, 13.4% у экземпляров со Шпицбергена, 13.8% у экземпляров из Северной Сибири). Также эти формы отличаются меньшей частотой ребер – в среднем ППР = 12 ребер (у печорской выборки ППР составляет 14 ребер, у экземпляров со Шпицбергена – 16 ребер, у экземпляров из Северной Сибири – 14 ребер). При этом A. harlandi sensu Poulton по частоте ребер принципиально не отличается от A. ishmae sensu Poulton относительно совокупной выборки, но существенно отличается от выборки типовой коллекции A. harlandi. Иными словами, между канадскими арктикоцерасами из разных слоев, относимыми к разным видам, различий существенно меньше, чем между канадской популяцией и популяциями других регионов, которые рассматриваются в объеме одного вида A. harlandi.
То же самое относится к экземплярам A. ishmae sensu Poulton. Они достаточно сильно отличаются от нормы печорских A. ishmae очень узким умбиликом, в то время как A. harlandi sensu Poulton по диаметру умбилика вполне сопоставим с A. ishmae печорских разрезов.
Все это показывает, что канадские арктикоцерасы статистически действительно отклоняются от нормы раннебатских арктикоцеросов, т.е. имеют как минимум провинциальные отличия.
С другой стороны, Arcticoceras из слоя 53 разреза каньона Салмон Кэш статистически отличаются от арктикоцерасов слоев 55–60. Разница проявляется только в диаметре умбилика: он существенно больше у базальных арктикоцерасов (12–18.6%, в среднем 14.12%), чем у более молодых форм (6.2–11%, в среднем 8.36%). Последние также характеризуются бо́льшим наклоном ребер (26°–29°), как у нормы печорских A. ishmae. У A. harlandi sensu Poulton это значение находится в интервале 7°–21°. Все это позволяет рассматривать эти формы как реально различающиеся разновидности, на основе которых могут быть выделены самостоятельные таксоны подвидового ранга.
В целом юконские популяции имеют некоторые статистические отличия по ряду признаков как от типовых A. harlandi (выборка слоя 53), так и от нормы A. ishmae (выборка слоев 55–60). Эти отличия весьма отчетливы, хотя и выявлены на небольшом материале – на экземплярах, изображенных Пултоном (Poulton, 1987, pls. 19–23). Несомненно, в более солидной выборке показатели изменчивости будут другими, что несколько снизит наблюдающиеся различия. Но, скорее всего, общая их особенность сохранится. Возможно, эти различия не только провинциальные, но и стратиграфические. В слое 53, помимо A. harlandi sensu Poulton, отмечаются последние Arctocephalites (Poulton, 1987, p. 12), что указывает на относительную древность комплекса арктикоцерасов этого слоя. Вероятно, они относятся к базальной последовательности этого рода. Арктикоцерасы из вышерасположенных слоев того же разреза составляют не менее специфический комплекс, вероятно, более древний, чем слои с A. ishmae типовой местности. Соответственно, последовательность Arcticoceras разреза каньона Салмон Кэш стратиграфически предшествует слоям зоны Ishmae бассейна р. Печора, а также Шпицбергена (островов Короля Карла) и может быть более или менее эквивалентна нижним биогоризонтам в гренландской шкале.
Таким образом, раннебатские арктикоцерасы образуют последовательность как минимум из трех хроновариаций A. ishmae. Исходя из границ изменчивости этих вариаций, их лучше относить к различным подвидам A. ishmae. Последовательность таких подвидов создает основу для инфразонального деления зоны Ishmae для всей Бореальной надобласти. Недостаток данных об изменчивости и географическом распространении не позволяет в полной мере очертить их границы, поэтому в настоящий момент данная последовательность может быть намечена только проспективно (рис. 7). Она имеет следующий вид:
Рис. 7.
Предлагаемый вариант инфразонального деления части бата Бореальной (Арктической) области по арктикоцерасам и его корреляция с различными регионами Арктики. Зоны, обозначенные в схеме, примерно соответствуют части нижнего и среднего бата. Поскольку точное соотношение зон бореальной (арктической) шкалы с зонами бата Северо-Западной Европы до сих пор не установлено, в данной схеме подъярусы бата не показаны. Сплошной заливкой обозначены интервалы высокого обоснования, штриховкой – предполагаемые.
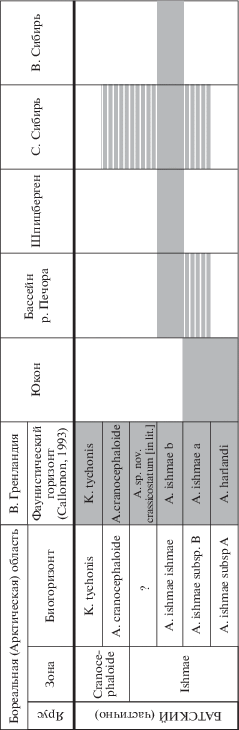
Arcticoceras ishmae (Keys.) subsp. A. Это самый ранний хроноподвид, соответствующий арктикоцерасам из слоя 53 разреза каньона Салмон Кэш Юкона. К нему могут относиться формы из базальной части зоны Ishmae Восточной Гренландии (фауна 14), обозначенные Дж. Калломоном (Callomon, 1993) как A. harlandi. Однако отсутствие их изображений и описаний в литературе оставляет это тождество под вопросом. Присутствие этого подвида в остальных регионах Панбореальной надобласти весьма вероятно, но пока окончательно не доказано. К нему могут быть отнесены некоторые арктикоцерасы из окрестностей Саратова, изображенные в работах В.В. Митта и описанные под названием A. harlandi (Митта, Сельцер, 2002, табл. 1, фиг. 1; Митта и др., 2011, табл. 1, фиг. 5; Mitta et al., 2014, fig. 6[1 ] ). Морфотип саратовских арктикоцерасов, представленных в выборке только экземплярами со взрослыми оборотами, имеет ряд свойств (грубая ребристость, высокий коэффициент ветвления), по которым их можно как относить к этому подвиду, так и рассматривать как проявление изменчивости более молодых подвидов A. ishmae.
Arcticoceras ishmae (Keys.) subsp. B. Соответствует арктикоцерасам из слоев 55–60 разреза каньона Салмон Кэш Юкона. По положению в восточногренландской стратиграфической шкале, ему отвечает форма A. ishmae α, однако и в этом случае, из-за отсутствия изображений соответствующих ей экземпляров, это остается на уровне предположения. Согласно краткой характеристике этой вариации, данной Калломоном, ей свойственны большие размеры терминальной раковины и позднее сглаживание ребер на КЖК. Но в первую очередь она отличается экстремальной инволютностью и минимальной плотностью скульптуры на средних и поздних оборотах, по сравнению с другими арктикоцерасами. В остальных регионах Арктики такие формы, у которых вышеописанный набор признаков проявлялся бы со статистической устойчивостью, не установлены. Предположительно, этот подвид может быть найден в основании арктикоцерасовых слоев в бассейне р. Печора и Северной Сибири, где редкоребристые арктикоцерасы встречаются весьма часто.
Arcticoceras ishmae ishmae (Keys.). Это номинальный подвид, характеризующийся наиболее полной выборкой. Особенности проявления изменчивости позволяют выделить в этом подвиде морфы, подробно рассмотренные выше (A. ishmae morpha ishmae, A. ishmae morpha stepankovi и A. ishmae morpha pseudishmae).
К этому подвиду относится большая часть, если не вся выборка арктикоцерасов из бассейна р. Печора. Синонимом этого подвида является A. harlandi. Соответственно, стратиграфическое положение известных арктикоцерасовых слоев Земли Короля Карла эквивалентно слоям с A. ishmae ishmae, а не базальным горизонтам зоны Ishmae. В Северной и Восточной Сибири к номинальному подвиду относятся большинство, если не все арктикоцерасы.
Данному подвиду соответствует разновидность A. ishmae β, которая рассматривается как индекс одноименного фаунистического горизонта восточно-гренландской шкалы.
ЗАКЛЮЧЕНИЕ
Выборки Arcticoceras раннего бата характеризуются настолько высокой изменчивостью, что по признакам, которые принято считать видоспецифическими, определять таксоны видового или подвидового ранга на единичном материале затруднительно или невозможно. Определение таксонов в пределах рода может быть осуществлено в основном на статистической основе. Таксономическое значение имеют не сами признаки, а статистически устойчивое их поведение в морфогенезе. В этой связи ограниченность материала, характерная для выборок арктикоцерасов большинства высокобореальных регионов, позволяет устанавливать только морфы, а не таксоны. Представители однотипных морф у разных подвидов могут быть неотличимы по морфологическим признакам, поэтому для диагностики таксонов на ограниченном материале не пригодны.
Большая часть пригодных видов Arcticoceras являются синонимами A. ishmae. В этой связи зона Ishmae может быть единственной зоной, выделяемой по арктоцефалитинам в интервале, где ранее были установлены зоны Harlandi и Ishmae. Зона Harlandi является младшим синонимом зоны Ishmae и должна быть упразднена. Инфразональное деление зоны Ishmae включает как минимум три биогоризонта, индексами которых являются подвиды A. ishmae. Из них номинальный подвид занимает верхнюю часть инфразональной шкалы. Выше него может располагаться четвертый биогоризонт, выделенный в Восточной Гренландии по неописанному новому виду или подвиду. Его положение в инфразональной шкале Арктического стандарта до описания этих новых форм исключительно гипотетическое.
Благодарности. Автор благодарит В.А. Захарова, М.А. Рогова и А.П. Ипполитова (ГИН РАН), А.С. Алифирова (ИНГГ СО РАН) за ценные замечания, сделанные при подготовке рукописи.
Источники финансирования. Работа выполнена при поддержке РФФИ (грант 18-05-01070).
Список литературы
Аркадьев В.В. Каталог коллекции к монографии Э.И. Эйхвальда “Lethaea Rossica ou Paléontologie de la Russie”. 1865–1868 (аммониты). СПб.: ЛЕМА, 2018. 168 с.
Бодылевский В.И. Келловейские аммониты Северной Сибири // Записки Ленинградского горного ин-та. 1960. Т. 37. С. 49–82.
Воронец Н.С. Стратиграфия и головоногие моллюски юрских и нижнемеловых отложений Лено-Анабарского района // Тр. НИИГА. 1962. Т. 110. 237 с.
Воронец Н.С., Ершова Е.С. Аммониты и некоторые двустворчатые моллюски верхней юры и нижнего валанжина Лено-Оленекского района. Ленинград, 1963, не опубликовано. 155 с.
Гуляев Д.Б. О зональной аммонитовой шкале верхов байоса, бата и низов келловея Восточно-Европейской платформы // Юрская система России: проблемы стратиграфии и палеогеографии. V Всероссийское совещание: научные материалы. Отв. ред. Захаров В.А. Екатеринбург: ООО “Издательский дом “ИздатНаукаСервис”, 2013. С. 58–64.
Ефимова А.Ф., Кинасов В.П., Паракецов К.В., Полуботко И.В., Репин Ю.С., Дагис А.С. Полевой атлас юрской фауны и флоры Северо-Востока СССР. Магадан: Магаданское книжное изд-во, 1968. 382 с.
Захаров В.А., Богомолов Ю.И., Ильина В.И., Константинов А.Г., Курушин Н.И., Лебедева Н.К., Меледина С.В., Никитенко Б.Л., Соболев Е.С., Шурыгин Б.Н. Бореальный зональный стандарт и биостратиграфия мезозоя Сибири // Геология и геофизика. 1997. Т. 38. № 5. С. 927–956.
Захаров В.А., Б.Н. Шурыгин, С.В. Меледина, М.А. Рогов, Киселев Д.Н., Никитенко Б.Л., Дзюба О.С., Ильина В.И. Бореальный зональный стандарт юры: обсуждение новой версии // Материалы первой Всероссийского совещания “Юрская система России: проблемы стратиграфии и палеогеографии” / Захаров В.А., Рогов М.А., Дзюба О.С. (ред.) М.: ГИН РАН. 2005. с. 88–96.
Меледина С.В. Аммониты и зональная стратиграфия байоса–бата Сибири // Тр. ИГиГ СО АН СССР. 1973. Вып. 153. 152 с.
Меледина С.В. Аммониты и зональная стратиграфия келловея суббореальных районов СССР // Тр. ИГиГ СО АН СССР. 1987. Вып. 691. 182 с.
Меледина С.В. Зональная схема “бореального бата”–нижнего келловея Сибири // Тр. ИГиГ СО АН СССР. 1991. Вып. 769. С. 125–154.
Меледина С.В. Бореальная средняя юра России // Тр. ИГиГ СО РАН. 1994. Вып. 819. 184 с.
Меледина С.В., Шурыгин Б.Н., Злобина О.Н., Левчук М.А., Нальняева Т.Н., Никитенко Б.Л. Чекуровская свита (бат–келловей) в стратотипе // Тр. ИГиГ СО АН СССР. 1991. Вып. 769. С. 5–37.
Меледина С.В., Никитенко Б.Л., Шурыгин Б.Н., Дзюба О.С., Князев В.Г. Современная комплексная зональная шкала юры Сибири и бореальный стандарт // Новости палеонтологии и стратиграфии. 2011. Вып. 16–17. С. 17–40. Приложение к журналу “Геология и геофизика”. Т. 52.
Митта В.В. Верхний байос и нижний бат бассейна Печоры и бореально-тетическая корреляция // Стратиграфия. Геол. корреляция. 2009. Т. 17. № 1. С. 77–87.
Митта В.В. О филогении ранних Cardioceratidae (Ammonoidea) и среднерусских представителях Cadoceratinae на рубеже бата и келловея // Палеонтол. журн. 2016. № 4. С. 42–51.
Митта В.В., Альсен П. Аммониты и зональная шкала батского яруса Гренландии // Юрская система России: проблемы стратиграфии и палеогеографии. V Всероссийское совещание: научные материалы. Отв. ред. Захаров В.А. Екатеринбург: ООО “Издательский дом “ИздатНаукаСервис”, 2013. С. 149–151.
Митта В.В., Сельцер В.Б. Первые находки Arctocephalitinae (Ammonoidea) в юре юго-востока Русской платформы и корреляция бореального батского яруса со стандартной шкалой // Тр. Научно-исслед. ин-та геологии СГУ. Нов. сер. 2002. Т. X. С. 12–39.
Митта В.В., Захаров В.А., Барсков И.С., Сельцер В.Б., Иванов А.В. Верхний байос и нижний бат окрестностей Саратова: малакологическая характеристика и биостратиграфия // Стратиграфия. Геол. корреляция. 2011. Т. 19. № 5. С. 32–45.
Никитенко Б.Л., Шурыгин Б.Н., Князев В.Г., Меледина С.В., Дзюба О.С., Лебедева Н.К., Пещевицкая Е.Б., Глинских Л.А., Горячева А.А., Хафаева С.Н. Стратиграфия юры и мела Анабарского района (Арктическая Сибирь, побережье моря Лаптевых) и бореальный зональный стандарт // Геология и геофизика. 2013. Т. 54. № 8. С. 1047–1082.
Репин Ю.С. Новые аммониты из Печорского верхнего келловея // Палеонтол. журн. 2002. № 5. С. 33–38.
Репин Ю.С. Новые данные по стратиграфии юры Печорской синеклизы // Бюлл. МОИП. Отд. геол. 2005. Т. 80. Вып. 3. С. 17–25.
Репин Ю.С. Аммонитовая шкала Печорской юры // Бюлл. МОИП. Отд. геол. 2007. Вып. 2. С. 24–31.
Репин Ю.С., Захаров В.А., Меледина С.В., Нальняева Т.И. Атлас моллюсков Печорской юры // Бюлл. Палеонтологического и литологического коллективного фонда ВНИГРИ. 2006. № 3. 262 с.
Репин Ю.С., Федорова A.A., Быстрова В.В., Куликова Н.А., Полуботко И.В. Мезозой Баренцевоморского седиментационного бассейна // Стратиграфия и ее роль в развитии нефтегазового комплекса России. Ред. Киричкова А.В., Дмитриева T.В. СПб.: ВНИГРИ, 2007. С. 112–161.
Решения Всесоюзного совещания по разработке унифицированной схемы стратиграфии мезозойских отложений Русской платформы. Л.: Гостоптехиздат, 1955.
Сазонов Н.Т. Юрские отложения Центральных областей Русской платформы. Л.: Гостоптехиздат, 1957. 155 с.
Сакс В.Н. (ред.) Стратиграфия юрской системы севера СССР. М.: Наука, 1976. 436 с.
Соколов Д.Н. К аммонитовой фауне Печорской юры // Тр. Геол. ком. Нов. Сер. 1912. Вып. 76. 65 с.
Тучков И.И. Юрские аммониты и белемниты Северо-Востока СССР // Материалы по геологии и полезным ископаемым Северо-Востока СССР. 1954. Вып. 8. С. 96–125.
Шурыгин Б.Н., Никитенко Б.Л., Меледина С.В., Дзюба О.С., Князев В.Г. Комплексные зональные шкалы юры Сибири и их значение для циркумарктических корреляций // Геология и геофизика. 2011. Т. 52. № 8. С. 1051–1074.
Callomon J.H. The ammonite zones of the Middle Jurassic beds of East Greenland // Geol. Mag. 1959. V. XCVI. № 6. P. 505–513.
Callomon J.H. Jurassic ammonites from the northern North Sea // Norsk Geologisk Tidsskrift. 1975. V. 55. P. 373–386.
Callomon J.H. Marine boreal Bathonian fossils from the northern North Sea and their palaeogeographical significance // Proc. Geol. Assoc. 1979. V. 90. № 4. P. 163–169.
Callomon J.H. A review of the biostratigraphy of the post-Lower Bajocian Jurassic ammonites of the western and northern North America // Geol. Assoc. Can. Spec. Pap. 1984. № 27. P. 143–174.
Callomon J.H. The evolution of the Jurassic ammonite family Cardioceratidae // Palaeontology. Spec. Pap. 1985. № 35. P. 49–90.
Callomon J.H. The ammonite succession in the Middle Jurassic of East Greenland // Bull. Geol. Soc. Denmark. 1993. V. 40. P. 83–113.
Callomon J.H., Wright J.K. Cardioceratid and kosmoceratid ammonites from the Callovian of Yorkshire // Palaeontology. 1989. V. 32. Pt. 4. P. 799–836.
De Baets K., Bert D., Hofmann R., Monnet C., Yacobucci M.M., Klug Ch. Ammonoid intraspecific variability // Ammonoid Paleobiology: from Anatomy to Ecology. Topics in Geobiology. V. 43. Springer US, 2015. P. 359–426.
Donovan D.T. The Jurassic and Cretaceous stratigraphy and palaeontology of Traill Ø, East Greenland // Meddl. om Grønland. 1953. Bd. 111. Nr. 4. 150 p.
Frebold H. The Jurassic faunas of the Canadian Arctic. Middle and Upper Jurassic ammonites // Bull. Geol. Surv. Canada. 1961. № 74. 43 p.
Frebold H. The Jurassic faunas of the Canadian Arctic. Cadoceratinae // Bull. Geol. Surv. Canada. 1964. № 119. 27 p.
Imlay R.W. Middle Jurassic (Bajocian and Bathonian) ammonites from Northern Alaska // U.S. Geol. Surv. Prof. Pap. 1976. № 854. P. 1–21.
Mitta V., Kostyleva V., Dzyuba O., Glinskikh L., Shurygin B., Seltzer V., Ivanov A., Urman O. Biostratigraphy and sedimentary settings of the Upper Bajocian–Lower Bathonian in the vicinity of Saratov (Central Russia) // Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen. 2014. V. 271. № 1. P. 95–121.
Mitta V., Glinskikh L., Kostyleva V., Dzyuba O., Shurygin B., Nikitenko B. Biostratigraphy and sedimentary settings of the Bajocian–Bathonian beds of the Izhma River basin (European North of Russia) // Neues Jahrbuch für Geologie und Paläontologie-Abhandlungen. 2015. V. 277/3. P. 307–335.
Poulton T.P. Zonation and correlation of Middle Boreal Bathonian to Lower Callovian (Jurassic) ammonites, Salmon Cache Canyon, Porcupine river, Northern Yukon // Bull. Geol. Surv. Canada. 1987. № 358. P. 1–155.
Rawson P.F. New Arctocephalitinae (Ammonoidea) from the Middle Jurassic of Kong Karls Land, Svalbard // Geol. Mag. 1982. V. 119. P. 95–100.
Smith D.G., Harland W.B., Hughes N.F., Pickton C.A.G. The geology of Kong Karls Land, Svalbard // Geol. Mag. 1976. V. 113. № 3. P. 193–232.
Spath L.F. The invertebrate faunas of the Bathonian–Callovian deposits of Jameson Land (East Greenland) // Medd. om Grønl. 1932. Bd. 87. Nr. 7. 158 p.
Zakharov V., Rogov M. Review of the Jurassic System of Russia: stages, boundaries, and perspectives // STRATI 2013. First Int. Congress on Stratigraphy. At the Cutting Edge of Stratigraphy. Springer Geology, 2014. P. 629–634.
Дополнительные материалы
- скачать Supplementary.doc
- Таблица (приложение). Размеры в мм и их соотношения: Д – диаметр раковины, мм; В/Ш – отношение высоты оборота к ширине; У% – относительный диаметр умбилика, в %; пР – количество первичных ребер на полуоборот; РО – реберное отношение
Инструменты
Стратиграфия. Геологическая корреляция