

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 4, стр. 142-158
Биостратиграфия палеогена Сирии по мелким бентосным фораминиферам
Э. М. Бугрова *
Всероссийский научно-исследовательский геологический институт им. А.П. Карпинского
Санкт-Петербург, Россия
* E-mail: Eleonora_Bugrova@vsegei.ru
Поступила в редакцию 10.11.2018
После доработки 07.02.2019
Принята к публикации 28.03.2019
Аннотация
Представлены сведения о таксономическом составе и стратиграфическом распространении мелких бентосных фораминифер в датском–приабонском ярусах палеогена Сирии по результатам изучения коллекций В.А. Крашенинникова. В рамках зональности по планктонным фораминиферам, разработанной этим исследователем и скоррелированной с зональным стандартом Международной стратиграфической шкалы (зоны Р1–Р17), в разрезах Пальмирид по бентосной группе выделены: зона Stensioeina beccariiformis–Anomalina danica (P1а–P2); зона Vaginulina robusta–Pyramidina crassa (P3); зона Reussella paleocenica (P4–P5); зона Pseudogaudryina soldadoensis–Cibicidoides lybicus (P6–P7); зона Hydromylinella wittpyuti (P8–Р9); слои с Paragaudryina dalmatina–Hydromylinella wittpyuti (P10 и P11); зона Falsoplanulina ammophilа (P12 и P13); слои с Queraltina epistominoides (P14); зона Planulina costata (Р15–Р17). По публикациям В.А. Крашенинникова выявлено присутствие видов этих зон в других регионах Сирии и Египта. Проведено сравнение состава и развития фораминифер палеогена Сирии и Крымско-Кавказской области, как представителей биот Тетиса и северо-восточного Перитетиса соответственно.
Посвящается памяти Валерия Аркадьевича Крашенинникова
ВВЕДЕНИЕ
Первые материалы по стратиграфии и тектонике Сирии появились в середине XIX в., но планомерное геологическое изучение этой территории началось лишь в 50-е годы XX в. Среди полученных к тому времени палеонтологических сведений мелкие бентосные фораминиферы (МБФ) не упоминались. В 1958–1963 гг. при геологической съемке территории Сирии в масштабе 1 : 200 000, проводившейся впервые группой советских геологов, начались комплексные стратиграфические работы. “Особая роль в разработке стратиграфии датского и маастрихтского ярусов, палеогена и неогена принадлежит В.А. Крашенинникову, принимавшему непосредственное участие и в полевых исследованиях. Им впервые для территории Сирии создана зональная стратиграфическая шкала палеогеновых отложений и проведено широкомасштабное сопоставление этих исследований со многими районами мира” (Геология…, 1969, с. 6). По развитию планктонных фораминифер (ПФ) в палеогеновых отложениях были выделены 12 зон (Крашенинников, 1964, табл. 2 ; 1965а, 1965б, 1965в и др.). Благодаря их корреляции с близкими по составу зонами Унифицированной схемы палеогена Крымско-Кавказской области, были приведены в соответствие объемы отделов, подотделов и ярусов последней с подразделениями палеогена западноевропейской и океанической шкал (Крашенинников, 1964, 1965в; Крашенинников, Басов, 2007). Позднее (Krasheninnikov et al., 1996; Krasheninnikov, 2005; Крашенинников, Басов, 2007) для региональных зон Сирии была использована нумерация зон Р1–Р17 тропической–субтропической шкалы по ПФ (Berggren et al., 1995), принятой в качестве стандартной для палеогеновой системы Международной стратиграфической шкалы (Luterbacher et al., 2004); при этом положение некоторых зон, обозначенных числами, в пределах ярусов отличалось от стандарта.
Сведения о бентосных фораминиферах палеогена Сирии были отрывочными, за исключением нуммулитид (Абдулрахим, Немков, 1972; Крашенинников, Немков, 1975; Krasheninnikov, 2005). В результате корреляции пелагических фаций с мелководными стал известен состав крупных фораминифер в зонах Р4–Р17 по планктону (Krasheninnikov, Ashourov, 1995). Краткие списки мелких бентосных фораминифер приводились в публикациях В.А. Крашенинникова при характеристике разрезов, и только в отложениях олигоцена им были выделены три региональные зоны.
Хотя таксоны мелких бентосных фораминифер “не являются носителями субглобальной информации” (Крашенинников, Басов, 2007, с. 288), в настоящее время они широко используются в работах по региональной стратиграфии и палеобиогеографии. По замыслу В.А. Крашенинникова, предполагалось наше совместное изучение распределения МБФ в рамках зональности по планктону в разрезах Пальмирид в Центральной Сирии, где представлена наиболее полная последовательность зон по ПФ. В основном с этой территории мне и были переданы отмытые порошки из глинисто-карбонатных пород (коллекция ГИН РАН) с указанием зональной принадлежности по ПФ каждого из них, но без определенного положения в интервале зоны. К сожалению, идея совместной работы осталась неосуществленной, хотя предварительные данные изучения бентоса В.А. Крашенинниковым были просмотрены.
Задачей настоящей работы являлось изучение состава мелких бентосных фораминифер из разрезов Центральной Сирии, расчлененных В.А. Крашенинниковым на зоны по планктонным фораминиферам; выделение биостратиграфических подразделений по МБФ; выявление сходных комплексов в отложениях других регионов Сирии (по публикациям); сравнение состава комплексов и развития фораминифер более тепловодного Тетического бассейна Сирии и умеренно-тепловодного Крымско-Кавказского бассейна (северо-восточного Перитетиса).
МЕТОДИКА
В.А. Крашенинников рекомендовал и сам проводил “расчленение разрезов с помощью комплексов видов, а не отдельных таксонов” (Крашенинников, Басов, 2007, с. 289). Того же принципа придерживается и автор статьи при изучении фораминифер (Бугрова, 2013).
По изученным материалам были выделены “слои” и комплексные зоны – “совокупность слоев, охарактеризованных комплексом древних организмов из трех или более таксонов, отличным от комплексов подстилающих и перекрывающих слоев” (Стратиграфический…, 2006, Ст. VII. 4е). Существенные изменения состава МБФ трактуются как границы в стратиграфической последовательности этих подразделений. По изученным образцам можно говорить о присутствии того или иного вида МБФ в зоне по ПФ, но не всегда можно судить о продолжительности его существования. Очевидно, что первое появление или исчезновение видов в разрезах зависело от частоты взятия образцов и сохранности материала, а не только от фаций. Поэтому совпадение границ зон по ПФ и зон, выделяемых по МБФ, принимается автором условно. Состав зональных комплексов ПФ известен из многих публикаций В.А. Крашенинникова. В представляемой статье сохранено расчленение разрезов на подотделы и зоны по планктону в его понимании (Krasheninnikov et al., 1996, fig. 12 ; Крашенинников, Басов, 2007, рис. 3.2 ), которое несколько отличается от современного варианта, принятого в качестве стандартного (Berggren et al., 1995; Vandenberghe et al., 2012).
Сведения о МБФ палеоцена и эоцена Сирии включают результаты изучения переданных автору образцов и дополнены ревизованными данными из публикаций В.А. Крашенинникова по Сирии и Египту. В его работах отмечалось присутствие видов из палеогена Крымско-Кавказского бассейна; число последних увеличилось после просмотра сирийской коллекции.
С сожалением можно отметить, что у исследователей пока отсутствует единообразное понимание и наименование многих родов и видов МБФ, от чего зависят точность биостратиграфических и палеобиогеографических построений. При определении родовой принадлежности бентоса автор придерживается модифицированной систематики А.В. Фурсенко (Введение…, 1981; Саидова, 1981; Бугрова в Практическое…, 2005, с. 17–38). В отличие от американской системы (Loeblich, Tappan, 1988), в ней преимущественное значение придается плану строения и морфологии раковин, как признакам более филогенетически значимым, чем строение стенки раковины, а также несколько по-иному понимаются иерархия таксонов и диагнозы некоторых родов. В результате был предложен первоначальный вариант деления разрезов Сирии по МБФ (Бугрова, 2016) в интервале датского–приабонского ярусов на региональные комплексные зоны и “слои” с видами-индексами; в настоящей статье в него внесены некоторые изменения. Посредством корреляции с зонами по ПФ зональность по МБФ соотнесена с Международной стратиграфической шкалой (Vandenberghe et al., 2012).
Изображения многих общих видов из Сирии и Крымско-Кавказской области приведены в палеонтологических таблицах (Практическое…, 2005), а также в публикациях автора (Бугрова, 1985, 1989, 2001а, 2004 и др.). Все авторские коллекции фораминифер Северо-Восточного Перитетиса хранятся в ЦНИГР музее им. Ф.Н. Чернышева (г. Санкт-Петербург).
МАТЕРИАЛ
Материалом для данной работы послужили образцы (более 160) из 9 разрезов в основном в центральной части Пальмирид (рис. 1): Айн-Бáрде (Wadi El Bardeh), Джебель Абуад (Jabal El-Abuad), Арак (Arak) или Вади Харрар (Wadi Harrar), Джебель Антар (Jabal Antar), Джебель Коле (Jabal Kohle), Джебель Накнакие (Jabal El-Naknaqiyeh), Хмер (Hmair) и р. Ярмук (The Nahr El-Yarmouk river), Иорданское поднятие. Образцы отобраны с разной степенью детальности из палеоцена–низов верхнего эоцена перечисленных выше разрезов (рис. 2).
Рис. 1.
Схема местонахождений палеогеновых разрезов на территории Сирии (по Krasheninnikov et al., 1996, fig. 2, с сохранением нумерации разрезов). 3 – Мнин (Mnin), 5 – Джебель Маалул (Jabal Maaloula), 10 – Джебель Коле (Jabal Kohle), 11 – Джебель Аада (Jabal Aade), 13 – Хмер (Hmair), 15 – Кариатейн (Al-Qaryatein), 16 – Вади Эль Бáрде или Айн-Бáрде (Wadi El Bardeh), 17 – Вади Эрзек (Wadi Erzeq), 18 – Джебель Хайан (Jabal Hayyan), 19 – Арак (Arak) или Вади Харрар (Wadi Harrar), 24 – Хеит (Heite) или Вади Аль Джизель (Wadi Al Jizel), 25 – Джебель Эль-Абуад (Jabal El-Abuad), 31 – район Джебель Дилаа (Jabal Dilaa), 33 – Джебель Аль-Бишри (Jabal Al-Bishri), 34 – р. Нахр-Эль-Ярмук (The Nahr El-Yarmouk river), 39 – Тагар (Jabal Fru Tagar El-Olab), 44 – Вади Эль-Миа (Wadi El-Miyah), 51 – Абу Дарен (Jabal Abou Darhen), 63 – Максар (Maksar), 67 – Мейданки (Meidanki), 70 – Дамата (Damata), 82 – Джебель Антар (Jabal Antar), 104 – Джебель Накнакие (Jabal El-Naknaqiyeh).
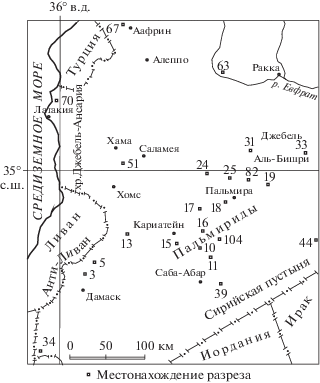
Рис. 2.
Стратиграфические уровни изученных образцов в разрезах палеогена Пальмирид (колл. В.А. Крашенинникова).

В ассоциациях фораминифер обычно преобладает планктон, МБФ составляют до 15% пробы, но при этом они достаточно разнообразны. Как правило, многочисленны секрецирующие формы и практически отсутствуют агглютинирующие, особенно примитивного строения. Литологическая характеристика отложений заимствована из публикаций В.А. Крашенинникова, в которых приведены и мощности зон по планктону (Krasheninnikov et al., 1996, figs. 5, 6 ; Крашенинников, Басов, 2007, рис. 3.3–3.5 ).
БИОСТРАТИГРАФИЯ ПАЛЕОГЕНОВЫХ ОТЛОЖЕНИЙ ПО БЕНТОСНЫМ ФОРАМИНИФЕРАМ
Палеоцен
На контакте мела и палеогена по всей территории Сирии обычно наблюдается несогласие и частичный перерыв.
Датский ярус представлен мягкими светло-серыми и зеленоватыми мергелями мощностью до 30–50 м. По ПФ выделены четыре подразделения (рис. 3).

Зона P1а Parvularugoglobigerina eugubina. Полный разрез от границы с маастрихтом указан лишь в разрезе Айн-Бáрде, где развиты относительно мелководные отложения. Здесь между зонами Abathomphalus mayaroensis (верхний маастрихт) и P1b Parasubbotina pseudobulloides залегает слой известняка (2–3 м), содержащий только бентосные фораминиферы: Clavulina aspera Cushm., Gaudryina retusa Cushm., Ammodiscus incertus d’Orb., Marssonella indentata (Cushm. et Jarv.), Spiroplectammina spectabilis (Grzyb.), Stensioeina caucasica (Subb.), Anomalina acuta (Plumm.), А. danica (Brotz.), Cibicides favorabilis Vassil., Nuttallides truempyi (Nutt.), Karreria fallax Rzehak, Alabamina wilcoxensis Toulmin, Loxostomum applinae Plumm., Gyroidina globosa Hagen., G. octocamerata Cushm. et Hanna, Flabellina rugosa d’Orb., Pullenia coryelli White (Krasheninnikov et al., 1996, в авторском написании таксонов), а также Plectina convergens Keller, Arenobulimina presli (Reuss), Repmanina charoides (Jones et Parker), Trochamminoides irregularis White, Textularia plummerae Lalicker (Крашенинников, Басов, 2007).
В.А. Крашенинников считал данный слой мелководным аналогом зоны P1а P. eugubina по его стратиграфическому положению, поскольку “вышележащие светло-серые мергели содержат микрофауну зоны P. eugubina”: вид-индекс, “Globoconusa daubjergensis, Eoglobigerina eobulloides, Praemurica taurica, Parasubbotina pseudobulloides, Chiloguembelina morsei” (Крашенинников, Басов, 2007, с. 57). Состав МБФ из стратотипического региона зоны P. eugubina (Центральные Апеннины, Италия; Luterbacher, Premoli Silva, 1964) автору неизвестен. В другом глубоководном разрезе (Agost, Испания; Alegret et al., 2003) выше принятого рубежа мела и палеогена преобладают секрецирующие фораминиферы, некоторые переходят этот рубеж; группа эпифауны доминирует или сосуществует с инфауной. В разрезе El Kef Туниса (Speijer, Van der Zwaan, 1996) наблюдаются те же изменения и состав МБФ в низах палеоцена. Часть этих видов встречена и в разрезах Сирии.
На территории северо-восточного Перитетиса выделение зоны P. eugubina обосновано нахождением ее вида-индекса в разрезах Керченского п-ова Крыма (Минтузова, 2008), Мангышлака (Pardo et al., 1999) и Туркменистана (Бугрова, Невзорова в Практическое…, 2005, с. 124–129). Повсеместно же в основании датского яруса фиксируется перерыв, и выше выделена региональная зона Eoglobigerina taurica (соответствующая зоне Р1b), в которой исчезают планктонные фораминиферы зоны Abathomphalus mayaroensis и группы наннопланктона маастрихта, а также значительно изменяется состав бентосных фораминифер (Bugrova, 1996).
Зона P1b Рarasubbotina pseudobulloides (= Turborotalia pseudobulloides; Krasheninnikov et al., 1996) установлена в нескольких изученных разрезах (рис. 2). Бентосная группа из этого интервала содержит Gaudryina pyramidata Cushm., G. gigantica (Subb.), Dorothia retusa (Cushm.), Saracenaria sp., Eponides plummerae Cushm., Osangularia plummerae (Brotz.), Stensioeina beccariiformis (White), Angulogavelinella avnimelechi (Reiss), Anomalina danica (Brotz.), Cibicidoides commatus (Moroz.), С. spiropunctatus (Galloway et Morrey), Brotzenella praeacuta (Vassil.), Stilostomella midwayensis (Cushm. et Todd), Loxostomum kolchidicum Moroz., Loxostomoides applinae (Plumm.) и др.
Зона P1c Praemurica inconstans (= Turborotalia trinidadensis; Krasheninnikov et al., 1996). Раковины МБФ немногочисленны, плохой сохранности, ожелезненные. Определены Bolivinopsis kurtishensis (Balakhm.), *Dorothia retusa Cushm., *Clavulinoides asperus (Cushm.), Nodosaria soluta (Reuss), Pseudonodosaria manifesta Reuss, Dentalina eocenica Cushm., D. colei Cushm. et Dusenb., Robulus cf. degolieri (Plumm.), Vaginulinopsis midwayana (Plumm.), Citharina plumoides (Plumm.), *Stensioeina beccariiformis (White), *Angulogavelinella avnimelechi (Reiss), *Eponides plummerae Cushm., *Anomalina danica (Brotz.), *Brotzenella praeacuta (Vassil.), B. acuta (Plumm.), *Cibicidoides spiropunctatus (Galloway et Morrey), *Loxostomoides applinae (Plumm.), Siphogeneroides elegantus (Plumm.), Stilostomella midwayensis (Cushm. et Todd) и др. (здесь и в следующей зоне Р2 знаком (*) отмечены виды из нижележащих отложений датского яруса).
Зона P2 Praemurica uncinata (= Acarinina uncinata; Krasheninnikov et al., 1996). В изученных разрезах на этом уровне содержатся МБФ Kolchidina sp., Tritaxia midwayensis (Cushm.), *Pseudoglandulina manifesta (Reuss), Robulus aff. midwayensis Plumm., Vaginulinopsis cf. midwayana (Plumm.), *Stensioeina beccariiformis (White), Gyroidinoides subangulatus (Plumm.), *Anomalina danica (Brotz.), Anomalinoides midwayensis (Plumm.), Brotzenella acuta (Plumm.), Karreria sp., Sporobulimina paleocenica Bugr., виды родов Nodosaria, Cibicidoides и др.
В интервале зон P1а–P2 по бентосной группе выделена единая региональная зона Stensioeina beccariiformis–Anomalina danica датского яруса (Бугрова, 2016).
Комплекс сходного состава присутствует в разрезах Египта: Textularia excolata (Cushm.), Dorothia retusa Cushm., Gaudryina austinana Cushm., Stensioeina beccariiformis (White), Anomalina danica (Brotz.), Anomalinoides welleri (Plumm.), Karreria fallax Rzehak, Gavelinella pertusa (Marsson), Alabamina midwayensis Brotz., Gyroidinoides octocameratus (Cushm. et Hanna), Valvulineria brotzeni Nakk. et Tall., Pullenia coryelli White и др. (Крашенинников, Поникаров, 1964; Krasheninnikov, Ponikarov, 1965). МБФ датского яруса принадлежат фауне “мидвейского типа”, расселившейся в начале палеоцена в шельфовых зонах многих бассейнов мира (Berggren, Aubert, 1975; Speijer, Van der Zwaan, 1996 и др.). Находки этой фауны в Сирии и в регионах Перитетиса (Bugrova, 1996; Бугрова, 1999; Beniamovskii, 1998) позволяют расширить ее ареал.
Зеландский ярус обычно сложен серыми тонкослоистыми мягкими мергелями, для которых характерно скопление планктона. В центральной части Пальмирид их мощность составляет 30–50 м. Эти отложения характеризует зона P3 Morozovella angulata s.l., принимаемая в составе зон M. angulata s. str. (P3a) и Igorina pusilla (P3b) (Крашенинников, Басов, 2007). По данным В.А. Крашенинникова (1965в), в отложениях Пальмирид и Анти-Ливана обе подзоны содержат единый таксономический комплекс, но в разрезах последнего МБФ преобладают.
Автором статьи изучены МБФ из зоны M. angulata s.l. Пальмирид (рис. 2). Бентосные формы на этом уровне редки, хотя и представлены более чем 20 родами. Часть видов встречается ниже по разрезу: Bolivinopsis kurtishensis, Dorothia retusa, Gaudryina gigantica, Marssonella indentata, Dentalina eocenica, Flabellina rugosa, Alabamina wilcoxensis, Gyroidinoides globosus, Stensioeina beccariiformis, Angulogavelinella avnimelechi, Anomalina danica, Brotzenella acuta, Karreria fallax, Loxostomoides applinae и некоторые др. Кроме того, появляются Vaginulina robusta Plumm., Robulus turbinatus Plumm., Anomalinoides welleri subsp. laevis (Vassil.), Falsoplanulina ekblomi (Brotz.), Pyramidina crassa Brotz., Ellipsonodosaria paleocenica Cushm., Allomorphina halli Jannings.
Виды этого комплекса распространены и за пределами Пальмирид. В разрезах зоны Р3 Анти-Ливана также отмечено присутствие большинства из них (Крашенинников, 1965в), включая Vaginulina robusta и Pyramidina crassa, а последний указан и в разрезах Месопотамии.
Отложения с приведенной выше фауной выделены в региональную зону Vaginulina robusta–Pyramidina crassa (Бугрова, 2016). Часть ее видов – Gaudryina pyramidata, Flabellina rugosa, Gyroidina globosa, Nuttallides truempyi, Anomalina danica, Loxostomum applinae, Pyramidina crassa – распространена на этом же уровне в разрезах Египта (Krasheninnikov, Ponikarov, 1965). По составу фауна близка к зоне Pyramidina crassa зеландского яруса Перитетиса (Бугрова в Практическое…, 2005, с. 46–78; Зональная…, 2006).
Состав фораминифер определенно свидетельствует о связях бассейнов Средиземноморья и Перитетиса (Атлас…, 1967). Судя по нахождению общих видов МБФ в разрезах Афганистана, Пакистана и некоторых крупных фораминифер (Lockhartia, Kathina, Rotalia, Nummulites) в Туркмении и Таджикистане (Бугрова, 2009, 2014; Бугрова, Старшинин, 2013), юго-восточные окраины Перитетиса были связаны системой проливов с тетическими морями.
Танетский ярус слагают серые и темно-серые мягкие мергели, мощность которых в центральной части Пальмирид составляет 80–150 м. По обильным ПФ выделены зоны P4 Planorotalites pseudomenardii и P5 Morozovella velascoensis (Krasheninnikov et al., 1996; Крашенинников, Басов, 2007). Таксономическое разнообразие МБФ при их малом количестве (0.35–5.0% комплекса) достаточно высоко: более 25 родов в зоне Р4 и около 40, в основном секрецирующих, в зоне Р5, но среди них трудно выбрать виды, характерные лишь для данного уровня. Среди агглютинирующих форм представлены эволюционно продвинутые роды. В.А. Крашенинников (1965в, с. 40) отмечает, что “примитивные песчанистые фораминиферы (ризамминиды, саккамминиды, гиперамминиды, реофациды) редки или отсутствуют”. В образцах встречаются гладкостенные остракоды, чешуя и зубы рыб.
В отложениях зоны P4, наряду с встречающимися ниже Marssonella indentata, Robulus turbinatus, Vaginulina robusta, Stensioeina beccariiformis, Angulogavelinella avnimelechi, Osangularia plummerae (Brotz.), Anomalinoides welleri subsp. laevis, Brotzenella ex gr. acuta, Cibicidoides spiropunctatus (Galloway et Morrey), появляются и новые виды МБФ: Textularia farafraensis LeRoy, Bolivinopsis kurtishensis aff. limbosa (Balakhm.), Vulvulina gracilima ten Dam et Sigal, Clavulinoides aff. midwayensis (Cushm.), Dorothia bulletta (Carsey), Lenticula aff. globulosa (ten Dam), Nodosaria velascoensis Cushm., Vaginulinoides aff. midwayana (Plumm.), Alabamina cf. wilcoxensis Toulmin, Gyroidinoides octocameratus (Cushm. et Hanna), Valvulineria esnahensis Nakkady, Anomalina ex gr. danica (Brotz.), Pleurostomella paleocenica Cushm., Reussella paleocenica (Brotz.), Bulimina midwayensis Plumm., В. farafraensis LeRoy, B. esnaensis LeRoy, Tappanina selmensis Cushm., Allomorphina allomorphinoides (Reuss), не встреченные ранее полиморфиниды, а также неопределенные виды родов Textularia, Gaudryina, Lingulonodosaria, Robulus, Frondicularia, Pullenia, Aragonia.
В отложениях зоны P5 распространены Spiroplectinella aff. tenera (J. Nikitina), Spiroplectammina richardi Martin, Bolivinopsis spectabilis (Grzyb.), Nodosaria aff. latejugata Guemb., Robulus insulsus Cushm., Neoflabellina delicatissima (Plumm.), Frondicularia frankei Cushm., Marginulinopsis tuberculata (Plumm.), Gyroidina subangulata (Plumm.), Alabamina midwayensis Brotz., Valvalabamina aff. depressaeformis (N. Byk.), Pullenia quinqueloba Reuss, Anomalina ex gr. danica (Brotz.), Cibicidoides sp., C. aff. howelli (Toulmin), Viveija cf. excavata (Brotz.), Siphogeneroides elegantus (Plumm.), Stilostomella midwayensis (Cushm. et Todd), Reussella paleocenica (Brotzen), Bulimina сf. midwayensis Cushm., B. quadrata (Plumm.), B. cacumenata Cushm. et Park., Tappanina selmensis Cushm., Bolivinoides delicatulus Cushm., Aragonia semireticulata (LeRoy), Globimorphina conica (Cushm. et Todd), а также редкие формы родов Clavulina, Gaudryina, Vulvulina, Pseudonodosaria, Lingulonodosaria, Saracenaria, Astacolus, Robulus, Bolivina.
Вблизи кровли разреза Айн-Бáрде встречены Arenobulimina sp., Gaudryina aff. pyramidata Cushm., Stensioeina cf. beccariiformis (White), Buliminella sp., Sporobulimina paleocenica Bugrova, Chilostomella eocenica (Cushm.). Из разрезов депрессии Латакия–Аафрин известны (Крашенинников, 1965в) Reussella paleocenica (Brotz.), Clavulina aspera White et Jones, Nuttallides trumpyi (Nutt.), Pullenia coryelli White, Globocassidulina globosa (Hantk.), Allomorphina halli (Jennings) и др.
По МБФ в объеме зон P4 и P5 выделена одна зона Reussella paleocenica. В мелководных известняках хребта Ансария (Эн-Нусайрия; Jabal Nusseiriyen) на этом уровне встречаются Nummulites, Operculina, Discocyclina и толстостенные скульптированные раковины роталиид, эльфидиид, дискорбид, аномалинид (Абдулрахим, Немков, 1972; Крашенинников, Немков, 1975; Krasheninnikov, Ashourov, 1995; Krasheninnikov, 2005).
Разнофациальные комплексы МБФ Перитетиса значительно отличаются по составу от одновозрастных сирийских. В глубоководных разрезах бассейна (Северный Кавказ и Западный Копетдаг) распространены в основном агглютинирующие формы; в его мелководных (платформенных) разрезах присутствуют R. рaleocenica (Brotz.) и другие виды, общие с этой зоной Сирии (Бугрова, 2004; Бугрова в Практическое…, 2005, с. 46–78).
Эоцен
Нижний эоцен. Отложения ипрского яруса залегают на верхнепалеоценовых согласно, а перерывы в осадконакоплении носят местный характер. Разрезы сложены мергелями и глинистыми известняками с обилием раковин ПФ, в основании некоторых разрезов наблюдается пласт, обогащенный глауконитом (Krasheninnikov et al., 1996). Мощность отложений в центральной части Пальмирид колеблется от 130 до 250 м, но иногда она сокращается до 40–80 м. В нижнем эоцене выделены 4 зоны по планктону (Krasheninnikov et al., 1996; Крашенинников, Басов, 2007).
Бентос из отложений почти всех зон эоцена изучен в разрезах Айн-Барде, Джебель Эль-Абуад и Джебель Антар, где были обнаружены зерна глауконита, обломки костей и чешуя рыб, фосфоритизированные копролиты, гладкие и скульптированные раковины остракод.
В Сирийском бассейне изменение состава МБФ на рубеже палеоцена и эоцена происходило в основном на видовом уровне. В Перитетисе на этом рубеже изменения коснулись состава родов и семейств, а процессы вымирания фораминифер и их восстановление протекали в регионах по-разному в зависимости от фациальной обстановки (Bugrova, 1996; Бугрова, 2000а).
Далее в списках МБФ зон Р6–Р9 Сирии отмечены знаком (+) формы, общие с фауной Перитетиса.
В зоне P6 Morozovella subbotinae бентос достаточно редок (в разрезе Джебель Антар 1.2–15.5%), хотя и разнообразен по составу (около 45 родов). Большинство видов появляется впервые, среди них много эндемичных. Комплекс включает Spiroplectinella desertorum (LeRoy), S. knobele (LeRoy), +Gaudryina spinulosa Bugrova, Pseudogaudryina soldadoensis (Cushm. et Renz), +P. externa Bugrova (= Gaudryina navarroana Cushm. sensu V. Krasheninnikov), +P. aff. concavilata Bugrova, Pseudoclavulina maqfiensis LeRoy, P. farafraensis LeRoy, Vulvulina sp., +Spirosigmoilina aff. decorata (Cushm.), Lagena spp., Lingulonodosaria sp. nov., Ellipsonodosaria granti (Plumm.), +Pseudoglandulina manifesta Reuss, Frondicularia sp., +Marginulinopsis eofragaria (Balakhm.) (=? Marginulina carri LeRoy), +Brotzenella ex gr. acuta (Plumm.), Anomalinoides desertorum (LeRoy), +Cibicidoides cf. lybicus (LeRoy), С. farafraensis (LeRoy), +C. aff. beatus (Martin), +Valvalabamina depressaeformis (N. Byk.) subsp. nov., +Cancris sp., Osangularia sp., +Falsoplanulina subbotinae (Balakhm.) [ex gr. Falsoplanulina ammophila (Guembel)], Bulimina esnaensis LeRoy, Eouvigerina aegyptiaca Nakkady, +Angulogerina wilcoxensis Cushm. et Appl., +Aragonia aragonensis (Nutt.), +Tappanina selmensis Cushm., Loxostomoides sp., +Allomorphina allomorphinoides (Reuss) и еще некоторые неопределимые формы. Из скважин на востоке Сирийской пустыни В.А. Крашенинниковым указан вид +Pseudogaudryina externa.
В зоне P7 Morozovella formosa бентосные виды по-прежнему редки (не более 10%); однако они принадлежат 35–40 родам, часть из которых представлена несколькими видами. Некоторые виды встречаются и в предыдущей зоне (Pseudogaudryina soldadoensis, P. externa, P. aff. concavilata, Pseudoclavulina maqfiensis, Spirosigmoilina aff. decorata, Marginulinopsis eofragaria, Valvalabamina depressaeformis subsp. nov., Cancris sp., Brotzenella ex gr. acuta, Falsoplanulina subbotinae, Eouvigerina aegyptiaca, Bulimina esnaensis, Tappanina selmensis). К ним добавляются +Martinottiella eocenica Cushm. et Berm., +Siphotextularia suleimanovi Bugrova, +Procerolagena axiformis (Matthes), +Orthomorphina aff. havanensis (Cushm. et Berm.) и представители других родов отряда Nodosariida, Valvulineria aegyptiana LeRoy, +Globorotalites sp. nov., +Siphonina wilcoxensis Cushm., +Cibicidoides lybicus (LeRoy), ?Karreria sp., +Globocassidulina aff. globosa (Hantk.), Loxostomoides sp., +Pleurostomella alternans Schwager, +Turillina brevispira ten Dam, Bulimina aff. aksuatica Moroz., Aragonia sp. nov. Среди вновь появившихся видов выделяется морфологически характерный Cibicidoides sp. 1 (с широким спиральным швом), находимый и выше по разрезу.
На уровне зон P6 и P7 выделяется региональная зона Pseudogaudryina soldadoensis–Cibicidoides lybicus по МБФ. По общности с фауной Перитетиса (+) подразделение соответствует зоне Pseudogaudryina externa низов эоцена СНГ (Бугрова, 1988б, 2004; Практическое…, 2005; Зональная…, 2006).
Широкое распространение теплолюбивого рода ПФ Morozovella и нуммулитид, разнообразие планктона и секрецирующих МБФ свидетельствуют о проявлении температурного максимума начала эоцена (PETM) и в бассейнах северо-восточного Перитетиса. Судя по нахождению многих общих видов, в раннем ипре бассейны Сирии, Египта и Перитетиса были тесно связаны (Атлас…, 1967), причем не только через западные акватории, но и, вероятно, через проливы в Средней (Центральной) Азии (Бугрова, 2014).
Зона Р8 Morozovella aragonensis верхнего ипра сложена известняками, глинистыми известняками, мергелями светлых тонов. Их мощность в центральной части Пальмирид колеблется от 100 до 180 м.
В комплексе фораминифер по-прежнему преобладает планктон (до 100% в разрезе Вади Хмер (= Вади Хабра (Wadi Khabra), № 13 на рис. 1), хотя содержание МБФ бывает более значительным (до 40% в разрезе Джебель Антар) и состав их разнообразнее (до 50 родов). Сохранность фауны недостаточно хорошая, из-за чего часть ее не была определена до вида.
В зоне Р8 изменяется родовой и видовой состав бентоса. Впервые появляются два рода спирально-плоскостных нодозариид – Hydromylinella Bugrova, 2001 (pro Hydromylina de Witt Pyut nom. nud.) и Turkmenicaella Bugrova, 1985 (pro Turkmeniella Bugrova nom. preoccup.) (Бугрова, 1985, 2001а), а также вид Robulus curvicameratus Bugrova, характерный в основном для отложений Средней (Центральной) Азии.
Комплексы включают Pseudogaudryina pseudonavarroana (Balakhm.), +P. aff. concavilata Bugrova, Clavulina sp., Dorothia aff. bulletta (Carsey), Marginulina pachigaster Guemb., +Robulus curvicameratus Bugrova, R. aff. limbosus (Hantk.), R. aff. princeps (Reuss), +Hydromylinella wittpyuti Bugrova forma typica [= Cristellaria iljini N. Byk. nom. nud. в публикациях В.А. Крашенинникова], Hydromylinella sp. nov., Turkmenicaella sp. nov., Astacolus praemexicanus (N. Byk.), Fursenkoinella aff. gorynica (A. et K. Furs.), Cibicidoides sp. 1, +C. aff. gigas (Hagn), C. aff. carinatus (Terquem), +Anomalina aff. capitata (Guemb.), +Brotzenella ex gr. acuta (Plumm.), +Falsoplanulina balakhmatovae Bugrova [сходная с “Cibicides” multifaria (Schwager) var. mellahensis Nakkady, 1950 из свиты Esna Египта], а также Falsoplanulina sp. nov. [из группы F. ammophila (Guembel)], Planulina sp., Alabamina aff. discana (Chalil.), +Baggina valvulineriaformis (N. Byk.), Cancris sp., Globobulimina ovata (Orb.), +Bulimina cf. mitgarziana Balakhm., +B. pseudoinflata Balakhm., Hopkinsina aff. compacta Balakhm., +Uvigerina bykovae (Balakhm.), +Loxostomoides adelae (Liebus), Aragonia aragonensis (Nuttall), виды родов Vulvulina, Nodosaria, Dentalina, Citharina, Marginulinopsis, Valvulineria, Turrilina, Fursenkoina, Pleurostomella, Globocassidulina и др. На западном склоне хр. Ансария (Эн-Нусайрия; Jabal Nusseiriyen) В.А. Крашенинниковым отмечен вид +Pseudogaudryina pseudonavarroana.
Кроме видов МБФ, общих с Перитетисом (отмеченных знаком +), в этой зоне Сирии, как и в Перитетисе, появляются обильные радиолярии, главным образом сферические и дискоидальные. В разрезе Вади Харар обнаружен род Voorthuyseniella Szczechura, 1969 (Incertae Sedis), стратиграфическое и географическое распространение которого не изучено; найденная “проблематика” морфологически несколько отличается от описанного из отложений Северного Кавказа вида V. tunica Bugrova (Бугрова, 2003). В образцах встречаются скульптированные раковины остракод и зубы рыб.
Следующая зона Р9 Acarinina pentacamerata заканчивает разрез ипрского яруса Сирии (Krasheninnikov et al., 1996). Она сложена чередованием светлых известняков и мергелей, мелоподобных или крепких известняков и многочисленных пластов черных и бурых кремней. Мощность толщи от 100 до 250 м.
Образцы на определение микрофауны отбирались из мягких прослоев, но сохранность материала оказалась не всегда хорошей. В комплексах фораминифер изученных разрезов увеличивается количество раковин бентоса, при этом сокращается число их родов (до 25). Некоторые формы известны из зоны Р8 по ПФ. Это Pseudogaudryina pseudonavarroana, P. aff. concavilata, Marginulina pachigaster, Astacolus praemexicanus, Robulus curvicameratus, Hydromylinella wittpyuti forma typica, Hydromylinella sp. nov., Cibicidoides sp. 1, C. aff. carinatus, C. aff. gigas, Baggina valvulineriaformis, Falsoplanulina sp. nov., Bulimina mitgarziana, Hopkinsina aff. compacta, Loxostomoides adelae.
Впервые появляются новые виды и роды Paragaudryina и Amphimorphina (оба редко в верхней части зоны). В этом интервале встречаются +Spiroplectammina carinatiformis Moroz., +Gaudryina aff. subbotinae Welmin, +Paragaudryina dalmatina (Liebus), Clavulina sp., Clavulinoides aff. golubjatnikovi (Schutzk.), Nodosaria sp., Amphimorphina sp., Robulus spp., +Heterolepa sp. (= Cibicides limarius N. Byk. nom. msc.), +Cancris turkmenensis (N. Byk.), Baggina bardehensis Bugrova nom. msc., B. aff. globula Balakhm., Valvulineria sp. (с натечными швами), +Falsoplanulina balakhmatovae Bugrova, +B. aksuatica Moroz., Hopkinsina sp. (мелкие раковины), Bolivina spp. (мелкие без скульптуры).
В зоне Р9 (разрез Арак) обнаружен планктонный вид Truncorotaloides arcanus Bugrova, который маркирует верхнюю границу ипрского яруса в Западном Туркменистане (Бугрова, 2009) и на Мангышлаке. Данный вид, возможно, является синонимом T. piparaensis Bronn. et Bermudez, известным в разрезах Израиля на том же уровне (Bendjamini, 1980).
Помимо фораминифер, в образцах присутствуют радиолярии, остракоды и зубы рыб.
Некоторые бентосные виды широко распространены в разрезах Сирии на уровне зон Р8 и Р9. Так, из публикаций В.А. Крашенинникова известны Hydromylinella wittpyuti, Bulimina mitgarziana, Cancris turkmenensis, Pseudogaudryina pseudonavarroana, Clavulinoides aff. golubjatnikovi в разрезах Пальмирид, массива Анти-Ливана и депрессии Латакия–Аафрин; Hydromylinella wittpyuti на правобережье р. Ярмук и на юго-западе в разрезе Мнин; вид P. pseudonavarroana на западном склоне хр. Ансария (Эн-Нусайрия; Jabal Nusseiriyen); на Алеппском плато найдены H. wittpyuti, B. mitgarziana, а комплекс видов P. pseudonavarroana, H. wittpyuti, B. mitgarziana, C. turkmenensis, B. valvulineriaformis – в разрезе Джебель Аль-Бишри; на левобережье р. Евфрат в Месопотамии (разрез Максар) отмечены H. wittpyuti, B. mitgarziana и Aragonia aragonensis. К мелководным отложениям приурочены крупные фораминиферы.
Такие характерные виды, как Hydromylinella wittpyuti, Bulimina mitgarziana, Clavulinoides aff. golubjatnikovi, встречаются в зонах M. aragonensis и A. pentacamerata разрезов Египта (Крашенинников, Поникаров, 1964).
В интервале зон Р8–Р9 выделяется зона Hydromylinella wittpyuti по МБФ (рис. 3).
По нахождению общих видов, отмеченных знаком (+), данная зона сопоставляется с верхнеипрской зоной Bulimina mitgarziana разрезов Перитетиса (Зональная…, 2006; Бугрова, 2014). В это время по-прежнему существовали широкие связи Перитетиса с бассейнами Средиземноморья (Атлас…, 1967).
Средний эоцен. Отложения повсеместно залегают согласно на кремнистой толще нижнего эоцена. Разрезы сложены белыми и светло-серыми мелоподобными известняками с обильным планктоном, которые в нижней части включают пачки более крепких слоистых известняков и глинистых известняков, а ближе к верхам – прослои окремненных известняков.
По ПФ в основании толщи В.А. Крашенинниковым выделены зоны P10 Hantkenina aragonensis и P11 Globigerapsis kugleri. Но поскольку “разграничение зон вызывает определенные трудности”, выделяется “нерасчлененный интервал… под условным названием зоны Acarinina bullbrooki” (Крашенинников, Басов, 2007, с. 62), относимой к лютетскому ярусу (Krasheninnikov et al., 1996).
Зона Acarinina bullbrooki. Максимальные мощности данной зоны приурочены к центральной части Пальмирид (150–200 м). В образцах из разрезов Айн-Бáрде и Арак присутствуют глауконит, желваки фосфоритов, галечки кремней и зубы рыб.
Из-за небольшого числа образцов и плохой сохранности фауны бентос этой части разреза охарактеризован недостаточно; возможно, в список со знаками открытой номенклатуры включены эндемичные формы. При визуальных различиях комплексов трудно передать их таксономические отличия.
Малочисленные МБФ принадлежат 22 родам. Среди них обнаружены Spiroplectammina aff. carinatiformis Moroz., Pseudogaudryina cf. pseudonavarroana (Balakhm.), Paragaudryina dalmatina (Liebus), Clavulinoides sp., Robulus spp. (крупные с выпуклыми швами), Marginulinopsis sp., Hydromylinella wittpyuti Bugrova forma typica и H. wittpyuti forma tumida (?= Robulus trompi Ansary), Eponides aff. jacksonensis Cushm., Valvulineria sp. (девятикамерная), Alabamina sp., Cibicidina aff. westi (Howe), Cibicides? aff. multifaria (Schwager), Uvigerina sp. (ребристая), Bulimina aff. sculptilis Cushm., Loxostomoides adelae (Liebus), Bolivina sp. и др. Распространены многочисленные остракоды.
В.А. Крашенинников указывал нахождение Paragaudryina dalmatina, Cibicidina westi, Nuttallides florealis на Алеппском плато; Paragaudryina dalmatina, Hydromylinella wittpyuti в разрезах Ант-и-Ливана.
В интервале зон Р10–Р11 определены в основном транзитные виды МБФ, по которым выделены слои с Paragaudryina dalmatina и Hydromylinella wittpyuti (рис. 3).
Обращает на себя внимание не только обедненность состава МБФ, но и редкость общих видов с фауной Перитетиса: это Paragaudryina dalmatina, Hydromylinella wittpyuti, Loxostomoides adelae, описанные из Средиземноморья (Liebus, 1911) и находимые в Крымско-Кавказской области. В зоне Р10 Сирии было отмечено (Krasheninnikov et al., 1996) присутствие родов Hantkenina (многочисленные H. aragonensis), Clavigerinella, Gumbelitrioides, Morozovelloides (= Morozovella spinulosa). В изученном образце из разреза Вади Харрар обнаружены Morozovelloides crassatus (Cushm.) и Hantkenina liebusi Shokhina. Эти роды и виды появляются в Перитетисе в низах среднего эоцена (Бугрова в Практическое…, 2005, с. 163–174; Зональная…, 2006) вследствие их иммиграции из Средиземноморья.
Вышележащие зоны P12 Morozovella lehneri и P13 Orbulinoides beckmanni В.А. Крашенинников относил к лютету и бартону соответственно (Krasheninnikov et al., 1996), что отличается от современного деления палеогена МСШ (Vandenberghe et al., 2012, fig. 28.1 ). Планктонные фораминиферы здесь обильны и разнообразны, ряд вновь появившихся видов распространен в обеих зонах. Как отмечал В.А. Крашенинников, вид-индекс верхней зоны встречается в ограниченном количестве экземпляров. В изученном материале Orbulinoides beckmanni обнаружен в разрезах Айн-Бáрде и Вади Харрар (?) (по одному экземпляру плохой сохранности). Ассоциации МБФ в отложениях этих зон сходны по видовому составу и далее рассматриваются совместно.
В основании разрезов зон P12 и Р13 содержится глауконит, желваки фосфоритов, встречаются зубы рыб, створки остракод. Максимальные мощности отложений в центральной части Пальмирид составляют 110–170 м (зона P12) и 80–160 м (зона P13).
МБФ малочисленны, недостаточно хорошей сохранности, иногда ожелезненные. Комплекс представлен 26 родами, в нем появляются некоторые новые виды, особенно среди нодозариид. В образцах обнаружены Paragaudryina dalmatina (Liebus) внизу, Clavulinoides triradiatus Minakova [?= C. szaboi (Hantken) sensu Krasheninnikov], Chrysalogonium leave Cushm. et Berm., крупные Robulus spp., R. laticostatus (Tutkow.), Hydromylinella sp., Turkmenicaella granata (Bugrova) (единственный экземпляр в зоне Р13 разреза Вади Харрар, T. sp. nov., T. sp. ind., Marginulinopsis fragaria (Guemb.), Brotzenеlla aff. turkmenica Bugrova, Heterolepa eocaena (Guemb.), Falsoplanulina ammophila (Guemb.), F. aff. tympaniformis Bugrova et Balakhm. [?= F. cubensis (Cushm. et Berm.)], Cibicidina westi (Howe), Globocassidulina subglobosa (Brady), Bulimina sculptilis Cushm., Uvigerina costellata Moroz., U. bykovae (Balakhm.), неопределенные виды родов Nodosaria, Dentalina, Marginulina, Baggina и др.
Из разрезов Пальмирид, Анти-Ливана, Алеппского плато и Сирийской пустыни В.А. Крашенинниковым указаны Paragaudryina dalmatina, Clavulinoides szaboi, Cylindroсlavulina cyclostomata, Marginulinopsis fragaria, Heterolepa eocaena [как Cibicides perlucidus Nutt.], Cibicidina westi, Bulimina sculptilis, Uvigerina bykovae.
На имеющемся материале не выявлено существенных отличий бентосных ассоциаций, приуроченных к каждой из зон по ПФ, поэтому по МБФ выделяется единая зона Falsoplanulina ammophilа. При наличии некоторых общих видов, различия бентосных ассоциаций Сирии и Перитетиса достаточно значительны.
Планктонный комплекс в Перитетисе беднее сирийского. В региональной зоне Acarinina rotundimarginata этого бассейна появляется указанный вид-индекс, Globigerinatheka subconglobata, Subbotina frontosa, более редкие Hantkenina liebusi, H. dumblei, Morozovelloides crassatus, M. spinulosa и M. crater (последний только на Северном Кавказе в опорном разрезе р. Хеу). В следующей зоне Hantkenina alabamensis появляется Truncorotaloides topilensis; вид-индекс зоны Р13 Orbulinoides beckmanni повсеместно отсутствует. Нахождение перечисленных родов ПФ в Перитетисе, как и в Сирийском бассейне, отмечает событие МЕСО (Mid Eocene climatic optimum).
Присутствие общих планктонных и в меньшей степени бентосных форм позволяет говорить о достаточно свободной связи Северокавказского бассейна с тетическими морями (Атлас…, 1967), о его нормальной солености, а развитие бентоса с известковой раковиной свидетельствует об аэрации придонных слоев.
Зона P14 Truncorotaloides rohri (бартон) на территории Пальмирид сложена белыми и светло-серыми мелоподобными известняками, которые по площади переходят в переслаивание мергелей и известняков, в том числе органогенных. В основании разрезов выделяется маркирующий глауконит-фосфоритовый пласт. Мощность отложений в центральной части Пальмирид достигает 102 м (разрез Джебель Эль-Абуад), сокращаясь на окраинах.
Сведения о МБФ этой части разреза собраны из публикаций В.А. Крашенинникова и лишь несколько дополнены данными изучения автором двух образцов из разрезов Айн-Бáрде и Арак.
Состав бентоса резко изменяется по площади в зависимости от фаций. Пелагические образования с преобладанием ПФ, развитые в центральной части Пальмирид, к бортам впадины замещаются мелководными органогенно-обломочными и нуммулитовыми известняками. В плотных известняковых породах (восточнее г. Саба-Абар) содержатся булиминиды, лагениды, аномалиниды, роталииды и дискорбиды, видовой состав которых не приводится. Сопутствующий им обедненный комплекс ПФ включает Truncorotaloides rohri (Bronn. et Berm.) и Subbotina turcmenica (Chalilov). Совместно с этим планктоном на правобережье Евфрата (скважина Суаб в приграничном с Ираком районе) отмечен бентосный вид Queraltina epistominoides Marie. По данным В.А. Крашенинникова, много МБФ содержится в пластах мягких известняков на восточном склоне Анти-Ливана. Отсюда известны Queraltina epistominoides Marie, Heterolepa eocaena (Guemb.), Robulus roemeri (Reuss), Baggina iphigenia (Samoil.), Bulimina sculptilis Cushm., Uvigerina jacksonensis Cushm. и др.; кроме них, Asterigerina varians Chal., A. contusa Chal., Discorbis schurutensis Chal., D. affinis Chal., Nonion sagittum Chal., Bolivina adziderensis Chal., Elphidium consociatum Chal., распространенные в зоне Subbotina turcmenica верхов среднего эоцена Азербайджана (Халилов, 1962).
Раковины МБФ, обнаруженные в образцах из разрезов Айн-Бáрде и Арак, имеют плохую сохранность. Они определены как Spiroplectammina sp., Nodosaria aff. elegantissima (dʾOrb.), Robulus chitanii Yabe et Asano, Marginulinopsis aff. fragaria (Guemb.) [?= Lenticulina? cf. waiparaensis (Finlay) (sensu Ansary)], Hydromylinella aff. trompi (Ansary), Frondicularia sp., Baggina bradyi (Brotz.) (sensu Ansary), Cancris cf. amplus Finlay, Globocassidulina sp., Hopkinsina? bartotara Finlay “var.” aegyptiaca Ansary, мелкие Bolivina spp. Эта фауна близка к описанной из эоценовых отложений Египта (Ansary, 1955), где также установлено совместное нахождение Subbotina turcmenica и Queraltina epistominoides (Крашенинников, Поникаров, 1964). Поскольку распространение бентосного вида Q. еpistominoides в разнофациальных условиях не имеет четких стратиграфических границ, на уровне зоны Р14 выделены слои с Queraltina epistominoides (Бугрова, 2016). Вид-индекс этих слоев в Западной Европе встречается на южном склоне плато Добруджа также совместно с обильными T. rohri (Крашенинников, 1965б).
Развитие северо-восточного Перитетиса в это время происходило в обстановке кислородной недостаточности (Бугрова, 2000б; Beniamovskii et al., 1999 и др.), вследствие чего резко изменился состав планктона и бентоса (биособытие вымирания). Появился комплекс зоны Subbotina turcmenica с двумя подзонами (Зональная…, 2006), включающий S. turcmenica (Chal.), S. azerbaidjanica (Chal.), Pseudohastigerina micra (Cole), Globigerina spp. мелких размеров, крайне редко Truncorotaloides rohri (Brönn. et Berm.) и в верхней подзоне G. praebulloides Blow, G. instabilis Korov., Acarinina rugosoaculeata Subb. (= A. medizzai Toumarkine et Bolli). На Северном Кавказе (р. Савердон; Бугрова, Письменная, 2009) в основании зоны обнаружены Hantkenina alabamensis Cushm., единичные H. australis Finlay, H. dumblei Weinz. et Applin, Truncorotaloides topilensis (Сushm.), Subbotina pseudocorpulenta Chal. Здесь в породе наблюдаются крупные фрагменты водорослей, определенные М.А. Ахметьевым (устное сообщение) как Azolla sp., которая существовала в условиях опреснения поверхностных вод. Об аноксической обстановке с признаками опреснения свидетельствует и присутствие празинофитов и акритарх (Попов и др., 2018). Более дробное деление этой части разреза по ПФ (Беньямовский, 2001, 2011) отражает особенности биоты глубоководного Северокавказского бассейна, и ареалы выделенных В.Н. Беньямовским биостратонов не охватывают северные и восточные регионы платформенного мелководья.
Среди МБФ в Перитетисе стали преобладать агглютинирующие виды. Так, в среднеазиатской части бассейна расселились эндемичные виды с песчанистой раковиной родов Proteonella, Saccammina, Hormosina, Evolutinella, Subtilina, Popovia, Trochammina, Spiroplectammina, Bolivinopsis, Schenсkiella, Gaudryinopsis; постоянно присутствовали радиолярии, причем иногда они составляли всю ассоциацию микрофауны (Бугрова, 2014). Появилось несколько новых секрецирующих видов – Turkmenicaella singularis Bugrova, Planulina tecta Rodionova, Siphonina praelata N. Byk., Uvigerina citae (Hagn), в том числе в Крыму (опорная скважина № 2, Бахчисарайский район; Бугрова, 1988а). В конце этого времени практически по всей территории Перитетиса распространился комплекс с видами родов Caucasina, Caucasinella и Bolivina (Практическое…, 2005; Бугрова, 2012), не встреченный в разрезах Сирии.
Верхний эоцен. Приабонский ярус на территории Пальмирид сложен известняками разных фациальных типов – от пелагических до мелководных органогенных (нуммулитовых, водорослевых и рифовых); в низах некоторых разрезов отмечены мягкие глинистые известняки. Их залегание на среднеэоценовых породах согласное, иногда с глауконитовым прослоем в основании. Мощность отложений колеблется в пределах 40–110 м.
ПФ численно преобладают. По этой группе В.А. Крашенинниковым выделены зоны Р15–Р17: Globigerapsis semiinvoluta, Turborotalia cocoaensis и Turborotalia centralis–Globigerina gortanii, которые не всегда можно разделить.
МБФ верхнего эоцена Сирии достаточно разнообразны по составу (табл. 1). По публикациям (Крашенинников, 1964, 1965в; Krasheninnikov et al., 1996) можно судить о распространении МБФ на территории Северных Пальмирид, восточного склона Анти-Ливана, Алеппского плато, Месопотамии и других местонахождений. Здесь встречаются виды Clavulinoides szaboi (Hantken), Karreriella siphonella (Reuss), Siphonodosaria annulifera (Cushm. et Berm.), Frondicularia budensis Hantk., Robulus limbosus (Reuss), Saracenaria arcuata d’Orb., Vaginulinopsis cumulicostatа (Guemb.), Marginulina boehmi (Reuss), Marginulinopsis fragaria (Guemb.), Anomalina granosa (Hantk.), Anomalinoides affinis (Hantk.), Heterolepa pygmea (Hantk.), Planulina costata (Hantk.), Bulimina sculptilis Cushm., Uvigerina jacksonensis Cushm., Bolivinoides reticulatus (Hantken), Bolivina antegressa Subb. и др.
Таблица 1.
Таксономический состав и распространение бентосных фораминифер зоны Planulina costata в разрезах Сирии
| Названия видов | Центральные Пальмириды | По В.А. Крашенинникову | ||||||
|---|---|---|---|---|---|---|---|---|
| Северные Пальмириды | Анти-Ливан | Алеппское плато | Плато Халаб | Латакия–Аафрин | Джебель Аль-Бишри | Месопотамия | ||
| Spiroplectammina azovensis J. Nikit. | + | + | ||||||
| Clavulinoides szaboi (Hantk.) | + | + | + | + | + | + | ||
| Clavulina cyclostomata (Gall. et Morr.) | + | + | + | |||||
| Karreriella siphonella (Reuss) | + | + | + | + | ||||
| Nodosaria bacillum Defr. | + | + | + | |||||
| Laevidentalina communis (d’Orb.) | + | |||||||
| Dentalina consobrina d’Orb. | + | + | ||||||
| Siphonodosaria exilis (Neugeb.) | + | + | + | |||||
| S. annulifera (Cushm. et Berm.) | + | + | + | |||||
| S. volgensis Samoil. | + | + | ||||||
| Robulus limbosus (Reuss) | + | + | ||||||
| Frondicularia budensis (Hantk.) | cf. | + | + | + | ||||
| Saracenaria arcuata (d’Orb.) | cf. | + | + | + | ||||
| Vaginulina mexicana Nuttall | + | + | + | + | + | |||
| Marginulina boehmi (Reuss) | + | + | + | + | + | + | ||
| Marginulinopsis fragaria (Guemb.) | + | + | + | + | + | |||
| Anomalina granosa (Hantk.) | + | + | + | + | + | + | + | + |
| A. hantkeni (Rzehak) | + | |||||||
| Anomalinoides affinis (Hantk.) | + | + | + | + | + | |||
| Cibicidoides aff. ungerianus (Cushm.) | + | + | + | |||||
| Heterolepa dutemplei (d’Orb.) | + | + | ||||||
| H. eocaena (Guemb.) | + | + | + | + | + | + | ||
| H. pygmea (Hantk.) | + | + | ||||||
| Alabamina almaensis (Samoil.) | + | + | ||||||
| Siphonina subreticulata (Mjatl.) | + | |||||||
| Falsoplanulina biconvexa Bugrova | + | + | ||||||
| Planulina costata (Hantk.) | cf. | + | + | + | + | + | + | + |
| Queraltina epistominoides Marie | + | |||||||
| Bulimina sculptilis Cushm. | + | + | + | + | + | + | ||
| Uvigerina hispida Schwag. | + | + | ||||||
| U. jacksonensis Cushm. | + | + | + | |||||
| U. nuda Gohrband | + | |||||||
| U. pygmea d’Orb. | + | + | + | |||||
| Bolivina antegressa Subb. | + | + | + | + | + | + | ||
| B. microlancetiformis Subb. | + | + | + | |||||
| Bolivinoides reticulatus (Hantk.) | + | + | + | + | + | |||
| Loxostomoides millepunctatus (Tutkow.) | + | + | + | |||||
Автором статьи бентос изучен из разрезов Джебель Коле и Вади Аль Джизель (центральная часть Пальмирид). В первом из них найдены окатанные раковины нуммулитов, чешуя, зубы и фрагменты костей рыб, копролиты. Раковины фораминифер плохой сохранности, часто раздробленные, трудно определимые. Из планктонных форм обнаружены Globigerinatheka tropicalis (Blow et Banner), Hantkenina sp. ind., Turborotalia sp. ind. Содержание МБФ значительное, и его состав обновляется; здесь присутствуют Laevidentalina communis (d’Orb.), Nodosaria bacillum Defr., Siphonodosaria exilis (Neugeb.), Plectofrondicularia cf. striata Hantken, Robulus limbosus (Reuss) (и еще не менее 6 видов этого рода), Vaginulinopsis cumulicostatа (Guemb.), Marginulina cf. boеhmi (Reuss), Мarginulinopsis fragaria (Guemb.), Anomalina granosa (Hantken), Heterolepa eocaena (Guemb.), H. dutemplei (Franzenau), Falsoplanulina biconvexa Bugrоva, Bulimina sculptilis Cushm., Uvigerina eocena Guemb., U. nuda Gohrband, U. aff. pygmea d’Orb. (разрез Джебель Коле). В разрезе Вади Аль Джизель обнаружены лишь Amphimorphina sp. ind., Cibicidoides sp. и мелкие Bolivina spp. К этой фауне центральной части Пальмирид следует добавить характерные виды Planulina costata, Clavulinoides szaboi, Bolivinoides reticulatus из приведенного выше списка В.А. Крашенинникова (табл. 1).
Отложениям зон Р15–Р17 соответствует региональная зона Planulina costata по МБФ (Бугрова, 2016). Ее руководящие виды известны в разрезах Египта (Крашенинников, Поникаров, 1964; Krasheninnikov, Ponikarov, 1965), широко распространены в приабонских отложениях Южной и Центральной Европы (Hantken, 1875; Grünig, 1985; Sztrаkos, 1987; Horváth-Kollányi, 1988; Popov et al., 2001; Cimerman et al., 2006; Стоjaнoвa, Петров, 2008 и др.), в Армении (Закревская и др., 2017). Они составляют зональный комплекс одноименной зоны верхнего эоцена Перитетиса (Бугрова, 1988б, 2001б, 2004; Практическое…, 2005; Зональная…, 2006). В приабонское время связь бассейнов Средиземноморья и Перитетиса была обширной и свободной (Попов и др., 2009, рис. 7 ).
Олигоцен
Сведения о бентосных фораминиферах олигоцена приведены по публикациям В.А. Крашенинникова (1964; Krasheninnikov et al., 1996). На территории Сирии в олигоценовое время преобладающими стали мелководные отложения: детритусовые, лепидоциклиновые и водорослево-коралловые известняки, в которых МБФ не обнаружены. К песчано-глинистым породам Пальмирид и некоторых других регионов приурочены остатки планктона нового состава. По ПФ В.А. Крашенинниковым были выделены зоны Globigerina tapuriensis, Globigerina selli, Globigerina ampliapertura (рюпельский ярус), а также зоны Turborotalia opima и Globigerina ciperoensis (хаттский ярус). Им соответствуют три региональные зоны по бентосу.
Зона Almaena taurica выделена по нахождению видов Spiroplectammina carinata (d’Orb.) sensu Krasheninnikov, Alabamina almaensis (Samоil.), Cibicidoides pseudoungerianus (Cushm.), Almaena taurica (Samoil.), Caucasina buliminoides Bogd., Bolivina microlancetiformis Subb. и др. Большинство из этих видов, включая зональный, присутствуют в верхах эоцена Перитетиса (Практическое…, 2005, табл. 3 ).
Зона Cibicidoides pseudoungerianus характеризуется нахождением Cibicidoides pseudoungerianus (Cushm.), C. oligocenicus (Samoil.), Heterolepa a-lmensis (Samoil.), Valvulineria petrolei (Andreae), Cibicidina amphysiliensis (Andreae), Melonis dosularensis Chalil., Ceratobulimina contraria (Reuss), Angulogerina gracilis (Reuss), Bolivina beyrichi Reuss. Из зонального комплекса практически исчезают эоценовые формы и преобладают олигоценовые виды Западной Европы. Обе зоны были отнесены к рюпельскому ярусу.
Зона Cibicides sigmoidalis содержит достаточно разнообразный комплекс, включающий виды Cibicides sigmoidalis Subb., C. tenellus (Reuss), Valvulineria petrolei (Andreae), Almaena osnabrugensis (Roemer), Elphidium inflatum (Reuss), E. minutum (Reuss), Bitubulogerina kesselensis (Batjes), Uvigerina longa Cushm. et Berm., Bolivina fastigia Cushm., Chilostomella cylindroides Reuss и др., характерные для хаттского яруса Западной Европы и крайне редко встречающиеся в разрезах Перитетиса. Кроме них, представлены виды тетических родов крупных и мелких бентосных фораминифер, которые в разрезах Перитетиса неизвестны.
Сирийский бассейн до конца олигоцена оставался тепловодным нормально-морским в отличие от сократившихся по площади морей Перитетиса (Попов и др., 2009, рис. 12, 13 ) с непостоянной соленостью вод.
В последние годы на территории Пальмирид отложения эоцена и олигоцена были вскрыты рядом скважин (Hernitz Kučenjak et al., 2006), в которых по ПФ выделены зоны P15–P22 и E15–O6. Зональные границы определялись по последнему нахождению вида-индекса. Олигоценовые отложения охарактеризованы пятью зонами: О1 Pseudohastigerina naguewichiensis, О2 Turborotalia ampliapertura, О3 Globigerina sellii, О4 Globigerina angulisuturalis/Chiloguembelina cubensis (рюпельский ярус) и О5 Paragloborotalia opima (хаттский ярус), сопоставление которых с зонами В.А. Крашенинникова не приводится. Сведений о МБФ в работе (Hernitz Kučenjak et al., 2006) не содержится.
Отложения олигоцена в Сирии трансгрессивно перекрыты осадками морского нижнего миоцена или континентальными осадками.
ЗАКЛЮЧЕНИЕ
1. Впервые проведено биостратиграфическое деление отложений палеогена Сирии по обобщенным сведениям о таксономическом составе и распространении мелких бентосных фораминифер в рамках зональности по планктонным фораминиферам, разработанной В.А. Крашенинниковым.
Выделены региональные подразделения – комплексные зоны и слои с видом-индексом (рис. 3). Палеоценовый отдел включает зону Stensioeina beccariiformis–Anomalina danica, зону Vaginulina robusta–Pyramidina crassa и зону Reussella paleocenica. К эоцену относятся зона Pseudogaudryina soldadoensis–Cibicidoides lybicus (низы ипрского яруса); зона Hydromylinella wittpyuti (верхи ипрского–низы лютетского ярусов); слои с Paragaudryina dalmatina–Hydromylinella wittpyuti (лютетский ярус); зона Falsoplanulina ammophila (верхи лютетского–низы бартонского ярусов); слои с Queraltina epistominoides (бартонский ярус); зона Planulina costata (приабонский ярус).
2. Проведено сравнение состава фораминифер палеогена Сирии и Крымско-Кавказской области, как представителей биот Тетиса и северо-восточного Перитетиса соответственно. Общие закономерности распространения планктонных фораминифер и одинаковая последовательность близких по составу их ассоциаций в субглобальных масштабах свидетельствуют об их эволюционной природе. Однако планктон умеренно тепловодного Перитетиса был менее разнообразен, а тейльзоны общих видов отличались по продолжительности от их биозон в приэкваториальных водах Сирийского бассейна. Вследствие этого установление границ зон “стандартной” шкалы современной МСШ по появлению индекс-таксонов в регионах Перитетиса не может дать однозначных результатов.
3. Состав мелких бентосных фораминифер и последовательность смены их комплексов в разрезах Сирии и Крымско-Кавказской области имеют много общего, что свидетельствует о связи этих бассейнов в течение палеоцена и эоцена, а также о корреляционном потенциале бентосной группы мелких фораминифер. В периоды региональных событий дефицита кислорода в Перитетисе (в позднем палеоцене и в конце бартона) бентосная фауна рассматриваемых бассейнов значительно отличалась по родовому и видовому составу.
По обновлению и общности состава бентосных фораминифер в обеих областях выделена единая зона Planulina costata приабона, содержащая многие виды, распространенные на том же стратиграфическом уровне в Западной Европе и Средиземноморье.
4. При единообразном понимании исследователями видов МБФ возрастет значение этой группы фауны для межрегиональных корреляций и палеобиогеографических построений.
Благодарности. Я сохраняю глубокую благодарность В.А. Крашенинникову за коллекционный материал, который был передан мне для изучения бентосных фораминифер палеогена Сирии.
Список литературы
Абдулрахим А.Х., Немков Г.И. Зональное расчленение палеогеновых отложений Дамасской области Сирии по нуммулитидам // Изв. высших учебных заведений. Геология и разведка. 1972. № 7. С. 27–31.
Атлас литолого-палеогеографических карт СССР. Т. 4. Палеогеновый, неогеновый и четвертичный периоды. М.: ГУГК, 1967.
Беньямовский В.Н. Обоснование детальной стратиграфической схемы нижнего палеогена Крымско-Кавказской области // Пути детализации стратиграфических схем и палеогеографические реконструкции. М.: ГЕОС, 2001. С. 210–223.
Беньямовский В.Н. Стратиграфическая шкала среднего эоцена Крымско-Кавказской области по планктонным фораминиферам: темпы эволюции, этапы и экология // Темпы эволюции органического мира и биостратиграфия. Материалы LVII сессии Палеонтол. общества при РАН. Санкт-Петербург, 2011. С. 29–30.
Бугрова Э.М. Новые спирально-плоскостные нодозарииды // Палеонтол. журн. 1985. № 1. С. 42–51.
Бугрова Э.М. Зональное деление эоцена Бахчисарайского района Крыма по мелким фораминиферам // Изв. АН СССР. Сер. геол. 1988а. № 1. С. 82–91.
Бугрова Э.М. Зональное деление эоцена юга СССР по бентосным фораминиферам // Докл. АН СССР. 1988б. Т. 300. № 1. С. 169–171.
Бугрова Э.М. Мелкие бентосные фораминиферы зональных комплексов эоцена юга СССР // Ежегодник ВПО. 1989. Т. 32. С. 13–29.
Бугрова Э.М. Развитие и расселение фораминифер в посткризисную эпоху раннего палеогена // Стратиграфия. Геол. корреляция. 1999. Т. 7. № 2. С. 54–60.
Бугрова Э.М. Позднепалеоценовое событие, Северный Кавказ // Использование событийно-стратиграфических уровней для межрегиональной корреляции фанерозоя России. Методическое пособие. СПб.: Изд-во ВСЕГЕИ, 2000а. С. 94–97.
Бугрова Э.М. Среднеэоценовое (кумское) событие, Северный Кавказ // Использование событийно-стратиграфических уровней для межрегиональной корреляции фанерозоя России. Методическое пособие. СПб.: Изд-во ВСЕГЕИ. 2000б. С. 97–99.
Бугрова Э.М. Новые таксоны бентосных фораминифер палеогена СНГ // Палеонтол. журн. 2001а. № 6. С. 3–9.
Бугрова Э.М. Стратиграфическое и географическое распространение верхнеэоценовых фораминифер на северных окраинах бассейна Тетис // Стратиграфия. Геол. корреляция. 2001б. Т. 9. № 2. С. 92–104.
Бугрова Э.М. Инфузории и проблематики палеогена северо-восточных окраин бассейна Тетис // Палеонтол. журн. 2003. № 1. С. 3–9.
Бугрова Э.М. Фораминиферы палеогена центральной части Северного Кавказа и Предкавказья // Атлас важнейших групп фауны мезозойско-кайнозойских отложений Северного Кавказа и Предкавказья. СПб.: Изд-во ВСЕГЕИ, 2004. С. 80–100.
Бугрова Э.М. Палеоген Туркменистана (Обновленная схема зонального расчленения и корреляции) // Нефтегазовая геология. Теория и практика. 2009. http://www.ngtp.ru
Бугрова Э.М. Новые данные о семействе Caucasinidae N. Bykova (фораминиферы позднего мела–миоцена) // Палеонтол. журн. 2012. № 2. С. 25–33.
Бугрова Э.М. Проблемы проведения границ подразделений МСШ при обновлении региональных стратиграфических схем палеогена России и СНГ // Общая стратиграфическая шкала России: состояние и перспективы обустройства. Сб. статей. Москва: ГИН РАН, 2013. С. 329–331.
Бугрова Э.М. Границы ярусов палеогена МСШ в региональной стратиграфической схеме Таджикистана // Геология и вопросы сейсмичности территории Таджикистана. Материалы научной конференции, посвященной 80-летию академика Джалилова Манзура Рахимовича. Душанбе, 2014. С. 36–43.
Бугрова Э.М. Бентосные фораминиферы палеогена Сирии (стратиграфия, сравнение с фауной Перитетиса) // Материалы LXII сессии Палеонтол. общества при РАН. Санкт-Петербург, 2016. С. 36–37.
Бугрова Э.М., Письменная Н.С. О положении границы лютетского и бартонского ярусов в регионах Северного Кавказа // Вестник Южно-Росс. гос. техн. университета. Сер. Науки о Земле. 2009. № 1. С. 155–162.
Бугрова Э.М., Старшинин А.С. Новое о развитии северо-восточной части Таджикской депрессии в раннепалеогеновое время // Стратиграфия. Геол. корреляция. 2013. Т. 21. № 6. С. 88–92.
Введение в изучение фораминифер. Классификация мелких фораминифер мезо-кайнозоя. Ред. Субботина Н.Н., Волошинова Н.А., Азбель А.Я. Л.: Недра, 1981. 209 с.
Геология и полезные ископаемые зарубежных стран. Сирия. Л.: Недра, 1969. 215 с. (Труды НИЛ Зарубежгеология. Вып. 18).
Закревская Е.Ю., Бугрова Э.М., Щербинина Е.А., Саакян Л.Г., Айрапетян Ф.А., Григорян Т.Е., Волошина О.В. Микропалеонтологическая характеристика среднеэоценовых–нижнеолигоценовых отложений разрезов Ланджар и Урцаландж в Южной Армении // Бюлл. МОИП. Отд. геол. 2017. Т. 92. Вып. 5. С. 60–85.
Зональная стратиграфия фанерозоя России. Палеогеновая система. Научн. ред. Корень Т.Н. СПб.: Изд-во ВСЕГЕИ, 2006. С. 172–193.
Крашенинников В.А. Значение фораминифер открытых тропических бассейнов датского и палеогенового времени для разработки Международной стратиграфической шкалы // Вопросы микропалеонтологии. 1964. Вып. 8. С. 190–213.
Крашенинников В.А. Зональная стратиграфия отложений датского яруса // Зональная стратиграфия палеогена Восточного Средиземноморья. М.: Наука, 1965а. С. 5–27 (Тр. ГИН АН СССР. Вып. 133).
Крашенинников В.А. Зональная стратиграфия палеогеновых отложений // Докл. советских геологов на XXII сессии Международного геол. конгресса. Проблема стратиграфии кайнозоя. М.: Недра, 1965б. С. 37–61.
Крашенинников В.А. Стратиграфия палеогеновых отложений Сирии // Зональная стратиграфия палеогена Восточного Средиземноморья. М.: Наука, 1965в. С. 28–75 (Тр. ГИН АН СССР. Вып. 133).
Крашенинников В.А., Басов И.А. Стратиграфия палеогеновых отложений Мирового океана и корреляция с разрезами на континентах. М.: Научный мир, 2007. 316 с. (Тр. ГИН АН СССР. Вып. 432).
Крашенинников В.А., Немков Г.И. Соотношение фаун планктонных фораминифер и нуммулитов в палеогеновых отложениях Сирии // Вопросы микропалеонтологии. 1975. Вып. 18. С. 179–211.
Крашенинников В.А., Поникаров В.П. Стратиграфия мезозойских и палеогеновых отложений Египта // Сов. геология. 1964. № 2. С. 42–71.
Минтузова Л.Г. Детальная стратиграфия палеоцен-э-оценовых отложений Керченского полуострова и их корреляция с прилегающими районами Крыма // Биостратиграфические основы создания стратиграфических схем фанерозоя Украины. Киев: Институт геологии НАН, 2008. С. 150–158 (на укр. яз.).
Попов С.В., Ахметьев М.А., Лопатин А.В. и др. Палеогеография и биогеография бассейнов Паратетиса. Часть 1. Поздний эоцен–ранний миоцен. М.: Научный мир, 2009. 200 с. (Тр. ПИН РАН. Т. 292).
Попов С.В., Табачникова И.П., Пинчук Т.Н., Ахметьев М.А., Запорожец Н.И. Опорный разрез эоцена р. Белая, Адыгея, Западное Предкавказье // Стратиграфия. Геол. корреляция. 2018. Т. 26. № 6. С. 65–79.
Практическое руководство по микрофауне. Т. 8. Фораминиферы кайнозоя. Научн. ред. Бугрова Э.М. СПб.: Изд-во ВСЕГЕИ, 2005. 324 с.
Саидова Х.М. О современном состоянии системы надвидовых таксонов кайнозойских бентосных фораминифер. М.: Институт океанологии им. П.П. Ширшова, 1981. 70 с.
Cтоjaнoвa В., Петров Г. Стратиграфско распространение на фораминиферите од Тиквешкиот Басен во Република Македонија // Posebno izdanie na Geologica Macedonica. 2008. V. 22. № 2. P. 63–74.
Стратиграфический кодекс России. Издание третье. Отв. ред. Жамойда А.И. СПб.: Межведомственный стратиграфический комитет России, ВСЕГЕИ, 2006. 96 с.
Халилов Д.М. Микрофауна и стратиграфия палеогеновых отложений Азербайджана. Баку: Изд-во АН Азерб. ССР, 1962. 325 с.
Alegret L., Molina E., Thomas E. Benthic foraminiferal turnover across the Cretaceous/Paleogene boundary at Agost (southeastern Spain). Paleoenvironmental inferences // Marine Micropaleontology. 2003. № 48. P. 251–279.
Ansary S.E. Report on the foraminiferal fauna from the Uhher Eocene of Egypt // Publ. Inst. Deserte Egypte. 1955. № 6. 106 p.
Bendjamini Ch. Planktonic foraminiferal biostratigraphy of the ’Avedat Group (Eocene) of the northern Negev, Israel // J. Paleontol. 1980. V. 54. № 2. P. 325–358.
Beniamovskii V. Dynamics of development of benthic Paleocene–Early Eocene foraminifera faunas of the European paleobiographic area (EPA) // STRATA. Sér. 1. 1998. V. 9. P. 29–32.
Beniamovskii V.N., Oberhäensli H., Alekseev A.S., Overchkina M.N. The anoxic Kuma Formation (Latest Lutetian–Earliest Priabonian) basin of Eastern Peri-Thetys: an environmental study // Early Paleogene warm climates and biosphere dynamics. Abstracts. Goteborg, 1999.
Benyamovskiy V.N. Detailed planktonic foraminiferal zonation of Middle Eocene in Crimean-Caucasus Region of Northeastern Peritethys // Berichte Geol. B.-A. 2011. V. 85. S. 38.
Berggren W.A., Aubert J. Paleocene benthic foraminiferal biostratigraphy, paleobiogeography and paleoecology of Atlantic-Tethyan region: Midway-type fauna // Palaeogeogr. Palaeoclimat. Palaeoecol. V. 18. 1975. P. 73–192.
Berggren W.A., Kent D.V., Swisher C.C., Aubry M.-P. A revised Cenozoic geochronology and chronostratigraphy // Soc. Econ. Pal. Miner. Spec. Publ. 1995. № 54. P. 129–212.
Bugrova E.M. Recovery of North Caucasus foraminiferal assemblages after the pre-Danian extinction // Biotic Recovery from Mass Extinction Events. London. Geol. Soc. Spec. Publ. 1996. № 102. P. 337–342.
Cimmerman F., Jelen B., Skaberne D. Late Eocene benthic foraminiferal fauna from clastic sequence of the Socka–Dobrna area and its chronostratigraphic importance (Slovenia) // Geologija (Ljubljana). 2006. V. 49. № 1. P. 7–44.
Grünig A. Systematical description of Eocene benthic Foraminifera of Possagno (Northern Italy), Sansoain (Northern Spain) and Biarritz (Aquitain, France) // Mem. Inst. Geol. Mineral. Univ. Padova. 1985. V. 37. P. 251–302.
Hantken M. Die Fauna der Clavulina szaboi – Schichten // Königlich-Ungarische Geol. Anst. 1875. Bd. 4. Th. 1. S. 1–93.
Hernitz Kučenjak M., Premec Fućek V., Slavković R., Mesić I.A. Planktonic Foraminiferal Biostratigraphy of the Late Eoc-ene and Oligocene in the Palmyride Area, Syria // Geologia Croatica. 2006. V. 59/1. P. 9–39.
Horváth-Kollányi K. Eocene bentonic smaller Foraminifera fauna from Dudar // Ann. Inst. Geol. Publici Hungarici. 1988. V. 63. Fasc. 4. P. 6–173.
Krasheninnikov V.A. Syria. Paleogene // Geological Framework of the Levant. V. I. Part II. Jerusalem, 2005. P. 299–342.
Krasheninnikov V.A., Ashourov A.A. Relation of pelagic and shallow-water (nummulitic) facies in the Paleogene of Syria and distribution of chertphosphate-oil shale rocks // II Int. Symp. of the Geology of the Eastern Mediterranian region. Jerusalem, 1995. P. 12.
Krasheninnikov V.A., Ponikarov V.P. Zonal stratigraphy of Paleogene in the Nile Valley // Geol. Surv. Min. Res. Depart. Cairo. 1965. № 32. P. 1–26.
Krasheninnikov V.A., Golovin D.I., Mouravyov V.I., with collaboration of Helon R., Nseir H., Shweki A. The Paleogene of Syria – Stratigraphy, Lithology, Geochronology // Geol. Jb., Reiche B. 1996. H. 86. S. 3–136.
Liebus A. Die Foraminiferenfauna der Mitteleocänen Mergel von Norddalmatien // K. Akad. Wiss. Wien. Math.-Naturwiss. Kl. Sitzungsber. 1911. Bd. 120. T. 1. S. 865–956.
Loeblich A.R., Tappan H. Foraminiferal genera and their classification. New York: Van Nostrand Rienhold Co, 1988. 970 p.
Luterbacher H.P., Premoli Silva I. Biostratigrafia del limite Cretaceo-Terziario nell’ Appennino centrale // Rivista Ital. Paleontol. Stratigr. 1964. V. 70. № 1. P. 67–128.
Luterbacher H.P., Ali J.R., Brinkhuis H. et al. The Paleogene period // A Geological Time Scale 2004. Eds. Gradstein F.M., Ogg J.G., Smith A.G. Cambridge: Cambridge University Press, 2004. P. 384–408.
Pardo A., Adatte Th., Keller G., Oberhänsli H. Paleoenvironmental changes across the Cretaceous–Tertiary boundary at Koshak, Kazakhstan, based on planktic foraminifera and clay mineralogy // Palaeogeogr. Palaeoclim. Palaeoecol. 1999. № 154. P. 247–273.
Popov S.V., Akhmetiev M.A., Bugrova E.M. et al. Biogeography of the Northern Peri-Tethys from the Late Eocene to the Early Miocene. Part 1. Late Eocene // Paleontol. J. 2001. V. 35. Suppl. 1. P. S1–S68.
Speijer R.P., Van der Zwaan G.J. Extinction and survivorship of southern Tethyan benthic foraminifera across the Cretaceous/Paleogene boundary // Biotic Recovery from Mass Extinction Events. Geol. Soc. Spec. Publ. 1996. № 102. P. 343–371.
Sztrakos K. Les Foraminiferes bartoniens et priaboniens des couches a “Tritaxia szaboi” de Hongrie et essai de reconstitution paleogeographique de la Montagne Centrale de Hongrie au Bartonien et Priabonien // Cahiers de Micropaleontologie. 1987. V. 2. № 1. P. 5–37.
Vandenberghe N., Higen F.J., Speijer R.P. et al. The Paleogene Period // The Geologic Time Scale 2012. Eds. Gradstein F.M. Cambridge: Cambridge University Press, 2012. P. 855–921.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция