

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 4, стр. 125-141
О возрасте буор-кемюсской флоры Северо-Востока Азии на основе материала из неморского мела Восточной Чукотки
С. В. Щепетов 1, *, А. Б. Герман 2, П. Л. Тихомиров 3, 4, А. В. Моисеев 2, С. Д. Соколов 2, Я. Хаясака 5
1 Ботанический институт РАН им. В.Л. Комарова
Санкт-Петербург, Россия
2 Геологический институт РАН
Москва, Россия
3 Московский государственный университет им. М.В. Ломоносова, Геологический факультет
Москва, Россия
4 Северо-Восточный комплексный научно-исследовательский институт ДВО РАН
Магадан, Россия
5 Graduate School of Science, Department of Earth and Planetary Systems Science, Hiroshima University
Hiroshima, Japan
* E-mail: shchepetov@mail.ru
Поступила в редакцию 14.03.2019
После доработки 11.04.2019
Принята к публикации 20.11.2019
Аннотация
На основании анализа литературных и картографических материалов, полевых наблюдений авторов, результатов U–Pb датирования цирконов и ознакомления с палеоботаническими коллекциями показано, что “мезофитная” папоротнико-голосеменная буор-кемюсская флора существовала на Восточной Чукотке дольше обычного в позднемеловую эпоху. Осадочные отложения ольховской свиты отчасти фациально замещают вулканогенные накопления нырвакинотской толщи. Оба стратона содержат остатки растений буор-кемюсской флоры. Вулканогенная амгеньская толща согласно без перерыва перекрывает нырвакинотскую и содержит “кайнофитные” комплексы растений со значительным количеством покрытосеменных, датируемые туроном–коньяком. Ранее эти три стратона входили в состав единой этелькуюмской свиты. U–Pb датировки цирконов из вулканогенных и вулканогенно-осадочных пород нырвакинотской толщи показали, что эта толща формировалась в позднем мелу и ее возраст примерно соответствует интервалу 93–87 млн лет (турон–коньяк). С учетом всех данных наиболее вероятным представляется, что буор-кемюсская флора (по крайней мере в пределах Восточно-Чукотского сегмента Охотско-Чукотского пояса) существовала в альбе, сеномане, туроне и, возможно, в начале коньяка.
ВВЕДЕНИЕ
На Северо-Востоке Азии широко распространены континентальные отложения мелового возраста. Они заполняют несколько осадочных бассейнов и входят в состав Удско-Мургальского и Охотско-Чукотского вулканогенных поясов (рис. 1).
Рис. 1.
Распространение образований Охотско-Чукотского вулканогенного пояса, а также местоположение захоронений остатков растений и возрастные соотношения основных палеофлор.
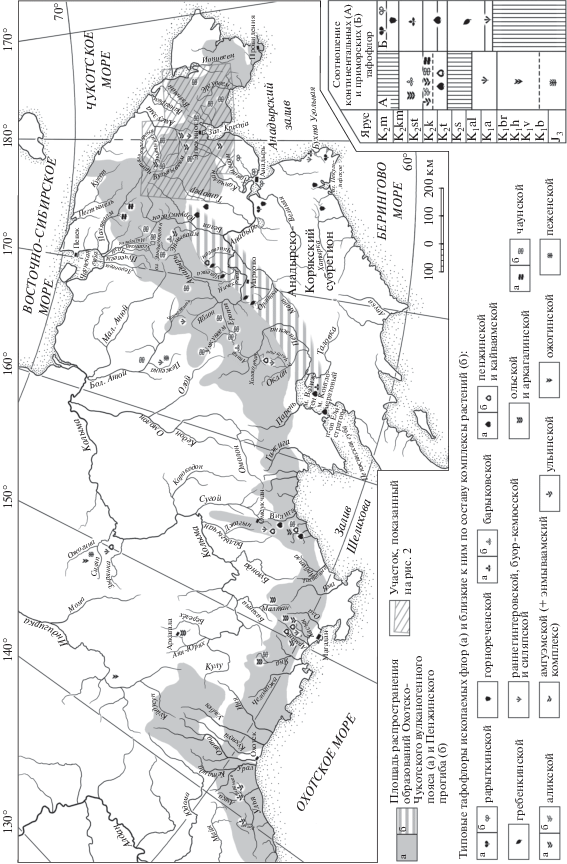
Охотско-Чукотский вулканический пояс (ОЧВП), при современном эрозионном срезе, имеет протяженность около 3200 км и представлен множеством более или менее изолированных полей вулканических пород, разделенных выходами интрузий и более древних осадочных отложений. Граница ОЧВП со стороны континента носит условный характер: вулканические поля есть и за ней, но их там значительно меньше. Планомерное изучение меловых вулканогенных образований началось в середине XX века, и сразу же встал вопрос об обосновании их возраста. Остатков морских моллюсков и фораминифер континентальные образования не содержат, споры и пыльца растений в вулканогенных толщах, как правило, не сохраняются, а изотопные методы тогда были мало пригодны для стратиграфических корреляций. Однако в терригенных отложениях и в туфовых прослоях вулканогенных толщ нечасто, но регулярно встречаются остатки ископаемых растений, которые широко использовались и используются для определения возраста вмещающих отложений.
На рисунках и в тексте мы используем термины “палеофлористический комплекс” или “тафофлора” – это элементарные палеофлористические единицы, представляющие собой “совокупность ископаемых растений из одного или нескольких территориально и стратиграфически близких местонахождений” (Герман, 2011, с. 10). Тафофлоры, или палеофлористические комплексы, обладающие существенно сходными чертами, т.е. характерным сочетанием таксонов, качественным и количественным соотношением групп растений и т.д., мы рассматриваем как ископаемую флору или палеофлору, характеризующую определенный этап развития древней флоры значительной территории (например, Северного Приохотья). Для обозначения совокупности растений, существовавших на каждом из региональных этапов развития флоры, В.А. Самылина (1974, 1976) использовала термин “стратофлора”.
К началу 70-х гг. ХХ века был накоплен значительный палеофлористический материал, который обобщила Самылина (1974). Она предложила для региона в целом базовую схему развития растительного мира в меловом периоде. Это дало возможность определять возраст флороносных отложений в относительно узких пределах и коррелировать эти отложения друг с другом. Основой построений Самылиной послужило представление о том, что в середине мелового периода происходил переход от флор мезофита с доминированием папоротников и голосеменных к флорам кайнофита, в которых существенную роль стали играть покрытосеменные растения. Растительный мир быстро эволюционировал, причем изменения состава флор происходили, по ее мнению, близко по времени или одновременно на всей территории региона.
Позже, однако, было установлено, что на территории Северо-Востока России меловая флора развивалась по-разному в зависимости от палеоландшафтной обстановки (Герман, 1993; Щепетов, 1995). Если во флорах приморских аллювиальных низменностей и равнин, располагавшихся к востоку от ОЧВП, в середине мела действительно шло быстрое нарастание элементов кайнофита, то рядом, в области вулканизма ОЧВП, количество покрытосеменных в некоторых районах заметно увеличилось, в других же продолжали доминировать древние группы растений, среди которых возникло множество новых таксонов.
В пределах ОЧВП наиболее древняя флора, в составе которой обильно представлены покрытосеменные растения, получила название “арманская” по имени наиболее представительной тафофлоры с р. Армань (Самылина, 1974). По современным данным ее возраст соответствует турону–коньяку (Herman et al., 2016). Ей предшествовала региональная буор-кемюсская флора, включающая многочисленные тафофлоры внутриматериковых впадин, ОЧВП и приморских низменностей. В ее комплексах растений покрытосеменные крайне редки или не встречены вовсе (Самылина, 1974; Герман, 2011). Представления о возрасте буор-кемюсской флоры не менялись со времен ее первоописания (Самылина, 1964, 1967, 1974, 1976): она и ныне датируется ранним–средним альбом и, вероятно, частью позднего альба (Герман, 2011; Решения…, 1978, 2009). Долгое время считалось, что между буор-кемюсской и арманской флорами существовала еще одна региональная флора, получившая название топтанской (Самылина, 1976). Однако, в конце концов, было признано, что самостоятельного топтанского этапа развития флоры Северо-Востока России не было, а флористический комплекс топтанской свиты следует рассматривать в составе буор-кемюсской флоры (Решения…, 2009).
Таким образом получается, что буор-кемюсская флора закончила свое существование в начале или середине позднего альба, а сменившая ее в пределах ОЧВП арманская флора появилась только в туроне или даже в коньяке. Даже с учетом некоторой условности наших привязок палеофлор к Общей стратиграфической шкале, между временем существования этих двух флор оказывается огромный перерыв. Причем признаков такого перерыва в разрезах ОЧВП, включающих флороносные отложения, не наблюдается; и более того, есть все основания предполагать более или менее непрерывное накопление вулканогенных и вулканогенно-осадочных отложений в позднеальбское–туронское время.
Чтобы объяснить такое противоречие, приходится предположить, что либо арманская флора могла начать свое существование уже в позднем альбе, либо буор-кемюсская флора в пределах ОЧВП продолжала существовать по крайней мере до начала турона включительно. Кроме того, нельзя исключать, что между буор-кемюсской и арманской флорами все-таки существовала еще какая-то переходная флора, которую палеоботаники просто не узнают при изучении небольших флористических комплексов, относя их либо к буор-кемюсской, либо к арманской флоре. В этой связи особый интерес приобретают данные о тафофлорах, которые могут оказаться представителями как раз такой – переходной, промежуточной – флоры. Именно в этом качестве Г.Г. Филиппова (1997, 2001) рассматривает флористические комплексы из нырвакинотской толщи Восточной Чукотки.
Начиная с 1990-х гг. для решения вопросов стратиграфии континентальных вулканогенных толщ Северо-Востока Азии успешно привлекаются результаты U–Pb и 40Ar/39Ar датирования мономинеральных фракций (Kelley et al., 1999; Акинин, Миллер, 2011; Tikhomirov et al., 2012). Прецизионные методы изотопного датирования позволяют определить возраст толщ, бедных растительными остатками, и в большинстве случаев обеспечивают меньшую погрешность оценки. Информация, полученная при изотопном датировании вулканических пород, уже использовалась для уточнения представлений о возрасте различных палеофлористических комплексов (Kelley et al., 1999; Spicer et al., 2002; Hourigan, Akinin, 2004; Герман, 2011; Herman et al., 2016; Моисеева, Соколова, 2014).
Цель настоящей статьи – установление возраста тафофлор района залива Креста на Восточной Чукотке на основе таксономического состава флористических комплексов, сведений о структурных взаимоотношениях различных флороносных стратонов и результатов датирования цирконов, извлеченных из вулканитов и вулканомиктовых осадочных пород.
СТРАТИГРАФИЧЕСКОЕ ПОЛОЖЕНИЕ ФЛОРОНОСНЫХ ОТЛОЖЕНИЙ ВОСТОЧНОЙ ЧУКОТКИ
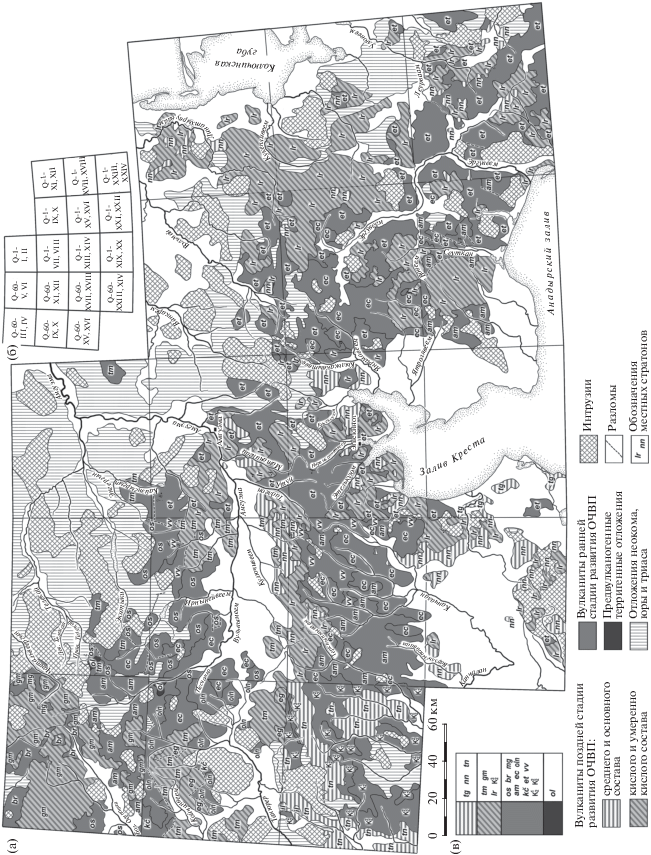
Первой публикацией о раннемеловой флоре Восточной Чукотки, вероятно, является работа В.Ф. Белого и его соавторов (Белый и др., 1965). В более поздней работе для района залива Креста и среднего течения р. Амгуэма Белый (1977) приводит близкий, но не идентичный комплекс растений, который, по его мнению (и, вероятно, мнению В.А. Самылиной), имеет несомненное сходство с буор-кемюсской “стратофлорой”. Остатки растений происходят из вулканогенно-осадочных образований нижней части разреза этелькуюмской свиты. Они замещаются в латеральном направлении и перекрываются вулканитами среднего состава (рис. 2, 3).
Рис. 2.
Распространение альбских–позднемеловых отложений на Восточной Чукотке по данным Государственных геологических карт масштаба 1: 200 000 (а), разграфка листов (б) и стратиграфическая последовательность меловых толщ и свит (в).

Самылина (1976, с. 110–111) приводит следующие данные: “Довольно богатый флористический комплекс собран в 1969 г. З.П. Потаповой и С.Н. Колобашкиным в районе залива Креста (п‑ов Амгень и бассейн р. Матачингай) в пачке алевролитов и песчаников, зажатых среди вулканогенных отложений. Отсюда мной определены Equisetites ramosus Samyl., Equisetites sp., Birisia alata (Pryn.) Samyl., B. onychioides (Vassilevsk. et Kara-Mursa) Samyl., Arctopteris rarinervis Samyl., Sphenopteris sp. cf. Coniopteris ketovae Vassilevsk., Sphenopteris sp., Nilssonia sp., Anomozamites aff. arcticus Vassilevsk., Williamsonia sp., Heilungia oloensis Samyl. et Philipp., H. tschuktschorum Samyl. et Philipp., Thaeniopteris lundgreni Nath., Ginkgo ex gr. adiantoides (Ung.) Heer, Sphenobaiera ex gr. longifolia (Pom.) Florin, Eretmophyllum cf. glandulosum (Samyl.) Krassil., Czekanowskia ex gr. rigida Heer, Phoenicopsis ex gr. angustifolia Heer, Ph. ex gr. speciosa Heer, Leptostrobus marginatus Samyl., Podozamites ex gr. eichwaldii Schimp., P. ex gr. lanceolatus (Lindl. et Hutt.) F. Braun, Pagiophyllum triangulare Pryn., Elatocladus manchurica (Yok.) Yabe, Cephalotaxopsis borealis Samyl., C. cf. intermedia Holl., Cephalotaxopsis sp., Pityophyllum ex gr. nordenskioldii (Heer) Nath., Athrotaxites sp., Parataxodium sp., Desmiophyllum magnum (Samyl.) Samyl., Desmiophyllum sp., Carpolithes sp., Thallites sp. Этот комплекс также может быть уверенно отнесен к буор-кемюсской стратофлоре. Аналогичная флора была определена А.Ф. Ефимовой по сборам С.В. Благодатского из вулканогенно-осадочных образований к северу от залива Креста (Белый и др., 1965)…”.
Самылина (1976) не указывает, из какого стратона происходят растительные остатки. Однако коллекция, определенная ею, хранится в Лаборатории палеоботаники БИН РАН (Санкт-Петербург) под № 3020. При образцах сохранились почти все этикетки, составленные коллекторами, что позволяет уточнить привязку. Сборы проводились в двух точках: на полуострове Амгень в бассейне р. Пийкун (название стратона не указано) и из уэленской свиты на р. Матачингай в месте впадения руч. Клад. Ни на одной из доступных нам карт этот ручей обнаружить не удалось, как и стратон “уэленская свита”, однако на р. Пийкун места сборов остатков растений легко опознаются: на Государственной геологической карте масштаба 1 : 200 000 (лист Q-1-XII, XIV), составленной С.В. Благодатским, флороносные отложения отнесены к этелькуюмской свите, которая включает здесь осадочные, вулканогенно-осадочные и вулканогенные образования среднего состава.
Северо-западнее этого района, на листе Q-60-XI, XII, по данным Е.Е. Петренко (Государственная…, 1982), на морских отложениях берриаса и валанжина с угловым несогласием залегает осадочная ольховская свита. В верховьях р. Бол. Покыткынваам в ее отложениях обнаружены Equisetites sp., Coniopteris setacea (Pryn.) Vachr., Cladophlebis ex gr. williamsonii (Brongn.) Brongn., Arctopteris sp. indet., Heilungia sp., Carpolithes sp., Pityophyllum nordenskioldii (Heer) Nath. В двух других обнажениях установлены Phoenicopsis ex gr. angustifolia Heer, Heilungia oloensis Samyl. et Philipp., Thaeniopteris sp., Ctenis sp. По заключению Г.Г. Филипповой, данные растительные остатки сопоставимы с таковыми из буор-кемюсской свиты альбского возраста.
Можно констатировать, что все известные к концу 70-х годов ХХ века раннемеловые комплексы растений происходят главным образом из отложений этелькуюмской свиты и, в меньшей степени, ольховской свиты.
В 1987–1988 гг. В.В. Романовой и Н.И. Романовым составлена опорная легенда Эгвекинотской серии листов Госгеолкарты-50, включающей и район залива Креста. Легенда обобщила результаты геологических съемок масштаба 1 : 50 000 и тематических работ. В 1992–1993 гг. было проведено геологическое доизучение масштаба 1 : 200 000 площади листа Q-1-XIII, XIV, включающее дополнительное изучение опорных разрезов стратиграфических подразделений. В результате Романова и Романов составили на эту территорию Госгеолкарту-200 второго издания (Санкт-Петербург, 1999 г.).
На новой карте стратон “этелькуюмская свита” не фигурирует (рис. 4). Ранее относимые к нему образования включены В.В. и Н.И. Романовыми в состав следующих стратиграфических подразделений (снизу вверх):
Рис. 4.
Схема распространения меловых отложений в районе залива Креста по данным В.В. Романовой и Н.И. Романова (а) и последовательность местных стратиграфических подразделений (б).

1. Ольховская свита – конгломераты, гравелиты, песчаники, алевролиты. Мощность 1100 м.
2. Нырвакинотская толща – роговообманковые туфы андезитов, андезиты, кварцевые латиты, реже дациандезиты и их туфы, туфоконгломераты, туфогравелиты, туфопесчаники, туфоалевролиты. Мощность 1100 м.
3. Амгеньская толща – игнимбриты и спекшиеся туфы, реже туфы риодацитов, риолитов, трахириолитов и трахидацитов, туффиты, туфопесчаники, туфоалевролиты. Мощность 1500 м.
4. Экитыкинская свита – двупироксеновые андезиты, кварцевые латиты, андезибазальты и их туфы, реже гиперстеновые базальты, туффиты, туфопесчаники, туфогравелиты, туфоалевролиты. Мощность 600 м.
Выше залегает леурваамская свита вулканитов кислого и умеренно кислого состава. Венчают меловой разрез района оливиновые базальты нунлигранской свиты.
Отметим, что в такой интерпретации меловой разрез Восточной Чукотки в целом соответствует разрезу Северного Приохотья (Щепетов, Герман, 2019), но в нем отсутствуют прямые аналоги арманской свиты.
Как уже отмечалось, В.Ф. Белый и В.А. Самылина, вслед за автором первого издания Госгеолкарты-200 С.В. Благодатским, считали терригенные и вулканогенные породы нижней части этелькуюмской свиты фациями и не относили их к разным стратиграфическим уровням. Растительные остатки из тех и других рассматривались совместно и сопоставлялись с буор-кемюсской флорой (Белый и др., 1965; Белый, 1977; Самылина, 1976). Принимая результаты ревизии стратиграфического разреза в районе залива Креста, тафофлоры из нижней части этелькуюмской свиты следует относить к двум разным флористическим комплексам. В частности, остатки растений, определенные Самылиной (1976) с полуострова Амгень, происходят из ольховской свиты, так как по данным В.В. и Н.И. Романовых только флороносные отложения этой свиты развиты в бассейне р. Пийкун.
Следует отметить, что во многих случаях соотношения осадочных отложений ольховской свиты и вулканитов нырвакинотской толщи, показанные на карте В.В. и Н.И. Романовых, кажутся неоднозначными. Авторы карты отмечают, что в двух обнажениях было описано угловое и даже азимутальное несогласие между ольховской свитой и нырвакинотской толщей. Однако толщи сложены разными породами, и механизм их формирования был не одинаков. Наверное, поэтому нам ни разу не приходилось наблюдать самим или встречать в пределах ОЧВП описания согласного залегания вулканогенной толщи на осадочной – контакт всегда в той или иной мере несогласный.
При внимательном изучении рисовки границ на карте В.В. и Н.И. Романовых складывается впечатление, что осадочные породы залегают стратиграфически ниже амфиболовых андезитов, на одном с ними уровне или выше их, то есть имеет место обычное переслаивание и фациальное замещение. Однако в соответствии с принятой легендой осадочные породы должны располагаться только стратиграфически ниже андезитов, и авторам карты пришлось нарисовать множество разломов, часто никак не выраженных в рельефе. По ним блоки горных пород были подняты или опущены на десятки и сотни метров.
Наиболее яркий пример такого “стратиграфического” разлома имеет место на юго-юго-восточном склоне г. Мал. Матачинай (верховья р. Прав. Татлыван (Тадлеоан)). Верхняя часть горы сложена субгоризонтально залегающими андезитами нырвакинотской толщи. Ниже по склонам выходят осадочные породы ольховской свиты. Еще ниже по склону вновь появляются туфы и лавы нырвакинотской толщи, которые опущены сюда по разлому северо-восточного направления. Этот разлом с вертикальным смещением как минимум на 200–300 м проходит поперек относительно пологой части склона, не создавая на нем даже уступа, он вкрест пересекает распадки и ручьи, которые его как бы “не замечают”. Чуть юго-западнее, на другом берегу оз. Резервное, возвышается одиноко стоящая сопка, сложенная осадочными породами. Чтобы они не нарушали принятую последовательность отложений, авторам карты пришлось “вырезать” сопку разломами, по которым блок пород был поднят как минимум на 250 м. По результатам полевых наблюдений П.Л. Тихомирова в 2006 г. представляется наиболее вероятным, что данная пачка осадочных пород расположена в разрезе между двумя горизонтами вулканитов. Отметим, что на геологической карте, составленной С.В. Благодатским, большинства этих разломов нет, а границы между терригенными и вулканогенными образованиями показаны как фациальные переходы.
Для амгеньской толщи на территории Эгвекинотской серии листов Г.Г. Филиппова (1997) приводит списки ископаемых растений из нескольких местонахождений. Среди этих растений присутствует хвойное Metasequoia и значительное количество остатков листьев покрытосеменных, в том числе Quereuxia. Казалось бы, такой состав флористических комплексов свидетельствует о том, что вмещающие отложения значительно моложе подстилающих образований нырвакинотской толщи, в которых не встречено ни одного отпечатка покрытосеменных, и между этими стратонами следует предполагать стратиграфический перерыв. Однако по данным В.В. и Н.И. Романовых амгеньская толща согласно залегает на нырвакинотской, что было зафиксировано в четырех опорных обнажениях (рис. 4). В двух из них – №№ 27 и 48 – предыдущие исследователи не отмечали смены толщ и все накопления относили к этелькуюмской свите. В обнажениях №№ 20 и 32 в соответствии с рисовкой карты С.В. Благодатского представлен контакт этелькуюмской и леурваамской свит. Причем в обнажении № 20 В.В. и Н.И. Романовы отмечают даже грубоволнистую поверхность размыва нижележащей толщи.
В настоящее время существуют две основные точки зрения на механизм формирования толщ субаэральных вулканитов ОЧВП. Согласно одной из них (Лебедев, 1987; Щепетов, 1995; Головнева, 2014), толщи отражают условия осадконакопления на вулканических плато, бронировавших верхний ярус рельефа. Согласно другой (Щепетов, Нешатаева, 2019), сохранившиеся в разрезах вулканиты заполняли центральные части вулканоструктур оседания и не образовывали высокогорного рельефа. Так или иначе, накопление вулканического материала и его эрозия происходили одновременно. В разрезах вулканогенных толщ стратификация обычно выражается сменой структуры и текстуры пород, вариациями их состава. Границы между такими “слоями” иногда имеют признаки локального размыва – следы перерыва между извержениями или сериями извержений ближайших вулканов. Судя по тому, что часть вулканитов амгеньской толщи первоначально рассматривалась вместе с накоплениями нырвакинотской толщи в составе одной – этелькуюмской – свиты, границу между ними первые исследователи восприняли именно как внутреннюю, локальную, не имеющую серьезного значения в строении разреза.
К изложенному выше можно добавить, что, по данным Г.И. Богомолова (Государственная…, 1983), восточнее рассматриваемой территории, на листе Q-1-XV, XVI, в разрезах этелькуюмской свиты наблюдается переслаивание осадочных пород не только с вулканитами среднего состава, но и с дацитами и риодацитами, а также переслаивание андезитов и дацитов.
Мы полагаем, что имеющиеся стратиграфические и картографические данные позволяют обсуждать правомерность отнесения образований, ранее выделявшихся как этелькуюмская свита, к четырем самостоятельным местным стратонам. Наблюдаемые структурные взаимоотношения геологических тел дают основания для вывода об отсутствии значительного перерыва, разделяющего формирование главных компонентов разреза бывшей этелькуюмской свиты.
СРАВНЕНИЕ ФЛОРИСТИЧЕСКИХ КОМПЛЕКСОВ
В своих работах Г.Г. Филиппова (1997, 2001) не рассматривает палеофлору ольховской свиты, а лишь отмечает: “Ольховская свита содержит богатый комплекс растительных остатков, близкий по составу буоркемюсскому флористическому комплексу…” (Филиппова, 2001, с. 96). Из накоплений нырвакинотской толщи она приводит следующий список растений: Coniopteris sp., Tchucotopteris ustinovii Vassilevsk., Asplenium cf. dicksonianum Heer, Cladophl-ebis sp., Taeniopteris aff. rhytidorahis Kryst., Taeniopteris sp. (много), Ginkgo ex gr. lepida Heer, Sphenobaiera sp., Phoenicopsis ex gr. angustifolia Heer, Desmiophyllum sp., Cephalotaxopsis (Taxites) magnifolia Font., C. microphylla Holl., Sequoia fastigiata (Sternb.) Heer, Elatocladus sp., Torreya gracillima (Holl.) Krysht. et Baik., Araucarites anadyrensis Kryst., Podozamites gramineus Heer, P. eichwaldii Schimp., Parataxodium sp., Pityophyllum nordenskioldii (Heer) Nathorst. Остатки этих растений были собраны В.П. Аркавым и А.В. Кукановым из четырех различных местонахождений, так что их совокупность по принятой нами терминологии следует считать палеофлорой.
В составе этой нырвакинотской флоры “по разнообразию родов доминируют хвойные позднемелового облика (Cephalotaxopsis, Sequoia, Torreya, Arauc-arites, Parataxodium), представители древних мезозойских форм имеют подчиненное положение (Sphenobaiera, Desmiophyllum, Podozamites, Pityophyllum)” (Филиппова, 1997, с. 18). Далее Филиппова (1997) отмечает, что папоротник Tchucotopteris ustinovii Vassilevsk. ранее она определяла как Birisia cf. onychioides (Vassikevsk. et K.-M.) Samyl. или Birisia alata Samyl.
“Как показал анализ растительных остатков из отложений нырвакинотской толщи, папоротники, цикадофиты и некоторые виды хвойных по морфологии листьев ближе стоят к палеофлорам из буоркемюсского горизонта (альб), а большинство облиственных побегов хвойных широко распространено в верхнемеловых отложениях Сибирско-Канадской палеофлористической области. Возраст нырвакинотского флористического комплекса определяется поздним альбом–сеноманом…” (Филиппова, 1997, с. 18).
Отметим, что, несмотря на позднемеловой, по мнению Филипповой, облик хвойных, род Cephalotaxopsis совершенно обычен для буор-кемюсских тафофлор Северо-Востока Азии. Остатки представителей родов Sequoia и Parataxodium встречаются в отложениях омсукчанской серии и буор-кемюсской свиты (Самылина, 1976). Представители родов Torreyа и Araucarites в буор-кемюсских тафофлорах действительно не отмечены, однако не следует забывать, что речь идет не о реальных родах, а о морфологических типах побегов, к ним относимых. Поэтому нельзя исключать, что подобные побеги в буор-кемюсских комплексах просто определены иначе, например как Elatocladus. Кроме того, род Araucarites входит в состав “нижней” тафофлоры района Как-Каолак Северной Аляски. Эта флора относится к этапу развития Какповрак, который достаточно надежно коррелируется с буор-кемюсским этапом развития палеофлоры Северо-Востока Азии (Герман, 2011; Herman, 2013).
Списки ископаемых растений из отложений, которые ранее включались в состав этелькуюмской свиты, приведены по данным В.В. и Н.И. Романовых в табл. 1. Эти списки, конечно, нельзя воспринимать буквально – в них, вероятно, собраны определения разных авторов, сделанные в разные годы. В накоплениях ОЧВП кутикулы на отпечатках растений обычно не сохраняются, так что определения были сделаны только по морфологическим признакам ископаемых растений. Вряд ли в нижней части разреза “этелькуюмской свиты” может содержаться сразу по 6 видов Ginkgo и Sphenobaiera и тем более 8 видов Cephalotaxopsis. Тем не менее мы полагаем, что списочные составы растений ольховской свиты и нырвакинотской толщи не имеют признаков разновозрастности. Каждый из них, взятый сам по себе, может быть уверенно отнесен к буор-кемюсской флоре.
Таблица 1.
Список ископаемых растений из отложений, ранее включавшихся в состав этелькуюмской свиты на территории листа Q-1-XIII, XIV (по материалам В.В. и Н.И. Романовых)
| Названия растений | Ольховская свита |
Нырвакинотская толща |
Амгеньская толща |
Экитыкинская свита |
|---|---|---|---|---|
| Equisetites ramosus Samyl. | + | |||
| Equisetites aff. rugosus Samyl. | + | |||
| Equisetites sp. | + | + | ||
| Asplenium dicksonianum Heer | + | |||
| Asplenium sp. indet. | + | |||
| Tchaunia tchaunensis Samyl. et Philipp. | + | + | + | |
| Coniopteris cf. nympharum (Heer) Vachr. | + | |||
| C. cf. setacea (Pryn.) Vachr. | + | |||
| Coniopteris sp. | + | + | + | |
| Cladophlebis denticulate (Brongn.) Font. | + | |||
| C. argutula (Heer) Font. | + | |||
| C. aff. frigida (Heer) Sew. | + | |||
| Cladophlebis sp. | + | + | + | + |
| Tchucotopteris ustinovii Vassilevsk. | + | + | ||
| Tchucotopteris sp. | + | |||
| Onichiopsis psilotoides (Stokes et Webb) Ward | + | + | ||
| O. elongata (Geyl.) Yok. | + | |||
| Onichiopsis sp. indet. | + | |||
| Sphenopteris sp. cf. Coniopteris ketovae Vassilevsk. | + | |||
| Sphenopteris sp. vel Arctopteris sp. | + | |||
| Sphenopteris sp. | + | + | + | + |
| Arctopteris rarinervis Samyl. | + | |||
| Arctopteris sp. | + | |||
| Filicales gen. indet. | + | |||
| Nilssonia af. canadensis Bell | + | |||
| N. cf. orientalis Heer | + | |||
| N. cf. serotina Heer | + | |||
| Nilssonia sp. | + | |||
| Nilssoniopteris cf. prinadae Samyl. | + | + | ||
| Anomozamites aff. arcticus Vassilevsk. | + | |||
| Jacutiella (?) amurensis (Novopokr.) Samyl. | + | + | ||
| Williamsonia sp. | + | |||
| Heilungia oloensis Samyl. et Philipp. | + | |||
| H. tschuktschorum Samyl. et Philipp. | + | |||
| H. cf. amurensis (Novopokr.) Pryn. | + | |||
| Heilungia sp. | + | + | ||
| Thaeniopteris lundgreni Nath. | + | + | ||
| T. cf. rhitidorahis Kryst. | + | + | ||
| T. cf. eurychoron (Schenk.) Pryn. | + | |||
| Thaeniopteris sp. | + | + | + | + |
| Ginkgo digitate (Brongn.) Heer | + | |||
| G. cf. sibirica Heer | + | + | ||
| G. cf. adiantoides (Ung.) Heer | + | + | ||
| G. cf. obovate (Nath.) Sew. | + | |||
| G. ex gr. lepida Heer | + | + | ||
| G. ex gr. huttonii (Sternb.) Heer | + | |||
| Baiera cf. polimorpha Samyl. | + | |||
| Baiera sp. | + | |||
| Sphenobaiera biloba Prin. | + | |||
| S. cf. angustiloba (Heer) Florin | + | |||
| S. cf. longifolia (Pom.) Florin | + | + | ||
| S. cf. uninervis Samyl. | + | |||
| S. ex gr. pulchella (Heer) Florin | + | + | ||
| Sphenobaiera sp. | + | + | ||
| Czekanowskia setacea Heer | + | |||
| Cz. ex gr. rigida Heer | + | |||
| Czekanowskia sp. | + | |||
| Phoenicopsis speciosa Heer | + | + | + | |
| Ph. cf. magnifolia Pryn. | + | |||
| Ph. cf. angustifolia Heer | + | + | + | + |
| Phoenicopsis sp. | + | + | + | |
| Podozamites cf. eichwaldii Schimp. | + | + | + | |
| P. cf. lanceolatus (Lindl. et Hutt.) F. Braun | + | |||
| P. latifolius (Schenk.) Heer | + | |||
| P. cf. distana (Presl.) Braun | + | |||
| P. gramineus Heer | + | |||
| Pagiophyllum triangulare Pryn. | + | |||
| P. ex gr. kryshtofovichii Samyl. | + | |||
| Elatocladus smittiana (Heer) Sew. | + | + | ||
| E. zheltovskii Philipp. | + | |||
| Elatocladus sp. | + | + | ||
| Pityophyllum nordenskioldii (Heer) Nath. | + | + | + | |
| Parataxodium aff. wigginsii Arn. et Lowth. | + | |||
| Parataxodium sp. | + | + | ||
| Cephalotaxopsis (Taxites) microphylla Holl. | + | + | ||
| C. intermedia Holl. | + | + | + | + |
| C. heterophylla Holl. | + | + | + | |
| C. ex gr. brevifolia Font. | + | + | ||
| C. cf. acuminate Krysht. et Pryn. | + | |||
| C. sangarensis Vassilevsk. | + | |||
| C. borealis Samyl. | + | |||
| Cephalotaxopsis sp. | + | + | ||
| Sequoia cf. fastigiata (Sternb.) Heer | + | |||
| S. cf. reichenbachii (Gein.) Heer | + | |||
| S. cf. heterophylla Velen. | + | |||
| Torreya gracillima (Holl.) Krysht. et Baik. | + | + | ||
| Torreya sp. | + | |||
| Araucarites cf. anadyrensis Krysht. | + | + | ||
| A. cf. subacutensis Philipp. | + | |||
| Araucarites sp. | + | + | + | |
| Pseudolarix sp. | + | |||
| Athrotaxites sp. | + | |||
| Picea sp. | + | |||
| Desmiophyllum magnum (Samyl.) Samyl. | + | |||
| Desmiophyllum sp. | + | + | + | + |
| Trochodendroides cf. richardsonii (Heer) Krysht. | + | |||
| Tr. cf. arctica (Heer) Berry | + | |||
| Paraprotophyllum aff. ignatianum (Krysht. et Baik.) Herman | + | |||
| Protophyllum sp. | + | |||
| Celastrophyllum sp. | + | |||
| Magnolia sp. | + | |||
| Zizyphoides sp. (Trochodendroides sp.) | + | |||
| Dicotylophyllum sp. | + | |||
| Phillites sp. | + | + | ||
| Grewiopsis (?) sp. | + | |||
| Carpolithes sp. | + | + |
“Флористический комплекс из амгеньской толщи включает около 30 видов ископаемых растений, собранных… в Вульвывеемской подзоне и на северном побережье залива Креста… В его составе присутствуют почти все таксоны, установленные в отложениях нырвакинотской толши, а также хвойные Elatocladus smittiana (Heer) Sew., E. cf. zheltovskii Philipp., Elatocladus sp., Sequoia ambigua Heer, S. obovata Knowlt., S. reichenbachii (Gein.) Heer, Sequoia sp., единичные отпечатки папоротников Sphenopteris sp., Tchaunia tchaunensis Samyl. et Philipp., Tchaunia sp., Sagenopteris sp., редкие находки покрытосеменных. Последние собраны В.П. Аркавым из стратотипического разреза толщи в бассейне р. Матачингай и представлены Trochodendroides cf. richsrdsonii (Heer) Kryst., Protophyllum aff. ignatianum Krysht. et Baik., Grewiopsis sp., Dicotylophyllum sp… В отложениях амгеньской толщи… на рр. Вульвывеем и Телеакай наряду с разнообразными хвойными присутствуют покрытосеменные Trochodendroides sp., Menispermites sp., Quereuxia angulata (Newb.) Krysht., Quereuxia sp…” (Филиппова, 1997, с. 18). В амгеньской толще встречаются, хотя и редко, остатки Metasequoia. На рассматриваемой территории Г.Г. Филиппова (1997) указывает присутствие этого хвойного лишь в одном захоронении – на руч. Медвежий.
“Таким образом, появление в составе амгеньского комплекса родов Metasequoia, Paraprotophyllum, Grewiopsis и Queureuxia свидетельствует о следующем за нырвакинотским этапе развития наземной растительности на территории Восточной Чукотки (турон)…” (Филиппова, 1997, с. 19). Таксономический состав растений амгеньского комплекса позволяет авторам настоящей статьи согласиться с выводом Филипповой. Этот “следующий этап” в пределах ОЧВП мы называем арманским и датируем туроном–коньяком (Herman et al., 2016). В это время на Северо-Востоке России существовали арманская, чинганджинская, пенжинская, кайваямская и ряд других флор (Герман, 2011; Щепетов, Герман, 2019).
ИЗОТОПНОЕ ДАТИРОВАНИЕ: МЕТОДИКА И РЕЗУЛЬТАТЫ
Изотопный U–Pb возраст определен для цирконов, извлеченных из двух образцов пород нырвакинотской толщи (обр. 09-308/1 – вулканомиктовый конгломерат, непосредственно подстилающий толщу андезитовых лав в верховьях р. Матачинай, и обр. T06-22 – дацитовый кристалловитрокластический туф из горизонта, залегающего между двумя потоками андезитовых лав в левом борту р. Правый Тадлеоан). Положение точек отбора проб показано на рис. 4. В шлифе обр. T06-22 отчетливо видны недеформированные частицы вулканического пепла, что практически исключает возможность субвулканической природы данного геологического тела. Цирконы из обоих образцов извлечены в соответствии со стандартной методикой (с использованием тяжелых жидкостей) и помещены в шашки из эпоксидной смолы.
Цирконы из обр. 09-308/1 изучены в Отделении планетологии и наук о Земле Университета Хиросимы (Япония) с использованием метода лазерной абляции и масс-спектрометрии (LA-ICPMS). Процедура анализа описана в работе (Katsube et al., 2012). Изучаемое вещество испаряли твердотельным Nd-YAG лазером с длиной волны 213 нм и анализировали в масс-спектрометре Agilent 7500. Диаметр области анализа составлял около 25 мкм. Результаты обрабатывали с помощью программ Pepi-AGE (Dunkl et al., 2008) и Isoplot/Ex 3.23 (Ludwig, 2003).
Цирконы из образца T06-22 анализировали на установке SIMS 1270 в Лаборатории PML Научного центра IPM Университета Окаяма (Япония). Процедура анализа и полученные результаты описаны в работе (Tikhomirov et al., 2012).
В обр. 09-308/1 (туфоконгломерат) проанализированы 74 индивидуальных зерна циркона. Полный комплект аналитических данных доступен по адресу: https://www.researchgate.net/publication/… Большая часть полученных изотопных отношений близки к конкордантным (рис. 5а). Вариации торий-уранового отношения (от 0.15 до 1.0, в единичных случаях до 2.8) не дают оснований для выделения каких-либо генетических групп цирконов. Диапазон полученных значений U–Pb возраста составляет от 3050 до 74 млн лет при относительной погрешности (1σ) около 5%. Цирконы с докембрийскими возрастами немногочисленны (7 зерен в интервале 3050–2550 млн лет, 10 зерен в интервале 2120–1710 млн лет, 3 зерна в интервале 770–680 млн лет; см. рис. 5б). Из фанерозойских цирконов наиболее многочисленная группа (38 зерен из 74) датирована интервалом 118–74 млн лет. Единственный отчетливый максимум кривой относительной плотности вероятности изотопного возраста (рис. 5б, 5в) соответствует значению 92.5 млн лет, то есть приходится на ранний турон (Ogg et al., 2008).
Рис. 5.
(а) Диаграмма Тера-Вассербурга для цирконов из обр. 09-308/1 (длина отрезков соответствует величине 2σ); (б) и (в) – гистограмма распределения значений U–Pb возраста цирконов и кривая относительной плотности вероятности распределения изотопных дат (б – для всего интервала полученных значений возраста, в – для цирконов фанерозойского возраста).

В обр. T06-22 (туф дацита) проанализированы семь кристаллов циркона, и для всех получены конкордантные возрасты с хорошей сходимостью (рис. 6). U–Pb возраст образца составил 88.1 ± 1.2 млн лет (2σ; СКВО = 0.87), что соответствует коньякскому веку (Ogg et al., 2008).

ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Можно констатировать, что на Восточной Чукотке в пределах одного стратона – ныне упраздненной (Решения…, 2009) этелькуюмской свиты – оказались проявлены два этапа развития меловой флоры Северо-Востока Азии: буор-кемюсский и арманский. При этом признаков сколько-нибудь значительного перерыва между породами, вмещающими остатки растений этих этапов, не наблюдается. По имеющимся ныне данным, буор-кемюсский этап датируется ранним–средним альбом (Решения…, 2009), арманский – туроном–коньяком (Герман, 2011; Herman et al., 2016; Щепетов, Герман, 2019). Будет логичным признать, что одна из этих датировок, скорее всего, неверна и нуждается в изменении.
Мог ли арманский этап начаться раньше, чем мы полагаем? Арманская флора одновозрастна с чинганджинской флорой Северного Приохотья (Решения…, 2009; Herman et al., 2016). Возраст этих двух флор ОЧВП устанавливается посредством их корреляции с ископаемыми флорами Анадырско-Корякского субрегиона, в которых континентальные флороносные отложения переслаиваются с морскими, содержащими остатки стратиграфически важных моллюсков и фораминифер. Арманская и чинганджинская флоры достаточно уверенно сопоставляются с фаунистически надежно датированными позднетуронской пенжинской и коньякской кайваямской флорами Анадырско-Корякского субрегиона (Герман, 2011). Пенжинской флоре предшествовала также надежно датированная различными методами гребенкинская флора, существовавшая с конца альба до начала турона (Щепетов и др., 1992; Spicer et al., 2002). И арманская, и чинганджинская флоры по систематическому составу явно моложе гребенкинской флоры (Герман, 2011), так что “удревнить” их возраст довольно трудно.
Для буор-кемюсской флоры надежно определена лишь нижняя возрастная граница: в бассейнах рек Бол. Анюй и Еропол флороносные слои айнахкургенской и умкувеемской свит согласно залегают на морских отложениях с остатками моллюсков позднего апта (Решения…, 1978, 2009). Иначе обстоит дело с верхней возрастной границей: “Возраст буор-кемюсской и топтанской стратофлор несомненно лежит еще в пределах раннего мела и условно рассматривается мной как альбский, по-видимому исключая поздний альб…” (Самылина, 1974, с. 25). За четыре десятилетия палеофлористических и стратиграфических исследований на Северо-Востоке Азии эта датировка ничем не была подтверждена, но не нашлось и повода в ней усомниться. В результате предположение В.А. Самылиной превратилось в аксиому и было отражено в решениях двух Межведомственных региональных стратиграфических совещаний (Решения…, 1978, 2009).
Результаты изотопного датирования цирконов, выделенных из пород нырвакинотской толщи, указывают на то, что данный стратон начал формироваться не раньше турона (93 млн лет). Для семи зерен из обр. 09-308/1 значения U–Pb возраста оказались моложе, чем у цирконов из обр. T06-22 (предположительно, отобранного выше по разрезу). Это можно объяснить тем, что туфоконгломераты, представленные в обр. 09-308/1, приурочены к депрессии палеорельефа и поэтому могут быть моложе вулканитов, обнажающихся на более высоких гипсометрических отметках. Кроме того, нельзя исключить частичную потерю радиогенного Pb некоторыми кристаллами циркона в результате наложенных процессов.
Учитывая изложенное выше, наиболее вероятным нам представляется, что буор-кемюсская флора (по крайней мере в пределах Восточно-Чукотского сегмента ОЧВП) существовала в альбе, сеномане, туроне и, возможно, начале коньяка: состав флористических комплексов, стратиграфические соотношения флороносных отложений в районе залива Креста и результаты U–Pb датирования цирконов свидетельствуют в пользу такого предположения (рис. 7). Именно в таких широких пределах – от альбского века до по крайней мере турона или даже начала коньякского века – мы датируем нырвакинотский флористический комплекс, таксономически идентичный “классической” буор-кемюсской флоре Северо-Востока России.
Рис. 7.
Соотношение специальных (биостратиграфических – слоев с флорой) и основных (горизонтов) стратиграфических подразделений средней части (апт–сеноман) неморского мела Верхояно-Охотско-Чукотского региона Северо-Востока России (а) по работе (Щепетов, Герман, 2019), (б) по представлениям авторов настоящей статьи (область изменений показана серым цветом).

ЗАКЛЮЧЕНИЕ
Покрытосеменные впервые появились в начале мелового периода в средних и низких широтах Азии и Северной Америки и достигли арктических регионов этих континентов в альбское время (Samylina, 1968; Smiley, 1969a, 1969b; Retallack, Dilcher, 1986; Herman, 2002, 2013; http://arcticfossils.nsii.org.cn/). В раннем и среднем альбе покрытосеменные были еще очень редки и, вероятно, населяли преимущественно нарушенные местообитания по берегам рек и морских бассейнов, что могло быть связано с хорошей защищенностью семезачатков, укороченным репродуктивным циклом и со склонностью к неотении и неспециализированным методам опыления и распространения семян (Stebbins, 1974) – с адаптациями, характерными для ранних покрытосеменных (Retallack, Dilcher, 1981). В течение позднего альба–позднего мела на Северо-Востоке Азии имело место постепенное замещение “мезофитных” растительных сообществ “кайнофитными” и сосуществование тех и других типов растительности во внутриматериковых районах Северной Азии.
Ранние покрытосеменные, однако, существовали не сами по себе, изолированно, а образовывали вместе с некоторыми папоротниками и хвойными продвинутые “кайнофитные” растительные сообщества. В течение позднего альба и позднего мела такие богатые покрытосеменными сообщества, расселившись сначала на приморских аллювиальных низменностях и равнинах Анадырско-Корякского субрегиона в восточной оконечности Азии, постепенно проникали по речным долинам и иным нарушенным (например, вулканической деятельностью в ОЧВП) местообитаниям вглубь материка, где они со временем вытесняли существовавшую ранее “мезофитную” папоротнико-голосеменную растительность, какое-то время, однако, сосуществуя с ней на ограниченной территории.
Большинство ископаемых флор из меловых образований ОЧВП по систематическому составу существенно отличается от одновозрастных флор приморских низменностей. Вполне вероятно, что растительный покров этой обширной вулканической области восстанавливался после массовых извержений в основном за счет пула местных видов, а отсутствие конкуренции со стороны покрытосеменных способствовало длительному сохранению в составе таких флор древних групп растений и появлению на их основе некоторых новых таксонов. Быстрое расселение кайнофитных растительных сообществ началось здесь лишь во второй половине турона и коньяке, однако и в позднем мелу в ряде районов ОЧВП развитие флоры шло более или менее независимо от такового прибрежных районов.
Приведенные выше данные свидетельствуют о том, что нырвакинотский флористический комплекс Восточной Чукотки, таксономически не отличающийся от типично “мезофитной” региональной буор-кемюсской флоры Северо-Востока Азии альбского возраста, в действительности существовал не только в альбское время, но и в позднем мелу, возможно вплоть до коньякского века. В это время растительность как приморских низменностей Анадырско-Корякского субрегиона, так и многих районов ОЧВП была уже типично “кайнофитной”, с доминированием покрытосеменных и сопутствующих им продвинутых хвойных. Вероятно, территория Восточной Чукотки позже других районов Северо-Востока Азии подверглась инвазии таких “кайнофитных” растительных сообществ, вследствие чего “мезофитная” папоротнико-голосеменная нырвакинотская флора существовала здесь дольше обычного в позднемеловую эпоху.
Благодарности. Авторы признательны рецензентам Н.К. Лебедевой (ИНГГ СО РАН) и А.М. Никишину (МГУ им. М.В. Ломоносова), чьи комментарии помогли улучшить статью.
Источники финансирования. Данная работа выполнена в рамках темы госзадания Ботанического института РАН “Ископаемые растения России и сопредельных территорий: систематика, филогения, палеофлористика и палеофитогеография” (№ АААА-А19-119021190031-8) (С.В. Щепетов), а также тем госзаданий Геологического института РАН (№ 0135-2019-0044 (А.Б. Герман) и № 0135-2016-0022 (А.В. Моисеев, С.Д. Соколов)). Работа поддержана грантом РФФИ № 19-05-00121 (А.Б. Герман, С.В. Щепетов), грантом РНФ №19-47-04110 (П.Л. Тихомиров, анализ изотопных данных) и программой НИР СВКНИИ ДВО РАН (№ 0288-2017-0002) (П.Л. Тихомиров). U–Pb датирование обр. 09-308/1 выполнено за счет средств гранта РФФИ мол_а_вед №18-35-20037.
Список литературы
Акинин В.В., Миллер Э.Л. Эволюция известково-щелочных магм Охотско-Чукотского вулканогенного пояса // Петрология. 2011. Т. 19. № 2. С. 1–42.
Белый В.Ф. Стратиграфия и структуры Охотско-Чукотского вулканогенного пояса. М.: Наука, 1977. 171 с.
Белый В.Ф., Ефимова А.Ф., Паракецов К.В. Нижний мел северо-восточной части Охотско-Чукотского вулканического пояса // Советская геология. 1965. № 10. С. 97–109.
Герман А.Б. Этапность и цикличность развития позднемеловой флоры Анадырско-Корякского субрегиона (Северо-Восток России) и их связь с климатическими изменениями // Стратиграфия. Геол. корреляция. 1993. Т. 1. № 1. С. 87–96.
Герман А.Б. Альбская–палеоценовая флора Северной Пацифики. М.: ГЕОС, 2011. 280 с. (Тр. ГИН РАН. Вып. 592).
Головнева Л.Б. Провинциальное деление азиатской части Сибирско-Канадской палеофлористической области в позднем мелу // Стратиграфия. Геол. корреляция. 2014. Т. 22. № 3. С. 64–74.
Государственная геологическая карта СССР масштаба 1 : 200 000. Серия Чукотская. Лист Q-1-XI, XII. Объяснительная записка. Составитель Петренко Е.Е. Москва: Союзгеолфонд, 1982. 85 с.
Государственная геологическая карта СССР масштаба 1 : 200 000. Серия Чукотская. Лист Q-1-XV, XVI. Объяснительная записка. Составитель Богомолов Г.И. Москва: Союзгеолфонд, 1983. 63 с.
Лебедев Е.Л. Стратиграфия и возраст Охотско-Чукотского вулканогенного пояса. М.: Наука, 1987. 175 с. (Тр. Геол. ин-та АН СССР. Вып. 421).
Моисеева М. Г., Соколова А.Б. Новые данные о составе и возрасте усть-эмунэрэтской флоры бассейна р. Энмываам (Центральная Чукотка) // Стратиграфия. Геол. корреляция. 2014. Т. 22. № 3. С. 45–63.
Решения 2-го Межведомственного регионального стратиграфического совещания по докембрию и фанерозою Северо-Востока СССР (Магадан, 1974–1975 гг.). Магадан: ГКП СВТГУ, 1978. 192 с.
Решения Третьего Межведомственного регионального стратиграфического совещания по докембрию, палеозою и мезозою Северо-Востока России (Санкт-Петербург, 2002). СПб.: Изд-во ВСЕГЕИ, 2009. 268 с.
Самылина В.А. Мезозойская флора левобережья р. Колымы (Зырянский угленосный бассейн). Часть I. Хвощевые, папоротники, цикадовые, беннеттитовые // Тр. Ботан. ин-та АН СССР. Сер. VIII. 1964. Вып. V. С. 40–79.
Самылина В.А. Мезозойская флора левобережья р. Колымы (Зырянский угленосный бассейн). Часть II. Гинкговые, хвойные. Общие главы // Тр. Ботан. ин-та АН СССР. Сер. VIII. 1967. Вып. VI. С. 133–176.
Самылина В.А. Раннемеловые флоры Северо-Востока СССР (к проблеме становления флоры кайнофита) // XXVII Комаровские чтения. Л.: Наука, 1974. 55 с.
Самылина В.А. Меловая флора Омсукчана. Л.: Наука, 1976. 207 с.
Филиппова Г.Г. Стратиграфия и возраст континентальных отложений бассейна реки Амгуэма и северного побережья залива Креста // Колыма. 1997. № 2. С. 12–23.
Филиппова Г.Г. Стратиграфия и возраст континентальных отложений Центральной и Восточной Чукотки // Тихоокеанская геология. 2001. Т. 20. № 1. С. 85–99.
Щепетов С.В. Стратиграфия континентального мела Северо-Востока России. Магадан: СВКНИИ ДВО РАН, 1995. 122 с.
Щепетов С.В., Герман А.Б. К вопросу о стратиграфии и флорах неморского мела Северо-Востока России // Стратиграфия. Геол. корреляция. 2019. Т. 27. № 3. С. 40–52.
Щепетов С.В., Нешатаева В.Ю. К проблеме корреляции толщ неморского мела Северо-Востока России: условия формирования флор вулканической области // Стратиграфия. Геол. корреляция. 2019. Т. 27. № 6. С. 41–54.
Щепетов С.В., Герман А.Б., Белая Б.В. Среднемеловая флора правобережья реки Анадырь (стратиграфическое положение, систематический состав, атлас ископаемых растений). Магадан: СВКНИИ ДВО АН СССР, 1992. 166 с.
Dunkl I., Mikes T., Simon K.E.A., Von Eynatten H. Brief introduction to the Windows program Pepita: data visualization, and reduction, outlier rejection, calculation of trace element ratios and concentration from LA-ICP-MS data // Laser Ablation ICP-MS in the earth sciences: current practices and outstanding issues. Ed. Sylvester P. Mineral. Asso-c. Canada Short Course. 2008. V. 40. P. 334–340.
Herman A.B. Late Early–Late Cretaceous floras of the North Pacific Region: florogenesis and early angiosperm invasion // Rev. Palaeobot. Palynol. 2002. V. 122. № 1–2. P. 1–11.
Herman A.B. Albian–Paleocene Flora of the North Pacific: systematic composition, palaeofloristics and phytostratigraphy // Stratigr. Geol. Correlation. 2013. V. 21. № 7. P. 689–747.
Herman A.B., Golovneva L.B., Shczepetov S.V., Grabovsky A.A. The Late Cretaceous Arman Flora of Magadan Oblast, Northeastern Russia // Stratigr. Geol. Correlation. 2016. V. 24. № 7. P. 651–760. https://doi.org/10.1134/S0869593816070029
Hourigan J.K., Akinin V.V. Tectonic and chronostratigraphic implications of new 40Ar/39Ar geochronology and geochemistry of the Arman and Maltan-Ola volcanic fields, Okhotsk-Chukotka volcanic belt, northeastern Russia // GSA Bull. 2004. V. 116. № 5/6. P. 637–654.
Katsube A., Hayasaka Y., Sakaguchi A., Takahashi Y. U–Pb zircon dating using Nd-YAG (213 nm) Laser ablation-ICP-MS, and evaluating the consistency with SHRIMP dating // J. Geol. Soc. Japan. 2012. V. 118(11). P. 762–767.
Kelley S.P., Spicer R.A., Herman A.B. New 40Ar/39Ar dates for Cretaceous Chauna Group tephra, North-Eastern Russia, and their implications for the geologic history and floral evolution of the North Pacific Region // Cretaceous Res. 1999. V. 20. № 1. P. 97–106.
Ludwig K.R. User’s Manual for Isoplot/Ex, Version 3.00. A geochronological toolkit for Microsoft Excel // Berkeley Geochronology Center Spec. Publ. 2003. V. 4. P. 25–32.
Ogg J.G., Ogg G., Gradstein F.M. The Concise Geologic Time Scale. Cambridge University Press, 2008. 150 p.
Retallack G.J., Dilcher D.L. A coastal hypothesis for the origin and rise to dominance of flowering plants // Palaeobotany, Palaeoecology and Evolution. V. 2. Ed. Niklas K.J. New York: Praeger Publ., 1981. P. 27–77.
Retallack G.J., Dilcher D.L. Cretaceous angiosperm invasion of North America // Cretaceous Res. 1986. V. 7. P. 227–252.
Samylina V.A. Early Cretaceous angiosperms of the Soviet Union based on leaf and fruit remains // J. Linnaean Soc. (Botany). 1968. V. 61. № 384. P. 207–218.
Smiley Ch.J. Cretaceous floras of Chandler-Colville region, Alaska: stratigraphy and preliminary floristics // Am. Assoc. Petrol. Geol. Bull. 1969a. V. 53. № 3. P. 482–502.
Smiley Ch.J. Floral zones and correlations of Cretaceous Kukpowruk and Corwin Formations, Northwestern Alaska // Am. Assoc. Petrol. Geol. Bull. 1969b. V. 53. № 10. P. 2079–2093.
Spicer R.A., Ahlberg A., Herman A.B., Kelley S.P., Raikevich M., Rees P.M. Palaeoenvironment and ecology of the middle Cretaceous Grebenka flora of northeastern Asia // Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002. V. 184. № 1–2. P. 65–105.
Stebbins G.L. Flowering plants: evolution above the species level. Cambridge, Massachusetts: Belknap Press of Harvard University Press, 1974. 399 p.
Tikhomirov P.L., Kalinina E.A., Moriguti T., Makishima A., Kobayashi K., Cherepanova I.Yu., Nakamura E. The Cretaceous Okhotsk-Chukotka Volcanic Belt (NE Russia): geology, geochronology, magma output rates, and implications on the genesis of silicic LIPs // J. Volcanol. Geothermal Res. 2012. V. 221–222. P. 14–32.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция