

Стратиграфия. Геологическая корреляция, 2020, T. 28, № 4, стр. 26-42
Микробиота баракунской свиты и биостратиграфическая характеристика дальнетайгинской серии: ранний венд Уринского поднятия Сибири
Н. Г. Воробьева 1, *, П. Ю. Петров 1
1 Геологический институт РАН
Москва, Россия
* E-mail: natali-geo@rambler.ru
Поступила в редакцию 13.06.2019
После доработки 21.08.2019
Принята к публикации 20.11.2019
Аннотация
В средней части мощной (2.2–2.7 км) глинисто-карбонатной толщи постледниковых отложений дальнетайгинской серии Уринского поднятия обнаружена богатая ассоциация раннеэдиакарской (ранневендской) акантоморфной палинофлоры (ЕСАР). Степень сохранности микрофоссилий определялась мгновенной скоростью их захоронения в пределах узких фациальных зон глубоководного бассейна, располагавшихся у подножий крупных проградировавших карбонатных тел. Сравнительный анализ микроостатков из баракунской свиты с известной ранее уринской ассоциацией микрофоссилий свидетельствует о принадлежности этих ассоциаций к единой комплексной палинозоне в пределах дальнетайгинской серии. Сравнительный анализ дальнетайгинской ассоциации с эдиакарскими палинофлорами других континентов показывает высокий уровень диверсификации микроорганизмов, характерный для верхних комплексных палинозон нижнего эдиакария Австралии и Китая, датированных интервалом 580–565 млн лет. Таким образом, верхняя большая часть дальнетайгинской серии имеет существенно более молодой возраст, чем это предполагалось ранее (<635 млн лет). Заметное сокращение стратиграфического объема в большей части постледниковых отложений дальнетайгинской серии предполагает наличие ощутимых стратиграфических перерывов в нижней части этой последовательности или пересмотр возраста большепатомского гляциогоризонта.
ВВЕДЕНИЕ
Разрез венда Уринского поднятия Сибири в последние годы играет заметную роль в построении глобальной биостратиграфической шкалы раннего эдиакария на микропалеонтологической основе (Чумаков и др., 2013; Воробьева, Сергеев, 2018). Однако до настоящего времени источником палеонтологической информации являлось единственное естественное обнажение, приуроченное к верхней части уринской свиты дальнетайгинской серии, а все известные региональные корреляции были выполнены по кернам буровых скважин (Голубкова и др., 2010; Кочнев и др., 2018). В этой связи получение микропалеонтологической информации по другим горизонтам дальнетайгинской серии и анализ этой информации на основе данных бассейновой (секвентной) стратиграфии представляется первоочередной задачей.
Дальнетайгинская серия мощностью более 3 км сформировалась в пределах Патомского бассейна, который представлял собой обширный залив Палеосибирского океана, возникшего после распада суперконтинента Родиния в конце неопротерозоя (Kuzmichev et al., 2001; Хераскова и др., 2010; Метелкин и др., 2012). В последовательности отложений этого бассейна терригенно-карбонатная толща дальнетайгинской серии залегает несогласно на породах терригенной баллаганахской серии и с размывом перекрывается преимущественно карбонатными отложениями жуинской серии (Чумаков и др., 2007, 2013). Вендский возраст этих отложений, впервые реконструированный на основе палеогеографических построений (Sovetov, 2002), позже был уточнен изотопными датировками, данными био- и хемостратиграфических исследований и палеотектонических построений (Покровский и др., 2006а, 2006б; Покровский, Буякайте, 2015; Melezhik et al., 2009; Голубкова и др., 2010; Чумаков и др., 2011; Sergeev et al., 2011; Moczydłowska, Nagovitsin, 2012; Powerman et al., 2015; Воробьева, Сергеев, 2018). По современным представлениям дальнетайгинская серия была сформирована в раннем венде в интервале 635–560 млн лет (Чумаков и др., 2013). Такой вывод основан на корреляции диамиктитов большепатомской свиты с глобальным оледенением Марино (Marinoan) в нижней части разреза и присутствием отрицательной аномалии δ13С Шурам-Вонока (EN-3) в перекрывающих известняках жуинской серии. Внушительная мощность отложений и отсутствие признаков длительного перерыва в основании жуинской серии (Чумаков и др., 2007) позволили рассматривать стратиграфический объем дальнетайгинской серии в границах всего нижнего венда (нижнего эдиакария) (Чумаков и др., 2013; Кочнев и др., 2018) (рис. 1). По современным оценкам этот интервал геологической истории насчитывает не менее 70 млн лет. Однако фациальная однородность и стратиграфическая непрерывность всей осадочной последовательности постледниковых отложений дальнетайгинской серии свидетельствуют о значительно более коротком временном интервале ее накопления (Рудько и др., 2017; Петров, 2018а, 2018б). Цель настоящей статьи – изучение новых местонахождений эдиакарской палинофлоры (ЕСАР) в отложениях баракунской и уринской свит на Уринском поднятии и анализ стратиграфического положения дальнетайгинской серии в границах нижнего венда.
Рис. 1.
(а) Географическое положение и геологическое строение района, (б) современная стратиграфическая схема отложений венда по (Чумаков и др., 2013), (в) секвентная стратиграфия по (Петров, 2018б) и положение органостенных микробиот в разрезе изученной части дальнетайгинского бассейна. 1 – диамиктиты; 2 – песчаники; 3 – аргиллиты; 4 – известковистые аргиллиты, мергели и глинистые известняки; 5 – известняки и доломиты; 6 – олистостромы, крупные тела известняковой брекчии; 7 – горизонт венчающих доломитов (cap dolomites); 8 – изохроны; 9 – местонахождения акантоморфной палинофлоры баракунской (1*, 2*) и уринской (3*, 4*) свит; 10 – габбро-долериты (силл). Серии: Dl – дальнетайгинская, Zh – жуинская, Tr – трехверстная. Свиты: bp – большепатомская, br – баракунская, ur – уринская, kl – каланчевская. Секвентная стратиграфия: Sq – границы секвенций, mf – поверхности максимального подъема уровня моря, T – трансгрессивные границы, TS – трансгрессивный тракт, HS – тракт высокого положения уровня моря. ПБ – Патомский бассейн, УП – Уринское поднятие.
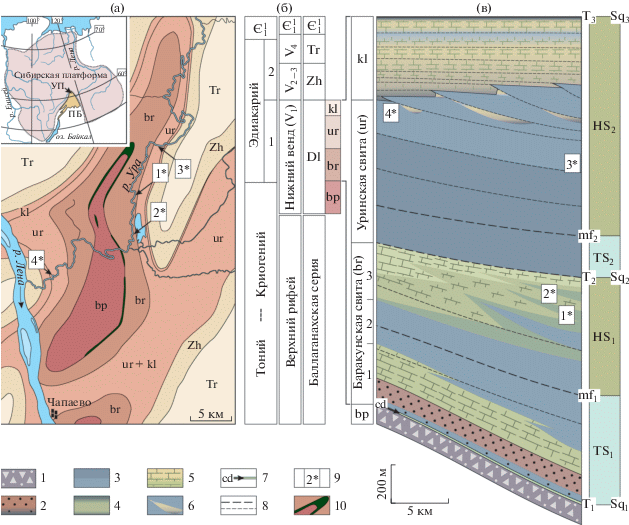
СЕКВЕНТНАЯ СТРАТИГРАФИЯ ДАЛЬНЕТАЙГИНСКОГО БАССЕЙНА И ФАЦИАЛЬНАЯ ХАРАКТЕРИСТИКА ПОСТЛЕДНИКОВЫХ ОТЛОЖЕНИЙ
Мощность большепатомского гляциогоризонта (большепатомская/джемкуканская свиты) достигает 1 км и более, что указывает на высокий темп прогибания ложа бассейна. Высокий уровень аккомодации сохранялся также и в течение постледниковой седиментации отложений дальнетайгинской серии. Шельфовая архитектура бассейна, мелководные обстановки сублиторали в большепатомское и в самое раннее баракунское время явились следствием компенсированного режима седиментации в условиях избытка осадков, мобилизованных ледниковой абразией на континенте. Минимальное положение уровня моря на границе большепатомской и баракунской свит, вероятно, было связано с кратковременным изостатическим подъемом области шельфа и прилегавших равнин, освободившихся от ледникового покрова. Последующее истощение потока поступавших в бассейн осадков привело к формированию глубоководных рампов баракунского и уринского времени, а замедление темпа прогибания на заключительной стадии – к проградации каланчевской карбонатной платформы (Петров, 2018а, 2018б). Быстрое и значительное по амплитуде падение уровня моря, приведшее к осушению всей территории бассейна, фиксируется в кровле дальнетайгинской серии. В такой последовательности событий был сформирован дальнетайгинский терригенно-карбонатный цикл.
В основании и в кровле дальнетайгинский цикл ограничен секвентными границами старшего порядка. Их природа различна. Происхождение нижней границы связано с эвстатическим падением уровня моря во время последнего ледникового максимума, а происхождение верхней границы – с перестройкой коллизионного процесса и подъемом территории. Внутри постледниковой последовательности отложений установлены две секвенции, разделенные согласной во внутренней части бассейна секвентной границей и представляющие собой два трансгрессивно-регрессивных цикла второго порядка (Петров, 2018б). Становление позднебаракунской карбонатной платформы в то время было прервано раннеуринской трансгрессией, которая, очевидно, также имела региональную тектоническую природу. Таким образом, осадочная летопись постледниковых отложений дальнетайгинской серии на территории современного Уринского поднятия была непрерывной.
Состав тонкозернистых силикокластических осадков дальнетайгинской серии предполагает значительную долю эоловой составляющей осадконакопления, которая стала наиболее ощутимой после раннеуринской трансгрессии, во время максимального удаления рассматриваемых фациальных зон палеобассейна от флювиальных источников сноса (Петров, 2018б). Эоловая пыль, несущая комплекс биологически важных элементов, обеспечила высокую биопродуктивность удаленных акваторий и повышенные скорости седиментации во внешних фациальных зонах палеобассейна. Массовое захоронение органического вещества, активизация метаногенных процессов, тиксотропное состояние осадков, насыщенных продуктами брожения, в условиях сейсмической активности бассейна способствовали почти непрерывному перемещению разжиженных осадочных масс вниз по склону. Очень малые (около 1°) углы наклона рампа исключили возможность формирования турбидитных потоков. Максимальные скорости оползневой седиментации наблюдались у подножия проградационных призм активно растущих карбонатных платформ, что, в частности, способствовало высокому тафономическому потенциалу ископаемых палинофлор. Кажется, деструкция оболочек микрофоссилий, и в первую очередь их тонких фрагментов, была чрезвычайно чувствительна даже к непродолжительной экспозиции в поверхностных слоях осадка. Таким образом, ассоциации микрофоссилий наилучшей сохранности как в уринской, так и в баракунской свите наблюдались непосредственно в основании проградационных фронтов крупных карбонатных тел (рис. 1). Даже на незначительном (первые километры) удалении от них органическая составляющая осадка, как правило, представляла собой ультрадисперсную высокоуглеродистую пыль.
МЕТОДИКА ИССЛЕДОВАНИЙ
Лабораторная обработка материала и изготовление препаратов проводились по щадящей методике выделения органических остатков из терригенных отложений и включали следующие этапы: (1) измельчение массивных фрагментов породы; (2) поверхностная очистка раздробленных фрагментов от загрязнений и удаление примеси обычно незначительного (менее 5%) карбонатного материала 20%-ным раствором соляной кислоты; (3) растворение очищенного материала в концентрированной плавиковой кислоте; (4) отделение заключающей микроостатки крупной фракции от глинистой взвеси на ситовой ткани (мельничный газ (GG) из полиэфирный нити) с диаметром ячеи 45 мкм; (5) ручной отбор органического материала из обогащенного осадка с помощью иглы под бинокулярным микроскопом МБС-9, что позволяет исключить центрифугирование органомацерата в тяжелой жидкости и дает возможность сохранить крупные фрагменты микроорганизмов неповрежденными; (6) изготовление препаратов на основе канадского бальзама с добавлением в качестве стабилизатора изоамилового эфира уксусной кислоты (C7H14O2). Микрофоссилии фотографировали в проходящем свете с помощью микроскопа Zeiss REM 5. Весь материал хранится в Геологическом институте РАН (Москва), палеонтологическая коллекция № 14713.
ХАРАКТЕРИСТИКА ИЗУЧЕННОГО МАТЕРИАЛА
В работе было проанализировано более 70 образцов, в 25 из них микрофоссилии выявлены не были. В мацерате остальных проб присутствует различное (иногда весьма значительное) количество сильно разложенного органического вещества. Большая часть проб содержит лишь единичные неопределимые микроостатки или их фрагменты, а общая сохранность микрофоссилий может быть оценена как плохая, реже как удовлетворительная. Микроостатки в основной массе подверглись существенной минеральной и/или бактериальной деструкции. Они имеют различные оттенки темно-коричневого (иногда до черного) цвета, приобретенного в результате температурных воздействий порядка 160–180°С и выше (шкала цветовых изменений докембрийских органостенных форм; Hаyes et al., 1983).
Наиболее обильные и разнообразные ассоциации микрофоссилий выявлены на двух практически идентичных по микрофитологическому составу уровнях, залегающих в 130 и 180 м ниже кровли баракунской свиты; их координаты 60.31040° с.ш., 117.28547° в.д. и 60.35165° с.ш., 117.29392° в.д. соответственно. Эти разрезы расположены в среднем течении р. Ура на расстоянии 4.1 км друг от друга (1* и 2* на рис. 1). Единичные остатки акантоморфит обнаружены также в средней части уринской свиты в 400–450 м от ее подошвы; координаты 60.40285° с.ш., 117.34530° в.д. (3* на рис. 1).
Все найденные микрофоссилии образуют единую, ограниченную по представительности, но показательную ассоциацию. О единстве этой ассоциации свидетельствует отсутствие каких-либо изменений таксономического разнообразия морфотипов, наблюдаемое по всему разрезу, а также сходство сохранности всех изученных микроостатков и повсеместное однотипное доминирование среднеразмерных акритарх. Таким образом, в пределах всего изученного интервала наблюдается отсутствие каких-либо эволюционно и экологически значимых трендов изменения обилия и таксономического разнообразия встреченных акантоморфит.
В состав баракунской ассоциации, помимо транзитных форм, представленных родами Leiosphaeridia, Siphonophycus, входят многочисленные Polythrichoides lineatus Hermann, 1974, Cephalophytarion laticellulosum Schoph et Blacic, 1971, Caudosphaera expansa Hermann et Timofeev, 1989, Obruchevella sp., единичные Palaeopleurocapsa wopfneri Knoll et al., 1975 и Palaeovolvox ooraminnus Zang et Walter, 1992, а также многочисленные микроорганизмы предположительно грибной природы (формы 3 и 4). Самыми значимыми являются находки акантоморфных акритарх, встреченные на двух упомянутых уровнях баракунской ассоциации, а также единичные экземпляры, обнаруженные в интервале между этими уровнями. Большая часть акантоморфит сохранилась настолько плохо, что невозможно провести их определение даже до рода. Пригодная для опознания часть выборки содержит микрофоссилии нескольких родов, характерных для нижнего венда. Самыми многочисленными являются представители видов Appendisphaera tenuis Moczydłowska et al., 1993, emend. Moczydłowska, 2005 и Asterocapsoides robustus Xiao et al., 2014 (последний был описан из уринской свиты как ?Sinosphaera rupina Zhang et al., 1998, emend. Liu et al., 2014 (Sergeev et al., 2011)). Ассоциация также включает Asseserium aff. piramidalis Nagovitsin et Moczydłowska, 2012, Knollisphaeridium maximum (Yin), emend. Willman et Moczydłowska, 2008, Multifronsphaeridium ramosum Nagovitsin et Moczydłowska, 2012, Tanarium conoideum Kolosova, 1991, emend. Moczydłowska et al., 1993, Cavaspina basiconica Moczydłowska et al., 1993, единичные Variomargosphaeridium sp., Tanarium pluriprotensum Grey, 2005, T. pycnacanthum Grey, 2005, Keltmia aff. cornifera Vorob’eva et al., 2009, а также неописанные Форму 1 и Форму 2. В интервале между упомянутыми уровнями обнаружены единичные эдиакарские акантоморфиты Appendisphaera tenuis, транзитные микрофоссилии Leiosphaeridia sp., Siphonophycus sp., а также неопределимые фрагменты орнаментированных и гладкостенных акритарх и нитчатых водорослей.
В целом баракунская микробиота является обедненным аналогом хорошо изученной ранее уринской ассоциации микрофоссилий (Голубкова и др., 2010; Sergeev et al., 2011; Moczydłowska, Nagovitsin, 2012; Воробьева, Сергеев, 2018) (4* на рис. 1). Отличия состоят в отсутствии в баракунском материале таких характерных для уринской свиты таксонов, как Ancorosphaeridium, Cavaspina, Eotylotopalla, и в наличии единичных Tanarium pluriprotensum, T. pycnacanthum. Кроме того, в баракунской ассоциации встречены Форма 1 и Форма 2, не обнаруженные в Lagerstätte терминального горизонта уринской свиты. Несколько найденных в баракунской свите экземпляров, схожих с Appendisphaera aff. tabifica и Tanarium aff. digitiformum, в силу своей плохой сохранности и возможности их неоднозначной трактовки, не были отнесены к этим таксонам. Вместе с тем нельзя исключить, что эти единичные остатки могут оказаться представителями перечисленных родов.
БИОСТРАТИГРАФИЧЕСКАЯ ХАРАКТЕРИСТИКА ДАЛЬНЕТАЙГИНСКОЙ СЕРИИ
Сравнительный анализ всех изученных ассоциаций микрофоссилий дальнетайгинской серии однозначно указывает на единство их таксономического состава. Здесь в обедненных выборках присутствуют лишь единичные таксоны наиболее распространенных форм, известных из богатых местонахождений, а самая представительная ассоциация баракунской свиты приближается к самой богатой и детально изученной ранее ассоциации терминального горизонта уринской свиты (рис. 2). Показательным является присутствие как в уринской, так и в баракунской ассоциации эндемичного таксона Asseserium aff. piramidalis. Обильно представленные в верхнеуринском местонахождении и не идентифицированные в нижележащих отложениях виды рода Ancorosphaeridium, по всей видимости, занимали очень узкие фациально-экологические зоны в бассейне. Вероятность сохранности фрагментов этих форм, перенесенных в сравнительно глубоководные зоны, была крайне незначительной. Отсутствие в баракунском материале характерного для ЕСАР сфероморфного космополита Schizofusa zangwenlongii Grey, 2005 может быть объяснено узкой избирательностью распространения этого таксона в разрезе. Принципиальным отличием баракунской ассоциации от уринской являются единичные находки в первой двух очень редких видов Tanarium pluriprotensum и T. pycnacanthum, пока не обнаруженных в уринской ассоциации. Таким образом, почти половина осадочной последовательности постледниковых отложений дальнетайгинской серии представляет единую комплексную зону акантоморфной палинофлоры нижнего венда (рис. 2).
Рис. 2.
Таксономическое разнообразие акантоморфных биот дальнетайгинской серии Уринского поднятия в сравнении с эдиакарским комплексом акантоморфной палинофлоры (ЕСАР) Австралии и Китая. 1–3 – количество найденных экземпляров: 1 – единичные находки, 2 – много (десятки-сотни экземпляров), 3 – очень много (сотни-тысячи экземпляров); 4 – гляциогоризонты (диамиктиты); 5 – изотопный возраст (млн лет). Lj/Lc – Leiosphaeridia jacutica–Leiosphaeridia crassa Assemblage Zone (по Grey, 2005); ELP – первая палинозона эдиакария (по Grey, 205). 1'–4' – новые палинозоны формации Доушаньтуо Ю. Китая (по Liu, Moczydłowska, 2019): 1' – Appendisphaera grandis–Weissiella grandistella–Tianzhushania spinosa, 2' – Tanarium tuberosum–Schizofusa zangwenlongii, 3' – Tanarium conoideum–Cavaspina basiconica, 4' – Tanarium pycnacanthum–Ceratosphaeridium glaberosum. Zh – жуинская серия. Свиты: bp – большепатомская, br – баракунская, ur – уринская, kl – каланчевская. EN-1, EN-2, EN-3, EN* – отрицательные аномалии δ13С эдиакария.

Сравнительный анализ дальнетайгинской ассоциации с наиболее изученными палинофлорами Австралии (Zang, Walter, 1992; Grey, 2005; Willman, Moczydłowska, 2008) и Южного Китая (Liu et al., 2014; Liu, Moczydłowska, 2019; Xiao et al., 2014; O-uyang et al., 2017, 2019) показывает высокий уровень диверсификации микроорганизмов, свойственный верхним горизонтам нижнего эдиакария, моложе 580 млн лет. Характерными представителями этой ассоциации являются Asterocapsoides robustus, Hocosphaeridium scaberfacium Zang, 1992, emend. Liu et al., 2014, Knollisphaeridium maximum, Tanarium conoideum, а также диноцистоподобные формы рода Variomargosphaeridium. Недавние находки в верхних горизонтах формации Доушаньтуо Южного Китая представителей рода Urasphaera (U. fungiformis и U. nupta) (Liu et al., 2014) и Ancorosphaeridium magnum (Liu, Moczydłowska, 2019), известных из уринской ассоциации, подтверждают такой вывод. Единичные экземпляры видов Tanarium pluriprotensum и T. pycnacanthum, обнаруженные в баракунской свите и известные в ЕСАР Китая (Liu et al., 2014; Liu, Moczydłowska, 2019) и Австралии (Grey, 2005; Willman, Moczydłowska, 2008), также указывают на принадлежность изученных горизонтов дальнетайгинской серии к верхним горизонтам нижнего эдиакария. В какой-то мере этот вывод подтверждают обнаруженные в уринской ассоциации микрофоссилии, близкие к роду Ceratosphaeridium (Sergeev et al., 2011; Moczydlowska, Nagovitsin, 2012), представители которого являются видом-индексом самой верхней зоны ЕСАР Австралии (Grey, 2005) и Китая (Liu et al., 2012). Помимо того, зона Tanarium pycnacanthum–Ceratosphaeridium glaberosum является последней комплексной зоной формации Доушаньтуо, датированной интервалом 580–565 млн лет, в современной биостратиграфической схеме нижнего эдиакария Южного Китая (Liu, Moczydłowska, 2019).
Таким образом, принимая существующую стратратиграфическую схему Патомского комплекса, основанную на трансрегиональных корреляциях и глобальном тренде δ13C (Чумаков и др., 2013; Чумаков, 2015), следует заключить, что большая (1.5–1.8 км) верхняя часть постледниковой осадочной последовательности дальнетайгинской серии, включая предуджинский стратиграфический перерыв, формировалась не более 20 млн лет, тогда как меньшая по мощности (0.7–0.9 км) нижняя часть этой последовательности накапливалась на протяжении как минимум 55 млн лет. Однако высокий уровень аккомодации, компенсированный режим седиментации и отсутствие стратиграфически значимых перерывов на всем протяжении этой последовательности предполагают в разы меньший временной интервал ее формирования (Петров, 2018б). Следовательно, если большепатомский гляциогоризонт синхронен диамиктитам Наньтуо Южного Китая (635 млн лет), то постледниковая последовательность нижней части баракунской свиты все же заключает, вероятно, серию скрытых стратиграфических перерывов. В противном случае большепатомский гляциогоризонт имеет более молодой возраст, соответствующий следовавшему за диамиктитами Наньтуо гляциопериоду Гаскье (580 млн лет), как предполагалось ранее (Рудько и др., 2017; Петров, 2018а, 2018б). Примечательно, что положение нижней границы эдиакария в стратотипе Южной Австралии имеет такую же возрастную неопределенность (Grey, Calver, 2007).
ЗАКЛЮЧЕНИЕ
В отложениях верхней части баракунской свиты дальнетайгинской серии Уринского поднятия обнаружена представительная ассоциация органостенных микрофоссилий, характерная для раннеэдиакарской (нижневендской) акантоморфной палинофлоры (ЕСАР). В пределах палеобассейна наиболее богатые уровни с наилучшей сохранностью форм приурочены к фронтальным фациальным зонам проградации крупных карбонатных тел, у подножия которых суммарная скорость суспензионной и оползневой седиментации была максимальной. Таким образом, скорость захоронения микрофоссилий являлась решающим тафономическим фактором, определявшим степень сохранности ископаемых палинофлор раннего венда. Единство таксономического состава всех изученных ассоциаций дальнетайгинской серии свидетельствует об их принадлежности к одной комплексной зоне акантоморфной палинофлоры нижнего эдиакария (нижнего венда). Сравнительный анализ дальнетайгинской ассоциации с наиболее изученными эдиакарскими палинофлорами других континентов показывает высокий уровень диверсификации микроорганизмов, характерный для верхних комплексных палинозон Австралии и Китая, датированных интервалом 580–565 млн лет. Таким образом, по крайней мере большая верхняя (1.5–1.8 км) часть мощной толщи постледниковых отложений дальнетайгинской серии имеет существенно более молодой возраст, чем это предполагалось ранее. Учитывая высокие скорости седиментации дальнетайгинского бассейна, такая поправка предполагает наличие ощутимых стратиграфических перерывов в нижней части баракунской свиты либо пересмотр возраста большепатомского гляциогоризонта.
ПАЛЕОНТОЛОГИЧЕСКОЕ ОПИСАНИЕ
ГРУППА ACRITARCHA EWITT 1963
ПОДГРУППА ACANTHOMORPHITAE DOWNIE, EVITT ET SARJEANT, 1963
Род Appendisphaera Moczydłowska, Vidal et Rudavskaya, 1993, emend. Moczydłowska, 2005
Appendisphaera tenuis Moczydłowska, Vidal et Rudavskaya, 1993, emend. Moczydłowska, 2005
Табл. I , фиг. 3, 4
Таблица I . Микрофоссилии баракунской свиты. 1, 2 – Tanarium pluriprotensum Grey, 2005: 2 – увеличенный фрагмент, экз. ГИН № 14713-353, препарат 17-261-1, т. 5; 3, 4 – Appendisphaera tenuis Moczydłowska, Vidal et Rudavskaya, 1993, emend. Moczydłowska, 2005: 4 – увеличенный фрагмент, экз. ГИН № 14713-26, препарат 17-335-3, т. 2; 5–7 – Asterocapsoides robustus Xiao, Zhou, Liu, Wang et Yuan, 2014: 6, 7 – увеличенные фрагменты, экз. ГИН № 14713-231, преп. 17211-5, т. 10; 8, 9 – Densisphaera cf. arista Nagovitsin et Moczydłowska, 2012: 9 – увеличенный фрагмент, экз. ГИН № 14713-225, препарат 17211-5, т. 4; 10–12 – Tanarium pycnacanthum Grey, 2005: 11, 12 – увеличенные фрагменты, экз. ГИН № 14713-444, препарат 17264-4, т. 2; 13, 14 – Форма 1: 14 – увеличенный фрагмент, экз. ГИН № 14713-170, препарат 17210-7, т. 3; 15 – Tanarium conoideum Kolosova, 1991, emend. Moczydłowska, Vidal et Rudavskaya, 1993, экз. ГИН № 14713-2, препарат 17-323-1, т. 2; 16 – Форма 2, экз. ГИН № 14713-429, препарат 17263-9, т. 3. Одинарная масштабная линейка равна 20 мкм, двойная – 50 мкм. Номера образцов соответствуют первым пяти цифрам в номерах препаратов.

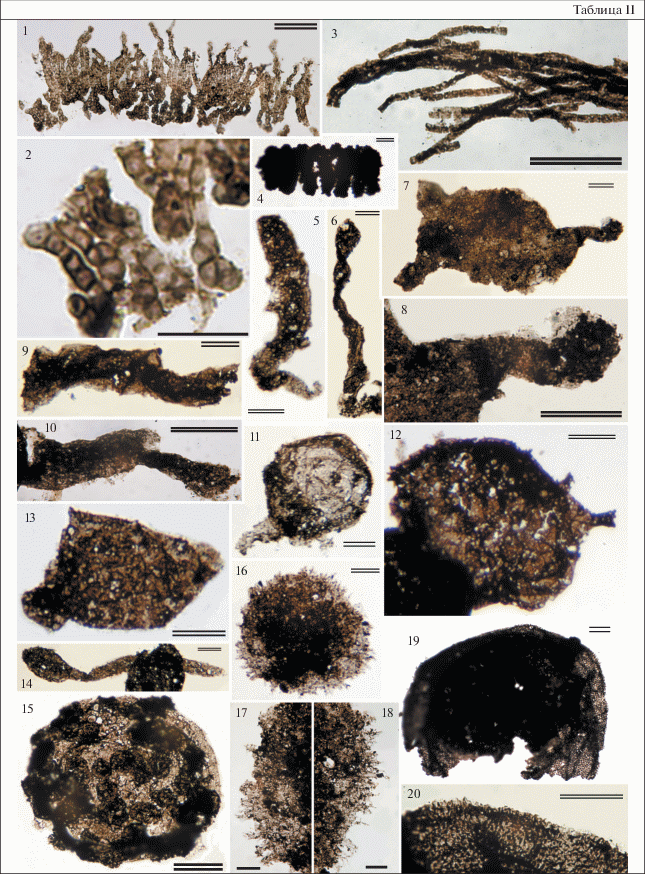
Таблица II. Микрофоссилии баракунской свиты. 1, 2 – Palaeopleurocapsa wopfnerii Knoll, Barghoorn et Golubic, 1975: 2 – увеличенный фрагмент, экз. ГИН № 14713-1, препарат 17-323-1, т. 1; 3 – Cehpalophytarion laticellulosum Schopf et Blacic, 1971, экз. ГИН № 14713-442, препарат 17265-3, т. 3; 4 – Obruchevella sp., экз. ГИН № 14713-293, препарат 17228-1, т. 5; 5, 6, 14 – Форма 3: 5 – экз. ГИН № 14713-380, препарат 17261-6, т. 5; 6 – экз. ГИН № 14713-116, препарат 17187-7, т. 1; 14 – экз. ГИН № 14713-324, препарат 17234-7, т. 2; 7, 8 – Keltmia aff. cornifera Vorob’eva, Sergeev et Knoll, 2009: 8 – увеличенный фрагмент, экз. ГИН № 14713-401, препарат 17263-3, т. 7; 9, 10 – Форма 4: 9 – экз. ГИН № 14713-249, препарат 17212-2, т. 5, 10 – экз. ГИН № 14713-320, преп. 17234-5, т. 1; 11 – Caudosphaera expansa Hermann et Timofeev, 1989, экз. ГИН № 14713-198, препарат 17211-1, т. 4; 12 – Variomargosphaeridium sp., экз. ГИН № 14713-3, препарат 17-324-1, т. 1; 13 – Asseserium aff. pyramidalis Nagovitsin et Moczydłowska, 2012, экз. ГИН № 14713-189, препарат 17210-20, т. 3; 15 – Palaeovolvox ooraminnus Zang et Walter, 1992, экз. ГИН № 14713-8, препарат 17-333-2, т. 1; 16–18 – Cavaspina basiconica Moczydlowska, Vidal et Rudavskaya, 1993: 17, 18 – увеличенные фрагменты, экз. ГИН № 14713-20, препарат 17-334-1, т. 5; 19, 20 – Knollisphaeridium maximum (Yin L., 1987), emend. Willman et Moczydłowska, 2008: 20 – увеличенный фрагмент, экз. ГИН № 14713-397, препарат 17263-3, т. 3. Одинарная масштабная линейка равна 20 мкм, двойная – 50 мкм. Номера образцов соответствуют первым пяти цифрам в номерах препаратов.
Appendisphaera tenuis: Moczydłowska et al., 1993, p. 506, text-fig. 7; Moczydłowska, 2005, p. 296, fig. 5 ; Grey, 2005, p. 224, fig. 113B ; Yin et al., 2007, fig. 1b; Willman, Moczydłowska, 2008, p. 520, figs. 7B, 7C ; Воробьева и др., 2008, фиг. 2k,l; Голубкова и др., 2010, табл. 1 , фиг. 2, табл. 3 , фиг. 5, 6 ; Sergeev et al., 2011, figs. 5.4 –6.; Xiao et al., 2014, pp. 9–10, fig. 3.4 .; Shukla, Tiwari, 2014, p. 215, figs. 4D–4E ; Ye et al., 2015, p. 48, pl. 1, figs. 9–14 ; Prasad, Asher, 2016, p. 44, pl. 3, figs. 3–6 , p. 42, pl. 2, figs. 1, 2, p. 52, pl. 6, figs. 3–6 , pp. 54, 56, pl. 7, figs. 6, 7 ; Anderson et al., 2019, pp. 8–9, figs. 6H–6I ; Liu, Moczydłowska, 2019, pp. 61–65, figs. 29, 30 .
Описание. Оболочки сфероидальной или эллипсоидальной формы, тонкие, полупрозрачные. Поверхность оболочки равномерно покрыта многочисленными выростами гомоморфными, неветвящимися, короткими, полыми, тонкими. Окончания выростов заостренные. Выросты сообщаются с полостью оболочки, четко отделены друг от друга в основании. Размер оболочек от 180 до 290 мкм, выросты длиной до 25 мкм, шириной менее 1 мкм.
Распространение. Паршинская свита непского горизонта, уринская и баракунская свиты дальнетайгинского горизонта Восточной Сибири; венд. Формации Pertatataka, Tanana, Karlaya Австралии, формация Doushantuo Южного Китая, формация Krol’A и группа Lower Vindhyan Индии, формация Khesen Северной Монголии; эдиакарий.
Материал. Тридцать восемь экземпляров из баракунской свиты.
Род Asseserium Nagovitsin et Moczydłowska, 2012
Asseserium aff. pyramidalis Nagovitsin et Moczydłowska, 2012
Табл. II, фиг. 13
Veryhachium sp.: Файзуллин, 1998, с. 334, табл. 2 , фиг. 7 .
Polygonium sp.: Наговицин и др., 2004, с. 13, 19, табл. II, фиг. 12–15 .
Gen. et sp. indet.: Голубкова и др., 2010, с. 20, табл. 4 , фиг. 4 .
Asseserium pyramidalis Nagovitsin et Moczydłowska: Moczydłowska, Nagovitsin, 2012, p. 13, fig. 5G .
Описание. Оболочки среднего размера (130–160 × 150–180 мкм), несущие небольшое количество (от 3 до 7) коротких выростов, расположенных полярно. Выросты полые, сообщающиеся с полостью оболочки, конические, имеют очень широкие основания и короткие узкие окончания. Ширина их основания от 35 до 60 мкм, высота от 10 до 35 мкм. Основания выростов не имеют четкой границы с поверхностью оболочки, что делает ее контур многоугольным.
Распространение. Уринская и баракунская свиты дальнетайгинского горизонта Восточной Сибири; венд.
Материал. Три экземпляра из баракунской свиты.
Род Asterocapsoides L. Yin et Li, 1978, emend. Xiao, Zhou, Liu, Wang et Yuan, 2014
Asterocapsoides robustus Xiao, Zhou, Liu, Wang et Yuan, 2014
Табл. I , фиг. 5, 6, 7
?Sinosphaera rupina: Zhang et al., 1998; Grey, 2005, p. 291, fig. 201 ; Willman et al., 2006, p. 32, pl. 2, fig. 4 ; Willman, Moczydłowska, 2008, p. 526, figs. 11D–11F ; Воробьева и др., 2008, фиг. 2i; Голубкова и др., 2010, табл. 4 , фиг. 5 ; Sergeev et al., 2011, p. 1005, figs. 7.2 –4.
Meghystrichosphaeridium chadianensis: Zhang et al., 1998, p. 34, figs. 3.6, 10.1–10.4 ; Воробьева и др., 2008, фиг. 2b.
Asterocapsoides robustus: Xiao et al., 2014, pp. 11–12, figs. 4.1 –6.
Описание. Сфероидальные или эллипсоидальные оболочки среднего размера, диаметром от 150 до 245 мкм, несущие многочисленные, равномерно распределенные по поверхности выросты. Выросты свободно сообщаются с полостью оболочки, полые, неветвящиеся, сужающиеся к острому замкнутому окончанию. Их основания сближены и почти соприкасаются. Ширина выростов в основании составляет от 11 до 21 мкм, длина от 14 до 25 мкм.
Замечание. Для рода Sinosphaera характерны два типа выростов: небольшие и плотно расположенные, перемежающиеся с крупными длинными выростами. Акантоморфные акритархи, описанные как ?Sinosphaera rupina из формаций Pertatataka, Dey Dey Mudstone и Tanana Австралии и из уринской свиты Восточной Сибири, имеют выросты одинакового размера и формы, что не соответствует диагнозу рода. На этом основании они были исключены из состава рода Sinosphaera и перемещены в Asterocapsoides robustus (Xiao et al., 2014).
Распространение. Формация Doushantuo Южного Китая, формации Pertatataka, Dey Dey Mudstone и Tanana Австралии; эдиакарий. Уринская и баракунская свиты дальнетайгинского горизонта Восточной Сибири; венд.
Материал. Девять экземпляров плохой и три экземпляра удовлетворительной сохранности из баракунской свиты.
Род Cavaspina Moczydłowska, Vidal et Rudavskaya, 1993
Cavaspina basiconica Moczydłowska, Vidal et Rudavskaya, 1993
Табл. II, фиг. 16–18
Baltisphaeridium (?) strigosum Jankauskas, 1976: Пятилетов, 1980, с. 11, табл. 1 , фиг. 5–8 , табл. 2 , фиг. 1–4 ; Пятилетов, Рудавская, 1985, с. 152, табл. 63 , фиг. 8 ; Рудавская, Васильева, 1989, табл. 1 , фиг. 2–4 , 6 , табл. 2 , фиг. 1, 2.
Tanarium perfectum (Kolosova): Колосова, 1991, figs. 6.1 –6.
Comasphaeridium sp. B: Zang, Walter, 1992, p. 34, figs. 28F, 28G .
Meghystrichosphaeridium perfectum (Kolosova, 1991): Zhang et al., 1998, p. 36, figs. 10.7 –8; Yuan et al., 2002, p. 78, fig. 108 ; Yin et al., 2007, pl. 11, fig. 5 .
Meghystrichosphaeridium chadianensis (Chen et Liu, 1986): Zhou et al., 2001, p. 1166, pl. 2, figs. 5, 6 .
Cavaspina basiconica sp. nov.: Moczydłowska et al., 1993, p. 510–512, text-fig. 11; Moczydłowska, 2005, p. 300, fig. 6C ; Willman et al., 2006, p. 25, pl. 1, figs. 3, 4 ; Willman, Moczydłowska, 2008, p. 522, figs. 9D–9F ; Willman, Moczydłowska, 2011, p. 25, pl. 2, figs. 1, 2; Xiao et al., 2014, pp. 16–18, figs. 8.1 –8.
Описание. Сфероидальные оболочки 160–200 мкм в диаметре, несущие многочисленные выросты. Выросты полые, 10–15 мкм в длину, сообщающиеся с полостью оболочки, конические, заостренные. Окончания выростов тупые или сужающиеся до тонкого волоска, основания выростов расширенные до 3–6 мкм. Выросты очень густо и равномерно покрывают всю поверхность оболочки.
Замечание. Данные экземпляры сходны с формами, отнесенными к Appendisphaera tenuis. Но так как выросты у этих экземпляров короче, толще и имеют более широкие основания, они описаны как Cavaspina basiconica.
Распространение. Паршинская свита непского горизонта, баракунская свита дальнетайгинского горизонта Восточной Сибири; венд. Формации Pertatataka, Tanana, Karlaya Австралии, формация Doushantuo Южного Китая; эдиакарий.
Материал. Четыре полных экземпляра и пять фрагментов плохой сохранности из баракунской свиты.
Род Densisphaera Nagovitsin et Moczydłowska, 2012
Densisphaera cf. arista Nagovitsin et Moczydłowska, 2012
Табл. I , фиг. 8, 9
Форма 3: Наговицин и др., 2004, с. 14, табл. 1 , фиг. 8, 9 .
Archaeotunisphaeridium aff. fimbriatum: Sergeev et al., 2011, p. 994, figs. 6.7 –9.
Densisphaera arista Nagovitsin et Moczydłowska: Moczydłowska, Nagovitsin, 2012, pp. 15, 16, figs. 6A–6F .
Описание. Оболочки среднего размера, имеющие неправильный контур из-за сильного смятия и частичного разрушения и несущие несколько выростов удовлетворительной сохранности. Выросты гетероморфные, полые, цилиндрической или конической формы. Выросты разделены на дистальные ветви второго и третьего порядка. Часть дистальных ветвей выростов покрыты остатками внешней полупрозрачной мембраны. Размеры оболочки 240 × 180 мкм, выросты длиной до 40 мкм, ширина основного выроста до 18 мкм, ширина вторичных ответвлений до 2 мкм.
Замечание. Внутреннее плотное тело, характерное для данного рода, отсутствует. Внешняя мембрана фрагментарно различима на дистальных частях выростов (табл. II, фиг. 9 ). Наличие этой мембраны, размер и форма выростов позволяют с большой долей вероятности отнести эти экземпляры к Densisphaera arista.
Распространение. Уринская и баракунская свиты дальнетайгинского горизонта Восточной Сибири; венд.
Материал. Два экземпляра плохой сохранности из баракунской свиты.
Род Knollisphaeridium Willman et Moczydłowska, 2008
Knollisphaeridium maximum (Yin L., 1987), emend. Willman et Moczydłowska, 2008
Табл. II, фиг. 19, 20
Baltisphaeridium maximum: Yin, 1987, pp. 439–440, pl. 14, figs. 14, 15 .
Baltisphaeridium maximum sp. nov.: Yin, 1987, pp. 439, 440, pl. 14, figs. 14, 15 .
Echinosphaeridium maximum (Yin, 1987): Knoll, 1992, p. 765, pl. 5, figs. 5, 6 ; Tiwari, Knoll, 1994, p. 198, pl. 1, fig. 3 ; Zhang et al., 1998, figs. 6.9 –10, 7.1–2; Tiwari, Pant, 2004, p. 10, figs. 7A–7F ; Zhou et al., 2004, p. 353, pl. VI, figs. 1–4 ; Yin et al., 2007, pl. 5, figs. 5, 6 ; Yin et al., 2009, pl. 1, fig. 5 .
Form 2: Наговицин и др., 2004, с. 14, табл. 2 , фиг. 6 .
Knollisphaeridium maximum: Willman, Moczydłowska, 2008, p. 523, figs. 5E–5F ; Sergeev et al., 2011, pp. 995, 1004, figs. 7.5 –5a; Yin et al., 2011, p. 292, fig. 6e ; Liu et al., 2012, p. 860, figs. 5.14 –15; Liu et al., 2013, figs. 11N, 11O ; Xiao et al., 2014, pp. 30, 33, figs. 19.1 –7; Liu et al., 2014, p. 52, figs. 46.1 –12, p. 53, figs. 47.1 –7; Ouyang et al., 2017, fig. 9A–9H .; Ouyang et al., 2019, р. 185, figs. 13A–13D ; Liu, Moczydłowska, 2019, pp. 118–123, figs. 64, 65 .
Описание. Крупные сфероидальные оболочки 320–550 мкм в диаметре, покрытые многочисленными мелкими (до 4 мкм в длину и 2 мкм в поперечнике у основания) выростами, равномерно и плотно распределенными по всей поверхности. Расстояние между основаниями выростов 1–2 мкм. Выросты полые, простые, остроконические, свободно сообщающиеся с полостью оболочки.
Распространение. Уринская и баракунская свиты дальнетайгинского горизонта Восточной Сибири; венд. Формация Infrakrol Малых Гималаев Индии, формация Doushantuo Южного Китая, формация Tanana Австралии, группа Scotia Шпицбергена; эдиакарий.
Материал. Пять экземпляров удовлетворительной и десятки крупных фрагментов плохой сохранности из баракунской свиты.
Род Tanarium Kolosova, 1991, emend. Moczydłowska, Vidal et Rudavskaya, 1993
Tanarium conoideum Kolosova, 1991, emend. Moczydłowska, Vidal et Rudavskaya, 1993
Табл. I , фиг. 15
Tanarium conoideum: Kолосова, 1991, с. 57, фиг. 5.1 –3; Moczydłowska et al., 1993, pp. 514–516, figs. 10C–10D ; Knoll, 1994, fig. 4G ; Файзуллин, 1998, табл. 2 , фиг. 2–4 , 8 ; Grey, 2005, pp. 299–303, figs. 212A–212D, 213A–213D ; Moczydłowska, 2005, pp. 302, 303, figs. 7A, 7C, 7E ; Willman et al., 2006, p. 32, pl. 6, figs. 1, 2; Vorob’eva et al., 2007, pl. 1, fig. D ; Голубкова, Раевская, 2007, табл. 1 , фиг. 7 ; Willman, Moczydłowska, 2008, pp. 526, 527, fig. 12C ; Воробьева и др., 2008, фиг. 2a; Vorob’eva et al., 2009a, pp. 180, 182, figs. 7.4 –7; Vorob’eva et al., 2009b, p. 166, fig. 4e ; Голубкова и др., 2010, табл. 3 , фиг. 7 , табл. 4 , фиг. 2; Сергеев и др., 2010, табл. 2 , фиг. 9, 12 ; Moczydłowska, Nagovitsin, 2012, p. 18, fig. 8k ; Xiao et al., 2014, pp. 51–53, figs. 33.1 –6; Liu et al., 2014, p. 84, fig. 77 ; Голубкова и др., 2015, с. 447, рис. 2в.
Hocosphaeridium scaberfacium Zang: Zang, Walter, 1992, p. 61, figs. 45A–45E, 45G ; Gravestock et al., 1997, p. 91, fig. E .
Goniosphaeridium sp. A.: Zang, Walter, 1992, p. 54, figs. 45H–45I .
Goniosphaeridium conoideum: Zhang et al., 1998, p. 32, figs. 9.1 –4.
Tanarium sp.: Вейс и др., 2006, табл. 2 , фиг. 4 ; Воробьева и др., 2006, фиг. 2s.
Cavaspina sp.: Вейс и др., 2006, табл. 1 , фиг. 7 , табл. 2 , фиг. 5, 5a .
Описание. Сфероидальные оболочки полупрозрачные, тонкие, несущие конусовидные выросты, заостренные к окончаниям и расширенные в основании. Выросты полые, прозрачные, сообщающиеся с полостью оболочки, их основания четко отделены друг от друга. Большинство выростов обломаны, некоторые имеют загнутые окончания. Диаметр оболочек 150–200 мкм. Длина выростов до 35 мкм, ширина их основания 10–15 мкм. Количество видимых выростов по контуру оболочки до 20.
Распространение. Вычегодская свита Восточно-Европейской платформы; курсовская, паршинская свиты непского горизонта, уринская и баракунская свиты дальнетайгинского горизонта Восточной Сибири; венд. Формации Pertatataka, Tanana, Dey Dey Mudstone Австралии; формация Doushantuo Южного Китая; эдиакарий.
Материал. Шесть экземпляров плохой сохранности из баракунской свиты.
Tanarium pluriprotensum Grey, 2005
Табл. I , фиг. 1, 2
Tanarium pluriprotensum Grey: Grey, 2005, pp. 320–322, figs. 240A–240D, 241A–241B ; Willman et al., 2006, p. 36, pl. VII, fig. 5 ; Willman, Moczydłowska, 2008, pp. 527, 518, figs. 14A–14F ; Голубкова и др., 2015, с. 447, рис. 2b.
Описание. Сфероидальная оболочка размером 210 × 184 мкм, с тонкой полупрозрачной стенкой, несущей многочисленные выросты. Выросты одиночные, полые, соединяющиеся с полостью оболочки, довольно жесткие. Форма выростов цилиндрическая, их основания четко отделены друг от друга, окончания сужены и плавно закруглены (в большинстве случаев обломаны). Длина сохранившихся выростов от 29 до 34 мкм, ширина их основания от 4 до 7 мкм, ширина окончаний выростов 2–3 мкм. Выросты равномерно распределены по поверхности оболочки, наблюдаемое количество выростов 60.
Распространение. Баракунская свита дальнетайгинского горизонта Восточной Сибири; венд. Формации Pertatataka, Tanana, Dey Dey Mudstone Австралии, формация Doushantuo Южного Китая; эдиакарий.
Материал. Один экземпляр из баракунской свиты.
Tanarium pycnacanthum Grey, 2005
Табл. I , фиг. 10–12
Tanarium pycnacanthum: Grey, 2005, pp. 322–326, figs. 245A–245D, 246A–246E .
Solisphaeridium? sp.: Zang, Walter, 1992, p. 61; Grey, 2005, p. 322, figs. 245A–245D, 246A–246E ; Willman, Moczydłowska, 2008, p. 527, figs. 12D–12E .
Tanarium pluriprotensum: Liu et al., 2013, fig. 13A ; Liu et al., 2014, p. 113, figs. 84.1 –7.
Описание. Фрагмент оболочки, несущий многочисленные простые, длинные, тонкие выросты. Стенка оболочки, вероятно, изначально была прозрачной и тонкой, но из-за плохой сохранности виден только ее край. Выросты полые, открывающиеся в полость оболочки, слегка расширенные в основании и сужающиеся к окончаниям. Окончания заостренные, иногда изогнутые, часто обломанные. Основания выростов сближены, но четко разделены. Размер видимого фрагмента оболочки составляет 90 × 95 мкм. Длина выростов от 20 до 35 мкм, ширина их оснований до 3 мкм.
Распространение. Баракунская свита дальнетайгинского горизонта Восточной Сибири; венд. Формации Pertatataka, Tanana, Dey Dey Mudstone Австралии; эдиакарий.
Материал. Один неполный экземпляр из баракунской свиты.
Variomargosphaeridium sp.
Табл. II, фиг. 12
Описание. Темные непрозрачные сферические оболочки диаметром 140–230 мкм, несущие фрагментарно сохранившиеся единичные выросты (1–2 на каждом экземпляре). Выросты цилиндрические, трубчатые, с расширенным основанием, полые, сообщающиеся с полостью оболочки. Разветвленные окончания выростов сохранились плохо или не сохранились вовсе. Длина сохранившихся фрагментов выростов 15–25 мкм, ширина основания 7–18 мкм, ширина средней части выростов 5–15 мкм.
Материал. Четыре экземпляра плохой сохранности из баракунской свиты.
Форма 1
Табл. I , фиг. 13, 14
Описание. Оболочка сфероидальная, полупрозрачная, тонкая, возможно двухслойная, несущая один короткий широкий вырост. Диаметр оболочки 130–170 мкм. Вырост полый, сообщающийся с полостью оболочки, трубчатый, слегка расширяющийся от основания к окончанию в виде воронки. Длина выроста 20 мкм, ширина основания 18 мкм, ширина окончания 28 мкм.
Материал. Два экземпляра из баракунской свиты.
Форма 2
Табл. I , фиг. 16
Описание. Тонкая оболочка, несущая пять широко и регулярно разнесенных крупных выростов. Выросты цилиндрические, полые, соединяющиеся с полостью оболочки, длинные, гибкие, расширенные в основании и немного суженные к закрытым скругленным окончаниям. Из-за относительно небольшого размера оболочки и весьма крупных выростов вся форма имеет вид морской звезды. Размер оболочки 55 × 70 мкм, длина выростов 80–145 мкм, ширина выростов 15–18 мкм, ширина основания 26–28 мкм.
Материал. Один экземпляр из баракунской свиты.
ИНФРАГРУППА KELTMIIDES VOROB’EVA ET SERGEEV, 2018
Род Keltmia Vorob’eva, Sergeev et Knoll, 2009
Keltmia aff. cornifera Vorob’eva, Sergeev et Knoll, 2009
Табл. II, фиг. 7, 8
Неназванная оболочка: Воробьева и др., 2006, с. 369, рис. 2н.
Unnamed form 2: Vorob’eva et al., 2007, pl. 1, fig. G .
Keltmia cornifera: Vorob’eva et al., 2009a, pp. 180–181, figs. 11.1 –2.
Описание. Крупный фрагмент оболочки с одним полным и двумя частично сохранившимися выростами. Размер неполной оболочки 280 × × 200 мкм. Вырост полый, отделен от полости оболочки, в средней части имеет ширину 40 мкм, увеличивающуюся к основанию до 100 мкм, к дистальному концу сужается до 20 мкм, образуя “шейку”, несущую округлое, до 45 мкм в диаметре окончание.
Распространение. Вычегодская свита Восточно-Европейской платформы, баракунская свита дальнетайгинского горизонта Восточной Сибири; венд.
Материал. Один неполный экземпляр и два фрагмента плохой сохранности из баракунской свиты.
Род Palaeovolvox Zang et Walter, 1992
Palaeovolvox ooraminnus Zang et Walter, 1992
Табл. II, фиг. 15
Palaeovolvox ooraminnus: Zang, Walter, 1992, pp. 88, 91, figs. 67E–67G .
Описание. Крупная (200 × 170 мкм) сфероидальная оболочка с тонкой полупрозрачной стенкой. На внешней стороне оболочки расположено около сорока тесно связанных с ней более мелких (диаметром 20–28 мкм) темных сфероидальных клеток. Мелкие клетки расположены на поверхности оболочки нерегулярно, над уровнем “материнской” оболочки приподнимаются на 10–18 мкм.
Замечание. От формы с подобной морфологией Bullatosphaera velata Vorob’eva et al., 2009, описанной из уринской свиты и вендских отложений скв. Кельтменская-1 Мезенской синеклизы Восточно-Европейской платформы (Vorob’eva et al., 2009; Sergeev et al., 2011), баракунские микрофоссилии отличаются расположением мелких “дочерних клеток” на внешней поверхности оболочки и отсутствием внешней прозрачной пленки, покрывающей клетки.
Распространение. Баракунская свита дальнетайгинского горизонта Восточной Сибири; венд. Формация Pertatataka Австралии; эдиакарий.
Материал. Один экземпляр удовлетворительной сохранности из баракунской свиты.
Форма 3
Табл. II, фиг. 5, 6, 14
Описание. Трубчатый таллом простой, неветвящийся, расширенный с одного конца и сужающийся к другому, эластичный, часто перекрученный. Расширенное окончание имеет немного заостренную, эллипсоидальную или сфероидальную форму. Узкие окончания талломов часто обломаны. Длина талломов до 600 мкм, ширина 20–45 мкм, размеры расширенных частей 45–200 мкм.
Замечание. Плохая сохранность материала, фрагментированность микроостатков, отсутствие среди большого количества экземпляров даже частей ветвящихся ризоидов не позволяют уверенно отнести эту группу микроостатков к роду Vendomyces Burzin, 1993.
Материал. Десятки экземпляров плохой сохранности из баракунской свиты.
Форма 4
Табл. II, фиг. 9, 10
Описание. Фрагменты трубчатых нитей, состоящие из двух вложенных один в другой сегментов. Первая часть, цилиндрическая, является базальной, из которой вырастает вторая, воронковидная, часть. Ширина нижних сегментов у разных экземпляров колеблется от 38–45 до 55–70 мкм, ширина верхних сегментов у основания от 18 до 28 мкм, у расширенного окончания от 30 до 48 мкм, общая длина фрагментов 150–250 мкм.
Замечание. Похожее строение нитей было описано у таксона Cheilofilum hysteriopsis (Butterfield, 2005) из ранненеопротерозойской формации Wynniatt Арктической Канады и у неназванной Формы D из раннемезопротерозойской формации Gaoyuzhuang Китая (Shi et al., 2017). Оба эти морфотипа предположительно имеют грибную природу и сравниваются авторами с гифомицетовыми грибами. Плохая сохранность и фрагментированность микроостатков из баракунской свиты не позволяют отнести данную форму к одному из этих таксонов.
Материал. Три экземпляра из баракунской свиты.
ЦИАНОБАКТЕРИИ
Род Palaeopleurocapsa Knoll, Barghoorn et Golubic, 1975
Palaeopleurocapsa wopfnerii Knoll, Barghoorn et Golubic, 1975
Табл. II, фиг. 1, 2
Palaeopleurocapsa wopfnerii: Knoll et al., 1975, p. 2491, figs. 1a, 2a–2f; Nautiyal, 1983, pp. 177, 178, pl. 1, figs. 28–31 ; Nautiyal, 1984, p. 33, pl. 1, figs. 33–35 ; Buick, Knoll, 1999, pp. 756, 760, figs. 8.1 –4.
Описание. Мелкие сферические, эллипсоидальные клетки, собранные в параллельные ряды и образующие псевдонитчатую колонию. У линейно выстроенных в ряды клеток наблюдается ложное дихотомическое ветвление за счет разделения клеток, их ряды при этом остаются субпараллельными. Размер клеток от 4 до 10 мкм, некоторые из них содержат непрозрачные включения диаметром 0.5 мкм, толщина стенки от 0.5 до 1 мкм. Группы клеток упакованы в общую тонкую оболочку. Размер фрагмента колонии 120 × × 340 мкм. Хорошо различаются дистальная и проксимальная части колонии. В дистальной части клетки светлее, крупнее и свободнее расположены внутри “нитей”, в проксимальной – клетки темнее, мельче, их оболочка толще, а расположены они теснее.
Распространение. Формация Skillogalee Dolomite Австралии; неопротерозой. Группа Bangemall Австралии; мезопротерозой. Группа Semri Индии; нижний рифей.
Материал. Один экземпляр хорошей сохранности из баракунской свиты.
Род Obruchevella Reitlinger, 1948 emend. Yakshin et Luchinina, 1981
Obruchevella sp.
Табл. II, фиг. 4
Описание. Непрозрачные темно-коричневые до черных трихомы, свернутые в цилиндрическую спираль. Сохранившиеся фрагменты длиной 120–200 мкм и шириной 70–100 мкм. Ширина трихома постоянна и составляет около 20 мкм, расстояние между витками спирали 5–10 мкм.
Материал. Четыре экземпляра плохой сохранности из баракунской свиты.
Род Cehpalophytarion Scopf, 1968
Cehpalophytarion laticellulosum Schopf et Blacic, 1971
Табл. II, фиг. 3
Cehpalophytarion laticellulosum: Schopf, Blacic, 1971, p. 944, pl. 5, figs. 2,6 , pl. 6, fig. 3 .
Описание. Многоклеточные прямые или изогнутые неразветвленные трихомы без оболочки, состоящие из цилиндрических или почти квадратных клеток. Ширина клеток 3–4 мкм, длина 4–7 мкм, длина фрагментов нитей до 300 мкм. Нити образуют скопления в виде дерновин.
Распространение. Формация Bitter Springs Австралии; неопротерозой. Баракунская свита дальнетайгинского горизонта Восточной Сибири; венд.
Материал. Три экземпляра из баракунской свиты.
Благодарности. Авторы выражают искреннюю признательность Е.Ю. Голубковой (ИГГД РАН, г. Санкт-Петербург) за рецензирование статьи и ценные замечания, которые значительно улучшили данную публикацию.
Источники финансирования. Исследования проведены при поддержке гранта РФФИ 19-05-00155.
Конфликт интересов. Авторы заявляют, что у них нет конфликтов интересов.
Список литературы
Вейс А.Ф., Воробьева Н.Г., Голубкова Е.Ю. Первые находки нижневендских микрофоссилий на Русской плите: таксономический состав и биостратиграфическое значение // Стратиграфия. Геол. корреляция. 2006. Т. 14. № 4. С. 28–46.
Воробьева Н.Г., Сергеев В.Н. Stellarossica gen. nov. и инфрагруппа Keltmiides infragr.nov.: аномально крупные акантоморфные акритархи венда Сибири и Восточно-Европейской платформы // Палеонтол. журн. 2018. № 5. С. 91–100.
Воробьева Н.Г., Сергеев В.Н., Семихатов М.А. Уникальная нижневендская кельтминская микробиота Тимана: новые данные о палеонтологии венда и его глобальной характеристике // Докл. АН. 2006. Т. 410. № 3. С. 366–371.
Воробьева Н.Г., Сергеев В.Н., Чумаков Н.М. Новые находки нижневендских микрофоссилий в уринской свите: пересмотр возраста патомского комплекса Средней Сибири // Докл. АН. 2008. Т. 419. № 6. С. 782–787.
Голубкова Е.Ю., Раевская Е.Г. Нижневендский комплекс микрофоссилий внутренних районов Сибирской платформы // Тр. Международной конференции “Рассвет и закат вендской (эдиакарской) биоты. Происхождение современной биосферы”. М.: Геос, 2007. С. 39–42.
Голубкова Е.Ю., Раевская Е.Г., Кузнецов А.Б. Нижневендские комплексы микрофоссилий Восточной Сибири в решении стратиграфических проблем региона // Стратиграфия. Геол. корреляция. 2010. Т. 18. № 4. С. 3–27.
Голубкова Е.Ю., Зайцева Т.С., Кузнецов А.Б., Довжикова Е.Г., Маслов А.В. Микрофоссилии и Rb–Sr возраст глауконитов в опорном разрезе верхнего протерозоя северо-востока Русской плиты (скв. Кельтменская-1) // Докл. АН. 2015. Т. 462. № 4. С. 444–448.
Колосова С.П. Позднедокембрийские шиповатые микрофоссилии востока Сибирской платформы // Альгология. 1991. Т. 1. № 2. С. 53–59.
Кочнев Б.Б., Покровский Б.Г., Кузнецов А.Б., Марусин В.В. С- и Sr-изотопная хемостратиграфия карбонатных отложений венда–нижнего кембрия центральных районов Сибирской платформы // Геология и геофизика. 2018. Т. 59. № 6. С. 731–755.
Метелкин Д.В., Верниковский В.А., Казанский А.Ю. Тектоническая эволюция Сибирского палеоконтинента от неопротерозоя до позднего мезозоя: палеомагнитная запись и реконструкции // Геология и геофизика. 2012. Т. 53. № 7. С. 883–899.
Наговицин К.Е., Файзуллин М.Ш., Якшин М.С. Новые формы акантоморфных акритарх байкалия Патомского нагорья (уринская свита, Восточная Сибирь) // Новости палеонтологии и стратиграфии. Приложение к журналу “Геология и геофизика”. 2004. Т. 45. Вып. 6–7. С. 7–19.
Петров П.Ю. Постледниковые отложения дальнетайгинской серии: ранний венд Уринского поднятия Сибири. Сообщение 1. Баракунская свита // Литология и полезн. ископаемые. 2018а. № 5. С. 459–472.
Петров П.Ю. Постледниковые отложения дальнетайгинской серии: ранний венд Уринского поднятия Сибири. Cообщение 2. Уринская и каланчевская свиты и история бассейна // Литология и полезн. ископаемые. 2018б. № 6. С. 521–538.
Покровский Б.Г., Буякайте М.И. Геохимия изотопов C, O и Sr в неопротерозойских карбонатах юго-западной части Патомского палеобассейна, юг Средней Сибири // Литология и полезн. ископаемые. 2015. № 2. С. 159–186.
Покровский Б.Г., Мележик В.А. Буякайте М.И. Изотопный состав С, O, Sr и S в позднедокембрийских отложениях патомского комплекса, Центральная Сибирь. Сообщение 1. Результаты, изотопная стратиграфия и проблемы датирования // Литология и полезн. ископаемые. 2006а. № 5. С. 505–530.
Покровский Б.Г., Мележик В.А., Буякайте М.И. Изотопный состав С, O, Sr и S в позднедокембрийских отложениях патомского комплекса, Центральная Сибирь. Сообщение 2. Природа карбонатов с ультранизкими и ультравысокими значениями δ13С // Литология и полезн. ископаемые. 2006б. № 6. С. 642–654.
Пятилетов В.Г. Юдомский комплекс микрофитофоссилий Южной Якутии // Геология и геофизика. 1980. № 7. С. 8–20.
Пятилетов В.Г., Рудавская В.А. Акритархи юдомского комплекса // Вендская система. М.: Наука, 1985. Т. 1. С. 151–158.
Рудавская В.А., Васильева Н.И. Талсинский комплекс акритарх Непско-Ботуобинской антеклизы // Фитостратиграфия и морфология спор древних растений нефтегазоносных провинций СССР. Л.: ВНИГРИ, 1989. С. 5–11.
Рудько С.В., Петров П.Ю., Кузнецов А.Б., Шацилло А.В., Петров О.Л. Уточненный тренд δ13С в дальнетайгинской серии Уринского поднятия (венд, юг Средней Сибири) // Докл. АН. 2017. Т. 477. № 5. С. 590–594.
Сергеев В.Н., Семихатов М.А., Федонкин М.А., Воробьева Н.Г. Основные этапы развития докембрийского органического мира: Сообщение 2. Поздний протерозой // Стратиграфия. Геол. корреляция. 2010. Т. 18. № 6. С. 3–34.
Файзуллин М.Ш. Новые данные о микрофоссилиях байкалия Патомского нагорья // Геология и геофизика. 1998. Т. 39. № 3. С. 328–337.
Хераскова Т.Н., Буш В.А., Диденко А.Н., Самыгин С.Г. Распад Родинии и ранние стадии развития Палеоазиатского океана // Геотектоника. 2010. № 1. С. 5–28.
Чумаков Н.М. Оледенения Земли: история, стратиграфическое значение и роль в биосфере. М.: ГЕОС, 2015. 160 с. (Труды ГИН РАН. Вып. 611).
Чумаков Н.М., Покровский Б.Г., Мележик В.А. Геологическая история патомского комплекса, поздний докембрий, Средняя Сибирь // Докл. АН. 2007. Т. 413. № 3. С. 379–383.
Чумаков Н.М., Капитонов И.Н., Семихатов М.А., Леонов М.В., Рудько С.В. Возраст верхней части патомского комплекса Средней Сибири: изотопные датировки обломочных цирконов никольской и жербинской свит // Стратиграфия. Геол. корреляция. 2011. Т. 18. № 2. С. 115–119.
Чумаков Н.М., Семихатов М.А., Сергеев В.Н. Опорный разрез вендских отложений юга Средней Сибири // Стратиграфия. Геол. корреляция. 2013. Т. 21. № 4. С. 26–51.
Anderson R.P., McMahon S., Macdonald F.A., Jones D.S., Briggs D.E.G. Palaeobiology of latest Ediacaran phosphorites from the upper Khesen Formation, Khuvsgul Group, northern Mongolia // J. Systematic Palaeontol. 2019. V. 17. P. 501–532.
Buick R., Knoll A.H. Acritarchs and microfossils from the Mesoproterozoic Bangemall Group, northwestern Australia // J. Paleontol. 1999. V. 73. P. 744–764.
Butterfield N.J. Reconstructing a complex early Neoproterozoic eukaryote, Wynniatt Formation, arctic Canada // Lethaia. 2005. V. 38. № 2. P. 155–169.
Gravestock D., Morton J., Zang W. Biostratigraphy and correlation // Petroleum Geol. South Australia. 1997. V. 3. P. 87–97.
Grey K. Ediacaran Palynology of Australia // Mem. Assoc. Australasian Palaeontol. 2005. V. 31. 439 p.
Grey K., Calver C.R. Correlating the Ediacaran of Australia // The Rise and Fall of the Ediacaran Biota. Geol. Soc. Spec. Publ. London. 2007. V. 286. P. 115–135.
Hayes J.M., Kaplan I.R., Wedeking W. Precambrian organic geochemistry. Reservation of the record // Earth’s Earliest Biosphere. Princeton, New Jersey: Princeton Univ. Press, 1983. P. 93–134.
Knoll A.H. Microfossils in metasedimentary cherts of the Scotia Group, Prins Karl Forland, western Svalbard // Palaeontology. 1992. V. 35. P. 751–774.
Knoll A.H. Proterozoic and Early Cambrian protists: evidence for accelerating evolutionary tempo // Proc. Natl. Acad. Sci. 1994. V. 91. P. 6743–6750.
Knoll A., Barghoorn E.S., Golubic S. Palaeopleurocapsa wopfneri gen. et sp. nov., a late-Precambrian blue-green alga and its modern counterpart // Proc. Natl. Acad. Sci. USA. 1975. V. 72. P. 2488–2492.
Kuzmichev A.B., Bibikova E.V., Zhuravlev D.Z. Neoproterozoic (~800 Ma) orogeny in the Tuva-Mongolia Massif (Siberia): island arc–continent collision at the northeast Rodinia margin // Precambrian Res. 2001. V. 110. P. 109–126.
Liu P., Moczydłowska M. Ediacaran microfossils from the Doushantuo Formation chert nodules in the Yangtze Gorges area, South China, and new biozones // Fossils Strata. 2019. № 65. P. 1–172.
Liu P., Yin C., Chen S., Tang F., Gao L. Discovery of Ceratosphaeridium (Acritarcha) from the Ediacaran Doushantuo Formation in Yangtze Gorges, South China and its biostratigraphic implication // Bull. Geosciences. 2012. V. 87. P. 195–200.
Liu P., Yin C., Chen S., Tang F., Gao L. The biostratigraphic succession of acanthomorphic acritarchs of the Ediacaran Doushantuo Formation in Yangtze Gorges, South China and its biostratigraphic correlation with Australia // Precambrian Res. 2013. V. 225. P. 29–43.
Liu P., Xiao S., Yin C., Chen S., Zhou C., Li M. Ediacaran acanthomorphic acritarchs and other microfossils from chert nodules of the Upper Doushantuo Formation in the Yangtze Gorges area, south China // Paleontol. Mem. 2014. V. 72. P. 1–139.
Melezhik V.A., Pokrovsky B.G., Fallick A.E., Kuznetsov A.B., Bujakaite M.I. Constraints on 87Sr/86Sr of Late Ediacaran seawater: insight from Siberian high-Sr limestones // J. Geol. Soc. London. 2009. V. 166. P. 183–191.
Moczydłowska M. Taxonomic review of some Ediacaran acritarchs from the Siberian Platform // Precambrian Res. 2005. V. 136. P. 283–307.
Moczydłowska M., Nagovitsin K. Ediacaran radiation of organic-walled microbiota recorded in the Ura Formation, Patom Uplift, East Siberia // Precambrian Res. 2012. V. 198–199. P. 1–24.
Moczydłowska M., Vidal G., Rudavskaya V.A. Neoproterozoic (Vendian) phytoplankton from the Siberian Platform, Yakutia // Palaeontology. 1993. V. 36. P. 495–521.
Nautiyal A.C. Algonkian (Upper to Middle) micro-organisms from the Semri Group of Son Valley (Mirzapur Distt), India // Geol. Sci. J. 1983. V. 6 (2). P. 169–198.
Nautiyal A.C. Morphological, study of Algonkian cyanophytes from Lesser Himalaya and Plains region, with biostratigraphic significance // Proc. 10th Int. Colloquium on Micropaleontology and Stratigraphy. 1984. P. 25–40.
Ouyang Q., Guan C., Zhou C., Xiao S. Acanthomorphic acritarchs of the Doushantuo Formation from an upper slope section in northwestern Hunan Province, South China, with implications for early–middle Ediacaran biostratigraphy // Precambrian Res. 2017. V. 298. P. 512–529.
Ouyang Q., Zhou C., Xiao S., Chen Zh., Shao Y. Acanthomorphic acritarchs from the Ediacaran Doushantuo Formation at Zhangcunping in South China, with implications for the evolution of early Ediacaran eukaryotes // Precambrian Res. 2019. V. 320. P. 171–192.
Powerman V., Shatsillo A., Chumakov N., Kapitonov I., Hourigan J. Interaction between the Central Asian Orogenic Belt (CAOB) and the Siberian craton as recorded by detrital zircon suites from Transbaikalia // Precambrian Res. 2015. V. 267. P. 39–71.
Prasad B., Asher R. Record of Ediacaran complex acanthomorphic acritarchs from the lower Vindhyan succession of the Chambal Valley (East Rajasthan), India and their biostratigraphic significance // J. Palaeontol. Soc. India. 2016. V. 61. P. 29–62.
Schopf J.W., Blacic B.M. New microorganisms from the Bitter Springs Formation (Late Precambrian) of the north-central Amadeus Basin, central Australia // J. Paleontol. 1971. V. 45. P. 925–960.
Sergeev V.N., Knoll A.H., Vorob’eva N.G. Ediacaran microfossils from the Ura Formation, Baikal-Patom Uplift, Siberia: taxonomy and biostratigraphic significance // J. Paleontol. 2011. V. 85. № 5. P. 987–1011.
Shi M., Feng Q., Khan M.Z., Zhu S. An eukaryote-bearing microbiota from the early Mesoproterozoic Gaoyuzhuang Formation, Tianjin, China and its significance // Precambrian Res. 2017. V. 303. P. 709–726.
Shukla R., Tiwari M. Ediacaran acanthomorphic acritarchs from the Outer Krol Belt, Lesser Himalaya, India: their significance for global correlation // Palaeoworld. 2014. V. 23. P. 209–224.
Sovetov J.K. Vendian foreland basin of the Siberian cratonic margin: Paleopangean accretionary phases // Rus. J. Earth Sci. 2002. V. 4. P. 363–387.
Tiwari M., Knoll A.H. Large acanthomorphic acritarchs from the Infrakrol Formation of the Lesser Himalaya and their stratigraphic significance // J. Himalayan Geol. 1994. V. 5. P. 193–201.
Tiwari M., Pant C. Neoproterozoic silicified microfossils in Infra Krol Formation, Lesser Himalaya, India // J. Himalayan Geology. 2004. V. 25. P. 1–21.
Vorob’eva N.G., Sergeev V.N., Knoll A.H. Microfossil assemblages from the Vychegda Formation of the East European Platform passive margin – a biostratigraphic model for the Upper Riphean (Crygenian)/Vendian (Ediacaran) boundary // The Rise and Fall of the Vendian (Ediacaran) biota. Origin of the Modern Biosphere. Transaction of the International Conference on the IGCP Project 493. Moscow: Geos, 2007. P. 42–46.
Vorob’eva N.G., Sergeev V.N., Knoll A.H. Neoproterozoic microfossils from the northeastern margin of the East European Platform // J. Paleontol. 2009a. V. 83. P. 161–196.
Vorob’eva N.G., Sergeev V.N., Knoll A.H. Neoproterozoic microfossils from the margin of the East European Platform and the search for a biostratigraphic model of lower Ediacaran rocks // Precambrian Res. 2009b. V. 173. P. 163–169.
Willman S., Moczydłowska M. Ediacaran acritarch biota from the Giles 1 drillhole, Officer Basin, Australia, and its potential for biostratigraphic correlation // Precambrian Res. 2008. V. 162. P. 498–530.
Willman S., Moczydłowska M. Acritarchs in the Ediacaran of Australia – local or global significance? Evidence from the Lake Maurice West 1 drillcore // Rev. Palaeobot. Palynol. 2011. V. 166. P. 12–18.
Willman S., Moczydłowska M., Grey K. Neoproterozoic (Ediacaran) diversification of acritarchs – a new record from the Murnaroo 1 drillcore, eastern Officer Basin, Australia // Rev. Palaeobot. Palynol. 2006. № 139. P. 17–39.
Xiao S., Zhou C., Liu P., Wang D., Yuan X. Phosphatized acanthomorphic acritarchs and related microfossils from the Ediacaran Doushantuo Formation at Weng’an (South China) and their implications for biostratigraphic correlation // J. Paleontol. 2014. V. 88. P. 1–67.
Ye Q., Tong J., An Z., Tian L., Zhao X., Zhou S. Phosphatized fossil assemblage from the Ediacaran Doushantuo Formation in Zhangcunping area, Yichang, Hubei province // Acta Palaeontol. Sinica. 2015. V. 54. P. 43–65.
Yin C., Liu Y., Gao L., Wang Z., Tang F., Liu P. Phosphatized Biota in Early Sinian (Ediacaran)–Weng’an Biota and Its Environment. Beijing: Geol. Publ. House, 2007. 132 p.
Yin C., Liu P., Chen S., Tang F., Gao L., Wang Z. Acritarch biostratigraphic succession of the Ediacaran Doushantuo Formation in the Yangtze Gorges // Acta Palaeontol. Sinica. 2009. V. 48. P. 146–154.
Yin C., Liu P., Awramik S.M., Chen S., Chen S., Tang F., Gao L., Wang Z., Riedman L.A. Acanthomorph biostratigraphic succession of the Ediacaran Doushantuo Formation in the East Yangtze Gorges, South China // Acta Geol. Sinica. 2011. V. 85. № 2. P. 283–295.
Yin L. Microbiotas of latest Precambrian sequences in China // Stratigraphy and Palaeontology of Systemic Boundaries in China, Precambrian–Cambrian Boundary. 1987. V. 1. P. 415–494.
Yuan X., Xiao S., Yin L., Knoll A.H., Zhou C., Mu X. Doushantuo Fossils: Life on the Eve of Animal Radiation. Hefei, China: Univ. Sci. and Technology of China Press, 2002. 171 p.
Zang W.-L., Walter M.R. Late Proterozoic and Early Cambrian microfossils and biostratigraphy, Amadeus Basin, central Australia // Mem. Assoc. Australasian Palaeontol. 1992. V. 12. 132 p.
Zhang Y., Yin L., Xiao S., Knoll A.H. Permineralised fossils from the terminal Proterozoic Doushantuo Formation, South China // Paleontol. Soc. Mem. 1998. V. 50. P. 1–52.
Zhou C., Brasier M.D., Xue Y. Three-dimensional phosphatic preservation of giant acritarchs from the terminal Proterozoic Doushantuo Formation in Guizhou and Hubei Provinces, South China // Palaeontology. 2001. V. 44. P. 1157–1178.
Zhou C., Yuan X., Xiao S., Chen Z., Xue Y. Phosphatized fossil assemblage from the Doushantuo Formation in Baokang, Hubei Province // Acta Micropalaeontol. Sinica. 2004. V. 21. P. 349–366.
Дополнительные материалы отсутствуют.
Инструменты
Стратиграфия. Геологическая корреляция