

Успехи современной биологии, 2019, T. 139, № 4, стр. 402-413
Биомы северной Евразии
Я. И. Старобогатов 1, В. Ф. Левченко 2, *
1 Зоологический институт РАН
Санкт-Петербург, Россия
2 Институт эволюционной физиологии и биохимии им. И.М. Сеченова РАН
Санкт-Петербург, Россия
* E-mail: lew@lew.spb.org
Поступила в редакцию 13.03.2019
После доработки 15.03.2019
Принята к публикации 18.03.2019
Аннотация
Обсуждаются теоретические вопросы системы биомов, весьма слабо разработанной как у нас, так и за рубежом. Даются краткие описания некоторых биомов. Я.И. Старобогатов задумывал написать очерк обо всех биомах планеты, однако, в связи со смертью в 2004 г., он не успел этого сделать. Здесь приводится отредактированный текст его рукописи, в которой описаны только биомы северной Евразии. В первом разделе рассмотрены также законы размещения жизни на Земле, что важно, поскольку они являются основой для построения полной системы биомов планеты.
ПРЕДИСЛОВИЕ
Когда с А.С. Керженцевым и А.В. Яблоковым мы писали наши “Очерки биосферологии” (Яблоков и др., 2018) мы испытывали трудности в поиске подходящих публикаций, где непротиворечиво, четко и кратко излагалась бы система классификации биомов, а также законы размещения жизни на планете. Чаще всего биомы определяются описательно, причем разные авторы подходят к этому с разных позиций, и поэтому их подходы не всегда согласуются между собой. Позднее среди архивов я нашел незаконченную рукопись Ярослава Игоревича Старобогатова, посвященную именно этим вопросам (рис. 1). В ней с позиций экосистемного подхода обсуждаются теоретические вопросы недостаточно разработанной как у нас, так и за рубежом системы биомов, а также коротко даются описания некоторых из них.

Насколько я помню, Ярослав Игоревич собирался написать очерк обо всех биомах планеты и издать небольшую книжку с условным названием “Биомы”. Однако, к большому сожалению, он не успел это сделать в связи со смертью в 2004 г. Настоящая статья – практически весь отредактированный текст этой незаконченной рукописи, в которой описаны биомы только северной Евразии.
В рукописи практически не было никаких ссылок на литературные источники. По опыту совместной работы с Я.И. Старобогатовым11, это был его обычный стиль – вставлять ссылки, когда текст уже практически готов. Я не решился импровизировать и добавлять ссылки самостоятельно, надеясь на глубочайшую эрудицию Ярослава Игоревича. Исключением являются ссылки только на самые общие руководства и монографии.
В.Ф. Левченко
ВВЕДЕНИЕ
Обычно термин биом (Одум, 1975; Уиттекер, 1980) не поясняют, а иллюстрируют. Под биомами суши понимают большие экосистемы, соответствующие почвенно-растительным зонам (тундра, тайга, широколиственные леса, степи, пустыни, саванны, дождевые тропические леса и др.) или, иными словами, главные экосистемы планеты или главные подразделения биосферы. Более точно биомы можно определить как ландшафтообразующие экосистемы, занимающие второе место по значению для подразделения биосферы после основных областей жизни (суша, океан, мировой сток). Поскольку ландшафты существуют не только на суше, имеются подводные ландшафты в море и в водоемах мирового стока, вполне справедливо говорить не только о биомах суши – геобиомах, но и о биомах мирового стока – реобиомах, биомах морского дна – талассобиомах, биомах морской пелагиали – пелагобиомах.
Размещение биомов по поверхности и в водной толще нашей планеты подчиняется четырем фундаментальным законам размещения любых проявлений жизни: законам широтной зональности, биологической антимерии, или секторности, вертикальной зональности, или поясности, провинциальности.
ЗАКОНЫ РАЗМЕЩЕНИЯ ЖИЗНИ НА ЗЕМЛЕ
Закон широтной зональности
Закон широтной зональности связан прежде всего со сфероидностью Земли. Действительно, каков бы ни был наклон земной оси по отношению к плоскости эклиптики, всегда будут существовать зоны, где поступление солнечной энергии меньше, чем в других зонах. Поэтому имеются пояса на поверхности Земного шара, где среднегодовые температуры, среднегодовая освещенность меньше, чем в других поясах.
На суше широтная зональность выражена отчетливо в виде почвенно-растительных зон, в море она проявляется не менее резко – как зоны полярные, умеренные и теплые (тропико-субтропические). В водоемах мирового стока она очень часто маскируется действием других законов, так что меридионально текущие реки не проявляют заметной широтной зональности.
Закон биологической антимерии
Закон биологической антимерии иногда называют законом биологической симметрии или противоположно – законом биологической асимметрии, или же законом секторности. Название еще не установилось, но ни симметрия, ни асимметрия не характеризуют однозначно действие этого закона. И в этом контексте, на наш взгляд, более приемлемо название закон секторности. Действительно, суша Северного полушария делится на сектора с разной влажностью и морозностью климата, а море, в силу направления теплых и холодных течений, делится в Северном полушарии на западные холодные акватории и восточные – теплые в пределах каждого океана. Особенно резко секторность проявляется в Северной Америке (в основном на территории США), где почвенно-растительные зоны сменяют одна другую с востока на запад, а не с севера на юг. Однако действие этого закона проявляется не только в образовании климатических и почвенно-растительных секторов. Оно проявляется также в определенном состоянии распределения жизни в Северном и Южном полушарии. Так, если бы Южное полушарие было целиком океаническим, а Северное – континентальным, такого соответствия не было бы. Это дает основание предпочесть название “закон биологической антимерии”, где последнее слово – основание для некоторого соответствия или даже совместимости зон Северного и Южного полушарий. Понятно, что действие этого закона связано с вращением Земли вокруг оси, что создает силы Кориолиса, отклоняющие теплые течения на восток (в Северном полушарии) или на запад (в Южном), а также перемещающие циклоны и определяющие направление пассатов. Это существенно влияет на климат, а в связи с этим, и на органический мир.
Закон вертикальной зональности
Закон вертикальной зональности (или поясности) проявляется на суше и в море несколько по-разному. На суше это некоторое соответствие вертикальных зон в горах широтным зонам на равнинах. При продвижении от подножия гор к вершинам почвенно-растительные зоны с похолоданием климата сменяются сходно со сменой зон от экваториальных и умеренных к полярным. Классический пример – гора Килиманджаро, расположенная на 3° к югу от экватора, где идет смена от тропических условий у подножия к вечным снегам на вершине. В водоемах мирового стока этот закон проявляется в размещении различных климатических типов озер в разных вертикальных зонах. Например, среди горных озер преобладают олиготрофные.
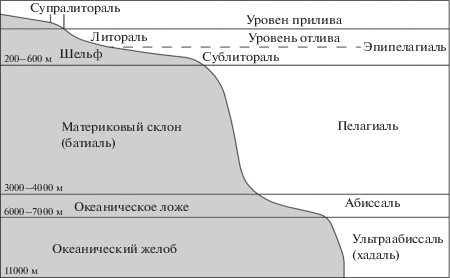
Иной тип вертикальной зональности в море. Наиболее крупными подразделениями тут являются: шельфовая зона – продолжение континентальной плиты под уровнем моря до глубин 200–600 м; материковый склон (батиаль) – от 200–600 до 3000–4000 м; ложе океана (абиссаль) – от 3000–4000 до 6000–7000 м. На это деление накладывается более дробное. Выделяется, например, приливо-отливная зона (в русской литературе – литораль, а в англоязычной – intertidal zone, поскольку под литоралью западные авторы понимают всю эвфотическую зону, примерно до 200 м). Выше приливо-отливной зоны располагается зона заплеска, увлажняемая морскими бризами (супралитораль). Эвфотическая зона обычно называется сублиторалью (не путать с супралиторалью), а шельфовая зона, расположенная глубже нижней границы эвфотической зоны, называется циркалиторалью. Если в пределах шельфа имеются впадины, характеризующиеся батиальными глубинами, эти акватории называются псевдобатиалями. На материковом склоне могут быть углубления с абиссальными глубинами – псевдоабиссаль. На океаническом ложе могут быть возвышенности с батиальными глубинами – талассобатиаль. Наконец, система желобов с глубинами до 11 000 м, особенно хорошо выраженными вдоль восточных берегов Азии, называется ультраабиссаль (у отечественных авторов), или хадаль (у западных авторов) (рис. 2).
БИОМЫ СУШИ
Биомы суши неоднократно подробно описывались (Вальтер, 1968, 1974, 1975), в связи с чем мы рассмотрим только некоторые из них, характерные для северной Евразии.
Тундра
Северная граница тундры, отделяющая ее от зоны арктических пустынь, совпадает с берегами арктических морей и островов, расположенных в пределах этих морей, за исключением Земли Франца Иосифа, Северной Земли и о. Северный Новой Земли, занятых арктическими пустынями. Южная граница отделяет северный берег Кольского п-ова и далее идет примерно по Полярному кругу, а восточнее р. Таз поднимается к северу до 70°–72° с.ш., а затем к востоку от Колымы спускается на юг вдоль 160–162 меридиана до берегов Охотского моря.
Физико-географические условия в тундре своеобразные – низкое положение солнца даже в день летнего солнцестояния, в связи с чем атмосфера задерживает значительную долю солнечных лучей. С другой стороны, длинный полярный день позволяет растениям в этот период осуществлять фотосинтез почти круглые сутки. В то же время низкая температура сильно замедляет обмен веществ. Первичная продукция в тундре довольно велика: вес биомассы – до 5 г/м2 сут. Большую роль играют ветра, скорость которых нередко достигает 10 м/с, а максимально – до 40 м/с. Это не позволяет развиваться древесной растительности и лишь по южной границе имеется разреженный древостой (лесотундра). Осадков мало – 200–300 мм в год, поэтому снеговой покров достигает лишь небольшой мощности и легко сдувается ветром. Мощным фактором является также вечная мерзлота. В европейской тундре поверхностный слой оттаивает летом до глубин 150 см (песок), 125 см (глина) и 35–40 см (торф). Восточнее на п-ове Гыдан на сухих отмирающих торфяниках оттаивающий слой не превышает 30 см, мерзлый слой препятствует проникновению воды, что приводит к заболачиванию местности. Низкие температуры замедляют или вовсе прекращают рост сфагновых мхов. Вместо них развиваются зеленые и печеночные мхи и лишайники (ягель или олений мох).
Типы тундр. Песчано-лишайниковая тундра тесно связана с песчаными почвами. В типичном покрове преобладают лишайники Cetraria, ягель Cladonia и зеленые мхи, в частности Polytrichum piliferum – один из видов кукушкина льна. Из высших растений более обычна ива травянистая Salix herbacea.
Кустарничково-лишайниковая тундра. Здесь доминируют те же виды лишайников и мхов, что и в предыдущем типе, но добавляются кустарнички шикши Empetrum nigrum, голубики Vaccinium uliginosum и некоторые другие, в том числе карликовая береза Betula nana. Ерники на песках занимают изолированные участки среди лишайниковых типов. Преобладает ерник – заросли карликовой березы, а наземный ярус занимают те же виды лишайников и мхов. Ерники и ивняки занимают те участки, где почва оттаивает на наибольшую глубину. Здесь образуются густые заросли нескольких видов карликовых ив и карликовой березы (еры).
Песчано-дернистые тундры располагаются пятнами на лишайниковом фоне. Здесь преобладают злаки, преимущественно овсяница овечья Festuca ovina, виды щучки Deschampsia, вейник лапландский Calamogrostis lapponica.
На суглинистых грунтах развиваются похожие типы тундр.
Это, в первую очередь, суглинисто-кочковатая тундра, которая имеет покров из зеленых мхов, а из высших растений здесь присутствуют горец живородящий Bistorta vivipara, вейник незамечаемый Calamagrostis neglecta и белокопытник холодный Nardosmia frigida.
В торфянистых тундрах развивается болотно-мочажинный комплекс – чередование мочажин с буграми. На мочажинах преобладают осоки и пушицы на полужидком торфе, а на буграх на торфе – мхи и ряд лишайников: цетрария и кладония (представители этих родов образуют ягель). Этот тип тундр связан с деградацией торфа и мохового покрова в связи с недостаточным для торфообразования поступлением воды.
Близ южной границы зоны тундр размещаются кустарничковые тундры и лесотундры. В первой из них преобладают заросли ерника (карликовой березы), кустарниковые ивы, багульник, голубика, брусника и др. В тундре этого типа много моховых и осоковых тундровых болот.
Лесотундра обычно рассматривается как экотон между биомами тундры и тайги. Характерным здесь является лесотундровое редколесье. В западных лесотундрах (Скандинавия) преобладает береза извилистая. К востоку от Белого моря основным видом становится ель сибирская. На восток от Урала до р. Пясина существенно преобладает сибирская лиственница, а еще восточнее – даурская лиственница.
Тайга
Северная граница тайги (зона хвойных и мелколиственных лесов) совпадает с южной границей тундры. Основные почвы – подзолистые. Южная граница тайги проходит от Стокгольма и Санкт-Петербурга–Новгорода примерно до русла р. Обь на уровне 55-й параллели. Далее она идет к северной части Монголии и крайнему северо-востоку Китая, а затем – к среднему течению р. Зея, после чего с севера и востока (по Сихотэ-Алиню) огибает широколиственные леса долин рр. Амур и Уссури. Этот биом занимает значительную часть площади России, причем в связи c большими размерами климатические условия в пределах этой обширной территории неоднородны. Средняя температура самого теплого месяца 10–19.5°C, среднегодовая – 2–5°C, годовое количество осадков 500 мм, реже – до 700 мм, причем по мере продвижения на восток количество осадков снижается. Разнообразие древесных пород очень невелико: европейская и сибирская ель, сибирская и даурская лиственницы, сибирская пихта, обыкновенная сосна, сибирская кедровая сосна и на крайнем востоке – саянская ель. Лиственные деревья – береза и осина распространены широко, но или составляют примесь к хвойным породам или приурочены определенным фазам яруса. Леса, как правило, 3–4-ярусные, но наиболее постоянными их компонентами являются хвойный и самый нижний напочвенный ярус. Это настолько бросается в глаза, что типы лесов обозначаются по древесному виду-эдификатору и растениям нижнего яруса.
Ельники подразделяют на: ельники-зеленомошники с нижним ярусом из зеленых блестящих мхов; ельники-долгомошники с нижним ярусом из кукушкина льна; ельники сфагновые с покровом из сфагнового мха; ельники болотно-травяные с высоким покровом из трав; ельники дубравные с подлеском из кустарников. По характеру травянистого яруса ельники зеленомошники подразделяют на ельники-черничники с травянистым ярусом из черники Vaccinium myrtillus, ельники-брусничники, ельники-кисличники и даже ельники с папоротником. Если к ели добавляется липа и дуб, то это ельники дубравные. Травянистый ярус дает основание для дальнейших подразделений. В одних случаях он представлен кислицей Oxalis acetosella с дубравными элементами, в других – звездчаткой Stellaria, в-третьих – медуницей неясной Pulmonaria obscura.
Боры (сосняки) по тому же принципу делят на несколько типов лесов. Это боры-беломошники с нижним ярусом из ягеля, боры-зеленомошники с нижним ярусом из зеленого мха, сосняки с покровом из трав, боры сфагнумовые с заболачивающимся покровом из сфагновых мхов, боры-долгомошники с нижним ярусом из кукушкина льна Polytrichum и боры дубравные на наиболее богатых почвах с примесью липы, орешника или дуба. Боры могут быть разделены более дробно по преобладающим группировкам травянистой растительности.
Как и предыдущие типы лесов, лиственничники могут быть разделены по составу нижних ярусов. Это лиственничники с толокнянкой Arctostaphylos uva-ursi, иногда замещающие сосняки с толокнянкой; лиственничники с брусникой Vaccinium vitis-idaea, в которых, кроме брусники, травянистый ярус представлен линнеей северной Linnaea borealis, грушанкой круглолистной Pyrola rotundifolia с примесью голубики Vaccinium uliginosum и брусники; лиственничники моховые – заболоченные с сильно развитым моховым покровом; лиственничники сфагновые – редкие сильно заболоченные мелколесья.
Стоит отметить, что хвойные леса на западе США резко отличаются от евразийских и представляют собой особый биом, развивающийся при более высокой температуре и влажности, за что их иногда называют дождевыми лесами умеренной зоны.
Степи
Для степей характерно большое видовое разнообразие, резко выраженные фенологические смены растительности и множественность жизненных форм. Различают два основных типа степей: северные (луговые, разнотравные) и южные (ковыльные). Количество осадков здесь существенно меньше, чем в тайге (250–450 мм в год).
Характерна смена сезонных аспектов в северной степи. Самая ранняя фаза, после схода снега, характеризуется цветением прострела Pulsatilla (сон-трава). Во второй фазе, конец апреля, добавляется низкая осока Carex humilis. В следующей фазе, начало мая, продолжается цветение двух упомянутых видов, но начинают цвести горицвет весенний Adonis vernalis и представители семейства гиацинтовых. В четвертой фазе, конец мая, зацветают сочевичник венгерский Lathyrus pannonicus, ирис безлистный Iris aphylla и лесная анемона Anemone sylvestris, в пятой фазе, конец мая– начало июня, цветут незабудки Myosotis. После отцветания растений пятой фазы, первая половина июня, начинается лето. Шестая фаза знаменуется цветением шалфея Salvia, который в шестой фазе, середина июня, дополняется козлобородником восточным Tragopogon orietalis. В седьмой фазе, вторая половина июня, основной фон степи становится белым из-за цветения горного клевера Trifolium montanum, нивяника обыкновенного Leucanthemum vulgare, таволги шестилепестной Filipendula hexapetala. К белому фону добавляются лиловые вкрапления – цветут колокольчики Campanula и короставник полевой Knautia arvensis. В восьмой фазе, первая половина июля, цветут эспарцет виколистный Onobrychis viciifolia и желтый подмаренник Galium verum. Во второй половине лета цветет дельфиниум Delphinium – это девятая фаза, вторая половина июля. Далее в десятой фазе, конец июля–начало августа, цветет чемерица черная Veratrum nigrum. В августе и сентябре зацветающих растений уже нет, и такой облик северной степи остается до выпадения снега. Таким образом, для северной степи характерна смена многочисленных красочных аспектов, господство разнотравных незлаковых видов, а среди злаковых – преобладание широколистных, рыхлокустовых и корневищных видов, а также господство многолистных видов при почти полном отсутствии эфемеров и эфемероидов (исключая, конечно, гиацинтовые).
Отличие южных степей от северных всецело связано с изменением климатических условий. Выделяют два варианта ковыльных (южных) степей: более северный – красочный и более южный – бескрасочный. В отличие от северных степей, разнообразие фаз цветения заметно меньше, и они не столь ярки. Ранневесенняя фаза, вторая половина марта – первая половина апреля, характеризуется развитием напочвенной синезеленой водоросли Stratonostoc и оживанием мха Barbula. В средневесеннюю фазу, вторая половина апреля, степь зеленеет, и зацветают ирисы, горицвет, гусиные луки Gagea и тюльпаны Tulipa. В поздневесеннюю фазу, первая половина мая, интенсивно цветут сочевичник венгерский, лесная анемона, лапчатки Potentila, пион тонколистный Paeonia tenuifolia, а из кустарников – терн Prunus spinosa, миндаль Amygdalus nana, карагана кустистая Caragana frutex. В раннелетнюю фазу, вторая половина мая–первая половина июня, цветут злаки, в первую очередь ковыли Stipa, типчак Festuca valesiaca, тонконог гребенчатый Koeleria cristata, кострец береговой Bromopsis riparia. Из двудольных в эту пору цветут клевер горный, таволга шестилепестная, незабудки, колокольчики. Количество цветущих двудольных резко уменьшается в среднелетнюю фазу, вторая половина июня–первая половина июля, степь буреет от отмирающих растений, цветут подорожник средний Plantago media, зопник Phlomoides, шалфей дубравный Salvia nemorosa и многие виды, образующие осенью перекати-поле. В позднелетнюю–раннеосеннюю фазы доминируют ковыль волосовидный Stipa capillata, или тырса, несколько видов зонтичных и астровых. В позднеосеннюю фазу, с начала сентября до выпадения снега, растения уже отцвели, и осенняя степь вновь вызывает к жизни синезеленые водоросли и мхи. Если сравнить с картиной, которая приведена для северных степей, то обнаруживается наличие фаз с господством ковыля. Разнообразие двудольных уменьшается, и остаются более ксерофильные виды, появляется ряд растений, образующих перекати-поле, заметно, особенно в весенние фазы, множество однолетников, сменяется набор видов мхов, и большую роль играют синезеленые водоросли Stratonostoc. Как легко видеть, смена сезонных аспектов выражается в замещении цветущих видов. Наблюдается согласование периодичности цветения с развитием насекомых-опылителей.
МОРСКИЕ БИОМЫ
Морские биомы (и пелагобиомы, и талассобиомы), как и наземные (геобиомы), характеризуются тем, что сукцессионные процессы происходят внутри биома и направлены на ликвидацию нарушений. В морских биомах в той или иной мере проявляются главным образом сукцессионно-переходные процессы, как правило, связанные с изменением гидрологических условий.
Пелагобиомы
В качестве примера пелагобиома приведем центральную акваторию субтропического антициклонического круговорота в Тихом океане. Эта акватория окаймлена системой течений Куросиа: северо-тихоокеанским, калифорнийским и северным пассатным. Водная масса внутри этого колодца малоподвижна, обладает сравнительно постоянной слегка повышенной соленостью 95.5‰ и в продукционно-биологическом отношении напоминает пустыню, что связано с недостатком биогенных элементов. Основные первичные продуценты – диатомовые водоросли и перидинеи, например Ceratium deflexum. Несомненно, существенную роль в продукции играет нанопланктон, не улавливаемый планктонными сетями. Общая среднегодовая первичная продукция этого биома менее 10 мг C/м2 × сут. Столь низкая продуктивность объясняется тем, что биогены выносятся из эвфотической зоны с “дождем трупов” (то есть опадом биогенов на дно), вымершие клетки фитопланктона перерабатываются бактериями, максимум концентрации биомассы которых почти совпадает с верхней границей термоклина (40–60 м). Диатомовые служат пищей веслоногих рачков – каланоидов, например Clausocalanus paululus, и эуфаузиид Euphausia brevis. Планктонных ракообразных поедают планктоноядные рыбы: Opostomias mitsuii, Idiacanthus antrostomus, Protomyctophum crockeri. Большинство их видов распространено и за пределами обсуждаемой области – или по всей тропической зоне, или, по крайней мере, в областях нескольких основных субтропических круговоротов. Напомним, что их пять (по два в Тихом и Атлантическом океанах и один – в Индийском). Планктоноядные рыбы служат пищевым объектом для хищных рыб и акул (синяя акула, сайра, макрелещука, тунец), а также кальмара Ommastrephes bartrami. Китообразные в силу своей подвижности, как и хищные рыбы, распространены более широко. В обсуждаемой акватории обычны кашалот, питающийся кальмарами, полосатики и горбач, питающиеся эуфаузиидами.
Биомы апвеллинга
Апвеллингом называется процесс, при котором глубинные, более холодные, но богатые биогенами воды выносятся к поверхности моря. Это происходит вследствие ветрового сгона поверхностных вод. Основные апвеллинги существуют там, где холодные части антициклонических круговоротов расположены вблизи берега, так что ветры, сгоняющие поверхностный слой воды, почти постоянно дуют с берега в море. Это приводит к тому, что суша, расположенная вблизи апвеллинга, испытывает дефицит влаги. На западе США этот дефицит компенсируется туманами. Процесс подъема глубинных вод очень медленный: его скорость по вертикали около 20 м в месяц. Сама акватория апвеллинга невелика, но связанное с ней повышение продуктивности охватывает более широкую акваторию. Усиленное перемешивание вблизи поверхности обогащает эвфотический слой биогенами и препятствует быстрому осаждению детрита. С высокой продуктивностью связано и богатство органического вещества в донных отложениях. По сравнению с центральными акваториями круговоротов, первичная продукция здесь много выше 250–500 мг C/м2 × сут. Набор массовых видов и пищевые связи – почти такие же, как и в центральных акваториях субтропических круговоротов. Первичная продукция создается диатомовыми водорослями (в том числе и массовыми, перечисленными выше), перидинеями и нанопланктонными фотосинтезирующими организмами. То же можно сказать и о веслоногих ракообразных – каланоидах, эуфаузиидах, планктоноядных рыбах и их потребителях – хищных рыбах, морских птицах, которых в подобных районах очень много. Их может быть столь много (например, в районе Перуанского апвеллинга), что их помет образует промышленно используемые запасы гуано. В районе этого апвеллинга преобладают рыбоядные птицы из отряда веслоногих, что связано с обилием мелких пелагических рыб. В противоположность этому, в районе Калифорнийского апвеллинга много планктоноядных (мелкие чистиковые и трубконосые), рыбоядных (средние и крупные чистиковые и бакланы) и бентосоядных птиц (нырковые утки и гаги).
Высокая продуктивность апвеллингов делает акватории, связанные с ними, одними из наиболее богатых промысловых районов.
Талассобиомы шельфовой зоны
Как пример биомов шельфа рассмотрим Карское море (рис. 3). Это море занимает обширный участок шельфа западной Азии и граничит с запада с Новой Землей и Землей Франца-Иосифа, с северо-востока – с Северной Землей, с юго-востока и юга – с п-овами Таймыр, Гыдан и Ямал. Море сообщается с центральными частями Северного Ледовитого океана через широкий пролив между Землей Франца-Иосифа и Северной Землей. Как и в других морях северных районов Азии, поверхностный слой воды в Карском море заметно опреснен за счет стока рр. Обь, Таз, Енисей, и вблизи выхода из Обской губы поверхностная соленость достигает 7–8‰, при этом летняя температура там 3–4°С. По мере приближения к полярному бассейну соленость возрастает до 32–34‰. На глубине 50–100 м располагается охлажденный слой, возникающий за счет погружения сильно охлажденных вод в зимнее время. Глубины занимает холодная вода с температурой от –1.6 до –1.7°С. Более соленые и относительно теплые воды проникают с севера из центральной Арктики, с запада – в обход северной части Новой Земли и через пролив Карские ворота. Поверхность дна относительно ровная с преобладающими глубинами 50–80 м, далее она заметно углубляется к северу, достигая между Землей Франца-Иосифа и Северной Землей глубин в 500–800 м. Вдоль восточного берега Новой Земли тянется восточно-новоземельский желоб с глубинами 300–400 м, дно юго-восточной половины моря покрыто песчанистыми грунтами (песок, илистый песок), образующимися под действием рек. Дно северо-западной половины и восточно-новоземельского желоба покрыто коричневыми илами с высоким содержанием железа и марганца. Первичную продукцию в Карском море создает фитопланктон, в котором резко преобладают диатомовые водоросли и перидинеи, а также макрофиты – по преимуществу бурые и красные водоросли. Гидрологические условия позволяют образоваться здесь характерной приливно-отливной зоне (литорали) со специфическим набором растений и животных.
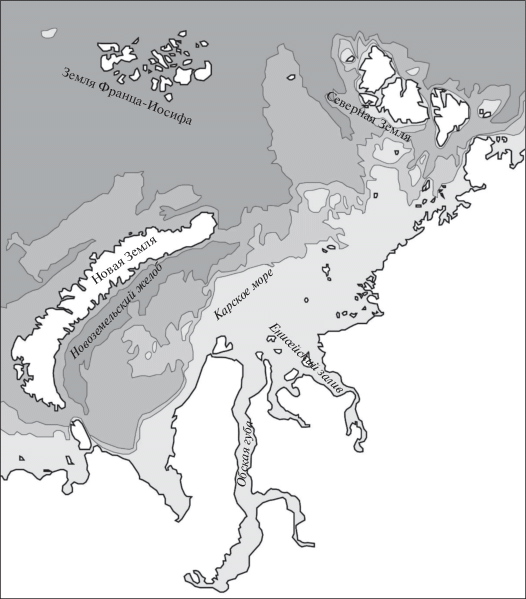
Виды, характерные для баренцевоморской литорали обитают в Карском море в верхней сублиторали. Из бурых водорослей характерны три вида ламинарий, например Laminaria agardhii, фукоиды Fucus evanescens и F. inflatus, Desmarestia aculeata, а из красных – Rhodymenia palmata, Delesseria sinuosa, Rhodomela lycopodioides и ряд других. Распространены и некоторые зеленые водоросли, например Chaetomorpha melagonium. В бентосе малых и средних глубин нередки виды, выносящие заметное опреснение, например двустворчатый моллюск Portlandia arctica. Здесь же встречается Serripes groenlandicus и ряд других арктических сублиторальных видов, приуроченных к более высоким соленостям. На коричневых илах более глубоких участков моря многочисленны фораминиферы Hormosina globulifera, Saccorhiza ramosa, крупный восьмилучевой коралл Umbellula еncrinus, ряд полихет, приуроченных к илам, например Nephthys ciliata, Thelepus cincinnatus, Actinula, а из двустворчатых моллюсков-фильтраторов – Serripes groenlandicus, Astarte crebricostata. Ими питаются морские звезды Icasterias panopla, Urasterias lincki и др. Обычны равноногие раки, в частности морские тараканы (виды рода Saduria) и креветки, например Sclerocrangon ferox. Изменения видового богатства связаны с характером грунта и соленостью. Довольно обильны животными (биомасса – до 10–50 г/м2) мало опресняющиеся каменистые побережья Новой Земли и почти полносоленые участки со стороны северной части моря. Много беднее населены коричневые илы (биомасса не превышает 10 г/м2), в то время как части, подверженные действию речного стока, демонстрируют наибольшие величины биомассы (50–100 г/м2 и более).
Эстуарные биомы
Этот тип биомов можно отнести к талассобиомам и к реобиомам. Представляется, что первое название правильнее, поскольку тут не оговаривается эрозионная деятельность рек. К категории эстуарных акваторий относятся собственно эстуарии, такие как Невская губа (эстуарий Невы), дельты рек (например, Дона и Волги), лиманы (Днепро-Бугский лиман, Куршский залив) и лагуны (вроде лагуны Лебяжьей на юге Приморского края). Хотя очевидно, что происхождение эстуарных участков и их история весьма различны, их всех объединяет кратковременность существования в геологических масштабах времени. Так, дельта Волги за четвертичное время перемещалась вверх и вниз, так что остатки самых верхних положений дельты находятся у оз. Сарпа к югу от Волгограда (Сарпинская дельта), а самых нижних – под водами у северного Каспия.
Коса, отделяющая Куршский залив, перемещалась, по крайней мере, в прошлом в сторону коренного берега. Это хорошо видно, если сравнивать Куршский и Вислинский заливы. Лиманы Причерноморья формировались при колебаниях уровня моря, так что нижнюю часть дельты реки подпруживала образующаяся морем пересыпь, что превратило некоторые лиманы, например Куяльницкий лиман под Одессой, в соленые озера.
Общей чертой всех эстуарных акваторий является то, что они располагаются на контакте реки и моря. Внешний район устья находится на одном уровне с базисом эрозии, что приводит к резкому замедлению течения и выпадению в осадок даже самых тонких взвесей. Это в свою очередь означает обогащение осадков детритом. Поступает из рек и растворенное органическое вещество. Другая особенность эстуарных акваторий – пониженная соленость в результате смешения речной воды с морской. Соленость также непостоянна и зависит от количества воды, сбрасываемой рекой. В реках, впадающих в моря с высокими приливами, приливная волна может подниматься вверх по реке на несколько километров, что ведет к относительно кратковременным (полусуточным) колебаниям солености. Нередко изменения солености превышают критический диапазон колебаний солености для ряда организмов (5–6‰), что, например, делает невозможным обитание здесь неэвригалинных морских и пресноводных животных. С другой стороны, постоянный приток биогенов, взвешенного и растворенного органического вещества стимулирует высокую продуктивность этой зоны. Этот поток органического вещества и биогенов вместе с органическим веществом, приносимым с моря (особенно при сильном волнении), создает мощную энергетическую субсидию, обеспечивающую постоянную высокую продуктивность. Первичная продукция вырабатывается обширной прибрежной растительностью, одноклеточными водорослями. Так, в дельтах рек, впадающих в Азово-Черноморский бассейн и Каспий, массовым прибрежным видом является тростник Phragmites, к которому добавляются рогоз Typha, сусак Butomus, ежеголовник Sparganium. Отмирающая растительная масса перерабатывается бактериями вместе с органическими остатками, поступающими с суши.
Из животных организмов здесь могут обитать эвригалинные формы, способные переносить колебания солености. Это либо физиологически пресноводные организмы, способные жить в опресненной воде во взрослом состоянии, но размножающиеся в полносоленой морской воде, либо формы, способные на некоторое время изолироваться от окружающей среды с неблагоприятной для них соленостью.
Одной из важнейших групп, использующих взвешенное органическое вещество, являются двустворчатые моллюски. В Азово-Черноморском лимане это солоноватоводные виды родов Hypanis и Adacna, а в эстуариях южного Приморья – Laternula limicola, Nuttallia olivacea и Macoma tokyoensis. Здесь же живут многочисленные брюхоногие, питающиеся осевшим детритом и одноклеточными водорослями. В лиманах Причерноморья – это виды семейства Pyrgulidae, а в южном Приморье – Assimineidae, Iravadiidae и Battilariidae. На илистых грунтах обитают полихеты: в южном Приморье – это Nereis japonica, а в Причерноморье – виды родов Hypania и Hypaniola. На растительности живут многочисленные личинки растительноядных насекомых. Есть личинки насекомых и на грунте – ручейники и Chironomidae. Наконец, эстуарные акватории служат местом нагула мальков рыб. Это богатство жизни и высокая продуктивность характеризуют эстуарные акватории как особые биомы, весьма важные во многих отношениях, в том числе и для хозяйственной деятельности.
Реобиомы
Реобиомы принципиально отличаются от биомов суши и мирового океана характером происходящих в них сукцессионных процессов. Если на суше и в море сукцессионные процессы ведут к восстановлению разрушенных его участков и лишь сукцессионно-переходные процессы (например, смена одного эдафического климакса другим) могут в той или иной мере влиять на облик биома, то реобиомы постоянно подвержены аллогенным сукцессионным процессам (которые вызваны внешними факторами, в том числе влиянием колебаний уровня воды, меандрированием рек и т.п.), а в некоторых частях биома – аутогенным сукцессионным процессам. Это, в конечном итоге, является следствием непрерывности мирового стока.
Основой процесса стока является река. Течение реки хотя и непрерывное, но скорости его меняются в зависимости от поперечного сечения потока и возвышения верховьев над базисом эрозии. Непрерывный речной поток (речной континуум) распадается на участки с быстрым течением – перекаты – и с медленным – плесы. На участках с быстрым течением тонкие взвеси, как правило, не осаждаются, быстрый поток размывает дно и, по преимуществу, правый берег (в северном полушарии). Дно на таких участках каменистое, галечное или гравийное. Фотосинтез обеспечивается водорослями (диатомовыми и зелеными), а животные обладают различными приспособлениями для прикрепления к твердому субстрату, чтобы противостоять стоку. В таких условиях преобладают личинки поденок, веснянок, мошек, комаров (блефароцерид), клопов (афелохирусы), а также раки-бокоплавы. Реже встречаются моллюски, способные плотно присасываться к камням, например, речная чашечка Ancylus fluviatilis. Некоторые группы насекомых во взрослом состоянии мигрируют вверх по течению, что компенсирует снос личинок. Собственного планктона на таких участках рек нет, планктонные организмы, поступающие из озер и более мелких водоемов речной системы, проносятся вниз, по крайней мере, до участков с замедленным течением. Иначе говоря, в ряде случаев в таких фрагментах реобиомов мы имеем относительно постоянные сообщества, но не полноценные экосистемы с замкнутым круговоротом.
Плесы характеризуются в связи с замедленным течением более тонкодисперсными осадками, наличием прибрежных зарослей, использующих оседающую взвесь и биогены, как приносимые, так и освобождающиеся при разложении отмерших растений. Переработанные остатки растений и отмерших организмов образуют взвешенное и растворенное органическое вещество, постепенно перерабатываемое бактериями. Характерно, что пресноводные животные, в отличие от морских, не способны усваивать растворенное органическое вещество поверхностью тела, так как их покровы, в связи необходимостью ионной регуляции внутри тела, надежно защищены от проникновения веществ извне.
Сообщества (иногда неточно называемые экосистемами), расположенные на участках с быстрым течением, называются лотическими (от лат. lotus – омываться). В противоположность этому, непроточные и слабопроточные водоемы речного бассейна обладают совершенно иными первичными продуцентами, иным характером осадков и иным ходом сукцессионных процессов. Такие экосистемы принято называть лентическими (от лат. lente – спокойно).
К числу аутогенных сукцессионных процессов относится зарастание озер. Нормально в эвфатической зоне озера фотосинтезирующие организмы располагаются в виде нескольких поясов, сменяющих друг друга по глубине мест обитания (Логинова, Лопух, 2011). Самый верхний пояс (у уреза воды) занят мелководными растениями: частухой Alisma, стрелолистом Sagittaria, водяной сосенкой Hippuris vulgaris, дербенником иволистным Lythrum salicaria, а также некоторыми видами осок, например Carex, и другими растениями, у которых находится под поверхностью воды низ стебля и корни (последние – в грунте). Далее вглубь располагается пояс камышей – озерный камыш Scirpus lacustris, тростник, хвощ Equisetum и др., причем эти растения образуют сплошные заросли, соотносимые с разными глубинами. В целом этот пояс приурочен к глубинам до 1.5–3 м, так что большая часть растения возвышается над водой. Еще глубже располагается пояс нимфей. Обитающие здесь растения – желтая кубышка Nuphar lutea, кувшинка Nymphaea, плавающий рдест Potamogeton natans – целиком находятся под водой на глубине до 2.5–4 м и лишь их листья плавают на поверхности воды. В следующем поясе все растения полностью находятся под водой на глубине не менее 3 м. Здесь обитают рдест блестящий Potamogeton lucens, рдест пронзеннолистный Potamogeton perfoliatus и некоторые другие виды этого рода, а также перистолистник Myriophyllum (уруть). Далее в глубину (до 6 м) идет пояс, часто называемый поясом погруженных макрофитов, или поясом харовых водорослей. Здесь обитают харовые: хара Chara, нителла Nitella; узколистные рдесты, например, рдест туполистный Potamogeton obtusifolius и рдест фриза Potamogeton friesii; роголистник погруженный Ceratophyllum demersum. Характерно, что представители последнего рода цветут под водой. Наконец вплоть до границы зоны распространения света располагаются только одноклеточные и нитчатые водоросли, прежде всего диатомовые, и цианобактерии.
Отмершие части всех этих растений пополняют донные отложения в пределах каждого пояса и тем самым постепенно повышают уровень дна. Кроме того, значительная часть осадков особенно при слабом развитии растительности образована за счет планктона. Отмершие планктонные организмы дают богатый органикой сапропель, или гиттию. Накопление осадков ведет к смене перечисленных выше поясов – более глубоководные из них сменяются более мелководными. Обилие остатков высших растений приводит к образованию торфа с заметной примесью минерального вещества. Дальнейшее развитие водоема, когда остается только пояс мелководной растительности, приводит к его заболачиванию. При этом сначала сохраняется грунтовое питание (поступление биогенов с почвенными водами), а далее происходит переход в фазу атмосферного питания. Эти последние фазы связаны с поселением на месте озера наземной растительности. Сначала здесь обычно поселяется ивняк, сменяемый ольхой, а затем березой и сосной. Когда болото разрастается настолько, что не может более использовать грунтовое питание, главную роль начинают играть мхи, в первую очередь торфяные (сфагновые), широко использующие атмосферное питание, а из деревьев начинает широко распространяться сосна. Как легко видеть, сначала идет сукцессия в водоеме, а затем она переходит в наземную сукцессию, завершающуюся в условиях зоны хвойных лесов климаксом – сосняком. Такой переход от пресноводного биома к сухопутному очень характерен для лесной зоны.
Зарастание озера может ускоряться образованием сплавин. В этом случае с берега нарастает плавающий ковер, состоящий из растений с мощными корневищами (вахта, белокрыльник, сабельник). Отмирающие нижние части этих растений формируют торф сплавник. Сплавины сильно уменьшают зеркало озера и нередко озерные заросли начинаются с пояса нимфей. Размывание нижних слоев сплавины приводит к поступлению в озерную котловину торфогенного ила.
Совершенно иные сукцессионные процессы, уже чисто аллогенные, происходят в речной долине. Здесь все зависит от эрозионной деятельности реки, меняющей ход русла, создающей и уничтожающей пойменные водоемы и обусловливающей рельеф поймы, а также от образования/исчезновения русловых петель (меандрирования реки).
ЗАКЛЮЧЕНИЕ
К сожалению, на этом рукопись работы Я.И. Старобогатова, посвященной биомам, заканчивается. Фактически, в ней описаны закономерности распространения жизни на планете, а также основные биомы северной Евразии, находящиеся на территории России. Это для нас имеет самостоятельный интерес.
В этих же рукописных черновиках содержится небольшой фрагмент более детального описания сукцессионных процессов, происходящих в водных экосистемах. На наш взгляд, он непосредственно относится к теме биомов, но также имеет самостоятельную ценность. Этот фрагмент приводится ниже.
ДОПОЛНЕНИЕ
О сукцессионных процессах в водных экосистемах
Для начала рассмотрим строение поймы. Поперечное сечение поймы любой реки ассиметрично. Правый коренной берег речной долины в северном полушарии обычно крутой и часто приближенный к руслу, что соответствует закону Бэра. Непосредственное русло окаймляет прирусловый вал (бечевник), далее поверхность центральной поймы располагается ниже вала, а на участке приближенном ко второй террасе (первой считается сама пойма) имеются выходы родниковых вод, обусловливающих наличие здесь притеррасных болот или притеррасной речки. При разливе в первую очередь заливается прирусловый вал, и далее вода выходит на центральную пойму. При этом течение воды, попавшей на центральную пойму, замедляется, что способствует выпадению осадков, влекомых паводковой водой. На луговых поймах осадки накапливаются слоями (слоистая пойма), погребая предшествующие им, тогда как на облесенной пойме скорость движения паводковых вод сильно замедляется, и здесь выпадает уже только тонкодисперсная часть взвеси; образуется слой полужидкого наилка, который по высыхании распадается на комки (зернистая пойма). Не надо забывать, что во время половодья долина заливается почти целиком, и вода идет не вдоль русла (которое может быть очень извилистым), а вдоль оси долины (вдоль тальвега).
В ходе речной эрозии русло реки сильно меняется: образуются и со временем теряются петли русла. Этот процесс очень важен для понимания аллогенной сукцессии водоемов речной долины. Петлеобразный водоем, еще сохранивший открытую связь с рекой, называется затоном. Он имеет озерный режим и мог бы развиваться по описанному выше пути, если бы не пребывало значительного количества речных наносов. Поступление последних превращает затон в старицу – водоем более мелкий и слабо связанный (по преимуществу стоком) с рекой. Следующее за этим заиление водоема осадками превращает его в полупостоянный, а затем и в пересыхающий (периодически) водоем. Дальнейшая его судьба – или превратиться в участок пойменного луга, или из-за перемещения речного русла вновь оказаться фрагментом затона или старицы. Впрочем, и на лугах центральной поймы имеются периодические и даже эфемерные водоемы. Все эти типы водоемов обладают характерными для них водным режимом, растительностью и животным миром. Так эфемерные водоемы высыхают более чем на три месяца в году, зарастают луговыми травами (мятлики болотный Poа palustris и луговой Poa pratensis, ситник жабий Juncus bufonius) и обзаводятся своим комплексом моллюсков, способных переносить длительное высыхание. Периодические водоемы высыхают на 1–3 мес. и зарастают влаголюбивыми растениями (осоки заячья Carex leporina и сероватая Carex canescens, горец водяной перечный Polygonum hydropiper, щучка дернистая Deschampsia cespitosa). Из моллюсков тут встречаются только виды, легко переносящие высыхание, – группа малых прудовиков Galba truncatula и группа катушек Anisus spirorbis. Оба обсуждавшихся выше типа водоемов имеют только паводковое и дождевое питание. Наконец, полупостоянные водоемы, пересыхающие на срок менее одного месяца, в связи с тем, что они дополнительно используют грунтовое питание, покрыты более разнообразной растительностью: рогоз широколистный, ежеголовники, камыш лесной, пушицы, несколько видов влаголюбивых осок, ряски и еще целый ряд видов растений. Кроме немногочисленных обитателей пересыхающих водоемов, здесь представлены и виды, характерные для прибрежных зарослей постоянных водоемов.
Эрозионная деятельность реки определяется высотой ее истоков над базисом эрозии. Эти величины могут быть весьма различны. Так, у р. Днепр высота истоков над базисом эрозии 226 м, тогда как у р. Амударья – свыше 5000 м. У р. Волга эта величина подвержена постоянным изменениям, связанным с многолетними колебаниями уровня Каспия. Эрозионная деятельность реки прекращается, когда высота верховьев над базисом эрозии становится нулевой. Однако такая ситуация возникает крайне редко, поскольку прежде чем упомянутая разница высот станет нулевой, тектонические процессы ее восстановят или как ту же речную систему, или в видоизмененной форме, о чем свидетельствуют данные по кайнозойской истории гидрографической сети.
Наконец, есть еще один ряд очень длительных сукцессионных изменений. Озера, расположенные в рифтовых долинах, с течением времени (за очень большой срок) могут превращаться в части океанов за счет увеличения протяженности рифтовых долин. Какие сукцессионные процессы тут будут происходить, пока неизвестно, но понятно, что если такое рифтовое озеро будет осолоняться за счет испарения (как Мертвое море или озера-предшественники Красного моря), то пресноводный биом такого озера в конце концов превратится в гипергалинный морской. Эта судьба, в частности, уготована через много миллионов лет Байкалу.
Список литературы
Вальтер Г. Растительность земного шара: эколого-физиологическая характеристика. Т. 1. Тропические и субтропические зоны / Ред. П.Б. Виппер. М.: Прогресс, 1968. 552 с. Т. 2. Леса умеренной зоны / Ред. П.А. Работнов. М.: Прогресс, 1974. 434 с. Т. 3. Тундры, луга, степи, внетропические пустыни / Ред. П.А. Работнов. М.: Прогресс, 1975. 429 с.
Логинова Е.В., Лопух П.С. Гидроэкология: курс лекций. Минск: БГУ, 2011. 300 с.
Одум Ю. Основы экологии. М.: Мир, 1975. 740 с.
Уиттекер Р. Сообщества и экосистемы. М.: Прогресс, 1980. 327 с.
Яблоков А.В., Левченко В.Ф., Керженцев А.С. Очерки биосферологии. СПб.: Cвое издательство, 2018. 150 с.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии