

Успехи современной биологии, 2020, T. 140, № 1, стр. 87-104
Совершенствование анализа ареалов и экологических ниш животных (на примере рептилий) с применением компьютерных ГИС-программ
В. А. Черлин *
Дагестанский государственный университет
Махачкала, Россия
* E-mail: cherlin51@mail.ru
Поступила в редакцию 09.04.2019
После доработки 03.08.2019
Принята к публикации 03.08.2019
Аннотация
Рассматриваются компьютерные ГИС-программы, предназначенные для моделирования ареалов животных и их экологических ниш. Материалы статьи на конкретных примерах показывают, что попытки этих программ выявить маркерные характеристики внешней среды и на их основе построить вероятные ареалы для отдельных видов животных малоперспективны. Для того, чтобы эффективность этих программ существенно увеличилась, в них необходимо включить комплекс физиологических требований животных данных видов к среде обитания. После модернизации, эти программы могут стать реально полезными.
ВВЕДЕНИЕ
В свое время среди научных направлений герпетологических исследований экология рептилий занимала одно из ведущих мест. Но затем, особенно в настоящее время, в герпетологии “центр тяжести” исследований сместился в сторону изучения систематики, биохимии, генетики, влияния загрязнения среды и других современных направлений. Экология сохраняет свое значение как важное направление изучения биологии рептилий, но в основном как описательная, “устаревающая” наука. Многими, особенно молодыми исследователями, она начинает рассматриваться, как некий “атавизм”. Конечно, она существует и требует определенного внимания, но недостаточно интересна с точки зрения современной, технологичной, требующей балльной аттестации науки.
Однако, три обстоятельства подталкивают нас не принимать такое положение дел:
1) изучение научного наследия замечательного русского ученого Ивана Дмитриевича Стрельникова, который сформировал научное направление, названное им “экспериментальной экологией”, но которое по сути дела является физиологической экологией (Черлин, 2019); это направление продемонстрировало возможность нового, более совершенного, “продвинутого” подхода к изучению и описанию экологии животных, выявляющего в экологии причинно-следственные связи, механизмы реализации закономерностей, и т.п.; но оно, несмотря на свою очевидную важность и перспективность, к сожалению, не было подхвачено последующими учеными;
2) наши собственные исследования по термобиологии рептилий, которые, как позже выяснилось, являлись именно физиолого-экологическими;
3) наши представления об экологии рептилий и их физиологической экологии не совсем укладываются в те результаты, которые получены при применении компьютерных ГИС11-технологий в изучении их биологии.
В последнее время в мире широко применяются компьютерные ГИС-технологии моделирования ареалов разных видов рептилий и расчета их экологических ниш. Основной подход этих работ заключается в том, что специальные компьютерные программы, анализируя конкретные точки ареалов данных видов рептилий и огромный массив географических, климатических и пр. данных по этим точкам, определяют общие характеристики, присущие ареалу, после чего экстраполируют полученные данные на окружающие территории и строят потенциальный ареал.
Анализируя создавшуюся ситуацию, можно сделать вывод о необходимости пересмотра некоторых основных методологических подходов к экологии пресмыкающихся (а возможно и не только их). Несмотря на то, что эта статья, безусловно, пойдет вразрез с общепринятым научным трендом биологических исследований, в ней предлагаются вниманию читателей некоторые размышления на эту тему.
КАК ФОРМИРУЮТСЯ АРЕАЛЫ ЖИВОТНЫХ РАЗЛИЧНЫХ ВИДОВ?
Для того, чтобы оценить и смоделировать ареалы разных видов живых организмов, необходимо, прежде всего, разобраться в том, как формируются видовые ареалы вообще, и какие факторы на это влияют. Без учета этих обстоятельств никакое моделирование не будет представлять ни научного, ни прикладного интереса.
Важнейшие факторы, влияющие на формирование ареалов:
1. Историко-биогеографический аспект. Каждый вид живых организмов образовался тем или иным способом в процессе филогенеза из некоторой предковой группы. Эта предковая форма обитала в некоем географическом регионе, располагавшемся в определенном месте на карте той исторической эпохи с присущими ему климатическими и другими экологическими условиями. В процессе развития Земли и эволюции групп живых организмов, новый вид распространялся из центра своего образования по допустимым для себя географическим направлениям и “коридорам” и в соответствии с присущими ему морфологическими, физиологическими и пр. эндогенными требованиями к среде обитания.
2. Экологический аспект. Каждый вид живых организмов, сформировавшись в процессе филогенеза, обладает определенным набором эндогенных свойств, которыми он связан со средой своего обитания, то есть обладает вполне конкретными требованиями к экологическим особенностям внешней среды: качество субстрата, травянистой растительности, наличие и качество пространственной многомерности среды (ровные пространства, вертикальные каменные поверхности, кустарники и деревья, возможность устраивать соответствующие убежища, гнезда для периодического суточного отдыха, связанные с размножением, подходящие для эстивации и гибернации), подходящие качественные и количественные характеристики кормовой базы, степень увлажненности субстрата и воздуха, и т.п.
3. Физиолого-экологический аспект. Эти эндогенные требования к среде обитания – очень важная сторона проблемы, без которой невозможно понять как, за счет чего, благодаря чему и по каким закономерностям происходило формирование ареалов в прошлом и происходит в настоящее время.
4. Климатический аспект. Каждый вид живых организмов чувствует себя комфортно и способен полноценно реализовывать все необходимые процессы индивидуального, популяционного и видового жизнеобеспечения лишь в местах с определенными климатическими условиями, прямо или косвенно влияющими на его физиологические процессы. Говоря о климате с физиолого-экологической точки зрения, нужно разделить факторы внешней среды на две части: 1) факторы абиотической среды, определяемые эмпирическим путем (табличные характеристики), важные для биологии животных данного вида: среда обитания – суша, вода; климатические (тип климата, температура воздуха на разной высоте, почвы на разной глубине, сезонный ход температур, осадки, влажность, облачность, инсоляция, продолжительность холодного периода, и т.п.), орографические, эдафические, пространственная структура зоны активности, убежища, и т.п.; 2) факторы абиотической среды расчетные, важные для биологии животных данного вида, для разных географических точек: потенциальные структуры суточной активности в разные сезоны → показатель суммарного количества теплоты за год в разных географических точках → сравнение показателей суммарного количества теплоты за год в разных географических точках (где животные данного вида реально живут на территории своего ареала и где их нет); индексы аридности или наоборот “мягкости” климата, показатель суммарного количества теплоты за год в разных географических точках, проективное покрытие поверхности и т.п.
Есть и другие механизмы формирования ареалов (например, антропологический), но мы здесь о них говорить не будем, потому что это выходит за рамки заявленной для данной статьи темы. Мало того, мы не будем здесь рассматривать отдельно историко-биогеографический и экологический аспекты (пункты 1 и 2). И это не потому, что эти пункты имеют меньшее значение, но просто их анализ – отдельная, специфическая научная работа, которая практически не имеет отношения к компьютерному моделированию, то есть к теме нашей статьи. Может быть в будущем и эти темы будут включены в компьютерный анализ ареалов, но похоже, что время еще пока не пришло…
Необходимо основное внимание обращать на физиолого-экологический и климатический аспекты (пункты 3 и 4), которые в настоящее время уже связаны с компьютерными программами анализа ареалов.
ВАЖНЫЕ ПРОБЛЕМЫ КОМПЬЮТЕРНОГО МОДЕЛИРОВАНИЯ
О физиолого-экологическом взгляде на проблему было уже сказано немного выше. Здесь имеет смысл лишь коротко акцентировать внимание на том, что он безусловно должен объединять внутренние потребности животных с внешними условиями, которые способны или не способны обеспечить удовлетворение внутренних потребностей и реализацию важнейших внутренних циклов жизнеобеспечения через использование факторов внешней среды.
Климатический аспект является одним из наиболее значимых для формирования ареалов эктотермных животных, особенно рептилий. Это происходит потому, что пресмыкающиеся – высшие эктотермные позвоночные, вся биология которых в значительной степени настроена на использование температурного фактора, у которых связь эндогенных особенностей с климатическими (физиолого-экологические связи) проявляется наиболее отчетливо.
Рассмотрим несколько примеров таких связей.
1) Суточная и сезонная динамика температур тела рептилий является для этих животных важнейшим фактором, регулирующим и определяющим режим их жизнедеятельности. Следовательно, возможности организовывать и поддерживать у себя необходимую динамику для этих животных критически важна. Но закономерности связи температуры тела с условиями внешней среды не в эксперименте, а в сложных, разнообразных, мозаичных по факторам среды природных биотопах, оказывается весьма сложной и неоднозначной. Так насколько сами по себе факторы внешней среды, полученные из климатических таблиц и компьютерных баз данных, способны охарактеризовать возможности этих животных поддерживать необходимый уровень температуры тела и формировать ее суточную и сезонную динамику в данном регионе? Другими словами: могут ли рептилии, обладая видоспецифичными особенностями морфологии, физиологии и поведения в данных климатических условиях региона поддерживать необходимую для нормальной жизнедеятельности температуру тела и ее динамику?
Как показывают термобиологические исследования, ни одна температура во внешней среде (ни температура воздуха, ни температура почвы и т.п.), ни их суточная динамика не совпадают полностью с температурой тела рептилий. Это происходит по двум основным причинам: а) температура тела – показатель температурного баланса тела, то есть результат совместного действия ряда тепловых факторов среды, соотношение величины которых очень различно в разных географических точках, в разных биотопах и микробиотопах, при разной погоде, в разное время дня и в разные сезоны года и т.п.; б) большое значение имеют регулирующие поведенческие реакции животных, способные сформировать сильно различающиеся варианты суточной динамики температуры тела в одних и тех же экологических условиях.
При этом, в ряде случаев температура тела некоторых видов рептилий может быть вычислена более или менее определенным образом. В нашей работе с серым вараном (Varanus griseus) измерения показали, что температура тела этих животных в период их активности на поверхности почвы в значительной степени соответствует среднему арифметическому значению температур поверхности почвы и воздуха в зоне активности (Целлариус Е., Целлариус А., 1997). Такую же закономерность отметили и другие исследователи для некоторых ящериц (Литвинов, 2015, и др.).
Но в некоторых случаях большу́ю долю тепла рептилии получают от солнечного излучения. Так, ряд видов ящериц, обитая в горах и оказываясь при температурах воздуха 5–10°С, могут исключительно за счет солнечного излучения поддерживать температуру своего тела от 30–32°С (прыткая ящерица, Lacerta agilis – Стрельников, 1944) до 34–36°С (агама Чернова, Laudakia chernovi – Черлин, Чикин, 1991; Черлин, 2014). Такая же ситуация характерна и для горных насекомых (Стрельников, 1940).
И еще одна ситуация. Крохотная, тоненькая, очень подвижная полосатая ящурка (Eremias scripta) в Каракумах очень сильно зависима от внешних условий. Но при этом она то недолго лежит на поверхности песка на открытом солнце, то перебегает в тень, то “взлетает” на нижние ветки кустов или, стремительно забравшись на вершину песчаной акации (Ammodendron), буквально “перепархивает” на соседнее дерево, а потом снова спускается на почву и бегает по горячему песку. В какие-то моменты, когда она лежит на песке, температура ее тела максимально зависит именно от нее, но когда она взбирается на ветки – температура ее тела больше всего связана с температурой воздуха. Она одна из самых термофильных ящурок (Черлин, 1988). Какая температура может считаться “маркерной” для ее обитания, если она за три минуты оказывалась в разных микроклиматических условиях? Мало того, на этой же территории вместе с ней живет и мезофильная средняя ящурка (Eremias intermedia). Температура тела у нее при активности заметно ниже, чем у полосатой ящурки. Она также очень зависима от внешних тепловых воздействий, но, в отличие от полосатой ящурки, она постоянно выбирает такие теневые места и такое время суток, где и когда она может поддерживать более низкую температуру тела, а в наиболее жаркое время она вообще переходит на жизнь в норах песчанки (Черлин, 2019). Какой климатический показатель может характеризовать место ее обитания, и будет ли он отличаться от показателя для полосатой ящурки? А если учесть, что ареалы этих ящурок во многом пересекаются и с ареалом сугубо ночного мезофила сцинкового геккона (Teratoscincus scincus), температура тела которого намного ниже, чем у ящурок, то ситуация еще больше осложнится.
Таким образом, учет механизмов формирования теплового баланса тела для удовлетворения эндогенных потребностей организма может помочь в описании “разрешающих” условий активности рептилий определенных видов, но вряд ли этому может способствовать даже компьютерный анализ климатических и пр. характеристик на данных территориях без учета физиологических потребностей животных.
В разных условиях внешней среды зависимости температуры тела от внешних температур оказываются разными. Следовательно, маркерного показателя условий внешней среды, способного определить возможную температуру тела рептилии или хотя бы определенно ее охарактеризовать, то есть показателя, пригодного к использованию в компьютерной программе моделирования, предложить просто невозможно.
2) Для того, чтобы эктотермное животное могло поддерживать необходимую динамику температуры тела, оно должно с помощью разнообразных поведенческих приемов взаимодействовать с тепловыми факторами внешней среды. Одним из аспектов этого взаимодействия является формирование определенных диапазонов внешних факторов, в рамках которых животные, применяя специфические для своего вида морфологические особенности, поведенческие и физиологические реакции, могут организовывать эту необходимую для себя динамику температуры тела.
Физиолого-экологические исследования по термобиологии рептилий показали, что стабильный, видоспецифичный комплекс термофизиологических показателей в значительной степени регулирует и определяет режим суточной и сезонной активности этих животных, их биотопическое распределение и географическое распространение (рис. 1, 2; Черлин, 2014).
Рис. 1.
Схема расчета продолжительности периода полной активности серого варана по метеоданным (по Целлариус, Целлариус, 1997). По оси ординат (T) – температура в °С, по оси абсцисс – время суток. Минимальная температура тела серых варанов при полной активности – 33°С, максимальная – 41.5°С; Tп – температура поверхности песка, Tв – температура приземного слоя воздуха, Tр – рассчитанная температура тела, полученная из полевых данных по времени как средняя между температурами поверхности почвы и воздуха на высоте 5 см на метеоплощадке.
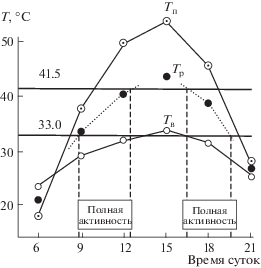
Рис. 2.
Динамика температур среды и тела ящериц в Каракумах в летний период и использование разных микроклиматических параметров для поддержания необходимой температуры тела (Черлин, 2015). Т – температура. (а) – полосатая ящурка, Eremias scripta (диапазон температур тела при полной активности примерно 39–43°С – Черлин, 1988); (б) – песчаная круглоголовка, Phrynocephalus interscapularis (диапазон температуры тела при полной активности примерно 38–44°С – Черлин, Музыченко, 1983). Общие формы поведения ящериц: 1 – ночной сон, 2 – нагревание, 3 – термостабилизирующее поведение, 4 – остывание, 5 – добровольный перегрев, 6 – дневной отдых. Температуры: 7 – тела ящериц; 8 – поверхности почвы на открытом месте; 9 – воздуха в приземном слое (на высоте 2 см) на открытом месте; 10 – воздуха в кроне дерева на высоте 2 м; 11 – поверхности почвы в тени кустов; 12 – почвы на глубине 5 см; 13 – почвы на глубине 10 см; 14 – почвы на глубине 15 см.

Серые вараны (рис. 1) могут быть активны на поверхности только в периоды времени, когда температура их тела, закономерно завися от температур поверхности почвы и воздуха, может находиться примерно в диапазоне между 33.0 и 41.5°С (Целлариус Е., Целлариус А., 1997).
Такая же закономерность наблюдается и на рис. 2. Полосатая ящурка (Eremias scripta) и песчаная круглоголовка (Phrynocephalus interscapularis) занимают разные микробиотопы в одном и том же регионе и имеют различную пространственную структуру активности (Черлин, 1988). Для необходимой обеим ящерицам динамики температуры тела и с учетом различных поведенческих видовых особенностей они используют разные тепловые факторы среды: ящурки – в основном температуру поверхности почвы на солнце и в тени, температуры воздуха в кронах кустов и деревьев на разных высотах; круглоголовки – только температуру поверхности почвы на солнце и в тени и температуру воздуха на высоте около 1 см над почвой. Рисунок 2 иллюстрирует, что динамика всех этих температур определяет то, в какое время и где могут находиться эти ящерицы.
Также этот комплекс определяет и режимы протекания сезонных физиологических циклов питания и размножения у среднеазиатских эф (Echis multisquamatus) (рис. 3, 4; Черлин, Целлариус, 1981).
Рис. 3.
Микроклиматические условия, определяющие сроки протекания половой активности у среднеазиатской эфы (по Черлин, Целлариус, 1981). N – частота встреч эф (в % от общего числа встреч), T – температура почвы на глубине 20 см; 1 – температура почвы в норах, при которой эфы могут свободно передвигаться по поверхности; 2 – температура почвы в норах, при которой может протекать гаметогенез; 3 – частота встреч эф на поверхности в светлое время суток в зависимости от температуры почвы в норах (в % от общего числа встреч); 4 – интервал температур почвы в норах, при котором возможна половая активность.

Рис. 4.
Микроклиматические условия, определяющие время протекания пищевой активности у среднеазиатской эфы (по Черлин, Целлариус, 1981). T – температура почвы на глубине 20 см; 1 – температура почвы в норах, при которой возможно переваривание пищи; 2 – температура почвы в норах, при которой возможна охота; 3 – температура почвы в норах после завершения половой активности; а – интервал температур почвы в норах, при которых возможно питание не размножавшихся в данный сезон особей, б – интервал температур почвы в норах, при котором возможно питание размножавшихся в данный сезон особей.

Эти рисунки иллюстрируют, как зависят сезонные циклы питания и размножения от годичных изменений температурных условий среды. И в том, и в другом случае зависимость определяется по отношению к температурам почвы в норах. И этому есть две основных причины:
а) дело в том, что температура поверхности почвы, особенно весной и осенью, очень изменчива, поскольку погода в это время неустойчива. А температуры в глубине почвы, и, следовательно, и в норах (на глубине около 20 см, то есть там, где в основном держатся змеи) не реагируют на быстрые изменения погодных условий. Они демонстрируют плавные сезонные изменения, характеризующие медленные, климатические, сезонные изменения, закономерную смену сезонов года, то есть именно температура почвы в норах (температура почвы на глубине 20 см) может являться неким показателем сезонной фазы состояния климата и фаз физиологических циклов;
б) но, кроме этого, температурные условия в норах влияют на сезонные физиологические циклы непосредственно. Именно температура в норах определяет диапазон суточных колебаний температуры тела змей, который чрезвычайно важен в реализации разных физиологических циклов. Это связано с тем, что верхний уровень температуры тела у эф, как и у всех других видов рептилий, всегда и во всех условиях видоспецифично одинаков (для эф – 33°С), о чем заботятся их системы терморегуляции (Черлин, 2014). А вот нижние границы температуры тела определяются доступными в природных условиях уровнями температур. Чаще всего этот уровень определяется условиями ночевок, то есть для эф – температурами почвы в норах. В весенний период уровень температуры почвы в норах еще достаточно низок (от 10°С и выше). Значит, весной амплитуда суточных колебаний температуры тела эф составляет примерно 23°С и с течением сезона постепенно уменьшается. Кроме этого, именно когда температура почвы в норах находится в интервале примерно 12–24°С, то есть в первой половине весны, температурные условия складываются так, что у эф есть возможность быть активными в светлое время суток, то есть для них в это время сильно возрастает период световой фазы, интенсивность поступления ультрафиолетового излучения при активности и увеличивается относительная влажность воздуха. А для реализации полового цикла (рис. 3) для большинства рептилий как раз и нужно, чтобы весной в течение суток у них увеличивался световой день, ультрафиолетовое излучение и влажность воздуха, и был период с пониженными температурами тела для правильного течения гаметогенеза, для запуска половой активности, и т.п. Ближе к лету, наоборот, температура почвы в норах повышается примерно до 30°С и даже иногда немного выше. Это уменьшает амплитуду суточных колебаний температуры тела эф почти до 0°С, что змеям важно для успешного и максимально быстрого течения восстановительного периода у самцов и для успешного окончания беременности у самок. Для реализации пищевого цикла важно, чтобы климатические условия позволяли эфам иметь достаточно времени, для охоты (рис. 4, п. 2), чтобы закончился период половой активности, в течение которого эфы не питаются (рис. 4, п. 3), и чтобы они могли успешно переваривать пищу, что возможно только когда суточная амплитуда колебаний температуры тела не превышает 10–12°С (то есть когда температура почвы в норах становится выше 20°С; рис. 4, п. 1).
Следовательно, именно взаимодействие видоспецифичных эндогенных характеристик термального гомеостаза и внешних температурных условий формируют особые условия для успешного течения сезонных физиологических циклов. И если какой-то параметр внешней среды теоретически мог бы являться маркером возможного обитания некоторого вида рептилий на данной территории (для эф – это динамика температуры почвы на глубине 20 см), то это характерно только для данного вида рептилий и только на данной территории с определенными характеристиками почвы, наличием нор песчанок и др. сопутствующими обстоятельствами. И принимать во внимание его необходимо определенным образом – с учетом физиологических условий, требований и ограничений для данного вида. Никакая компьютерная программа не в состоянии все это учесть и вычислить, только исходя из анализа огромного набора, по сути, случайных характеристик точек на карте ареала данного вида. Важные для данного вида параметры должны быть заложены человеком, который для этого должен провести предварительные, специальные экологические, физиологические и физиолого-экологические исследования.
3) Одной из важных характеристик, определяющих способность обитания рептилий данного вида на некоторых территориях, является возможность набрать за период активности определенное количество тепловой энергии, выраженное, к примеру, в днях-градусах, или по нашей методике – в часах-градусах (Черлин, 2014). Это количество тепловой энергии нужно животным данного вида для того, чтобы начать и полностью завершить такие важнейшие для его жизнедеятельности сезонные физиологические циклы, как обеспечение энергией, питание, размножение. Мы применили сравнительный показатель количества полученной животным тепловой энергии в часах-градусах. В целом – это температура тела животного, умноженная на количество часов, которые животное проводит при данной температуре тела.
Необходимо понимать, что, строго говоря, произведение температуры тела животного на количество часов, которые оно проводит при данной температуре тела, по сути – не количество энергии, а лишь некий показатель этого количества. Но если во всех случаях вычисления производить по единой стандартной методике, то этот показатель становится вполне приемлемым для сравнения количества тепловой энергии, полученной животными разных видов в различных географических зонах, в разном физиологическом состоянии и т.п.
Расчет сравнительного показателя суммы тепловой энергии, полученной рептилией за год (ΣTe), достаточно сложен (если мы, конечно, хотим, чтобы эта величина была не абстрактной, а действительно приближенной к реальной). Мы предложили вариант такого расчета (Черлин, 2014). Вычисление этой характеристики должно учитывать необходимую суточную динамику температуры тела, тепловой баланс тела в данных условиях, динамику факторов внешней среды, расчетный возможный режим активности, количество солнечных дней за сезонный период активности животных в данной географической точке, температуру в убежищах и ее сезонную динамику, другие климатические характеристики региона обитания, и ряд других показателей, которые должны иметь значение именно для данного вида животных в данных экологических, географических и климатических условиях.
Вначале необходимо вычислить сравнительный показатель количества тепловой энергии, полученной животным данного вида (Те) за сутки. Наше основное исходное положение заключается в том, что сутки должны быть разделены на отдельные отрезки времени, при которых температура тела животного примерно одинаковая. В каждом таком отрезке времени мы вычисляем Те (температура тела умноженная на длительность отрезка), после чего Те всех отрезков за сутки суммируем.
Год делится для животного на разные периоды – сезоны. В целом это периоды активности и покоя (эстивация, гибернация и т.п.). Получив информацию об образе жизни животных в данном географическом регионе (из литературы, из собственных экологических наблюдений, рассчитанные по определенной физиолого-экологической методике), можно разделить все сезоны на отдельные более или менее однородные периоды для данного вида животных в данной географической точке. После этого можно рассчитать Те для каждого такого периода, а потом уже и ΣTe за весь год.
Методика не простая, но вполне понятная и реализуемая. Приведем несколько примеров таких расчетов (табл. 1–5).
Таблица 1
-1. Расчет ΣTe за год для обыкновенной гадюки Vipera berus (Якутия, Оленминский район)2
| Месяц | Ясные дни | |||||||||||
| на поверхности | в сумерках | в убежище | за день | кол. дней | за мес. | |||||||
| кол. часов | Т тела | ΣTe | кол. часов | Т тела | ΣTe | кол. часов | Т тела | ΣTe | ΣTe | ΣTe | ||
| I | – | – | – | – | – | – | – | – | – | – | – | – |
| II | – | – | – | – | – | – | – | – | – | – | – | – |
| III | – | – | – | – | – | – | – | – | – | – | – | – |
| IV | – | – | – | – | – | – | – | – | – | – | – | – |
| V | 6 | 28 | 168 | – | – | – | 18 | 5 | 90 | 258 | 19 | 4902 |
| VI | 9 | 28 | 252 | 3 | 18 | 54 | 12 | 12 | 144 | 450 | 17 | 7650 |
| VII | 9 | 28 | 252 | 3 | 18 | 54 | 12 | 12 | 144 | 450 | 11 | 4950 |
| VIII | 6 | 28 | 168 | – | – | – | 18 | 5 | 90 | 258 | 17 | 4384 |
| IX | – | – | – | – | – | – | – | – | – | – | – | – |
| X | – | – | – | – | – | – | – | – | – | – | – | – |
| XI | – | – | – | – | – | – | – | – | – | – | – | – |
| XII | – | – | – | – | – | – | – | – | – | – | – | – |
| Месяц | Пасмурные дни | В холодовом оцепенении | За мес. | |||||||||
| в убежище | кол. дней | за мес. | в убежище | кол. дней | за мес. | ΣTe | ||||||
| кол. часов | Т тела | ΣTe | ΣTe | кол. часов | Т тела | ΣTe | ΣTe | |||||
| I | – | – | – | – | – | 24 | 2 | 48 | 31 | 1488 | 1488 | |
| II | – | – | – | – | – | 24 | 2 | 48 | 28 | 344 | 1344 | |
| III | – | – | – | – | – | 24 | 1 | 24 | 31 | 744 | 744 | |
| IV | – | – | – | – | – | 24 | 3 | 72 | 30 | 2160 | 2160 | |
| V | 24 | 5 | 120 | 12 | 1440 | – | – | – | – | – | – | |
| VI | 24 | 12 | 288 | 13 | 3744 | – | – | – | – | – | – | |
| VII | 24 | 12 | 288 | 20 | 5760 | – | – | – | – | – | – | |
| VIII | 24 | 5 | 120 | 13 | 1560 | – | – | – | – | – | – | |
| IX | – | – | – | – | – | 24 | 3 | 72 | 30 | 2160 | 2160 | |
| X | – | – | – | – | – | 24 | 2 | 48 | 31 | 1488 | 1488 | |
| XI | – | – | – | – | – | 24 | 2 | 48 | 30 | 1440 | 1440 | |
| XII | – | – | – | – | – | 24 | 1 | 24 | 31 | 744 | 744 | |
| ΣTe за год | 45 960 | |||||||||||
| ΣTe за активный период | 34 392 | |||||||||||
| Месяц | Ясные дни | |||||||||||
| на поверхности | в убежище | за день | кол. дней | за мес. | ||||||||
| кол. часов | Т тела | ΣTe | кол. часов | Т тела | ΣTe | ΣTe | ΣTe | ΣTe | ||||
| I | – | – | – | – | – | – | – | – | – | |||
| II | – | – | – | – | – | – | – | – | – | |||
| III | – | – | – | – | – | – | – | – | – | |||
| IV | 4 | 29 | 116 | 20 | 2 | 40 | 156 | 18 | 2808 | |||
| V | 5 | 29 | 145 | 19 | 5 | 95 | 240 | 21 | 5040 | |||
| VI | 6 | 30 | 180 | 18 | 10 | 180 | 360 | 21 | 7560 | |||
| VII | 6 | 30 | 180 | 18 | 12.5 | 225 | 405 | 20 | 8100 | |||
| VIII | 6 | 30 | 180 | 18 | 13.5 | 243 | 423 | 20 | 8460 | |||
| IX | 5 | 29 | 145 | 19 | 12 | 228 | 373 | 20 | 7460 | |||
| X | 3 | 28 | 84 | 21 | 9 | 189 | 273 | 8 | 4914 | |||
| XI | – | – | – | – | – | – | – | – | – | |||
| XII | – | – | – | – | – | – | – | – | – | |||
| Месяц | Пасмурные дни | В холодовом оцепенении | За мес. | |||||||||
| в убежище | кол. дней | за мес. | в убежище | кол. дней | за мес. | ΣTe | ||||||
| кол. часов | Т тела | ΣTe | кол. часов | Т тела | ΣTe | Т тела | ΣTe | ΣTe | ||||
| I | – | – | – | – | – | 24 | 4 | 96 | 31 | 2976 | 2976 | |
| II | – | – | – | – | – | 24 | 3 | 72 | 28 | 2016 | 2016 | |
| III | – | – | – | – | – | 24 | 2 | 48 | 31 | 1488 | 1488 | |
| IV | 24 | 2 | 48 | 12 | 576 | – | – | – | – | – | – | |
| V | 244 | 5 | 120 | 10 | 1200 | – | – | – | – | – | – | |
| VI | 24 | 10 | 240 | 9 | 2160 | – | – | – | – | – | – | |
| VII | 24 | 12.5 | 300 | 11 | 3300 | – | – | – | – | – | – | |
| VIII | 24 | 13.5 | 324 | 11 | 3564 | – | – | – | – | – | – | |
| IX | 24 | 12 | 288 | 10 | 2880 | – | – | – | – | – | – | |
| X | 24 | 9 | 216 | 16 | 3456 | – | – | – | – | – | – | |
| XI | – | – | – | – | – | 24 | 6 | 144 | 30 | 4320 | 4320 | |
| XII | – | – | – | – | – | 24 | 4 | 96 | 31 | 2976 | 2976 | |
| ΣTe за год | 75 578 | |||||||||||
| ΣTe за активный период | 61 802 | |||||||||||
Примечание: здесь и в табл. 2–13 прочерки означают, что необходимых для заполнения ячеек данных в печатных источниках не найдено. 2Источник таблиц 1–5 – Черлин, 2014.
Таблица 2.
Расчет ΣTe для армянской гадюки Montivipera raddei (Ереван)
| Месяц | Ясные дни | ||||||||||||||
| утром на поверхности | днем в убежище | вечером на поверхности | ночью в убежище | за день | кол. дней | за мес. | |||||||||
| кол. часов | Т тела | ΣTe | кол. часов | Т тела | ΣTe | кол. часов | Т тела | ΣTe | кол. часов | Т тела | ΣTe | ΣTe | ΣTe | ||
| I | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| II | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| III | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| IV | 6 | 28 | 168 | – | – | – | – | – | – | 28 | 16 | 288 | 456 | 24 | 10 944 |
| V | 7 | 28 | 196 | – | – | – | – | – | – | 17 | 21 | 357 | 553 | 26 | 14 378 |
| VI | 3 | 30 | 90 | 5 | 25 | 125 | 3 | 28 | 84 | 13 | 25 | 325 | 624 | 26 | 16 224 |
| VII | 3 | 30 | 90 | 5 | 29 | 145 | 3 | 29 | 87 | 13 | 29 | 377 | 699 | 28 | 19 572 |
| VIII | 5 | 30 | 150 | 4 | 30 | 120 | 2 | 29 | 58 | 13 | 30 | 390 | 718 | 29 | 20 822 |
| IX | 6 | 30 | 180 | – | – | – | – | – | – | 18 | 25 | 450 | 630 | 28 | 17 640 |
| X | 6 | 28 | 168 | – | – | – | – | – | – | 18 | 21 | 378 | 546 | 27 | 14 742 |
| XI | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| XII | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| Месяц | Пасмурные дни | В холодовом оцепенении | За мес. | ||||||||||||
| в убежище | кол. дней | за мес. | в убежище | кол. дней | за мес. | ΣTe | |||||||||
| кол. часов | Т тела | ΣTe | ΣTe | кол. часов | Т тела | ΣTe | ΣTe | ||||||||
| I | – | – | – | – | – | 24 | 6 | 144 | 31 | 4464 | 4464 | ||||
| II | – | – | – | – | – | 24 | 6 | 144 | 28 | 4032 | 4032 | ||||
| III | – | – | – | – | – | 24 | 8 | 192 | 31 | 5952 | 5952 | ||||
| IV | 24 | 16 | 34 | 6 | 2304 | – | – | – | – | – | 13 248 | ||||
| V | 24 | 21 | 504 | 5 | 2520 | – | – | – | – | – | 16 898 | ||||
| VI | 24 | 25 | 600 | 4 | 2400 | – | – | – | – | – | 18 624 | ||||
| VII | 24 | 29 | 696 | 3 | 2088 | – | – | – | – | – | 21 660 | ||||
| VIII | 24 | 30 | 720 | 2 | 1440 | – | – | – | – | – | 22 262 | ||||
| IX | 24 | 25 | 600 | 2 | 1200 | – | – | – | – | – | 18 840 | ||||
| X | 24 | 21 | 504 | 4 | 1216 | – | – | – | – | – | 16 758 | ||||
| XI | – | – | – | – | – | 24 | 15 | 360 | 30 | 10 800 | 10 800 | ||||
| XII | – | – | – | – | – | 24 | 9 | 216 | 31 | 6696 | 6696 | ||||
| ΣTe за год | 160 234 | ||||||||||||||
| ΣTe за активный период | 128 290 | ||||||||||||||
Таблица 3.
Расчет ΣTe для среднеазиатской гюрзы Macrovipera lebetina turanica (Нурата, Узбекистан)
| Месяц | Ясные дни | ||||||||||||||
| днем на солнце при терминге3 | вечером без солнца | в убежище | ночевки на поверхности | за день | кол. дней | за мес. | |||||||||
| кол. часов | Т тела | ΣTe | кол. часов | Т тела | ΣTe | кол. часов | Т тела | ΣTe | кол. часов | Т тела | ΣTe | ΣTe | ΣTe | ||
| I | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| II | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| III | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| IV | 8 | 29 | 232 | – | – | – | – | – | – | 6 | 13 | 78 | 480 | 19 | 9120 |
| V | 8 | 30 | 240 | 4 | 22 | 88 | 4 | 20 | 80 | 8 | 20 | 160 | 480 | 26 | 12 480 |
| VI | 10 | 30 | 300 | 4 | 26 | 104 | – | – | 54 | 10 | 23 | 230 | 634 | 29 | 18 386 |
| VII | 10 | 30 | 300 | 4 | 26 | 104 | – | – | 345 | 10 | 23 | 230 | 634 | 31 | 19 654 |
| VIII | 10 | 30 | 300 | 4 | 25 | 100 | 2 | 27 | 306 | 8 | 20 | 160 | 614 | 31 | 19 034 |
| IX | 9 | 30 | 270 | – | – | – | 15 | 23 | – | – | – | – | 615 | 29 | 17 835 |
| X | 6 | 29 | 174 | – | – | – | 18 | 17 | – | – | – | – | 480 | 26 | 12 480 |
| XI | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| XII | – | – | – | – | – | – | – | – | – | – | – | – | – | – | – |
| Месяц | Пасмурные дни | ||||||||||||||
| днем на поверхности | в убежище | ночевки на поверхности | за день | кол. дней | за мес. | ||||||||||
| кол. часов | Т тела | ΣTe | кол. часов | Т тела | ΣTe | кол. часов | Т тела | ΣTe | ΣTe | ΣTe | |||||
| I | – | – | – | – | – | – | – | – | – | – | – | – | |||
| II | – | – | – | – | – | – | – | – | – | – | – | – | |||
| III | – | – | – | – | – | – | – | – | – | – | – | – | |||
| IV | 6 | 20 | 120 | 18 | 17 | 306 | – | – | – | 426 | 11 | 4686 | |||
| V | 10 | 25 | 250 | 10 | 20 | 200 | 4 | 17 | 686 | 518 | 5 | 2590 | |||
| VI | 10 | 25 | 250 | – | – | – | 14 | 22 | 308 | 558 | 1 | 558 | |||
| VII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| IX | – | – | – | 24 | 23 | 552 | – | – | – | 552 | 1 | 552 | |||
| X | – | – | – | 24 | 17 | 408 | – | – | – | 408 | 5 | 2040 | |||
| XI | – | – | – | – | – | – | – | – | – | – | – | – | |||
| XII | – | – | – | – | – | – | – | – | – | – | – | – | |||
| Месяц | В холодовом оцепенении | За мес. | |||||||||||||
| в убежище | кол. дней | за мес. | ΣTe | ||||||||||||
| кол. часов | Т тела | ΣTe | ΣTe | ||||||||||||
| I | 24 | 9 | 216 | 31 | 6696 | 6696 | |||||||||
| II | 24 | 8 | 192 | 28 | 5376 | 5376 | |||||||||
| III | 24 | 12 | 228 | 31 | 8928 | 8928 | |||||||||
| IV | – | – | – | – | – | 13 806 | |||||||||
| V | – | – | – | – | – | 15 070 | |||||||||
| VI | – | – | – | – | – | 18 944 | |||||||||
| VII | – | – | – | – | – | 19 654 | |||||||||
| VIII | – | – | – | – | – | 19 034 | |||||||||
| IX | – | – | – | – | – | 18 387 | |||||||||
| X | – | – | – | – | – | 14 520 | |||||||||
| XI | 24 | 15 | 360 | 30 | 10 800 | 10 800 | |||||||||
| XII | 24 | 10 | 240 | 31 | 7440 | 7440 | |||||||||
| ΣTe за год | 158 655 | ||||||||||||||
| ΣTe за активный период | 119 415 | ||||||||||||||
3 Терминг – форма активности рептилий, при которой они греются, крайне редко отвлекаясь на другие формы активности (питание, социальные контакты и т.п.), и удерживают температуру тела примерно на одном, достаточно высоком, предпочитаемом уровне, то есть реально заняты термостабилизацией. Терминг наблюдается не у всех рептилий, а только у термонейтрально активных – у гаттерии, гекконовых ящериц, змей, крокодилов и черепах (Черлин, 2014).
Таблица 4.
Расчет ΣTe для среднеазиатской эфы Echis multisquamatus (Кушка, Туркменистан)
| Месяц | Ясные дни | ||||||||||||||
| днем на солнце при терминге | вечером без солнца | в убежище | за день | кол. дней | за мес. | ||||||||||
| кол. часов | Т тела | ΣTe | кол. часов | Т тела | ΣTe | кол. часов | Т тела | ΣTe | ΣTe | ΣTe | |||||
| I | – | – | – | – | – | – | – | – | – | – | – | – | |||
| II | 2 | 30 | 60 | – | – | – | 22 | 10 | 220 | 280 | 17 | 4760 | |||
| III | 4 | 30 | 120 | – | – | – | 20 | 14 | 280 | 400 | 18 | 7200 | |||
| IV | 9 | 30 | 270 | – | – | – | 15 | 19 | 285 | 555 | 19 | 19 545 | |||
| V | 10.5 | 30 | 315 | – | – | – | 13.5 | 26 | 351 | 666 | 28 | 18 648 | |||
| VI | 4 | 30 | 120 | 4 | 27 | 108 | 16 | 29 | 464 | 692 | 30 | 20 760 | |||
| VII | 1 | 30 | 30 | 6 | 29 | 174 | 17 | 331 | 527 | 731 | 31 | 22 661 | |||
| VIII | 1 | 30 | 30 | 6 | 31 | 186 | 17 | 32 | 544 | 760 | 31 | 23 560 | |||
| IX | 6 | 30 | 180 | – | – | – | 18 | 25 | 450 | 630 | 30 | 8900 | |||
| X | 3 | 30 | 90 | – | – | – | 21 | 18 | 378 | 468 | 29 | 13 572 | |||
| XI | – | – | – | – | – | – | – | – | – | – | – | – | |||
| XII | – | – | – | – | – | – | – | – | – | – | – | – | |||
| Месяц | Пасмурные дни | В холодовом оцепенении | за мес. | ||||||||||||
| днем на поверхности | в убежище | за день | кол. дней | за мес. | в убежище | кол. дней | за мес. | ΣTe | |||||||
| кол. часов | Т тела | ΣTe | кол. дней | Т тела | ΣTe | ΣTe | ΣTe | кол. часов | Т тела | ΣTe | ΣTe | ||||
| I | – | – | – | – | – | – | – | – | – | 24 | 9 | 216 | 31 | 6696 | 6696 |
| II | – | – | – | 24 | 10 | 240 | 240 | 11 | 2640 | – | – | – | – | – | 7400 |
| III | 4 | 14 | 56 | 20 | 14 | 280 | 336 | 13 | 4368 | – | – | – | – | – | 11 568 |
| IV | 4 | 19 | 76 | 20 | 19 | 380 | 456 | 11 | 5016 | – | – | – | – | – | 15 561 |
| V | 5 | 25 | 125 | 19 | 26 | 494 | 619 | 3 | 1857 | – | – | – | – | – | 20 505 |
| VI | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | – | – | – | – | – | 20 760 |
| VII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | – | – | – | – | – | 2661 |
| VIII | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | – | – | – | – | – | 23 460 |
| IX | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | – | – | – | – | – | 18 900 |
| X | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 864 | – | – | – | – | – | 14 436 |
| XI | – | – | – | – | – | – | – | – | – | 24 | 15 | 360 | 30 | 10 800 | 10 800 |
| XII | – | – | – | – | – | – | – | – | – | 24 | 10 | 240 | 31 | 7440 | 7440 |
| ΣTe за год | 180 287 | ||||||||||||||
| ΣTe за активный период | 155 351 | ||||||||||||||
Таблица 5.
Расчет ΣTe для некоторых видов рептилий за год (средние по нескольким регионам)
| Вид рептилий | Min | Max | Среднее | % | |
|---|---|---|---|---|---|
| За весь год | |||||
| Среднеазиатская эфа | 179 697 | 180 969 | 180 213 | 324 | 114 |
| Среднеазиатская гюрза | – | – | 158 655 | 285 | 100 |
| Армянская гадюка | – | – | 160 234 | 288 | 101 |
| Обыкновенная гадюка | 45 960 | 85 137 | 69 458 | 100 | 44 |
| Тропические удавы* | – | – | 240 900 | 347 | 152 |
| Только за период активности (без зимовки) | |||||
| Среднеазиатская эфа | 148 041 | 156 033 | 151 245 | 272 | 127 |
| Среднеазиатская гюрза | – | – | 119 415 | 215 | 100 |
| Армянская гадюка | – | – | 128 290 | 231 | 107 |
| Обыкновенная гадюка | 34 392 | 71 052 | 55 649 | 100 | 47 |
| Тропические удавы* | – | – | 240 900 | 433 | 202 |
Эти зависимости и их расчеты еще намного сложнее, чем в пункте 1, и простой анализ только по базам климатических данных выявить реальные закономерности, влияющие на географическое распространение животных, не в состоянии.
4) Один из важных моментов, регулирующих географическое распространение разных видов рептилий в северной и умеренной зонах – условия зимовки. Так, рассмотрим для примера распространение ушастой круглоголовки на юге Российской Федерации. Зимовки располагаются у этого вида в песчаной почве на глубине около 1–1.5 м. Температура в холодное время года (с декабря по начало апреля) на этой глубине не должна опускаться ниже примерно от 6–8°С, что становится опасным для жизни ящериц. Это те условия гибернации, которые позволяют успешно протекать сезонному циклу размножения у ящериц, адекватно функционировать семенникам и запускают половую активность. Компьютерный анализ ареала для именно этого вида ящериц, если данный показатель физиологических потребностей животных заранее не заложить в программу, не сможет его выделить и учесть, как один из важнейших. Без учета данного показателя анализ климатических данных отодвинет границу ареала круглоголовки намного севернее, чем это есть на самом деле.
Таблица 6 показывает, что в регионах, расположенных ниже темно-серой разделительной полосы (Астрахань, Махачкала, равнинные Туркменистан и Узбекистан), в зимовках ушастых круглоголовок с января по начало апреля температура не опускается ниже примерно 6°С, а в ноябре–декабре она вообще находится в диапазоне 14–19°С. В более северных регионах (Саратов, Оренбург, Волгоград, Каракалпакия, Ставрополь, Краснодар), расположенных в таблице выше серой разделительной полосы, температура зимовок часто оказывается в опасном и даже смертельном диапазоне – примерно от 2–5°С.
Таблица 6.
Температура почвы на глубине расположения зимовальных камер ушастых круглоголовок (примерно 1.6 м)4
| Пункт | Месяц | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |
| Саратов | 3.7 | 2.7 | 2.2 | 2.4 | 5.3 | 8.8 | 11.4 | 12.9 | 12.9 | 11.0 | 8.3 | 5.7 |
| Оренбург | 4.1 | 2.6 | 1.9 | 2.2 | 4.9 | 8.0 | 10.7 | 12.4 | 12.6 | 11.2 | 8.8 | 6.0 |
| Волгоград | 4.3 | 2.9 | 2.6 | 4.9 | 10.2 | 15.2 | 18.8 | 20.4 | 19.2 | 15.6 | 11.1 | 6.8 |
| Каракалпакия | 6.6 | 5.0 | 4.2 | 5.2 | 8.5 | 11.9 | 15.0 | 17.1 | 17.5 | 15.8 | 12.8 | 9.5 |
| Ставрополь | 5.0 | 4.0 | 3.8 | 5.3 | 8.8 | 12.2 | 14.4 | 15.7 | 15.1 | 12.7 | 9.7 | 6.8 |
| Краснодар | 6.8 | 6.6 | 5.5 | 8.0 | 12.3 | 16.3 | 19.5 | 21.2 | 20.2 | 16.9 | 13.0 | 9.1 |
| Астрахань | 8.3 | 6.9 | 6.4 | 7.9 | 11.3 | 14.8 | 17.8 | 19.7 | 19.6 | 17.2 | 14.0 | 10.6 |
| Махачкала | 13.6 | 12.2 | 11.7 | 12.6 | 15.4 | 18.6 | 21.4 | 23.2 | 21.3 | 21.4 | 18.7 | 15.6 |
| Самарканд | 9.0 | 8.0 | 8.6 | 10.8 | 14.4 | 18.2 | 21.2 | 22.6 | 22.0 | 19.5 | 15.6 | 11.7 |
| Ашхабад | 12.7 | 11.7 | 12.2 | 14.1 | 17.5 | 21.3 | 24.3 | 25.9 | 25.3 | 23.0 | 19.4 | 15.5 |
Примечание: в регионах, находящихся в таблицах 6–9, 11 и 13 выше темно-серой полосы, ушастые круглоголовки не отмечены, ниже нее – встречаются. 4Все климатические данные в таблицах №№ 6–12 взяты из литературных источников (Климат Самарканда, 1983; Климат Ашхабада, 1984; Климат Баку, 1988; Научно-прикладной справочник по климату СССР… Вып. 12, 1988; Научно-прикладной справочник по климату СССР… Вып. 30, 1989; Научно-прикладной справочник по климату СССР… Вып. 19, 1989; Климат Краснодара, 1990; Научно-прикладной справочник по климату СССР… Вып. 13, 1990; Научно-прикладной справочник по климату СССР… Вып. 15, 1990; Научно-прикладной справочник по климату СССР… Вып. 28, 1990; и др.).
Средняя месячная температура поверхности почвы (табл. 7) в июне–августе в районах, где живут ушастые круглоголовки, обычно выше 28–29°С, а средние максимальные температуры поверхности почвы (табл. 8) в то же время и в тех же регионах оказываются выше 56–60°С. В некоторых южных регионах России температурные условия почти соответствуют этим требованиям, но обитание этих ящериц в указанных регионах блокируется другими обстоятельствами – например, условиями зимовок (табл. 6) и др.
Таблица 7.
Средняя месячная температура поверхности почвы5
| Пункт | Месяц | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |
| Саратов | –11 | –12 | –5 | 7 | 19 | 25 | 27 | 24 | 15 | 5 | –2 | –7 |
| Оренбург | –15 | –15 | –7 | 7 | 20 | 25 | 27 | 24 | 15 | 5 | –4 | –11 |
| Волгоград | –9 | –8 | –2 | 11 | 21 | 26 | 29 | 26 | 18 | 8 | 1 | –4 |
| Ставрополь | –4 | –3 | 1 | 11 | 18 | 25 | 27 | 25 | 18 | 10 | 4 | –1 |
| Краснодар | –3 | –1 | 4 | 14 | 22 | 28 | 30 | 28 | 21 | 12 | 6 | 1 |
| Баку | 4.2 | 4.4 | 7.9 | 15.1 | 23.7 | 28.8 | 32.9 | 30.6 | 24.6 | 18.0 | 11.2 | 6.8 |
| Астрахань | –5 | –5 | 2 | 13 | 23 | 29 | 31 | 28 | 20 | 10 | 3 | –2 |
| Махачкала | 0 | 1 | 5 | 13 | 21 | 27 | 30 | 29 | 22 | 14 | 8 | 3 |
| Самарканд | –1 | 2 | 8 | 16 | 24 | 31 | 34 | 30 | 23 | 14 | 6 | 1 |
| Ашхабад | 2 | 5 | 11 | 19 | 28 | 35 | 38 | 34 | 27 | 17 | 8 | 3 |
Таблица 8.
Средняя максимальная температура поверхности почвы
| Пункт | Месяц | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |
| Саратов | 1 | 3 | 11 | 39 | 51 | 57 | 58 | 55 | 45 | 27 | 13 | 3 |
| Оренбург | –1 | 1 | 10 | 41 | 54 | 58 | 56 | 56 | 48 | 39 | 30 | 10 |
| Волгоград | 2 | 6 | 20 | 41 | 53 | 58 | 59 | 55 | 48 | 32 | 17 | 6 |
| Краснодар | 14 | 19 | 34 | 46 | 55 | 59 | 60 | 58 | 51 | 39 | 26 | 17 |
| Баку | 27 | 30 | 38 | 50 | 58 | 64 | 64 | 67 | 56 | 58 | 38 | 26 |
| Астрахань | 8 | 13 | 9 | 46 | 56 | 61 | 62 | 60 | 51 | 35 | 22 | 11 |
| Махачкала | 13 | 17 | 29 | 41 | 50 | 56 | 56 | 55 | 48 | 37 | 26 | 17 |
| Ашхабад | 36 | 43 | 54 | 61 | 70 | 72 | 72 | 72 | 67 | 56 | 45 | 37 |
Температура воздуха играет не такую большую роль в экологии ушастой круглоголовки, поскольку она бо́льшую часть времени либо лежит на разогретом песке, либо в самую жару стоит на вытянутых лапах, приподняв туловище на 2–3 см над почвой. Именно в этом втором варианте температура тела ящериц становится в значительной степени зависимой от температуры воздуха. В большинстве случаев температура воздуха в самые жаркие месяцы года в регионах, где живут ушастые круглоголовки, немного выше, чем там, где их нет (табл. 9). Кроме этого, здесь надо учесть, что на самом деле имеет смысл учитывать температуру тела в период активности ящериц, то есть за период от мая до конца октября и с 9-00 до 19-00 включительно (табл. 10).
Таблица 9.
Средняя месячная температура воздуха
| Пункт | Месяц | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |
| Саратов | –11.0 | –11.4 | –4.8 | 6.6 | 15.0 | 19.4 | 21.9 | 19.9 | 14.0 | 5.4 | 2.0 | –8.3 |
| Оренбург | –14.8 | –14.2 | –7.3 | 5.2 | 15.0 | 19.7 | 21.9 | 20.0 | 13.4 | 4.5 | 4.0 | –11.2 |
| Волгоград | –9.1 | –7.6 | –1.4 | 10.0 | 17.0 | 21.0 | 23.4 | 22 | 16.2 | 7.5 | 1.4 | –4.2 |
| Ставрополь | 0.1 | 1.2 | 5.4 | 14.7 | 20.7 | 24.7 | 27.4 | 26.9 | 22.1 | 14.8 | 8.5 | 2.9 |
| Краснодар | –1.6 | –0.6 | 4.3 | 11.3 | 17.0 | 20.7 | 23.3 | 22.7 | 17.6 | 11.4 | 5.6 | 1.1 |
| Баку | 3.9 | 4.1 | 6.3 | 11.2 | 17.7 | 22.6 | 25.7 | 25.7 | 21.8 | 16.6 | 11.1 | 6.8 |
| Астрахань | –2.8 | –1.5 | 5.4 | 16.1 | 24.0 | 28.4 | 31.0 | 29.7 | 23.4 | 14.8 | 6.5 | 0.2 |
| Махачкала | –0.5 | 0.2 | 3.5 | 9.4 | 16.3 | 21.5 | 24.6 | 24.1 | 19.4 | 13.4 | 7.2 | 2.6 |
| Самарканд | 0.2 | 3.0 | 7.4 | 13.8 | 9.2 | 23.2 | 25.6 | 23.7 | 18.6 | 12.6 | 6.4 | 2.2 |
| Ашхабад | 1.4 | 4.4 | 9.4 | 16.2 | 23.1 | 28.2 | 30.7 | 30.1 | 23.8 | 16.5 | 8.7 | 3.7 |
Таблица 10.
Средняя месячная температура воздуха в Самарканде, вычисленная в двух вариантах
| Характеристика | Месяц | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |
| За целые сутки | 0.2 | 3.0 | 7.4 | 13.8 | 19.2 | 23.2 | 25.6 | 23.7 | 18.6 | 12.6 | 6.4 | 2.2 |
| С 9-00 до 19-00 включительно | – | – | – | – | 23.5 | 28.5 | 30.5 | 29.1 | 24.3 | 17.3 | – | – |
В местах, где ушастые круглоголовки живут, средние максимальные температуры воздуха в жаркие месяцы обычно превышают 30°С (табл. 11). В Ставрополе, где их нет, температуры воздуха в жаркое время года высокие, но другие климатические характеристики (например, температуры зимовок – табл. 6) не позволяют им там быть. А в полутора десятков километров от Махачкалы на бархане Сарыкум эти круглоголовки есть. И хотя по чисто климатическим параметрам там вроде бы оказывается прохладнее, чем надо для этих ящериц, на са́мом деле температуры воздуха на высоте 2–3 см над песком на само́м бархане будут выше (к сожалению, конкретных данных по этому участку у нас пока нет, но в Каракумах и Кызылкумах дело обстоит именно так).
Таблица 11.
Средняя максимальная температура воздуха
| Пункт | Месяц | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |
| Саратов | –7.6 | –7.6 | –1.5 | 11.3 | 21.0 | 25.4 | 27.5 | 26.2 | 19.5 | 10.0 | 1.0 | –5.5 |
| Оренбург | –10.7 | –10.0 | –2.9 | 11.4 | 21.8 | 26.4 | 28.5 | 27.0 | 20.3 | 9.7 | –0.2 | –7.7 |
| Волгоград | –5.7 | –5.2 | 1.7 | 14.6 | 23.1 | 27.4 | 30.0 | 28.8 | 22.2 | 12.7 | 3.8 | –2.7 |
| Ставрополь | 10 | 11 | 17 | 25 | 28 | 31 | 35 | 33 | 29 | 24 | 18 | 13 |
| Краснодар | 2.8. | 3.6 | 9.5 | 17.7 | 23.3 | 27.0 | 29.8 | 29.5 | 24.7 | 18.0 | 11.1 | 5.2 |
| Баку | 6.8 | 7.1 | 9.5 | 15.6 | 22.3 | 27.1 | 30.1 | 30.0 | 25.4 | 19.6 | 13.9 | 9.4 |
| Астрахань | 5 | 6 | 15 | 25 | 31 | 34 | 36 | 35 | 31 | 23 | 15 | 8 |
| Махачкала | 2.8 | 3.0 | 6.8 | 13.8 | 20.8 | 25.8 | 28.6 | 28.0 | 23.2 | 16.8 | 10.5 | 5.5 |
| Самарканд | 5.3 | 8.2 | 13.3 | 20.1 | 26.4 | 31.1 | 33.4 | 31.9 | 27.7 | 21.4 | 13.9 | 7.9 |
| Ашхабад | 6.8 | 9.5 | 14.8 | 22.5 | 30.0 | 35.2 | 37.6 | 36.6 | 31.0 | 23.4 | 15.7 | 7.9 |
5) Еще один важный аспект, позволяющий ушастым круглоголовкам вести нормальную активность – температуры в дневных убежищах, которые к тому же определенным образом коррелируют с температурными условиями на поверхности. Обычно ящерицы в дневное время находятся в норах на глубине 0.2–0.4 м. Температура в этих убежищах с июня по сентябрь должна быть примерно в диапазоне от 24–25 до 30°С (табл. 12, 13). Это необходимо ящерицам для уменьшения амплитуды суточных колебаний температуры тела и приближения ее к высокой температуре термостабилизации, чтобы успешно протекали сезонные циклы гаметогенеза, беременность, чтобы было возможно активное питание и т.п.
Таблица 12.
Средняя температура почвы на глубине расположения дневных убежищ ушастых круглоголовок (примерно 0.4 м)
| Пункт | Месяц | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |
| Махачкала | 6.3 | 6.5 | 8.5 | 13.9 | 20.8 | 26.0 | 29.1 | 29.4 | 25.6 | 19.6 | 13.5 | 8.6 |
| Самарканд | 2.9 | 4.3 | 7.9 | 13.5 | 20.0 | 25.4 | 28.7 | 28.0 | 23.7 | 17.0 | 9.5 | 4.7 |
Таблица 13.
Средняя температура почвы на глубине расположения дневных убежищ ушастых круглоголовок (примерно 0.2 м)
| Пункт | Месяц | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | VII | VIII | IX | X | XI | XII | |
| Оренбург | 14.0 | 19.3 | 22.7 | 21.9 | 16.1 | 8.5 | ||||||
| Саратовская обл., Александров Гай | 15.1 | 20.6 | 22.8 | 22.7 | 16.1 | 8.5 | ||||||
| Ставрополь | 8.6 | 15.2 | 20.0 | 23.1 | 22.7 | 18.2 | 12.2 | |||||
| Краснодар | 11.6 | 18.1 | 22.9 | 25.7 | 25.3 | 20.7 | 13.8 | |||||
| Волгоград | 17.7 | 22.7 | 25.8 | 24.6 | 19.6 | 10.6 | ||||||
| Астрахань | 11.1 | 18.7 | 24.0 | 26.9 | 26.0 | 20.7 | 13.1 | |||||
| Махачкала | 12.4 | 19.4 | 24.1 | 27.0 | 26.8 | 22.0 | 15.6 | |||||
| Самарканд | 0.9 | 3.1 | 8.4 | 14.4 | 20.7 | 26.6 | 29.5 | 28.3 | 23.7 | 16.5 | 8.7 | 4.3 |
Таблицы 12 и 13 показывают, что в районах Астрахани, Махачкалы и равнинной Средней Азии в теплый сезон (с мая по конец сентября) температуры в дневных убежищах составляют примерно 21–29.5°С (24.0 ± 3.27°С). В Оренбургской, Саратовской, Ставропольской областях, Краснодарском крае и Каракалпакии температуры в зоне дневных убежищ были 16–26°С (21.7 ± 4.13°С), то есть более низкими, чем те, которые необходимы для этих ящериц.
Климатические условия зимовок ушастых круглоголовок в Краснодаре (если бы там присутствовали биотопические и другие экологические условия для их обитания) очень близко подходят к допустимым, но эти условия не позволяют им там зимовать и быть активными днем.
Эта ситуация касается и многих других видов рептилий, и ряда других климатических и географических показателей.
Обращаем внимание на то, что характеристики, приведенные в указанных выше примерах, являются не “одними из многих”, которые можно учесть, а можно и проигнорировать. Каждая из них даже в отдельности жестко определяет распространение данного вида рептилий. И подобных примеров для других видов можно привести десятки и даже более.
Все это говорит о том, что маркерные характеристики, которые важны именно для данных видов рептилий и теоретически могли бы выявить компьютерные программы, неоднозначны, чаще всего не регистрируются на стандартных метеорологических площадках, и, скорее всего, ими на самом деле не выявляются. Да, к тому же, они неодинаковы для разных видов, даже обитающих на одних и тех же территориях. Именно поэтому смоделированные с помощью компьютерных ГИС-программ ареалы оказываются лишь приблизительными, вероятными, зачастую далекими от фактических, а не реально возможными, и порой существенно отличаются от реально существующих.
ПУТИ РЕШЕНИЯ ПРОБЛЕМ
Логика действующего ныне подхода к компьютерному моделированию ареалов животных и расчета экологических ниш вполне понятна, но, по нашему мнению, она содержит в себе серьезный недостаток, который требует корректировки. Проблема заключается в том, что при анализе ареалов и экологических ниш в расчет берутся только характеристики внешней среды, но совершенно не учитываются потребности сами́х животных, их реальная экология, врожденные физиологические и др. требования к внешней среде, необходимые для их нормальной жизнедеятельности. Связи между физиологическими потребностями и характеристиками среды часто могут быть очень сложными. Для того, чтобы связать все части этой зависимости воедино и облечь ее в цифры и формулы, необходимо произвести предварительные исследования экологии интересующих видов, их физиологии, физиологической экологии, выявить набор видоспецифичных и важных в данных условиях среды критических характеристик, определяющих, регулирующих и формирующих режимы их жизнедеятельности – активности, индивидуального выживания, протекания сезонных физиологических циклов, выживания и процветания популяций и вида в целом. Просто по анализу многих точек ареала вида даже по большому набору случайных, стандартных характеристик, заложенных в базы данных компьютерных ГИС-программ, без понимания физиологических, климатических, экологических и пр. приоритетов, получить удовлетворительные результаты вряд ли получится. Другими словами, определить физиологические требования животных к среде обитания через анализ только самой среды обитания этих животных не выйдет.
Проведенные исследования по термобиологии рептилий показали, что стабильный, видоспецифичный комплекс термофизиологических показателей определенным образом регулирует и определяет режим суточной и сезонной активности этих животных, их биотопическое распределение и географическое распространение, режимы протекания у них сезонных физиологических циклов питания и размножения (Черлин, 2014). Определив этот комплекс эндогенных характеристик, можно сформировать физиолого-экологическую матрицу (ФЭМ) отдельных видов. С помощью нее, зная определенные характеристики внешней среды, есть возможность с большой вероятностью просчитать указанные выше экологические особенности для данных видов рептилий (Cherlin, 1991, 2015).
Эти физиолого-экологические закономерности дают основание полагать, что без их учета, то есть без понимания того, на основе чего формируются экологические закономерности, достаточно удовлетворительно связать экологические проявления вида и его географическое распространение с определенными, отдельными, единичными “маркерными” характеристиками внешней среды будет крайне затруднительно и малоэффективно. Для того, чтобы реально выделить важные именно для данного случая характеристики, необходимо знать экологические и физиологические особенности животных данного вида, их эндогенные требования к внешней среде и вполне определенный набор факторов внешней среды, важный и характерный именно для этих животных.
И, наконец, хотелось бы окончательно определиться с тем, зачем вообще все эти компьютерные программы нужны? Ведь любые программы создаются для чего-то. Какие цели они преследуют?
1) Одна из возможных целей – смоделировать потенциальный ареал, чтобы иметь возможность поиска новых регионов, где данный вид возможно было бы отыскать. Цель, вероятно, интересная и заманчивая. Однако, одно то, что на рассчитанных исследователями по таким методикам площадях, не входящих в истинный ареал животных, этих видов не найдено, должно было бы, скорее, насторожить, чем радовать… Мало того, практика показывает, что никто, кроме хорошего эколога, знакомого с биологией данного вида, не сможет лучше отыскать те микростации, которые подходят для его обитания. И зачастую, осуществляет он этот поиск, исходя не из показателей микроклимата и прочих маркеров, а из знания и понимания тех требований к внешней среде, которые важны для вида, биологию которого он знает и понимает. Кстати, это именно та сторона биологии животных, которую мы и призываем добавить в компьютерные ГИС-программы. Возможно, тогда они станут реальными помощниками биологам в решении данной задачи. Пока же данная цель, практически, даже приблизительно не достижима.
2) Другая возможная цель – смоделировать предположительные ареалы отдельных видов в прошлом в случае наличия информации о климатических и других экологических условиях на определенных территориях в эти прошедшие периоды времени. Это может иметь значение при изучении возможных путей распространения видов из зон их возникновения. Это красивая цель, но, к сожалению, она вряд ли достижима. Положительного результата можно было бы ожидать только если бы базы данных этих компьютерных программ смогли выдавать подробные климатические и др. характеристики отдельных точек на поверхности Земли в прошлом, иногда – весьма далеком… Но это, практически, нереально, так как такой компьютерный анализ сможет дать только весьма приближенные результаты, которые вряд ли могут быть квалифицированы как достоверные.
3) Третья возможная цель – в процессе моделирования ареалов выявить те важнейшие характеристики, которые могут определять экологию видов, их географическое распространение. Видимо, именно эти характеристики, с большой степенью неопределенности, полученные с помощью программ, авторы называют характеристиками экологических ниш животных. Но, как показали материалы данной статьи, это “с натяжкой” можно было бы назвать так только в случае, если в программы были бы внесены предложенные нами дополнения – данные о биологии изучаемых видов, об их требованиях к условиям внешней среды.
4) Но по анализу точек в ареале, скорее всего, доработанные программы могли бы более или менее достоверно показать не маркерные характеристики внешней среды, а выявить, насколько достоверно определены эндогенные требования животных данных видов к внешней среде. Это вполне достойная задача.
Получается, что ни одна из трех первых отмеченных выше целей, которые могли бы быть решены в процессе компьютерного моделирования ареалов или расчета экологических ниш с помощью компьютерных ГИС-программ, не может быть решена ими в их современной форме. Похоже, что только последняя (четвертая) из приведенных выше целей, вполне серьезная и важная, наиболее адекватно могла бы быть достигнута с помощью этих программ, но лишь в случае их корректного усовершенствования. Таким образом, сами программы, их задачи и пути их применения требуют серьезной, принципиальной доработки.
ЗАКЛЮЧЕНИЕ
Все, изложенное выше, показывает, что в компьютерные программы, анализирующие ареалы и экологические ниши рептилий разных видов, должны быть обязательно включены физиологические требования данных видов к среде обитания. Набор этих характеристик видоспецифичный, и получается он при специальных экологических и физиолого-экологических исследованиях (Черлин, 2014; Cherlin, 1991, 2015; и др.), которые необходимо предварительно провести и затем ввести их определенным образом в программу. Только в таком случае эти программы значительно приблизят результаты своего анализа к реальному положению дел в природе. Кроме того, следует, видимо, внести коррективы и в понимание целей использования этих программ. После модернизации, они могут быть наиболее полезны для выявления того, насколько достоверно определены эндогенные требования животных данных видов к внешней среде.
Список литературы
Климат Ашхабада. Л.: Гидрометеоиздат, 1984. 192 с.
Климат Баку. Л.: Гидрометеоиздат, 1988. 109 с.
Климат Краснодара. Л.: Гидрометеоиздат, 1990. 109 с.
Климат Самарканда. Л.: Гидрометеоиздат, 1983. 190 с.
Литвинов Н.А. К вопросу об оценке температуры тела рептилий, ее соотношении с внешней температурой и адаптивности размеров и окраски // Принципы экологии. 2015. Т. 4. № 1. С. 4–16.
Научно-прикладной справочник по климату СССР. Сер. 3. Многолетние данные. Ч. 1–6. Вып. 12. Татарская АССР, Ульяновская, Куйбышевская, Пензенская, Оренбургская, Саратовская обл. Л.: Гидрометеоиздат, 1988. 548 с.
Научно-прикладной справочник по климату СССР. Сер. 3. Многолетние данные. Ч. 1–6. Вып. 13. Волгоградская, Ростовская и Астраханская области, Краснодарский и Ставропольский край, Калмыцкая, Кабардино-Балкарская, Чечено-Ингушская и Северо-Осетинская респ. Л.: Гидрометеоиздат, 1990. 724 с.
Научно-прикладной справочник по климату СССР. Сер. 3. Многолетние данные. Ч. 1–6. Вып. 15. Дагестанская АССР, Азербайджанская ССР, Нахичеванская АССР. Л.: Гидрометеоиздат, 1990. 264 с.
Научно-прикладной справочник по климату СССР. Сер. 3. Многолетние данные. Ч. 1–6. Вып. 19. Узбекская ССР. Кн. 2. 1989. Л.: Гидрометеоиздат, 349 с.
Научно-прикладной справочник по климату СССР. Сер. 3. Многолетние данные. Ч. 1–6. Вып. 28. Калужская, Тульская, Тамбовская, Брянская, Липецкая, Орловская, Курская, Воронежская, Белгородская обл. Л.: Гидрометеоиздат, 1990. 366 с.
Научно-прикладной справочник по климату СССР. Сер. 3. Многолетние данные. Ч. 1–6. Вып. 30. Туркменская ССР. Л.: Гидрометеоиздат, 1989. 501 с.
Стрельников И.Д. Значение солнечной радиации в экологии высокогорных насекомых // Зоол. журн. 1940. Т. 19. № 2. С. 218–237.
Стрельников И.Д. Значение солнечной радиации в экологии высокогорных рептилий // Зоол. журн. 1944. Т. 23. № 5. С. 250–256.
Целлариус Е.Ю., Целлариус А.Ю. Температурные условия активности серого варана (Varanus griseus, Reptilia, Sauria) // Зоол. журн. 1997. Т. 76. № 2. С. 206–211.
Черлин В.А. К термобиологии серого геккона (Cyrtopodion russowi), полосатой ящурки (Eremias scripta) и степной агамы (Trapelus sanguinolentus) в Восточных Каракумах // Изв. АН Туркм. ССР, Серия биол. наук. 1988. № 5. С. 36–43.
Черлин В.А. Рептилии: температура и экология. Saarbrücken: Lambert Acad. Publishing, 2014. 442 с.
Черлин В.А. Тепловые адаптации рептилий и механизмы их формирования // Принципы экологии. 2015. Т. 4. № 1. С. 17–76.
Черлин В.А. Иван Дмитриевич Стрельников. Часть 2. Научная деятельность // Принципы экологии. 2019. Т. 8. № 1. С. 3–39.
Черлин В.А., Музыченко И.В. Термобиология и экология сетчатой ящурки (Eremias grammica), ушастой (Phrynocephalus mystaceus) и песчаной (Ph. interscapularis) круглоголовок летом в Каракумах // Зоол. журн. 1983. Т. 62. № 6. С. 897–908.
Черлин В.А., Целлариус А.Ю. Зависимость поведения песчаной эфы, Echis multisquamatus Cherlin 1981 от температурных условий в Южной Туркмении // Фауна и экология амфибий и рептилий палеарктической Азии / Тр. Зоол. ин-та АН СССР. Т. 101. Л.: Наука, 1981. С. 96–108.
Черлин В.А., Чикин Ю.А. К термобиологии ящериц горных районов Узбекистана // Герпетол. исследования. Л.: ЛИСС, 1991. № 1. С. 119–129.
Cherlin V.A. Method of ecology-physiological matrix of species in researches on reptile biology // Герпетол. исследования. Л.: ЛИСС, 1991. № 1. С. 138–146.
Cherlin V.A. Physiology-ecological matrix and its significance in biology of reptiles // Some important aspects of thermal biology of reptiles. Saarbrücken: Lambert Acad. Publishing, 2015. P. 117–125.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии