

Успехи современной биологии, 2020, T. 140, № 2, стр. 107-115
Экологическая генетика и современные проблемы биосферы
И. А. Захаров *
Институт общей генетики им. Н.И. Вавилова РАН
Москва, Россия
* E-mail: iaz34@mail.ru
Поступила в редакцию 17.01.2020
После доработки 17.01.2020
Принята к публикации 17.01.2020
Аннотация
Рассмотрены проблемы биосферы – глобальное потепление, загрязнение окружающей среды, последствия экономической глобализации, сокращение биоразнообразия, получение и выращивание генетически-модифицированных организмов, и связанные с этими проблемами задачи экологической генетики как научного направления, занимающегося изучением генетических процессов в популяциях с учетом влияний окружающей среды.
ВВЕДЕНИЕ
Земля – единственная планета Солнечной системы, где существует жизнь. Окружающие Землю условия космоса и условия на других планетах – вакуум, низкие или высокие температуры, высокий уровень радиации – не пригодны для появления и сохранения жизни. Ее нет и в глубоких слоях Земли с их темнотой, высокой температурой, большими давлениями. Наша планета, таким образом, окружена слоем, довольно тонким, единственно пригодным для обитания организмов, – это вода и донные осадки, поверхность суши и нижние пласты атмосферы. Этот слой – не только место обитания всех известных нам организмов. Он буквально насыщен ими: достаточно сказать, что ежегодная продукция органического вещества, создаваемого живущими в этом слое растениями, животными и микроорганизмами, достигает колоссальной величины, значимой и в геологическом масштабе, – общая годовая продукция сухого органического вещества на Земле составляет примерно 2 × 1011 т. Живые организмы не только поддерживают свое существование в поверхностном слое Земли – они активно влияют на его свойства – структуру, химический состав. Поверхностный слой Земли – это биосфера, сфера обитания живых организмов, постоянно перерабатываемая ими и включающая их самих.
Современная биосфера складывалась в результате протекавшей многие миллионы лет эволюции. В ходе нее возникла та гармония природы, которая давно поражала наблюдателей. Если же подойти к процессам биосферы не с позиции эмоционального восприятия, а с позиции научного анализа, то можно сказать, что эти процессы прекрасно сбалансированы, протекают устойчиво, и в основном в биосфере поддерживается равновесие в поглощении и выделении веществ и энергии. Мы говорим “в основном”, так как происходит определенное запасание энергии в форме органического вещества нефти, каменного угля, торфа.
Точнее, однако, было бы сказать не “поддерживается” равновесие, а “поддерживалось”, потому что существовавшее миллионы лет положение значительно изменилось с появлением человека. Даже первобытный человек, вооружившись огнем и примитивными орудиями, существенно влиял на природу, истребив или ускорив гибель отдельных видов животных, уничтожив лес на некоторых территориях. В основном же деятельность человека до недавнего времени если и не вполне вписывалась в природное равновесие, то и не очень сильно его нарушала.
Природа стала драматически изменяться с середины XX века. Развитие человеческой цивилизации, идущее по своим законам, привело к двум явлениям. Во-первых, к бурному росту населения Земли от 100–230 млн в начале нашей эры до 1 млрд в 1850 г., 2 млрд – в 1930, 3 млрд – в 1960, 4 млрд – в 1975 и 6 млрд человек к 2000 г. (Капица, 1999). В настоящее время на Земле уже живет 7 млрд 756.5 млн человек (https://www.worldometers.info/world-population/). Огромное народонаселение нуждается в пище и в удовлетворении многих других потребностей и производит большое количество отходов, многие из которых трудно перерабатываемы или опасны для живых организмов. Во-вторых, человечество оказалось вооруженным мощной техникой, возможности которой с каждым десятилетием увеличиваются.
Для удовлетворения потребности в пище человек все больше уничтожает природные ландшафты, заменяя их посевами культурных растений, а для удовлетворения потребности в энергии расходует накопленные в прошлые геологические эпохи нефть, газ и уголь. Города, индустриальные комплексы, дороги занимают большие территории и выключают их из естественного биологического круговорота веществ и энергии. Как следствие, большие территории, занятые урбанизированными и техногенными ландшафтами, перестают быть пригодными для существования животных и растений. В возрастающих масштабах меняется химический состав биосферы – из-за выделения отходов производства, транспорта, быта, использования химикатов в сельском хозяйстве. Объемы внесения в биосферу отдельных элементов, например тяжелых металлов, сопоставимы с последствиями геологических процессов.
Отдельные виды животных и растений исчезли, другим грозит гибель; снижается общая продуктивность биосферы – уменьшаются выделение кислорода и накопление органического вещества и, может быть, главное – нарушается равновесие, которое до сих пор было характерно для природных процессов. От изучения последствий этих экологических явлений не может оказаться в стороне и генетика.
Экологическая генетика – раздел науки о наследственности и изменчивости, который изучает процессы в популяциях в связи с экологическими условиями их существования. Естественно, когда экологическая ситуация во многих регионах мира или даже глобально изменяется в результате антропогенных влияний, перед экологической генетикой встают новые задачи и интересные возможности.
Далее я рассмотрю несколько ситуаций, с которыми сталкивается биосфера и человечество, и приведу примеры эколого-генетических исследований, в которых генетическими методами были изучены последствия этих новых вызовов.
ГЛОБАЛЬНОЕ ПОТЕПЛЕНИЕ
Глобальное потепление – это долгосрочное повышение средней температуры климатической системы Земли, которое наблюдается на протяжении последних более чем ста лет, и особенно выражено в последние десятилетия. Его причиной обычно считают человеческую деятельность, то есть антропогенный фактор. Помимо самого изменения средних температур потепление вызывает, особенно в северных регионах, разрушение сложившейся экологической среды – деградацию ледников, оттаивание “вечной” мерзлоты и т.п.
Вызванные глобальным потеплением климатические изменения приводят к изменениям видового состава фауны и флоры – исчезновению одних видов, появлению других, ранее не обитавших на той или иной территории.
Нами было замечено, что многочисленный в Архангельске или даже преобладающий вид кокцинеллид Adalia frigida после 2008 г. здесь больше не встречался; не удалось его найти (при высокой численности близкого, обитающего обычно в средней полосе, вида A. bipunctata) на Кольском полуострове и в Северной Норвегии (города Тромсе, Альта). При этом оказалось, что при распространении A. bipunctata на север клинально изменяется соотношение митохондриальных гаплотипов (Паленко и др., 2018).
Генетические изменения, связанные с климатическими, были описаны при изучении веснянок Lednia tumana (Plecoptera, Nemouridae) в горах штата Монтана (США); личинки этого вида обитают в холодной воде ручьев, стекающих с ледников. Было проведено сравнение генофондов популяций, собранных в 1997, 1998 и 2005 годах и в 2010 году. Было обнаружено снижение генетического разнообразия, в частности, – исчезновение одного, достаточно часто встречавшегося в старых выборках гаплотипа мтДНК. Снижение генетического разнообразия может свидетельствовать о снижении адаптивного потенциала изученной популяции (Jordan et al., 2016).
Исследования, проведенные на космополитическом виде дрозофилы, D. subobscura, четко выявили последствия глобального потепления. Авторы работы сравнили хромосомный полиморфизм в популяциях этого вида, современных и собранных на протяжении предыдущих 24 лет на трех континентах. В 21 из 22 изученных популяций полиморфизм изменился в сторону, характерную для более низкоширотных (южных) популяций. Наблюдаемое явление может быть следствием как адаптивной перестройки генофонда существующей в данной местности популяции, так и миграцией особей из более южных популяций (Balanyá et al., 2006). Подобные изменения генофонда были описаны и для популяций других видов дрозофил (van Heerwaarden, Hoffmann, 2007).
Возможность генетического приспособления организмов к изменившимся температурным условиям обитания и те обстоятельства, которые препятствуют такому быстрому приспособлению, обсуждаются в статье бельгийских авторов (De Meester et al., 2018).
Глобальное потепление не только угрожает многим представителям дикой флоры и фауны, но и создает проблемы для сельского хозяйства во многих регионах. В редакционной статье в журнале Nature Genetics (Genetics…, 2019) рассмотрены задачи генетики культурных растений в связи с драматическим изменением условий их выращивания.
Глобальное потепление способствует биологическим инвазиям – расселению южных видов и их укоренению в новых территориях. Вселение в области с умеренным климатом новых видов насекомых-переносчиков уже сейчас приводит к появлению тропических болезней там, где раньше они не проявлялись.
ХИМИЧЕСКОЕ И РАДИАЦИОННОЕ ЗАГРЯЗНЕНИЯ
Одни из первых исследований в области экологической генетики были посвящены изучению так называемого индустриального меланизма у насекомых отряда Lepidoptera. Энтомологи в Англии обратили внимание на накопление темно окрашенных особей в популяциях березовой пяденицы Biston betularia в городах и их окрестностях, где наиболее сильно проявляется промышленное загрязнение атмосферы (Majerus, 1998).
С начала ХХ века в огромном числе работ изучалось влияние загрязнения окружающей среды на генетические процессы в популяциях самых разных организмов. При этом разрабатывались следующие проблемы: 1) косвенное действие загрязнения – состав популяции модельного организма меняется как реакция на изменение состава биоты в районе загрязнения (именно сюда относится классический случай меланизма у бабочки B. betularia, см. ниже); 2) возрастание устойчивости к токсическому действию загрязнителя или суммы загрязнений; 3) увеличение частоты мутаций при попадании в среду химических или физических мутагенов.
Появление темных особей в популяциях березовой пяденицы B. betularia было замечено еще в середине ХIХ в., а в середине ХХ в. было экспериментально показано, что меланисты и не меланисты (светло-окрашенные бабочки) оказываются с разной частотой жертвами насекомоядных птиц. Многочисленные исследования, выполненные на B. betularia, дали следующие основные результаты (Majerus, 1998):
1. Бабочки служат пищей для насекомоядных птиц, склевывающих их со стволов деревьев, на которых проводят светлое время суток эти ночные бабочки; светлые бабочки менее заметны на стволах, покрытых лишайниками, а темные – на стволах, лишенных лишайников;
2. Лишайники чувствительны к промышленному загрязнению атмосферы, особенно к SO2 (двуокись серы), соответственно в промышленных районах стволы деревьев оказываются непокрытыми лишайниками, и здесь получает преимущество форма carbonaria;
3. Состав популяции в данной местности определяется двумя факторами – величиной селективного преимущества той или другой формы в конкретных условиях и миграцией бабочек из районов с другим составом популяций.
Такое описание истории березовой пяденицы оказалось, однако, чересчур упрощенным. В результате интенсивных исследований, проводившихся преимущественно в Великобритании, было показано сложное взаимодействие генетических и средовых факторов, которое привело к возрастанию доли меланистов в популяциях, а в последние десятилетия, напротив, – к их снижению (Majerus, 1998).
Иной механизм имеет явление накопления меланистов в популяциях некоторых городов у божьей коровки Adalia bipunctata.
Я.Я. Лусис первый обратил внимание на то, что в ближайших окрестностях Ленинграда – в Петергофе и Гатчине, черных форм встречается значительно меньше, чем в самом Ленинграде (1961). Мы дополнили это наблюдение, на протяжении 10 лет обследовав 17 популяций пригородов Санкт-Петербурга и населенных пунктов Ленинградской области (рис. 1). Корреляция между расстоянием от центра Петербурга и долей меланистов в популяции (рис. 2) высоко значима (r = = –0.75, P = 0.00036).
Рис. 1.
Состав популяций Adalia bipunctata Ленинградской области (собственные данные). Размер черного сектора соответствует процентному содержанию черных форм в популяции. I, II, III – районы Санкт-Петербурга (соответственно, Центральный, Южный, Юго-Западный).
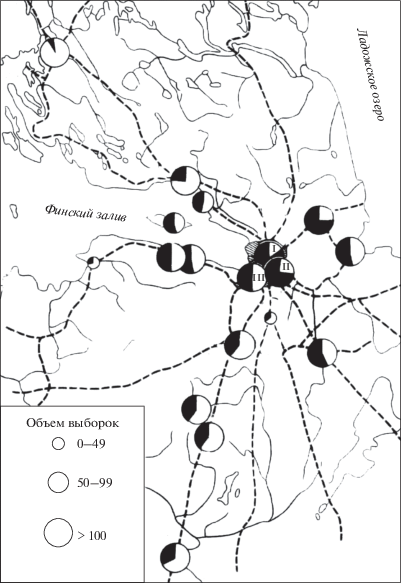
Рис. 2.
Зависимость частоты черных форм в популяциях Ленинградской области от расстояния мест сборов до центра Санкт-Петербурга. Указан 95%-ный доверительный интервал линии регрессии.
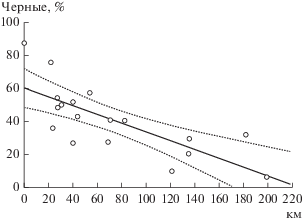
Обнаруженная зависимость может иметь двоякую основу. На территории Ленинградской области наиболее крупный индустриальный центр – Санкт-Петербург. В нем должно быть наибольшее загрязнение атмосферы промышленными и транспортными выбросами. Движением воздуха эти загрязнения распространяются по окрестностям, их концентрация уменьшается по мере удаления от Санкт-Петербурга. С другой стороны, можно ожидать расселения коровок (пассивного и активного) из города в область, и в этом случае меланизация областных популяций есть следствие не естественного отбора, а миграций. Вероятно, действуют оба механизма, и наблюдаемая картина является результатом взаимодействия по крайней мере двух факторов – распространения загрязнений и миграций жуков.
Подробные исследования, проведенные Кридом (Creed, 1966, 1971a) в Великобритании (было обследовано более 100 популяций), подтвердили существование связи между распространением меланизма и центрами промышленности. Автором получена высокая положительная корреляция между частотой меланистов и уровнем загрязненности атмосферы, в частности задымленностью воздуха.
Крид (Creed, 1971b) в Южном Уэльсе, для которого характерна низкая концентрация меланистов, обнаружил близ одного промышленного предприятия популяцию с высокой частотой меланистов (53.7%), которая постепенно, по мере удаления от фабрики, понижалась и в 35 км от нее составляла лишь 10%. Автор рассматривает это наблюдение как весомое доказательство того, что меланизм у адалии связан с промышленным загрязнением атмосферы.
Приведенные наблюдения говорят в пользу того, что высокая частота меланистов в некоторых крупных городах действительно является результатом влияния не каких-то климатических факторов, а антропогенных воздействий, вероятнее всего – загрязнения атмосферы промышленными предприятиями, а также печами, использовавшимися для отопления домов. При этом надо подчеркнуть, что божьи коровки, в отличие от бабочек, не часто подвергаются нападению хищных птиц, и преимущество меланистов не может быть объяснено их лучшей маскировкой, а связано, вероятно, с их большей физиологической устойчивостью.
Накопление токсических тяжелых металлов вблизи металлургических предприятий вызывает исчезновение растительного покрова при высоком загрязнении почвы, а при более низком – стимулирует процессы приспособления растений к токсическому агенту (Antonovics, 1975; Ernst, 2006).
В.А. Шевченко показал, что при существовании в условиях загрязнения имеет место повышение устойчивости и к такому физическому фактору, как радиация. Это было четко показано при изучении одноклеточных водорослей (Шевченко, 1979) и нескольких видов цветковых растений (Centaurea scabiosa, Lilium martagon, Crepis tectorum и др.), собранных на загрязненных участках (Шевченко, Померанцева, 1985; Шевченко и др., 1992). В первом случае – по увеличению выживаемости клеток при внешнем облучении, во втором случае – по тесту хромосомных аберраций.
ПОСЛЕДСТВИЯ ЭКОНОМИЧЕСКОЙ ГЛОБАЛИЗАЦИИ
Проявлением и условием экономической глобализации является расширение сети транспортных путей и резкое увеличение потоков перевозимых товаров. И то, и другое создало благоприятные возможности для неконтролируемого перемещения и расселения представителей видов флоры и фауны. Генетическое изучение аборигенных и инвазивных популяций позволило в ряде случаев установить источники и пути инвазии.
В 2005 г. было отмечено появление азиатской желтоногой осы Vespa velutina во Франции. В течение 5 лет этот вид занял уже территории размером 190 000 кв. км. В дальнейшем желтоногая оса была обнаружена в Испании, Португалии, Италии, Бельгии. Крупная хищная оса стала наносить заметный вред пчеловодству, укусы ее представляют опасность и для человека. Естественным ареалом вида является Юго-Восточная Азия. В Азии инвазивные популяции были обнаружены в Корее и в Японии.
Среди особей из области естественного распространения и из инвазивных популяций Франции и Кореи был изучен полиморфизм по митохондриальной ДНК (мтДНК) и по ядерным маркерам (22 микросателлитам) (Arca et al., 2015). По мтДНК в инвазивных популяциях разнообразия не было: во Франции все осы имели митотип F, в Корее – митотип K. На всей территории Юго-Восточной Азии присутствовало много различных митотипов, F и K были преобладающими в популяциях Южного Китая. Был сделан вывод, что именно эта область явилась источником инвазии как в Европу, так и в Корею. Вывод был подтвержден и при изучении микросателлитов. Авторы заключили также, что французская популяция взяла начало от одной (!) самки, оплодотворенной несколькими (по расчетам – 2–4) самцами (Arca et al., 2015).
Божья коровка Harmonia axyridis, другой азиатский вид, в последние десятилетия расселился по четырем континентам – Европе, Северной и Южной Америкам, Африке.
В 1988 году появилось сообщение об укоренении азиатской божьей коровки на юго-востоке штата Луизиана и на востоке штата Миссисипи. Были высказаны предположения о случайном характере интродукции H. axyridis в этом штате морским транспортом, поскольку все первые находки шести экзотических для Соединенных Штатов видов кокцинеллид к началу 1990-х годов, в число которых входила и H. axyridis, были сделаны в непосредственной близости от морских портов восточного побережья. Вторая инвазивная популяция была обнаружена в 1980-х годах на северо-западе Тихоокеанского побережья США. Эта популяция, как предполагают, появилась независимо от первой. К 1994 году H. axyridis находили уже в 24 штатах, большинство из которых прилегало к западному и восточному побережьям Соединенных Штатов, а к 2007 г. число штатов, где находили гармонию, возросло до 45. В настоящее время вид не выявлен лишь в Вайоминге и на Аляске (Андрианов и др., 2018).
В Европе инвазивные популяции H. axyridis появились в 2000 году, где быстро закрепились и распространились. К 2005 году вид появился в 27 государствах Западной и Восточной Европы, в том числе в Германии, Бельгии, Нидерландах, Англии, Швейцарии и Франции. В последующие годы H. axyridis была зарегистрирована практически во всех странах западной и центральной Европы: в Италии, Польше, Чехии, Австрии, Дании, Норвегии, Испании, Швеции, Венгрии, Сербии, Хорватии, Словакии, Латвии, Румынии, Украине, Болгарии, Черногории и Словении. Самые северные находки H. axyridis зафиксированы в Норвегии 2010 году. Автором этой статьи в 2018 г. размножение H. axyridis было замечено в Финляндии (г. Оулу).
В России H. axyridis была впервые найдена нами в Калининграде в 2010 году; после этого сообщалось о находках в других регионах: в Краснодарском крае – в 2011 году, и в Крыму – в 2013 году (Андрианов и др., 2018).
Изучение полиморфизма аборигенных и инвазивных популяций позволило определить пути глобального расселения H. axyridis. Развитие методик микросателлитного генотипирования и статистической обработки получаемых результатов значительно расширили возможности изучения генетической структуры популяций и их происхождения. Генетическая структура нативных, инвазивных и биоконтрольных (искусственно разводимых и используемых для подавления вредителей) популяций H. axyridis была изучена с использованием селективно нейтральных маркеров – 18 микросателлитных локусов (Lombaert et al., 2010, 2014). На основании полученных данных была проведена оценка вероятности различных сценариев расселения H. axyridis.
Проведенный анализ однозначно поддержал сценарий, в соответствии с которым Северная Америка оказалась первым континентом, который был захвачен H. axyridis в результате двух независимых событий инвазии из азиатского региона, по всей видимости, с северо-востока Китая (рис. 3). Первое вселение имело место в 1988 году на востоке США (ENA), второе – в 1991 году на западе Соединенных штатов. Значительное смешивание генофондов западной и восточной плацдармных популяций произошло в зоне их контакта в штатах Юта и Колорадо. Восточный плацдарм стал вероятным источником четырех инвазий на другие континенты – Южную Америку, Африку и Европу. В Южной Америке по крайней мере две инвазивные популяции – в Бразилии/Аргентине и в Чили были образованы в результате двух независимых событий интродукции из ENA. Один из отводков ENA проник и закрепился на западе Европы – в Бельгии и Франции, причем укоренение сопровождалось смешиванием с европейской биоконтрольной линией. Плацдармная популяция запада Северной Америки – WNA-популяция – оказалась источником по крайней мере двух независимых интродукций в Европу. Следствием одной из них стало укоренение “дикой” популяции H. axyridis в Северной Италии, тогда как вторая, вероятно, привела к смешиванию с западноевропейской инвазивной популяцией при распространении обеих популяций – отводков ENA и WNA в восточную и северную части материка – в Германию, Польшу, Чехию, Венгрию, Данию, Норвегию. Вполне вероятно, что в области смешивания могли иметь место и дополнительные интродукции.
Рис. 3.
Вероятный сценарий глобальной инвазии Harmonia axyridis по данным микросателлитного анализа. Указаны годы первого появления H. axyridis. AF – Африка, E – Европа, EBC– европейская биоконтрольная популяция, ENA – восток Северной Америки, SA – Южная Америка, WNA – запад Северной Америки (из Андрианов и др., 2018, по Lombaert et al., 2014).
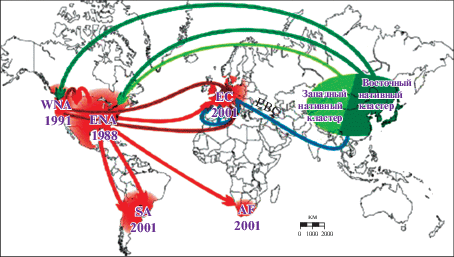
Неоднократно было показано, что вселение на новые территории крупной и агрессивной азиатской коровки вызывает снижение численности аборигенных видов кокцинеллид, с которыми вид-вселенец конкурирует.
Приведенные на рис. 3 пути расселения H. axyridis хорошо совпадают с основными мировыми путями торговли, по которым развозятся товары вместе со случайно попавшими в контейнеры живыми организмами.
ГЕНЕТИЧЕСКИ-МОДИФИЦИРОВАННЫЕ ОРГАНИЗМЫ
Широкое использование в сельском хозяйстве генетически-модифицированных организмов (ГМО) вызывает в обществе значительное беспокойство. Убедительных научных данных о вреде ГМО для здоровья человека не получено. Тем не менее, следует изучать и оценивать возможное влияние выращиваемых ГМО на дикую флору и фауну, то есть на состояние экологии, в тех регионах, где выращиваются генетически модифицированные сельскохозяйственные культуры.
Беспокойство вызывает, прежде всего, возможное неблагоприятное действие ГМ-растений на полезных (или безвредных) насекомых. Оценка опасности ГМО для биоразнообразия показала, что опасения здесь преувеличены. При получении устойчивых к насекомым-вредителям растений используются гены кристаллического белкового токсина из генома разных штаммов бактерии Bacillus thuringiensis. Этот, так называемый Bt-токсин, высокоспецифичен, и его варианты действуют на представителей одних, но не других отрядов насекомых. Bt-токсины, летальные для представителей отрядов Чешуекрылые или Жесткокрылые, оказываются безвредными для перепончатокрылых, в частности, для пчел (Duan et al., 2008).
Даже при потенциальной токсичности, ГМО-растения не повреждают не питающихся ими насекомых. Личинки популярной в США бабочки-монарха Danaus plexippus питаются листьями не кукурузы, а ваточника, растения рода Asclepias spp., (сем. Кутровых). Пыльца растений кукурузы, устойчивых к чешуекрылым насекомым, может попадать на другие, растущие близко от кукурузного поля, растения. Однако, наиболее широко возделываемые в США гибриды кукурузы заметного количества токсина в пыльце не образуют (хотя есть и формы, в которых токсин в пыльце присутствует). И, в целом, не получено данных, говорящих о вредных последствиях ГМО-кукурузы для бабочек-монархов, личинки которых питались растущими близ кукурузных полей кормовыми растениями (Sear et al., 2001).
При обсуждении экологических опасностей культивирования трансгенных сельскохозяйственных растений указывают на возможность распространения чужеродных, повышающих устойчивость, генов среди диких родственников культивируемого растения. В результате гербициды, предназначенные для борьбы с сорняками, могут оказаться неэффективными (Tsatsakis et al., 2017). Такая опасность действительно существует, но ее масштабы еще не определены. Более реальны экологические последствия увеличения количества используемых пестицидов (гербицидов), возможность чего создается устойчивостью к гербицидам ГМ-культур. Увеличение вброса в среду пестицидов создает угрозу здоровью людей и нарушает сложившуюся структуру биоты. В процитированном выше обзоре приводятся также важные сведения о законодательном регулировании использования ГМО в Европейском союзе и в России (Tsatsakis et al., 2017).
СОКРАЩЕНИЕ БИОРАЗНООБРАЗИЯ И ВЫМИРАНИЕ ВИДОВ
Все рассмотренные выше процессы, происходящие под влиянием человеческой деятельности в биосфере, ведут к сокращению биоразнообразия. Такое сокращение подрывает сбалансированность функционирования биосферы, и на каком-то этапе может привести к ее деградации с самыми печальными последствиями для человечества. Считается, что на Земле каждый год исчезает от нескольких тысяч до ста тысяч видов. Вымирание того или иного вида может быть быстрым, но это не мгновенный процесс. Очевидные этапы его – уменьшение общей численности особей, сокращение и фрагментация ареала. Эти этапы сопровождаются изменениями генетических характеристик популяции/популяций – уменьшением генетического разнообразия, возрастанием гомозиготности и инбредности. На последних этапах сокращения численности популяции увеличение частоты инбридинга в случае перекрестно-оплодотворяющихся организмов само становится фактором, способствующим или вызывающим гибель популяции. Популяции малой численности могут исчезать и по стохастическим причинам.
В работе Спилмена с соавт. (Spielman et al., 2004) проведено сравнение по уровню гетерозиготности популяций 170 видов, находящихся в угрожаемом состоянии, с популяциями родственных видов, которым вымирание не угрожает. Объекты сравнительного анализа – растения, позвоночные и беспозвоночные животные. Оказалось, что в 77% сравниваемых пар находящиеся по угрозой виды показали более низкую гетерозиготность, в среднем на 35%, чем их партнеры, родственные виды, находящиеся в благополучном состоянии. Таким образом, снижение генетического разнообразия не только сопутствуют движению популяции (вида) к исчезновению, но и, вероятно, является одним из факторов, способствующих вымиранию.
Роль генетических факторов в сохранении и вымирании вида рассмотрена в обзоре Франкхэма (Frankham, 2005).
Генетика не только описывает процессы, происходящие в ходе вымирания, но может дать рекомендации по поводу того, как его замедлить или предотвратить, поддержав жизнеспособность популяции. Соответствующее направление популяционной генетики было названо Ю.П. Алтуховым “природоохранной генетикой” (Алтухов, 1995, 2002); в англоязычной литературе используется термин “conservation genetics”.
ЗАКЛЮЧЕНИЕ
В настоящей статье были кратко рассмотрены особо острые проблемы, которые сейчас стоят перед экологической генетикой в связи с происходящими в биосфере изменениями. На некоторые из этих проблем автор настоящей статьи обратил внимание еще в 1984 году (Захаров, 1984). Каждая из кратко рассмотренных здесь проблем могла бы быть предметом отдельной большой статьи. Здесь моя задача была лишь наметить ключевые вопросы и дать примеры того, как они обсуждаются или решаются в литературе.
Генетическое изучение природных популяций было в свое время начато русскими учеными – С.С. Четвериковым в Москве и Ф.Г. Добржанским в США. Оба исследователя работали с “дикими” видами дрозофил и если С.С. Четверикову не удалось развить свои исследования, то Ф.Г. Добржанский, изучавший популяции на обширных территориях Северной и Центральной Америк, естественно перешел к анализу влияния условий обитания на генетическую структуру природных популяций, то есть к изучению экологической генетики. В СССР Я.Я. Лусис с середины 1920-х годов проводил классические исследования по экологической генетике на материале популяций жука Adalia bipunctata (Лусис, 1961). С 1920-х годов на различных объектах исследования, которые можно отнести к области экологической генетики, начали проводиться в Великобритании и в других западных странах. Их значимость особенно возрастала и осознавалась по мере понимания неблагополучного и тревожно ухудшающегося состояния биосферы.
Под воздействием все усиливающегося антропогенного пресса происходит не только деградация биосферы. Человек резко ускоряет темпы эволюции биоты Земли. Ускорение это в ничтожной степени связано с мутагенными эффектами антропогенных загрязнений, а в решающей степени – с появлением новых факторов естественного отбора. В этих условиях задача экологической генетики – анализ генетических процессов, происходящих в популяциях возможно большего числа видов-моделей, и разработка на этой основе прогноза развития эволюционного процесса в биосфере.
Список литературы
Алтухов Ю.П. Генетика популяций и сохранение биоразнообразия // Соросовский образоват. журн. 1995. № 1. С. 32–43.
Алтухов Ю.П. Природоохранная генетика // Наука в России. 2002. № 3. С. 25–32.
Андрианов Б.В., Блехман А.В., Горячева И.И. и др. Азиатская божья коровка Harmonia axyridis: глобальная инвазия. М.: Тов-во науч. изданий КМК, 2018. 143 с.
Захаров И.А. Экологическая генетика и проблемы биосферы. Л.: Знание, 1984. 32 с.
Капица С.П. Сколько людей жило, живет и будет жить на земле. Очерк теории роста человечества. М.: Наука, 1999. 136 с.
Лусис Я.Я. О биологическом значении полиморфизма окраски у двуточечной коровки Adalia bipunctata L. // Latvijas Entomologs. 1961. № 4. С. 3–29.
Паленко М.В., Андрианов Б.В., Романов Д.А. и др. Географический клинальный полиморфизм распределения митохондриальных гаплотипов Adalia bipunctata Linneaus, 1758 (Coleoptera: Coccinellidae) Норвегии // Генетика. 2018. Т. 54 (4). С. 456–461. https://doi.org/10.7868/S0016675818040070
Шевченко В.И. Радиационная генетика одноклеточных водорослей. М.: Наука, 1979. 254 с.
Шевченко В.А., Померанцева М.Д. Генетические последствия действия ионизирующих излучений. М.: Наука, 1985. 279 с.
Шевченко В.А., Печкуренков В.Л., Аврамов В.И. Радиационная генетика природных популяций. М.: Наука, 1992. 221 с.
Antonovics J. Metal tolerance in plants: perfecting an evolutionary paradigm // Intern. Conf. on heavy metals in the environment. Toronto, Ontario, 1975. P. 189–186.
Arca M., Mougel F., Guillemaud T. et al. Reconstructing the invasion and the demographic history of the yellow-legged hornet, Vespa velutina, in Europe // Biol. Invasions. 2015. V. 17. P. 2357–2371.
Balanyá J., Oller J.M., Huey R.B. et al. Global genetic change tracks global climate warming in Drosophila subobscura // Science. 2006. V. 313 (5794). P. 1773–1775. https://doi.org/10.1126/science.1131002
Creed E.R. Geographic variation in the two-spot ladybird in England and Wales // Heredity. 1966. V. 21. P. 57–72.
Creed E.R. Melanism in the two-spot ladybird, Adalia bipunctata, in Great Britain // Ecological genetics and evolution / Ed. E.R. Creed. Oxford: Blackwell, 1971a. P. 134–151.
Creed E.R. Industrial melanism in the two-spot ladybird and smoke abatement // Evolution. 1971b. V. 25. P. 290–293.
De Meester L., Stoks R., Brans K.I. Genetic adaptation as a biological buffer against climate change: potential and limitations // Integrat. Zool. 2018. V. 13. P. 372–391. https://doi.org/10.1111/1749-4877.12298
Duan J.J., Marvier M., Huesing J. et al. A meta-analysis of effects of Bt crops on honey bees (Hymenoptera: Apidae) // PLoS One. 2008. V. 3 (1). P. e1415. https://doi.org/10.1371/journal.pone.0001415
Ernst W.H.O. Evolution of metal tolerance in higher plants // For. Snow Landsc. Res. 2006. V. 80 (3). P. 251–274.
Frankham R. Conservation genetics // Ann. Rev. Genet. 1995. V. 29. P. 305–327. PMID https://doi.org/10.1146/annurev.ge.29.120195.0015138825477
Genetics for a warming world (Editorial) // Nat. Genet. 2019. V. 51. P. 1195. https://doi.org/10.1038/s41588-019-0488-6
Jordan S., Giersch J.J., Muhlfeld C.C. et al. Loss of genetic diversity and increased subdivision in an endemic alpine stonefly threatened by climate change // PLoS One. 2016. V. 11 (6). P. e0157386. https://doi.org/10.1371/ journal.pone.0157386
Lombaert E., Guillemaud T., Cornuet J-M. et al. Bridgehead effect in the worldwide invasion of the biocontrol harlequin ladybird // PLoS One. 2010. V. 5 (3). P. e9743.
Lombaert E., Guillemaud T., Lundgren J. et al. Complementarity of statistical treatments to reconstruct worldwide routes of invasion: the case of Asian ladybird Harmonia axyridis // Mol. Ecol. 2014. V. 23 (24). P. 5979–5997.
Majerus M.E.N. Melanism. Evolution in action. Oxford, New York, Tokyo: Oxford Univ. Press, 1998. 338 p.
Sear M., Helmich R.L., Stanley-Horn D.E. et al. Impact of Bt corn pollen on monarch butterfly // PNAS USA. 2001. V. 98 (21). P.11937–11942.
Spielman D., Brook B.W., Frankham R. Most species are not driven to extinction before genetic factors impact them // PNAS USA. 2004. V. 101 (42). P. 15261–15264. https://doi.org/10.1073/pnas.0403809101
Tsatsakis A., Nawaz M.A., Kouretas D. et al. Environmental impacts of genetically modified plants: a review // Environ. Res. 2017.https://doi.org/10.1016/j.envres.2017.03.011
van Heerwaarden B., Hoffmann A.A. Global warming: fly populations are responding rapidly to climate change // Curr. Biol. 2007. V. 17 (1). P. R16–R18. https://doi.org/10.1016/j.cub.2006.11.035
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии