

Успехи современной биологии, 2020, T. 140, № 3, стр. 294-305
О фазах вселения нового вида в пресноводные экосистемы
Ю. С. Решетников *
Институт проблем экологии и эволюции им. А.Н. Северцова Российской академии наук
Москва, Россия
* E-mail: ysreshetnikov@gmail.com
Поступила в редакцию 02.02.2020
После доработки 03.02.2020
Принята к публикации 03.02.2020
Аннотация
Проблема биологических инвазий чужеродных видов стала актуальной со второй половины прошлого столетия. Часто именно хозяйственная деятельность человека по деформации природных экосистем, в том числе и водных, приводит к гигантскому росту числа случаев расширения видами своих естественных ареалов. Известно, что саморасселение или интродукция экзотического вида в районы, где он ранее не встречался, часто порождает резкие перестройки в экосистеме водоема и снижение биологического разнообразия. В ряде случаев эффект от интродукции нового вида подобен экологическому стрессу для экосистемы. На основании длительных мониторинговых исследований на примере вселения трех видов рыб в пресноводные водоемы (озера и реки) прослежены основные изменения в структуре рыбного населения. Выделены четыре фазы вселения вида.
ВВЕДЕНИЕ
Проблема биологических инвазий чужеродных видов актуальна последние 50 лет, она стала ключевой в исследованиях экосистем всего Земного шара (Элтон, 1960; Дгебуадзе, 2000; Биологические инвазии…, 2004; Слынько и др., 2010; Odum, 1985; Williamson, 1996; Inderjit et al., 2005; Dgebuadze et al., 2017). Одна из главных задач – организация мониторинговых исследований видов-вселенцев с целью установления скорости их проникновения в новые экосистемы, разработки методов экологического прогнозирования инвазий и конкретных мер контроля и защиты от вселенцев. На примере рыб основные факторы, которые способствуют попаданию чужеродных видов в новые места обитания, можно суммировать следующим образом:
1) непреднамеренные действия человека (постройка каналов и плотин, сброс балластных вод и т.п.);
2) преднамеренная акклиматизация и пересадка рыб с помощью человека (в том числе и садковое выращивание рыбы в естественных водоемах, когда рыба уходит из садков);
3) саморасселение рыб (многие виды при высокой численности ищут новые места обитания): появление жереха, судака и белоглазки в бассейне р. Северная Двина, расселение корюшки и ряпушки вниз по р. Волга из оз. Белое, распространение вверх по р. Волга тюльки и каспийских бычков и т.п.;
4) выпуск рыбы в водоем рыболовами и аквариумистами (ротан, eрш, аквариумные рыбы, в том числе и экзотические виды: пираньи, панцирная щука, акулы).
В 1960-е гг. у нас в стране проводились многочисленные работы по акклиматизации рыб (Бурмакин, 1963; Карпевич, Бокова, 1963; Карпевич, 1975; Кудерский, 2001). Однако многие попытки по акклиматизации рыб провалились; по нашим подсчетам, успех был только в 17% всех попыток пересадки рыб, хотя все разрешения на акклиматизацию имели “биологическое обоснование” и проходили экологическую экспертизу. Поэтому часто трудно предсказать поведение всей экосистемы по отношению к новому пришельцу и его дальнейшую судьбу. Чаще сама экосистема активно сопротивляется появлению нового вида. Установлено, что продвижение новых видов протекает в несколько этапов, на каждом из которых происходит образование устойчивых самовоспроизводящихся популяций. В этом плане очень важны длительные мониторинговые наблюдения.
В данной статье мы остановимся лишь на одном интересном моменте при интродукции нового вида в пресноводную экосистему – это резкое первоначальное увеличение численности вселенца, а затем столь же резкое падение его численности и биомассы.
МАТЕРИАЛЫ И МЕТОДЫ
Основой выводов и заключений послужили наши многолетние совместные с карельскими ихтиологами исследования на оз. Сямозеро (Решетников и др., 1982; Стерлигова и др., 2002), экспедиционные работы совместно с норвежскими коллегами в бассейне пограничной р. Пасвик (Попова и др., 2005; Решетников и др., 2008, 2011; Решетников, Попова, 2011; Amundsen et al., 1999, 2003; Reshetnikov et al., 2010; Liso et al., 2011; Reshetnikov, Popova, 2017); анализ данных по вселению ерша в водоемы Северной Америки проведен на основе опубликованных работ (Пронин и др., 1998; Обыкновенный eрш…, 2016; Brazner et al., 1998; Gunderson et al., 1998; Popova et al., 1998). Для оценки численности и биомассы рыб использовали данные по промысловым и научно-исследовательским уловам рыбы, а также показатели численности в единицах на промысловое усилие CPUE (catch per unit effort) в экз. рыб за ночь лова в пересчете на 100 м2 сетного полотна. Анализ питания проводили по обычным ихтиологическим методикам: по весовому методу для рыб-планктофагов и бентофагов и по восстановленным весам для хищных рыб (Методическое пособие…, 1974). В таблице по питанию рыб используется предложенный нами индекс относительной значимости IR (index of relative significance), который объединяет частоту встречаемости F и долю по весу Р (Попова, Решетников, 2011).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЯ
Европейская корюшка Osmerus eperlanus – вид с широким ареалом в Северной Европе от Франции до морей Балтийского, Белого и Баренцева, включая юго-восточную часть Скандинавского п-ова и юго-запад Ирландии. В России корюшка отмечена в бассейнах всех этих морей. Северной границей ее распространения в России является оз. Имандра в Мурманской обл. и низовья р. Печора. Самые южные популяции в виде снетка обитают в оз. Псковское и оз. Белое (Берг, 1948; Рыбы в заповедниках…, 2010; Сидоров, Решетников, 2014).
Оз. Сямозеро (Южная Карелия) входит в ареал обитания корюшки. Она есть и в соседних озерах, но в самом Сямозере ее раньше никогда не было. Вселение вида в Сямозеро сначала дало резкую вспышку численности (рис. 1) и существенно трансформировало схему потоков энергии по пищевым цепям, а потом численность корюшки пошла на спад под воздействием хищников, паразитов и конкурентов (Решетников и др., 1982; Стерлигова и др., 2002).
Рис. 1.
Динамика уловов рыбы в Сямозере (т в год) после вселения корюшки (по: Решетников и др., 1982).
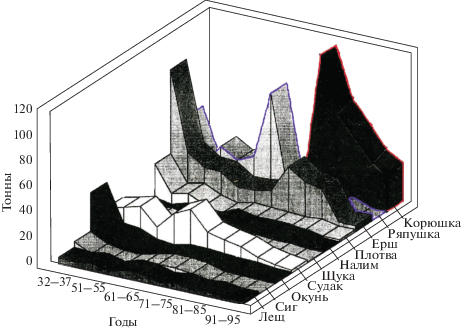
Основным конкурентом корюшки по питанию зоопланктоном в Сямозере была аборигенная европейская ряпушка Coregonus albula. Однако корюшка оказалась более сильным конкурентом, так как не только питалась планктоном, но частично выедала икру и личинок ряпушки. В одном желудке корюшки отмечалось до 40 личинок ряпушки и до 20 личинок сига (Стерлигова, 1979). Первые появления корюшки в Сямозере относятся к 1970-м гг., пик численности приходится на 1979–1980 гг. (точная причина появления не ясна, предполагают, что был непреднамеренный занос рыбаками).
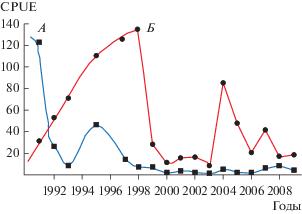
Численность корюшки быстро достигла высокого значения, выше реальной кормовой базы Сямозера, однако голодание и появление паразита Glugea привели к резкому падению ее численности. Уже к 1982–1984 гг. численность корюшки в озере сократилась втрое. В 2002–2009 гг. вновь было зафиксировано 100%-ное заражение корюшки этим же паразитом. Резко сократилась ее численность, что привело к падению уловов до 0.5 т/г. В 2013–2014 гг. как в водоеме, так и в питании хищных рыб корюшка отмечалась крайне редко, а с 2015 г. и по настоящее время и вовсе отсутствует, но ряпушка все еще не может восстановить свою прежнюю численность (рис. 2).
Рис. 2.
Динамика вылова вида-интродуцента европейской корюшки (А) и аборигенной ряпушки (Б) в 1970–2002 гг. в Сямозере (по: Стерлигова и др., 2002).

В последние десятилетия европейская корюшка существенно расширила границы области обитания. Ее распространение происходит разными путями: в результате рыбоводно-акклиматизационных мероприятий, саморасселения и случайного заноса.
В оз. Белое Вологодской обл. много столетий обитал снеток и далеких миграций не совершал. В 1943–1945 гг. его впервые отметили в Рыбинском вдхр., куда он попал из оз. Белое. Вероятно, он и раньше спускался вниз по р. Шексна до р. Волга, но в самой реке условия для его обитания были не очень благоприятными. После постройки плотины около г. Рыбинск в 1941 г. и начала заполнения Рыбинского вдхр. снеток был отмечен в первые годы его существования, и численность снетка быстро росла. В водохранилище снеток растет быстрее, живет дольше и частично меняет характер питания и фактически превращается в корюшку. Снеток и корюшка – это один и тот же вид Osmerus eperlanus, только снеток – мелкая форма – живет лишь в некоторых озерах Северо-Европейской части России, в частности в оз. Белое. Спускаясь вниз по Волге, он в новых водоемах по образу жизни становится типичной корюшкой (Иванова, 1982). Снеток был обнаружен в вдхр. Горьковское и Чебоксарское – в 1957 г., Куйбышевское – в 1958 г., Саратовское – в 1961 г. и Волгоградское – в 1970 г. В 1970-х гг. он пытался проникнуть в вдхр. Угличское и Иваньковское (рис. 3).
Рис. 3.
Распространение вниз по Волге снетка (корюшки) из оз. Белое (число обозначает год первой поимки корюшки в данном месте).

В 1990 г. в Рыбинском вдхр. корюшка была важным объектом промысла. При общем вылове рыбы в водоеме 1400–1800 т ее доля составила 40 т, что значительно превышало вылов многих видов карповых рыб, кроме плотвы Rutilus rutilus. В уловах экспериментального трала численность молоди корюшки на два порядка превосходила численность всех других видов рыб (Рыбы Рыбинского водохранилища…, 2015; Герасимов и др., 2018). Начиная с 1996 г. в водохранилище отмечается очень низкий уровень воды, это привело к осушению нерестилищ и гибели икры корюшки. Например, в 1994 г. за 10 мин траления учетным пелагическим тралом вылавливали в среднем 2120 экз. корюшки, в 1995 г. – 530 экз., в 1996 г. – 37 экз., в 1998−1999 гг. – всего 11 экз. В 2002 г. этот вид в Рыбинском вдхр. практически полностью исчез (Герасимов и др., 2018). Необходимо отметить, что в 1994 г. в Рыбинском вдхр. впервые была зарегистрирована каспийская тюлька Clupeonella cultriventris, которая поднялась по р. Волга из нижележащих водохранилищ. Она быстро освоила всю пелагиаль и заменила корюшку в качестве основного кормового объекта пелагических хищников.
Второй пример – это инвазия европейской ряпушки Coregonus albula в бассейн р. Пасвик. Ряпушка сначала была вселена в финское оз. Инари в 1960-е гг., первая ее поимка в озере относится к 1973 г., а с 1978 г. она стала регулярно встречаться в уловах (Salonen, 1998), резкая вспышка ее численности отмечена в 1989–1990 гг. (рис. 4). В эти годы уловы ряпушки достигали 300 т, затем последовал резкий спад, и уловы во все последующие годы не превышали 30 т/г.

Пасвик – пограничная река между Норвегией и Россией, вытекает из финского оз. Инари, течет на север и впадает в Баренцево море в районе г. Киркенес, река перегорожена рядом плотин, поэтому состоит из каскада водохранилищ. В пик своей максимальной численности в оз. Инари ряпушка стала распространяться вниз по всей р. Пасвик. Первая регистрация ряпушки в реке отмечена в 1989 г. (Hestefoss), в 1991 г. она ловилась уже ниже в районе оз. Ваггатем (Vaggatem) и оз. Рускебукта (Ruskebukta), в 1992 г. – в вдхр. Лангватн (Langvatn) и вдхр. Сванватн (Svanvatn) и в 1993 г. – около дер. Бьёрнватн (Bjornvatn) и в оз. Скруккебукта (Skrukkebukta), а к 1995 г. освоила весь бассейн реки (рис. 5).
Рис. 5.
Распространение ряпушки по системе р. Пасвик (число обозначает год ее появления в данном районе).

Рыбное население реки представлено восьмью видами рыб, причем сиг С. lavaretus имеет две формы. Малотычинковый сиг типа пыжьяна (с числом жаберных тычинок 15–30 с модой 24; длина тычинки 12–22% длины жаберной дуги) населяет зону литорали и профундаль, питается бентосными организмами и достигает крупных размеров, живет 12–15 лет, созревает в возрасте 3–4 лет. Среднетычинковый сиг (число жаберных тычинок 29–40 с модой 33; длина тычинки 15–25% длины жаберной дуги) занимает пелагическую зону и питается преимущественно зоопланктоном, он меньше по размеру, созревает в возрасте 2–3 лет, живет 10–13 лет. Эти две формы сигов широко представлены во многих северных водоемах Норвегии, Швеции, Финляндии и России. Южнее Кольского п-ова появляется еще третья форма – многотычинковый сиг (число жаберных тычинок 40–60 с модой 45; длина тычинки 20–36% длины жаберной дуги), в питании которого преобладают планктонные организмы.
Ряпушка активно вошла в состав рыбной части сообщества экосистем бассейна р. Пасвик: она быстро стала доминантным видом в новых для нее водоемах водохранилищного типа и заняла место планктофага в зоне пелагиали, потеснив оттуда и снизив численность среднетычинкового сига (рис. 6). Как планктофаг она имела явные преимущества в питании планктоном по сравнению со среднетычинковым сигом: верхний рот (у сига – нижний), длинные и тонкие жаберные тычинки (39–49 с модой 42), раннее созревание в возрасте 1–2 лет, продолжительность жизни не более 10 лет, размеры 10–17 см и масса до 40 г.
Рис. 6.
Численность среднетычинкового сига (А) и ряпушки (Б) в р. Пасвик (водоем Ваггатем) в первые годы после появления в нем ряпушки.
Не выдержав пищевой конкуренции, среднетычинковый сиг переселился в прибрежные биотопы и стал больше потреблять бентос, его численность резко снизилась (рис. 6). Влияние ряпушки сказалось и на видовом и размерном составе зоопланктона: снизился средний размер зоопланктеров, а вместо крупных форм (Daphnia) стали преобладать мелкие (Bosmina) (Amundsen et al., 2003).
В свою очередь на питание ряпушкой активно переключились некоторые хищные рыбы. Нарастание численности ряпушки шло по экспоненциальной кривой, максимум – в 1998 г. (CPUE = 135), однако на следующий год последовало столь же резкое падение численности (CPUE = 26). Второй всплеск численности был в 2004 г. (CPUE = 83), затем вновь резкое падение (CPUE = 19), после чего уловы не превышали 12–15 экз. за ночь. Как и в оз. Инари, после бума численности наступил резкий спад: уловы на усилие с 1998 г. снизились на 93% к 2000 г.
На снижение численности ряпушки повлияло и переключение хищных рыб с питания сигами на потребление ряпушки. Первыми на потребление ряпушки перешли кумжа Salmo trutta (пелагический хищник, она быстро начала питаться ряпушкой) и щука Esox lucius. В пищевом рационе этих хищников вместо прежних сигов стала преобладать ряпушка. В первые же годы появления ряпушки в водоеме на ее потребление перешел и окунь Perca fluviatilis, но ряпушка оказалась доступной только для крупного окуня. Налим Lota lota, как придонный хищник, живущий на больших глубинах, перешел на питание ряпушкой только спустя пять лет после ее появления (табл. 1).
Таблица 1.
Ряпушка в питании хищных рыб р. Пасвик в разные годы (F – частота встречаемости, %; Р – доля по весу, %; IR – комбинированный индекс, %)
| Годы | F | P | IR | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Щука | Окунь | Налим | Щука | Окунь | Налим | Щука | Окунь | Налим | |
| 1991–1992 | 1 | 2 | 0 | 0.8 | 1.0 | 0 | 0.1 | 0.7 | 0 |
| 1998–2005 | 13 | 6 | 0 | 10.8 | 4.6 | 0 | 8.3 | 2.0 | 0 |
| 2006–2010 | 31 | 4 | 43 | 34.5 | 7.6 | 58.2 | 39.8 | 1.5 | 57.9 |
После попадания ряпушки в р. Пасвик в структуре всего сообщества стали происходить существенные изменения, начиная с зоопланктона и кончая хищными рыбами (Попова и др., 2005; Решетников и др., 2008, 2011; Решетников, Попова, 2011; Amundsen et al., 1999, 2003; Bøhn et al., 2002; Liso et al., 2011; Popova et al., 2013; Reshetnikov, Popova, 2017).
Третий пример относится к обыкновенному ершу Gymnocephalus cernuus в водоемах Северной Америки. В начале 1980-х гг. ерш с балластными водами попал в систему Великих озер США (устье р. Сент-Луис в оз. Верхнее), где и образовал самовоспроизводящиеся популяции (Пронин и др., 1998; Обыкновенный ерш…, 2016; Brazner et al., 1998; Gunderson et al., 1998; Popova et al., 1998; Fullerton, Lamberti, 2006; Bauer et al., 2007). Точная дата попадания ерша в Америку неизвестна. Первые личинки были пойманы в 1986 г. (66 шт.). Уже в следующий 1987 г. было поймано много личинок и молоди ерша. В последующие годы ерш быстро увеличивал свою численность, и к 1991 г. он стал доминантным видом вдоль южного побережья оз. Верхнее на расстоянии около 200 миль. Нерестовое стадо ерша уже оценивалось в 200 тыс. рыб в 1989 г., а в 1991 г. оно достигло 1.8 млн. В эти годы ерш стал доминантным видом в траловых уловах в районе Сент-Луис (до 65% в уловах).
Одновременно с ростом численности ерша отмечалось падение численности некоторых аборигенных видов в 2–10 раз: желтый окунь Perca flavescens, лососеокунь Percopsis omiscomaycus, пестрый нотропис Notropis atherinoides, изумрудный нотропис N. hudsonius и др. (рис. 7).
Рис. 7.
Изменение плотности (экз./га) ерша и аборигенных видов рыб в эстуарии р. Сент-Луис (оз. Верхнее, США) (по: Пронин и др., 1998).

Это вызвало большое беспокойство среди американских ихтиологов, оценка ущерба оценивалась в 120–215 млн долл. Однако после того, как ерша стали активно потреблять хищные рыбы, его численность заметно снизилась (Обыкновенный ерш…, 2016; Gunderson et al., 1998). Тем не менее ерш уже сейчас встречается в трех озерах: Верхнее, Мичиган и Гурон.
Отметим, что и в случае попадания ерша в озера Северной Америки там последовала сначала вспышка численности вселенца, а потом ее падение. Однако, если нет ограничений в кормовой базе и есть возможность освоить новые биотопы и водоемы, то высокая численность нового вселенца может продолжаться дольше обычного, но в каждом из водоемов она имеет типичный характер резкого бума численности и затем столь же резкого падения. Это происходило и с ряпушкой во всей системе р. Пасвик, и с ершом в Северной Америке.
Можно привести много других примеров, когда появление нового вида сопровождается резкой вспышкой его численности, например бурное расселение ротана Perccottus glenii (Reshetnikov, 2010; Reshetnikov, Schliewen, 2013) или саморасселение тюльки Clupeonella cultriventris по системе волжских водохранилищ и т.п. (Слынько, Кияшко, 2012; Слынько, Терещенко, 2014). При проведении акклиматизационных работ в России это называлось эффектом акклиматизации (Карпевич, 1975; Решетников и др., 1982; Кудерский, 2001). Имеются примеры и по другим водоемам (Элтон, 1960; Биологические инвазии…, 2004; Odum, 1985; Moyle, Light, 1996; Williamson, 1996; Marco et al., 2002; Heger, Trepl, 2003; Inderjit et al., 2005). Некоторые авторы весь процесс вселения нового вида в экосистему подразделяли на несколько последовательных фаз (Карпевич, 1975; Слынько, Кияшко, 2012) или стадий (Inderjit et al., 2005): фаза проникновения, фаза обнаружения личинок, фаза быстрого роста численности и освоения биотопов и фаза прекращения экспоненциального роста численности и переход к фазе флуктуаций. В общем виде экологическая эффективность натурализации нового вида оценивалась по изменению разнообразия видов в сообществе, смене доминантных видов и по системе связей с доминантными видами (Одум, 1975; Слынько, Терещенко, 2014; Marco et al., 2002; Heger, Trepl, 2003; Wang et al., 2009).
Необходимо еще раз обратить внимание на то, что сама экосистема и ее жители активно сопротивляются и могут контролировать численность вида-вселенца. На рис. 8 представлены разные судьбы видов-вселенцев в новых экосистемах.
Рис. 8.
Численность вида-интродуцента в пресноводной экосистеме на примере рыб (по: Решетников и др., 1982). (а) – натурализация вида с относительно высокой численностью вселенца; (б) – натурализация вида с низкой численностью вселенца; (в) – медленное вытеснение вида из экосистемы при сильной конкуренции с местными видами и под воздействием хищников; (г) – новый вид быстро исчезает из экосистемы прежде всего из-за отсутствия условий для воспроизводства.

Можно привести не так много данных, когда пришелец приживался в экосистеме и его численность была на сравнительно высоком уровне (pис. 8а). Это зависит от его взаимодействия с другими членами сообщества, и должны быть пригодные места для нереста. Примером может служить удачная акклиматизация леща Abramis brama и судака Sander lucioperca в водоемах Западной Сибири, успешные работы по расширению ареала судака в Карелии и Вологодской обл. (Кудерский, 2001), саморасселение леща в р. Северная Двина, расширение ареала ротана Perccottus glenii в Европе (Reshetnikov, 2010) и др. (Рыбы в заповедниках…, 2010).
Если для нового вида характерна сильная конкуренция с аборигенными видами, или же он в значительной мере истребляется хищниками, то его численность держится на сравнительно невысоком уровне (рис. 8б). Если вселенец имеет возможность для воспроизводства в новом водоеме, но испытывает сильную конкуренцию со стороны аборигенных видов, а также если хищные рыбы активно переходят на питание новым видом, то вселенец или медленно исчезает, или его численность находится на предельно низком уровне, и в конце концов он обречен на вымирание (рис. 8в). Примером могут служить миграции из оз. Белое вниз по системе волжских водохранилищ ряпушки С. albula – до Саратовского и корюшки Osmerus eperlanus – до Волгоградского (Решетников, Попова, 2012). Если инвайдер не имеет хороших условий для размножения, то его численность быстро падает, и вид практически исчезает из экосистемы (рис. 8г). Иллюстрацией этого могут быть многие неудачные попытки вселения сиговых рыб в южные водоемы России. Существование здесь сиговых рыб возможно только при постоянной подсадке молоди (товарное сиговодство).
В последнее время нередко подчеркивается опасность от внедрения в экосистему чужеродного вида. Его появление рассматривается как инвазия или агрессия, способная разрушить экосистему. Всем хорошо известны плачевные результаты от инвазии гребневика мнемиопсиса Mnemiopsis leidyi в бассейны Каспийского и Черного морей, который вызвал резкие изменения во всей экосистеме, начиная с планктона и кончая рыбами (Shiganova, 2017). Хотелось бы еще раз подчеркнуть, что порой экосистема сама справляется с пришельцем, часто она сама сразу отторгает пришельца из-за несоответствия условий обитания в водоеме требованиям пришельца. Сегодня гребневик уже не так и страшен, нашлись организмы, способные сдерживать рост его численности. С другой стороны, акклиматизация камчатского краба Paralithodes camtschaticus в Баренцевом море и саморасселение тюльки по системе волжских водохранилищ имеет не только отрицательные, но и положительные стороны.
Имеется много примеров среди растений, животных и даже среди микроорганизмов, когда на первых этапах вселения наблюдается резкая вспышка численности (Элтон, 1960; Moyle, Light, 1996; Williamson, 1996; Begon et al., 2006). Все зависит от времени, через которое это наблюдается: для короткоцикловых видов это месяцы и годы, для длинноцикловых – десятки лет и даже столетия.
К сожалению, нет четкого понятия времени. Согласно энциклопедии (Новая философская…, 2018), время – это форма протекания всех механических, органических и психических процессов; это условие возможности движения, изменения и развития. Причем время тесно связано с пространством, и при искривлении пространства меняется и время в зависимости от скопления масс вещества и сил гравитации. По философским понятиям время не имеет ни начала ни конца, оно вечное, не исчезает и не появляется. Есть разные системы измерения времени (обыденное, солнечное, астрономическое, звездное, атомное). Отметим, что все живые организмы живут по своему внутреннему времени, конечно, они корректируют свою жизнь с сезонными и суточными ритмами (Алимов, Казанцева, 2004). Значит, у биологов должна быть своя единица времени для измерения продолжительности жизненных циклов животных, в экологии такой единицей измерения может служит смена поколения (τ-период). Если принять за единицу измерения время смены поколений τ, то у разных животных этот период имеет разную величину: у человека – это 18–20 лет, у крупных осетровых рыб типа белуги – 11–15 лет, у аквариумных рыб типа гуппи – 3–4 месяца, у микроорганизмов – часы и дни. Если для наших примеров принять эту единицу (τ-период), то все наши данные по изменениям биомассы и численности вида-вселенца в новом водоеме можно представить в виде общей кривой (рис. 9).
Рис. 9.
Изменение биомассы вида-вселенца в новом водоеме (время измеряется в τ-периодах, биомасса в условной величине уловов, римскими цифрами показаны фазы вселения).

Отметим, что экосистема переживает несколько различных фаз в соответствии с колебаниями численности нового вида в ходе его внедрения в экосистему водоема.
I. Латентный, или скрытый, период (медленное нарастание численности пришельца), освоение им разных биотопов и пищевых объектов, избегание хищников и нахождение пригодных для нереста мест. Этот период может быть довольно длительным: так, для тюльки в водохранилищах Волги – 5–10 лет, для ряпушки р. Пасвик он равен 10 годам, для сигов Севана – 40 годам. Чаще всего точная дата первого появления вселенца в водоеме остается неизвестной, поскольку мы судим только по первой поимке вида-вселенца. После первой поимки (начала увеличения численности) эта фаза длится примерно 4 τ-периода (рис. 9, I).
II. Фаза взрыва численности, или бума (период резкого увеличения численности и биомассы вселенца, так называемый эффект акклиматизации). Эта фаза также длится 3–5 τ-периодов (рис. 9, II).
III. Фаза резкого падения численности под воздействием хищников, паразитов и конкуренции с другими видами. Период столь же короток, как и фаза взрыва – примерно 4 τ-периода (рис. 9, III).
IV. Фаза вхождения пришельца в состав экосистемы (рис. 9, IV). Фаза длится несколько десятилетий или десятки τ-периодов; ее продолжительность зависит от длины жизненного цикла нового вида, кормовой базы и отношений с другими членами сообщества (обычно конкуренты, паразиты и хищники “ставят вселенца на место”).
Виды-вселенцы, которые не находят пригодных для нереста мест и не способны выдерживать конкуренцию с аборигенными видами, очень скоро исчезают из экосистемы. Таким образом, после вселения нового вида экосистема проходит несколько фаз, и все завершается “фазой стабилизации”: пришелец или совсем исчезает, или находит свое место среди других членов сообщества.
Отметим, что островершинную кривую (смена периодов II и III) можно наблюдать при ограниченной кормовой базе. Если корма много и вид осваивает все новые и новые биотопы или даже экосистемы, то фаза взрыва может продолжатся долго. Например, ерш в Великих озерах в каждом озере имел четкий пик, но в целом по всем Великим озерам пик получается растянутым. Аналогично и для ротана: в целом его завоевание Европы длится второе–третье десятилетие, но в каждом водоеме пик четко выражен.
Самый яркий пример – эпидемия холеры или чумы, которая сначала охватывает одну деревню, потом целый город, потом всю страну и даже целые континенты. И все это время численность микроба-интродуцента остается достаточно высокой, но в каждом населенном пункте вспышка длится недолго (пока не умрут или не уедут все люди). Нечто подобное отмечалось и на рыбах, например освоение тюлькой Сlupeinella cultriventris системы волжских водохранилищ или ротаном Perccottus glenii – мелких водоемов западной части России и всей Западной Европы (Reshetnikov, 2010). Если брать каждый отдельный водоем, то пик четко выражен, если брать район или регион, то максимум получается размытым.
Отметим еще одну особенность в жизни организмов. На примере многих видов растений и животных подмечено, что максимальная продолжительность жизни вида или популяции составляет примерно семь τ-периодов. Например, белуга Huso huso достигает половой зрелости в возрасте 14–17 лет, значит, ее максимальный возраст может быть 110–120 лет (в прошлом максимальный возраст отмечен в 118 лет, сейчас редко встречаются особи свыше 36 лет) (Рыбы в заповедниках…, 2010). Гуппи Poecilia reticulata созревает в возрасте 3–5 месяцев и живет 3–5 лет.
Отметим, что на основании этого соотношения между возрастом созревания и общей продолжительностью жизни (примерно семь τ-периодов) можно рассчитать и максимальный возраст человека. Если человек Homo sapiens достигает половой зрелости в 18–20 лет (ранние браки в 16 лет в счет не идут, так как даже у южных народов и в семьях аристократов в этом возрасте старались не иметь детей), то теоретически максимальный возраст человека может быть 130–140 лет (что совпадает с точкой зрения некоторых ученых-физиологов). Известны достоверные случаи, когда возраст жителей России превышал 120–130 лет: Тути Юсупова (1880 г.р.) из Каракалпакии умерла в 133 года (рекорд долголетия), Сархат Рашидова прожила в своем родном ауле 131 год, дедушка Магомед из Дагестана родился 1.05.1890 и умер в возрасте 123 лет (https://ru.wikipedia.org/wiki/Юсупова,_Тути_(долгожительница)).
ЗАКЛЮЧЕНИЕ
Таким образом, при инвазии нового вида в экосистему можно выделить 4 фазы вселения, причем заканчивается это или полным вытеснением вида из экосистемы, или вид переходит в фазу стабилизации. Весьма полезным оказалось применение τ-периода (время смены генераций) в качестве единицы времени, поскольку все живые организмы живут по своему внутреннему времени, конечно, согласовывая его с суточными и сезонными ритмами всей планеты. Но общая продолжительность жизни вида и даже каждой популяции зависит от времени смены поколений (τ-периода). Хотелось бы еще раз подчеркнуть, что порой экосистема сама справляется с пришельцем. После вселения нового вида экосистема проходит несколько фаз, и завершается все фазой стабилизации, когда экосистема “ставит пришельца на место”. Трудно предсказать заранее судьбу вида-пришельца в экосистеме. Мониторинговые исследования по инвазиям чужеродных видов предоставляют информацию не только о новом виде, но и о структуре и функционировании самой экосистемы, а также об адаптивных способностях видов-вселенцев – возможности изменять свою морфологию, жизненные циклы и генетическую структуру популяции в соответствии с новыми условиями обитания.
Список литературы
Алимов А.Ф., Казанцева Т.И. Рост животных и время // Докл. РАН. 2004. Т. 396. № 4. С. 561–563.
Берг Л.С. Рыбы пресных вод СССР и сопредельных стран. Ч. 1. М.; Л.: АН СССР, 1948. 467 с.
Биологические инвазии в водных и наземных экосистемах / Ред. А.Ф. Алимов, Н.Г. Богуцкая, М.И. Орлова. М.: КМК, 2004. 436 с.
Бурмакин Е.В. Акклиматизация пресноводных рыб в СССР. Л.: ГосНИОРХ, 1963. Т. 53. 317 с.
Герасимов Ю.В., Иванова М.Н., Свирская А.Н. Многолетние изменения роли местных и инвазийных видов рыб в питании хищных рыб Рыбинского водохранилища // Вопр. ихтиол. 2018. Т. 58. № 5. С. 507–522.
Дгебуадзе Ю.Ю. Экология инвазий и популяционных контактов животных: общие подходы // Виды-вселенцы в Европейских морях России / Ред. Г.Г. Матишов. Апатиты: КНЦ РАН, 2000. С. 35–50.
Иванова М.Н. Популяционная изменчивость пресноводных корюшек. Рыбинск: ИБВВ АН СССР, 1982. 145 с.
Карпевич А.Ф. Теория и практика акклиматизации водных организмов. М.: Пищевая промышленность, 1975. 342 с.
Карпевич А.Ф., Бокова Е.Н. Пересадка рыб и водных беспозвоночных, проведенная в СССР за 1960–1961 гг. // Вопр. ихтиол. 1963. Т. 3. С. 366–395.
Кудерский Л.А. Акклиматизация рыб в водоемах России: состояние и пути развития // Вопр. рыболов. 2001. Т. 2. № 1 (5). С. 6–85.
Методическое пособие по изучению питания и пищевых взаимоотношений рыб в естественных условиях / Ред. Е.В. Боруцкий. М.: Наука, 1974. 254 с.
Новая философская энциклопедия. 2018. https:// iphlib.ru/library/collection/newphilenc/document/ HASH-5940a150b1ca4a7297d259
Обыкновенный ерш Gymnocephalus cernuus (Linnaeus, 1758). Систематика, морфология, образ жизни и роль ерша в экосистемах / Ред. Ю.С. Решетников, О.А. Попова. М.: КМК, 2016. 279 с.
Одум Ю. Основы экологии. М.: Мир, 1975. 740 с.
Попова О.А., Решетников Ю.С. О комплексных индексах при изучении питания рыб // Вопр. ихтиол. 2011. Т. 51. № 5. С. 712–717. (Popova O.A., Reshetnikov Yu.S. On Complex indices in investigation of fish feeding // J. Ichthyol. 2011. V. 51. № 8. P. 686–691.)
Попова О.А., Решетников Ю.С., Амундсен П.-А. Включение вида-вселенца (ряпушка) в трофические цепи водоемов бассейна реки Пасвик // Чужеродные виды в Голарктике (Борок-2) / Междун. симп. по изучению инвазийных видов 27 сент.–1 окт. 2005. Борок: ИБВВ, 2005. С. 164–165.
Пронин Н.М., Селгеби Д.Х., Литвинов А.Г., Пронина С.В. Сравнительная экология и паразитофауна экзотических вселенцев в Великие озера мира: ротана-головешки (Perccottus glehnii) в оз. Байкал и ерша (Gymnocephalus cernuus) в оз. Верхнее // Сиб. экол. журнал. 1998. № 5. С. 397–406.
Решетников Ю.С., Попова О.А. Влияние вида-вселенца на экосистему реки Пасвик // Экологические проблемы пресноводных рыбохозяйственных водоемов России. СПб.: ГосНИОРХ, 2011. С. 294–298.
Решетников Ю.С., Попова О.А. Лососеобразные и корюшковые рыбы в водохранилищах Волги // Бассейн Волги в XXI-м веке: структура и функционирование экосистем водохранилищ. Ижевск: Издатель Пермяков, 2012. С. 236–239.
Решетников Ю.С., Попова О.А., Стерлигова О.П. и др. Изменение структуры рыбного населения эвтрофируемого озера. М.: Наука, 1982. 248 с.
Решетников Ю.С., Попова О.А., Амундсен П.-А. Влияние вида-вселенца на экосистему (на примере ряпушки р. Пасвик) // Водные экосистемы: трофические уровни проблемы поддержания биоразнообразия. Вологда: ВГПУ, 2008. С. 347–350.
Решетников Ю.С., Попова О.А., Амундсен П.-А. Структурные перестройки в экосистеме реки Пасвик // Биоиндикация в мониторинге пресноводных экосистем. СПб.: Любавич, 2011. С. 232–238.
Рыбы в заповедниках России. Т. 1. Пресноводные рыбы / Ред. Ю.С. Решетников. М.: КМК, 2010. 627 с.
Рыбы Рыбинского водохранилища: популяционная динамика и экология. Ярославль: Филигрань, 2015. 418 с.
Сидоров Г.П., Решетников Ю.С. Лососеобразные рыбы водоемов европейского северо-востока. М.: КМК, 2014. 346 с.
Слынько Ю.В., Кияшко В.И. Анализ эффективности инвазий пелагических видов рыб в водохранилища Волги // Рос. журн. биол. инвазий. 2012. Т. 3 (1). С. 73–87.
Слынько Ю.В., Терещенко В.Г. Рыбы пресных вод Понто-Каспийского бассейна. М.: Полиграф-Плюс, 2014. 328 с.
Слынько Ю.В., Дгебуадзе Ю.Ю., Новицкий Р.А., Христов О.А. Инвазии чужеродных рыб в бассейнах крупнейших рек Понто-Каспийского бассейна: состав, векторы, инвазионные пути и темпы // Рос. журн. биол. инвазий. 2010. Т. 1 (2). С. 119–126.
Стерлигова О.П. Корюшка Osmerus eperlanus и ее роль в ихтиофауне Сямозера // Вопр. ихтиол. 1979. Т. 19. Вып. 5. С. 793–800.
Стерлигова О.П., Павлов В.Н., Ильмаст Н.В. и др. Экосистема Сямозера: биологический режим, использование. Петрозаводск: КНЦ РАН, 2002. 119 с.
Элтон Ч. Экология нашествий животных и растений. М.: Иностранная литература, 1960. 232 с.
Amundsen P.-A., Staldvik F.J., Reshetnikov Yu.S., Popova O.A. Invasion of vendace Coregonus albula in a subarctic watercourse // Biol. Conserv. 1999. № 88. P. 405–413.
Amundsen P.-A., Bøhn T., Popova O.A. et al. Ontogenetic niche shifts and resource partitioning in a subarctic piscivore fish guild // Hydrobiologia. 2003. V. 497. P. 109–119.
Bauer C.R., Bobeldyk A.M., Lamberti G.A. Predicting habitat use and trophic interactions of Eurasian ruffe, round gobies, and zebra mussels in nearshore areas of the Great Lakes // Biol. Invasions. 2007. № 9. P. 667–678.
Begon M., Townsend C.R., Harper J.L. Ecology: from individuals to ecosystems. Oxford: Blackwell, 2006. 738 p.
Bøhn T., Amundsen P.-A., Popova O.A. et al. Predator avoidance by coregonids: can habitat choice be explained by size-related prey vulnerability // Adv. Limnol. 2002. V. 57. P. 183–197.
Brazner J.C., Tanner D.K., Jensen D.A., Lemke A. Relative abundance and distribution of ruffe (Gymnocephalus cernuus) in a Lake Superior coastal wetland fish assemblage // J. Great Lakes Res. 1998. V. 24. P. 293–303.
Dgebuadze Yu.Yu., Petrosyan V.G., Khlyap L.A. et al. Investigation of invasions of alien species, which are the greatest danger for ecosystems in the European part of Russia: pathways, vectors, biological features and methods of control // Invasion of alien species in Holarctic: book of abstracts. V Intern. Symp. Uglich–Borok, 25–30 sept., 2017 / Eds Yu.Yu. Dgebuadze, L.I. Tereshchenko, A.V. Krylov. Yaroslavl: Филигpaнь, 2017. P. 27.
Gunderson J.L., Klepinger M.R., Bronte C.R., Marsden J.E. Overview of the international symposium on Eurasian ruffe (Gymnocephalus cernuus) biology, impacts, and control // J. Great Lakes Res. 1998. V. 24. № 2. P. 165–169.
Fullerton A.H., Lamberti G.A. A comparison of habitat use and habitat-specific feeding efficiency by Eurasian ruffe (Gymnocephalus cernuus) and yellow perch (Perca flavescens) // Ecol. Freshwater Fish. 2006. V. 15. P. 1–9.
Heger T., Trepl L. Predicting biological invasions // Biol. Invasions. 2003. № 5. P. 313–321.
Inderjit, Cadotte M.W., Colautti R.I. The ecology of biological invasions: past, present and future // Invasive plants: ecological and agricultural aspects / Ed. Inderjit. Basel: Birkhäuser Verlag, 2005. P. 19–43.
Liso S., Gjeland R.Ø., Reshetnikov Y.S., Amundsen P.-A. A planktivorous specialist turns rapacious: piscivory in invading vendace Coregonus albula // J. Fish Biolog. 2011. V. 78. P. 332–337.
Marco D.E., Páez S.A., Cannas S.A. Species invasiveness in biological invasions: a modelling approach // Biol. Invasions. 2002. № 4. P. 193–205.
Moyle P.B., Light T.L. Biological invasions of freshwater: empirical rules and assembly theory // Biol. Conservat. 1996. V. 78. P. 149–161.
Odum E.P. Trends expected in stressed ecosystems // BioScience. 1985. V. 35. № 7. P. 419–422.
Popova O.A., Reshetnikov Yu.S., Kiyashko V.I. et al. Ruffe from the former USSR: variability within the largest part of its natural range // J. Great Lakes Res. 1998. V. 24 (2). P. 263–284.
Popova O.A., Reshetnikov Yu.S., Amundsen P.-A. Changes of “predator-prey” relations in the Pasvik river waterbodies after the vendace invasion // Invasion of alien species in Holarctic: book of abstracts / Eds Yu.Yu. Dgebuadze, Yu.V. Slynko, A.V. Krylov. Yaroslavl: Филигpaнь, 2013. P. 143.
Reshetnikov A.N. The current range of Amur sleeper Perccottus glenii Dybowski, 1877 (Odontobutidae, Pisces) in Eurasia // Rus. J. Biol. Invasions. 2010. № 1. P. 119–126.
Reshetnikov A.N., Schliewen U.K. First record of the invasive alien fish rotan Perccottus glenii Dybowski, 1877 (Odontobutidae) in the Upper Danube drainage (Bavaria, Germany) // J. App. Ichthyol. 2013. V. 29 (6). P. 1367–1369.
Reshetnikov Yu.S., Popova O.A. Phases of penetration of new species in freshwater ecosystems // Invasion of alien species in Holarctic: book of abstracts. V Intern. Symp. Uglich–Borok, 25–30 sept., 2017 / Eds Yu.Yu. Dgebuadze, L.I. Tereshchenko, A.V. Krylov. Yaroslavl: Филигpaнь, 2017. P. 102.
Reshetnikov Yu.S., Popova O.A., Amundsen P.-A. Introduction of vendace (Coregonus albula) in Pasvik river ecosystem // Invasion of alien species in Holartic: book of abstracts. III Intern. Symp. Borok–Myshkin, 5–9 oct., 2010. Yaroslavl, 2010. P. 79–80.
Salonen E. The vendace stock and fisheries in Lake Inari // Boreal Envir. Res. 1998. V. 3. P. 307–319.
Shiganova T.A. Dispersal of invasive ctenophores Mnemiopsis leidyi and Beroe spp. in the European seas // Invasion of alien species in Holarctic: book of abstracts. V Intern. Symp. Uglich–Borok, 25–30 sept., 2017 / Eds Yu.Yu. Dgebuadze, L.I. Tereshchenko, A.V. Krylov. Yaroslavl: Филигpaнь, 2017. P. 113.
Wang Q., Jin Sh., Ruan X. Ecological explanations for successful invasion of exotic plants // Front. Biol. China. 2009. V. 4. P. 271–281.
Williamson M.W. Biological invasions. L.: Chapman and Hall, 1996. 400 p.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии