

Успехи современной биологии, 2020, T. 140, № 6, стр. 616-623
Многолетние стационарные исследования в южнотаежных фитоценозах Ярославской области
Д. В. Татарников 1, *, А. А. Дерюгин 1
1 Институт лесоведения РАН
Московская обл., Одинцовский г.о., с. Успенское, Россия
* E-mail: orobus@land.ru
Поступила в редакцию 06.07.2020
После доработки 15.07.2020
Принята к публикации 15.07.2020
Аннотация
Охарактеризованы результаты многолетних исследований сотрудников Института лесоведения РАН в южнотаежных фитоценозах. При изучении факторов, определяющих типологическое разнообразие и различную продуктивность южнотаежных лесов, была установлена ведущая роль дефицита азота. Чем ниже было содержание азота в листьях или хвое деревьев по данным листового анализа, тем менее продуктивным был тип леса. Также была установлена тесная корреляция продуктивности местообитаний с длительностью и глубиной стояния грунтовых вод в почвенном профиле: чем лучше дренированность, тем выше продуктивность. Показана ограниченность корнеобитаемого объема почвы, его зависимость от гидрологического режима, концентрация сосущих корней в верхних горизонтах почвы. Определены запасы аммиачного азота в корнеобитаемом слое почвы основных типов леса, величины годичной аммонификации при минерализации опада. Анализ спонтанного хода демутационных процессов под пологом производных березняков показал, что рост подпологовой популяции ели протекает очень медленно и к моменту распада березового древостоя в возрасте естественной спелости (120 лет) из нее может образоваться разреженный, низкопродуктивный и низкотоварный ельник, состоящий преимущественно из ослабленных, пораженных стволовыми гнилями деревьев. Проведенные экспериментальные рубки березы в возрасте 50–60 лет с сохранением подроста ели, когда его численность была достаточной для формирования сомкнутого елового насаждения на большей части вырубленной площади, показали возможность в короткие сроки формирования высокопродуктивного хозяйственно ценного елового насаждения на месте проведенной рубки. Также установлено значительное влияние лося на ход сукцессионной динамики лесных фитоценозов в условиях его высокой численности.
ВВЕДЕНИЕ
Институт лесоведения РАН с момента своего основания проводит комплексные биогеоценологические исследования в разных природно-климатических условиях в пределах лесной зоны европейской части России. С этой целью была создана сеть стационаров в разных природных зонах как база для длительных наблюдений и долговременных экспериментов. В южной тайге с 1958 года функционирует Северная лесная опытная станция (ЛОС), объекты которой расположены в ряде районов Ярославской области. В данной статье предлагается краткий обзор основных результатов исследований, полученных на объектах Северной ЛОС, расположенных в Рыбинском районе Ярославской области, за время ее существования.
Основным направлением исследований южнотаежных фитоценозов на объектах Северной ЛОС в советское время была общая биогеоценотическая характеристика типологического разнообразия лесов с упором на детальное изучение факторов почвенного плодородия, так как стационары многие десятилетия были закреплены за отделом лесного почвоведения. Этими работами долгие годы руководил Александр Яковлевич Орлов.
В дальнейшем акцент был смещен на изучение сукцессионной динамики производных лесов, а также оценку эффективности вмешательства в ее ход с помощью экспериментальных рубок для получения значимых лесохозяйственных результатов. В этот период основные исследования на объектах Северной ЛОС осуществлялись силами отдела лесоводства под руководством Михаила Владимировича Рубцова.
ОБЪЕКТЫ И МЕТОДЫ
С момента основания Северной ЛОС в центре внимания было изучение типов леса, выявление тех абиотических факторов, которые определяют типологические различия лесов, их разную продуктивность. В 60–80-е гг. XX в. была изучена структура почвенного покрова, формирующегося в основных типах леса, рассмотрены особенности строения профиля почв и их физико-химические свойства. Проводилось исследование закономерностей размещения корневых систем в почвенном профиле, глубины распространения корней. В каждом типе леса был определен корнеобитаемый объем почвы. Дефицит элементов минерального питания в разных типах леса для основных лесообразующих пород был установлен по данным листового анализа. Большие работы проведены по изучению режима почвенно-грунтовых вод и динамики влажности почв в различных типах леса. Комплексные исследования позволили составить подробную характеристику типов лесных биоценозов южной тайги (Орлов и др., 1974; Абатуров и др., 1982) и разработать почвенно-экологические основы лесоводства для этих условий (Орлов, 1991).
В дальнейшем основным направлением исследований на объектах Северной ЛОС стал мониторинг долговременной спонтанной динамики производных лесов. В наиболее распространенных условиях местопроизрастания (кислично-черничная группа типов леса) была создана сеть постоянных пробных площадей (ПП) – 18 проб в березняках разного возраста (30–115 лет) с подпологовой елью и 8 – в насаждениях, формирующихся после рубки березовых древостоев. Березняки на ПП относятся к одному естественному возрастному ряду. В сеть ПП были включены ранее действовавшие опытные объекты, заложенные сотрудниками института во главе с А.Я. Орловым (Орлов, 1991) в период с 1977 по 1992 гг. На пробных площадях проводятся детальные учеты с периодичностью раз в десять лет. Для изучения хода роста деревьев ели под пологом березы и на вырубках после рубки березняков было взято соответственно 456 и 300 модельных деревьев. Более подробно методика исследований приведена в (Рубцов, Дерюгин, 2017; Дерюгин, 2017а).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В ходе обстоятельных исследований, продолжавшихся с 1958 года, были детально и тщательно изучены факторы, определяющие типологические различия южнотаежных лесов и их разную производительность. Для каждого типа еловых и сосновых лесов были определены запас доступных элементов минерального питания в лесных почвах, их распределение по горизонтам и обеспеченность ими фотосинтезирующих органов деревьев по данным листового анализа (Орлов и др., 1974). Полученные результаты показали, что основным почвенным ресурсом, определяющим различие южнотаежных типов леса по их производительности, является аммонийный азот. Листовой анализ показал, что в большинстве типов леса как ельников, так и сосняков, содержание азота в хвое ниже оптимального и снижалось от одного типа к другому параллельно с продуктивностью. Содержание других элементов не опускалось ниже нормы, лишь в сфагновом и долгомошном сосняках листовой анализ диагностировал также недостаток калия, но недостаток азота там был наибольшим. Оптимальное содержание азота в хвое было обнаружено только в неморальном ельнике.
Большое внимание было уделено изучению размещения в почве корневых систем. Оказалось, что большая часть корней и особенно их сосущих окончаний концентрируется в самых верхних горизонтах почвы. Так 100% сосущих окончаний ели в ельнике-черничнике сосредоточены в верхних 20 см почвы, в неморальном ельнике 99% сосущих корней ели располагаются в верхних 30 см. И в сосняках, несмотря на стержневой характер корневой системы сосны, картина существенно не меняется. В сосняке-кисличнике в верхних 30 см почвы находится 85% сосущих окончаний сосны, а глубже метра проникает только 3% корней (по весу). В сосняке-черничнике 80% корней располагается в слое 0–20 см, а глубже метра корни сосны вообще не проникают из-за периодического анаэробиоза (Орлов и др., 1974; Мамаев, 1977). Корни большинства травянистых растений также приурочены исключительно к поверхностным горизонтам (Татарников, 1996). В березняке-кисличнике виды травяно-кустарничкового покрова можно поделить на корнеподстилочные (звездчатка, кислица); поверхностнокорневые, чьи корневые системы почти не выходят за границы тонкого гумусаккумулятивного горизонта (майник, седмичник, голокучник, бор); неглубокоукорененные, чьи корневые системы ограниченны верхними 25 см почвы (черника, ожика волосистая, костяника); глубокоукорененные, активно осваивающие практически весь метровый слой почвы (щитовники игольчатый и австрийский, вейник наземный). Проведенные исследования позволили сформировать представление об ограниченности корнеобитаемого слоя почвы в условиях южной тайги.
Был подробно изучен водный режим почв (Орлов и др., 1974), оказывающий решающее влияние на размещение корней в почвенном профиле. Уровень стояния грунтовых вод во время вегетационного периода был тем выше, чем ниже была производительность типа леса, за исключением лишайниковых сосняков. Если в неморальном ельнике, ельниках и сосняках кисличных верхний слой почвы большую часть вегетационного сезона свободен от грунтовых вод и там складывается благоприятный для жизнедеятельности корней аэробный режим, то в ельниках и сосняках черничных уровень грунтовых вод стоит уже на глубине 70 см. При этом обогащение грунтовых вод кислородом происходит только после сильных дождей. Этот кислород быстро расходуется на дыхание корней и почвенных организмов, поэтому в теплое время года в переувлажненных почвах быстро возникают неблагоприятные для жизнедеятельности корней анаэробные условия. Во влажных типах леса, таких как сфагновые ельники и сосняки, грунтовые воды большую часть вегетационного сезона стоят недалеко от поверхности. В ряде экспериментальных работ была наглядно показана эффективность поверхностного осушения переувлажненных лесов для повышения их продуктивности (Вомперская, 1989; Орлов, 1991). Кроме того, почвенно-климатические исследования показали, что в условиях южной тайги на глубине больше метра температура почвы повышается до оптимальной для нормальной жизнедеятельности корней только в середине вегетационного сезона.
В ходе исследований азотного режима корнеобитаемого слоя почвы было установлено, что в слое 0–20 см ельника-черничника содержится 14 кг/га аммиачного азота (N), а в слое 0–30 см неморального ельника – 58 кг/га (Орлов и др., 1974). При недавних микробиологических исследованиях была определена годичная аммонификация в подстилке и верхних слоях почвы. В березняке-кисличнике она составила 58 кг/га N в год (Разгулин, 2017), а в неморальном березняке – 152 кг/га N в год (Разгулин, 2012). В неморальном березняке также была показана значительная роль нитрификации (19% от минерализованного азота), что необычно для зональных сообществ южной тайги. Эксперименты по внесению удобрений в березняке-кисличнике показали, что на внесение азотных удобрений (мочевины) неморальные травы реагировали положительно, тогда как бореальные – не показали значимой реакции (Татарников, 2017). На другие удобрения – фосфорные и магниевые – виды травяно-кустарничкового яруса не реагировали.
Изучение возрастной динамики производных березняков на разных объектах с учетом длительности наблюдений охватило период от возраста количественной спелости древостоя и завершения его интенсивного изреживания до достижения березовыми древостоями возраста естественной спелости перед началом распада (возрастной диапазон от 40 до 120 лет).
Естественная сукцессионная динамика производных лиственных древостоев в южной тайге связана с формированием под их пологом растущей генерации ели, которая в дальнейшем сменит лиственные породы. Появление подроста ели под пологом березняков зависит от наличия поблизости семеносящих генеративных деревьев ели. Их количество определяет активность обсеменения и скорость накопления численности растущего елового возобновления в березняках. Когда процесс возобновления ели протекает активно, к возрасту количественной спелости древостоев березы (45–50 лет) под ее пологом образуется густой (около 4.0 тыс. шт. га–1) подрост ели, в 70–80-летних березняках уже сформирован второй ярус ели с густотой около 1.2 тыс. шт. га–1, а к возрасту естественной спелости березы (120 лет) численность второго яруса ели значительно уменьшается, составляя всего 0.3–0.4 тыс. шт. га–1 при среднем возрасте елей 85 лет (Рубцов, Дерюгин, 2017). Такая динамика подпологовой генерации ели в березняках существенно отличается от динамики в зрелых ельниках. Там еловое возобновление, появляющееся под пологом материнского древостоя, обычно доживает только до 30–45 лет и сомкнутого яруса подроста не образует (Карпов и др., 1983).
Смыкание полога елового подроста ведет к обострению конкуренции и началу процесса изреживания растущей генерации ели. Установлено, что отпад ели преимущественно происходит за счет более молодых особей, которые появились позднее (Рубцов, Дерюгин, 2017). При сопоставлении изменений, происходящих в разных сообществах естественного ряда березняков кислично-черничной группы типов леса, обнаружено, что в среднем до 60% от всего количества погибших деревьев ели в процессе изреживания первой подпологовой генерации ели после ее смыкания приходится на подрост и 40% – уже на второй ярус древостоя, без учета гибели елового самосева в первые годы жизни.
Начало семеношения лидирующих деревьев ели первой генерации ведет к появлению под ее пологом всходов ели второй генерации. Однако их смертность очень высока из-за неблагоприятных фитоценотических условий под пологом двухярусного березово-елового древостоя. В березняках возрастом 110–120 лет численность второй генерации ели составляет лишь около 1.0 тыс. шт. га–1.
Для подробного анализа хода роста подпологовой генерации ели были изучены ростовые процессы у 456 модельных деревьев в интервале возрастов 11–77 лет, чей период возобновления (разница возрастов березы первого яруса и деревьев ели) изменялся от одного года до 58 лет. По периоду возобновления (Tw) все модели поделили на три группы с Tw 1–25, 26–40 и 41–60. Оказалось, что по росту в высоту деревья первой и второй групп почти не отличаются в их начальный период роста. Различия обнаруживаются только, когда деревья второй группы становятся старше 30 лет. Более молодые деревья третьей группы начинают испытывать прессинг со стороны ранее появившихся елей сразу и отстают в росте уже в первые годы жизни (Дерюгин, 2018).
Различия в росте деревьев этих трех групп, выделенных по периоду возобновления, становятся еще более очевидными при рассмотрении динамики текущего среднепериодического прироста в высоту (рис. 1). Все описанные различия наблюдаются в случае, когда в одном березняке появляются последовательно деревья всех этих трех групп.
Рис. 1.
Возрастная динамика текущего среднепериодического прироста деревьев ели в высоту в связи с периодом возобновления (Tw, лет) (Дерюгин, 2018).
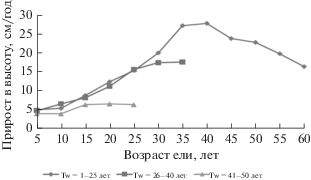
Приведенные данные свидетельствуют о том, что при спонтанном формировании подпологовой популяции ели доминируют деревья, появившиеся в числе первых. Они имеют наибольшие шансы дожить до распада березового древостоя. Из них преимущественно и формируется второй еловый ярус.
Значения всех показателей сомкнутого яруса ели под пологом березы существенно уступают значениям в свободнорастущих еловых древостоях того же возраста. Так, значения, характеризующие высоту в возрасте 60 лет, составляют у елей под пологом березы только 53–61% от значений в наиболее распространенных в регионе ельниках II класса бонитета. Значения площадей поперечного сечения стволов на высоте 1.3 м составляют у подпологовой ели в 60 лет только 25–38% от таковых в свободнорастущих ельниках. Еще более значительна разница средних объемов стволов (под пологом они составляют только 20–21% от значений в свободнорастущем ельнике того же возраста).
Кроме того, в подпологовой популяции ели деревья имеют более сбежистые стволы пониженной товарности. На состояние и сохранность деревьев подпологовой ели существенное влияние оказывают повреждения коры стволов лосем. Возникшие раны не зарастают в течение продолжительного времени и являются местом возникновения стволовой гнили, что в последующем часто становится причиной ветролома деревьев. Существенная часть деревьев ели под пологом поражается и комлевой гнилью.
Таким образом, к возрасту естественной спелости березняков (120 лет) под их пологом формируется разреженный (численностью около 400 шт. га–1), низкопродуктивный и низкотоварный второй ярус ели, из которого после распада березы не может сформироваться хозяйственно ценный высокопроизводительный еловый древостой, соответствующий условиям местопроизрастания.
Без проведения своевременных уходов ельники, спонтанно формирующиеся из подпологовой популяции, будут характеризоваться существенно меньшей производительностью (IV класса бонитета), чем ельники, изначально формирующиеся на вырубках без существенного участия мелколиственных пород. В таежной зоне европейской части страны, в условиях ограниченной возможности проведения лесовосстановительных работ, основным мероприятием по восстановлению на вырубках еловых древостоев является сохранение подроста и тонкомера ели предварительной генерации. В условиях Костромской и Кировской областей, расположенных в южной тайге, за счет сохранения при рубках подроста хвойные породы можно восстановить на 50% площади древостоев, поступающих в сплошную рубку (Побединский, 1973). Успешность данного мероприятия зависит от многих факторов. Это прежде всего технология лесосечных работ, количество, состояние и возраст ели предварительной генерации.
Какой эффект может дать своевременное удаление березы, когда под ее пологом имеется достаточное количество жизнеспособного елового подроста, показали результаты экспериментальных рубок, проведенных в рамках комплекса стационарных исследований на Северной ЛОС Института лесоведения РАН. Наиболее показательным оказался объект, где детальные наблюдения были начаты на второй год после рубки березы в возрасте 55 лет с сохранением елового подроста и продолжались двадцать лет. Технология рубок – метод узких лент, ширина пасек 35 м и волоков 5–6 м. Валку деревьев проводили бензиномоторными пилами под углом 35° к волоку, сучья обрубали на территории между волоками. Хлысты трелевали за вершину тракторами ТДТ-40.
Густота подроста ели сразу после рубки составляла около 3.5 тыс. шт. га–1, средние высота – 2 м и возраст – 25 лет. Наиболее представлены (около 40%) ели возрастом 21–40 лет. Возраст некоторых деревьев достигал 60 лет.
В начальный 10-летний период отпад главным образом определяется особенностью адаптации ели предварительной генерации к условиям вырубки и повреждением ее при рубке древостоев березы. Доля погибших деревьев увеличивается с повышением возраста и высоты ели. Так в возрастном интервале 61–70 лет отпад составлял 27%, а в интервале до 40 лет – 9–17%. Та же картина наблюдается при увеличении высоты деревьев ели (Рубцов, Дерюгин, 2015). При этом в отпаде преобладают деревья, которые изначально были наиболее представлены в возрастном и высотном распределениях.
При давности рубки больше 10 лет отпад деревьев ели предварительной генерации обусловлен главным образом повышением напряженности конкурентных отношений. Это объясняется интенсивным ростом ели, повышением сомкнутости крон и, как следствие, увеличением темпа дифференциации деревьев по росту и развитию. При этом в отпаде преобладают более молодые и отставшие в росте деревья ели.
Для оценки перспективности деревьев подроста и второго яруса для формирования будущего ельника был использован показатель Kh – отношение протяженности кроны к высоте дерева (Дерюгин, 2017б). Его рассчитывали по данным наблюдений через 12 лет после рубки березы. В интервале значений Kh меньше 0.70 преобладают особи, погибшие в первые 12 лет после рубки. К перспективным можно отнести деревья со значениями Kh > 0.70. Согласно выполненным расчетам, через 12 лет после рубки общее число перспективных деревьев на объекте исследований составляло 1.69 тыс. шт. га–1 (44% от общего числа елей).
По данным большого числа модельных деревьев был проведен анализ хода роста деревьев ели после рубки березового древостоя.
Время реакции деревьев ели на изменения условий среды в результате рубки березы в значительной мере определяется их возрастом. В первые 10 лет после рубки наилучшим образом реагируют деревья возрастом до 25 лет, их высота становится существенно больше, чем у деревьев под пологом. В последующие 10 лет это происходит с деревьями возрастом 26–40 лет. У деревьев более старших возрастов (41–60) лет увеличение прироста в высоту плавное и менее выраженное (Дерюгин, Глазунов, 2018).
Высота деревьев ели в год рубки, наряду с возрастом, также может быть использована для прогноза реакции деревьев ели на рубку мелколиственного древостоя. В результате анализа модельных деревьев по тенденции возрастной динамики высоты были выделены три высотные группы деревьев по высоте в год рубки березы (HR): 2.0 м и меньше (группа 1), 2.1–6.0 м (группа 2), больше 6.0 м (группа 3). Для выделенных высотных групп модельных деревьев была проанализирована динамика периодического (за 5 лет) прироста в высоту (ZH). Установлено, что деревья 1 и 2 групп реагируют на изменение условий среды в результате рубки уже в первые пять лет после ее проведения (рис. 2). Так, прирост деревьев группы 1 через 10 лет после рубки березы увеличивается почти в 7 раз, а у деревьев группы 2 – почти в 3 раза. У деревьев группы 3 реакция на рубку замедлена. В течение первых пяти лет их прирост в высоту оставался на уровне дорубочного периода. Максимальные значения периодического прироста (ZH = 160 см) и процента прироста (PH = 12%) наблюдались только через 15 лет после рубки (Дерюгин, Глазунов, 2018).
Рис. 2.
Динамика периодического (за 5 лет) прироста ели в высоту (ZH) до и после рубки березы (Дерюгин, Глазунов, 2018).
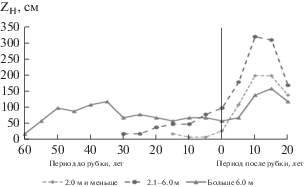
Существенное ослабление влияния рубки на рост ели предварительной генерации наблюдается во второе десятилетие после ее проведения. В последнее пятилетие ZH в выделенных высотных группах уменьшается в 1.3–1.8 раза. Сомкнутость крон деревьев ели увеличивается почти в 2 раза (с 0.40 до 0.79), что приводит к обострению внутривидовой конкуренции и, как следствие, к снижению прироста в высоту.
В формировании первого яруса будущих ельников преимущественно будут участвовать деревья второй и третьей высотных групп. Через 20 лет после рубки на их долю придется соответственно 62 и 37% численности деревьев первого яруса. Значительная часть деревьев первой высотной группы (45%) перейдет в отпад, несмотря на их бурный рост в первые годы после рубки березы.
Таким образом, через 22 года на объекте после рубки березы сформировалось высокополнотное (относительная полнота 0.78) 60-летнее елово-лиственное насаждение с общим запасом стволовой древесины 195 м3 га–1, из которых 158 м3 га–1 (81%) приходится на еловую древесину. Запас последней приближается к запасу среднеполнотного древостоя ели II класса бонитета. В этом возрасте запас ели, произрастающей под пологом березняка, составляет только 58 м3 га–1, а производительность соответствует IV–V классам бонитета.
Длительные наблюдения за ходом текущего возобновления ели в производных березняках с уже сформированным ярусом сомкнутого елового подроста, а также в молодых ельниках, возникших после рубки березового древостоя из подпологовой ели предварительной генерации, показали, что в этих фитоценозах дальнейшего пополнения существующей популяции ели новыми экземплярами не происходит ввиду крайне неблагоприятных фитоценотических условий для появляющегося там елового самосева. Весь самосев появляется там после семенных лет, когда происходит массовое семеношение ели. Но лишь малое число созревших семян получает шанс прорасти. Смертность появившегося самосева в первые годы жизни приближается к 100%. Для иллюстрации в таблице приведены данные об урожае семян в молодых ельниках, полученные на основе учета числа шишек в семенной год (приняв число зрелых семян в одной шишке примерно равным 200 штук), о числе появившихся там всходов ели на следующий год и о числе выживших экземпляров спустя три года (Татарников, 2019). Выжившие экземпляры приурочены исключительно к участкам, где есть разрывы в древесном пологе, а в сфагново‑черничном ельнике – и к участкам, где нет сомкнутого сфагнового покрова.
Таблица.
Урожай семян и число всходов ели в молодых ельниках (Татарников, 2019)
| Фитоценоз | Урожай семян ели в 2007 г., млн га–1 |
Число всходов ели в 2008 г., тыс.шт. га–1 |
Число трехлеток ели в 2011 г., тыс. шт. га–1 |
|---|---|---|---|
| Ельник кисличный | 3.6 | 330 | 11 |
| Ельник черничный | 6.6 | 330 | 13 |
| Ельник сфагново-черничный | 4.4 | 240 | 3.6 |
В результате многолетних наблюдений было выявлено разностороннее влияние лося на южнотаежные леса, их возрастную и сукцессионную динамику, особенно сильное в периоды его высокой численности. Наиболее сильными трансформирующими лесные фитоценозы воздействиями лося оказались:
1) массовое поедание лосем корневых отпрысков осины на молодых вырубках, что существенно меняет состав образующихся мелколиственных древостоев, усиливая преобладание березы над осиной;
2) постоянное объедание лосем молодых побегов рябины в южнотаежных лесах не дает сформироваться там сомкнутому пологу рябинового подлеска, в том числе в мелколиственных древостоях, что создает более благоприятные условия для поселения под их пологом елового подроста;
3) регулярные погрызы коры у молодых елей в результате приводят к значительному увеличению пораженности деревьев ели стволовыми гнилями (Смирнов, 1987, 2017).
Формирование в березняках сомкнутого полога елового подроста вызывает закономерные изменения травяно-кустарничкового и мохового ярусов. Смыкание крон растущего елового возобновления ведет к “бореализации” нижних ярусов: в их составе резко сокращается встречаемость и обилие светолюбивых и неморальных видов трав на фоне относительной стабильности видов бореального мелкотравья, кустарничков и папоротников. Таежные зеленые мхи – Pleurozium и Dicranum – увеличивают свою встречаемость и обилие (Татарников, 2017).
После полной вырубки сомкнутого елового подроста под пологом двух южнотаежных березняков-кисличников деградация травяно-кустарничкового покрова из-за жестких фитоценотических условий прекратилась. Наблюдения велись в течение 15 лет после вырубки подроста. Проективное покрытие травяно-кустарничкового яруса значительно выросло уже в первые годы после рубки, а затем стабилизировалось. Покрытие мхов сокращается вследствие разрастания черники и папоротников.
За период после рубки наблюдалось постепенное восстановление видовой насыщенности травяно-кустарничкового покрова, причем за счет тех видов, которые уже встречались на вырубленных участках в отдельных локациях. Примерно к десяти годам после рубки елового подроста флористический состав живого напочвенного покрова на отдельных площадках в пределах вырубленных участков существенно сближается.
Проведенные наблюдения динамики живого напочвенного покрова после вырубки сомкнутого елового подроста в южнотаежных березняках подтвердили, что наблюдающаяся в березняках южной тайги бореализация нижних ярусов является закономерным процессом, сопровождающим возрастное развитие березняков с еловым подростом, и связана с воздействием подпологовой ели на нижние ярусы фитоценоза (Татарников, 2018).
ЗАКЛЮЧЕНИЕ
Главным фактором, определяющим продуктивность южнотаежных лесов, является доступность минерального азота почвы, которая зависит от водного и воздушного режимов почв, определяющих условия функционирования корней и интенсивность процессов минерализации опада.
В березняках основа формирующегося под пологом ельника – первое поколение ели, возобновившееся в течение первых 25–30 лет жизни березняка. К возрасту естественной спелости березы (120 лет) из него формируется ельник низкой полноты (0.3) и производительности, соответствующей IV классу бонитета. Это низкопроизводительный и низкотоварный ельник, не соответствующий по производительности условиям произрастания.
Для ускорения смены березняков ельниками целесообразно проводить в березняках возрастом старше 50 лет рубку березы с сохранением ели первого поколения, если там имеется еловый подрост достаточной густоты. При оптимальном сочетании факторов (возраст насаждения, назначаемого в рубку; густота, состояние, возрастная и вертикальная структуры предварительной генерации ели, технологии лесосечных работ) такие рубки являются эффективным средством относительно быстрого восстановления высокопроизводительных хозяйственно ценных ельников в условиях южной тайги.
Список литературы
Абатуров Ю.Д., Зворыкина К.В., Ильюшенко А.Ф. Типы березовых лесов центральной части южной тайги. М.: Наука, 1982. 156 с.
Вомперская М.И. Поверхностное осушение временно переувлажненных лесных почв. М.: Наука, 1989. 134 с.
Дерюгин А.А. Некоторые результаты рубок березняков с сохранением подпологовой популяции ели // Комплексные стационарные исследования в лесах южной тайги (Памяти М.В. Рубцова). М.: КМК, 2017а. С. 172–187.
Дерюгин А.А. Динамика состояния популяции ели в насаждениях, формирующихся после рубки березовых древостоев с сохранением подроста // Лесохоз. информ. 2017б. № 1. С. 16–23. URL:http:// lhi.vniilm.ru/
Дерюгин А.А. Особенности роста популяции ели под пологом березняков в южной тайге Русской равнины // Лесохоз. информ. 2018. № 1. С. 21–30. URL:http://lhi.vniilm.ru/
Дерюгин А.А., Глазунов Ю.Б. Рост деревьев ели предварительного возобновления после рубки березняков в подзоне южной тайги Русской равнины // Вестн. Поволжского гос. технол. ун-та. Сер.: Лес. Экология. Природопользование. 2018. № 2 (38). С. 5–18.
Карпов В.Г., Пугачевский А.В., Трескин П.П. Возрастная структура популяции и динамика численности ели // Факторы регуляции экосистем еловых лесов. Л.: Наука, 1983. С. 35–62.
Мамаев В.В. Масса корней в сосняке и березняке кислично-черничном // Лесоводственные исследования в подзоне южной тайги. М.: Наука, 1977. С. 61–67.
Орлов А.Я. Почвенно-экологические основы лесоводства в южной тайге. М.: Наука, 1991. 104 с.
Орлов А.Я., Кошельков С.П., Осипов В.В., Соколов А.А. Типы лесных биогеоценозов южной тайги. М.: Наука, 1974. 232 с.
Побединский А.В. Рубки и возобновление в таежных лесах СССР. М.: Лесная пром-ть, 1973. 200 с.
Разгулин С.М. Минерализация азота в почве высокопродуктивного березняка южной тайги // Лесоведение. 2012. № 1. С. 65–71.
Разгулин С.М. Минерализация соединений азота в почве кислично-черничного березняка // Комплексные стационарные исследования в лесах южной тайги (Памяти М.В. Рубцова). М.: КМК, 2017. С. 47–61.
Рубцов М.В., Дерюгин А.А. Отпад деревьев после рубки древостоев березы с сохранением ели в южной тайге центральной части Русской равнины // Лесной журн. 2015. № 5. С. 9–25.
Рубцов М.В., Дерюгин А.А. Возрастная динамика морфоструктуры и состояния популяции ели под пологом южнотаежных березняков // Комплексные стационарные исследования в лесах южной тайги (Памяти М.В. Рубцова). М.: КМК, 2017. С. 125–154.
Смирнов К.А. Роль лося в биогеоценозах южной тайги. М.: Наука, 1987. 112 с.
Смирнов К.А. Трофическая деятельность лося как фактор формирования лесных экосистем // Комплексные стационарные исследования в лесах южной тайги (Памяти М.В. Рубцова). М.: КМК, 2017. С. 209–224.
Татарников Д.В. Вертикальная и временная структура травяно-кустарничкового покрова южнотаежного березняка в связи с конкуренцией // Экология. 1996. № 3. С. 225–227.
Татарников Д.В. Закономерности динамики травяно-кустарничкового покрова южнотаежных березняков // Комплексные стационарные исследования в лесах южной тайги (Памяти М.В. Рубцова). М.: КМК, 2017. С. 250–286.
Татарников Д.В. Динамика живого напочвенного покрова после рубки сомкнутого подроста ели под пологом двух южнотаежных березняков // Хвойные бореальной зоны. 2018. Т. 36. № 3. С. 238–242.
Татарников Д.В. Текущее возобновление ели в производных южнотаежных фитоценозах в ходе демутационных сукцессий // Хвойные бореальной зоны. 2019. Т. 37. № 6. С. 432–442.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии