

Успехи современной биологии, 2020, T. 140, № 6, стр. 601-615
Особенности структуры популяций карельской березы
Л. В. Ветчинникова 1, *, А. Ф. Титов 2, 3, **
1 Институт леса Федерального исследовательского центра “Карельский научный центр РАН”
Петрозаводск, Россия
2 Институт биологии Федерального исследовательского центра “Карельский научный центр РАН”
Петрозаводск, Россия
3 Отдел комплексных научных исследований Федерального исследовательского центра
“Карельский научный центр РАН”
Петрозаводск, Россия
* E-mail: vetchin@krc.karelia.ru
** E-mail: titov@krc.karelia.ru
Поступила в редакцию 24.04.2020
После доработки 06.05.2020
Принята к публикации 06.05.2020
Аннотация
Обобщены и систематизированы результаты исследований, касающиеся структуры популяций карельской березы Betula pendula Roth var. carelica (Mercklin) Hämet-Ahti, являющейся уникальным представителем лесной дендрофлоры на территории стран Балтийского макрорегиона. Дана характеристика пространственной, экологической, возрастной, половой и генетической структуры популяций карельской березы. Ее сравнительный анализ с близкородственными видами (березой повислой и березой пушистой) по рассмотренным показателям свидетельствует о том, что карельская береза существенно отличается от них, хотя и считается разновидностью березы повислой. По мнению авторов, это является еще одним важным аргументом в пользу ранее сделанного вывода о том, что карельская береза соответствует тем требованиям, выполнение которых позволяет рассматривать ее в качестве самостоятельного биологического вида.
ВВЕДЕНИЕ
На территории Евразии среди лиственных древесных пород наибольшее распространение получила береза повислая Betula pendula L., которая формирует здесь мелколиственные леса во всех климатических зонах, кроме тундры. Почти повсеместно березе повислой сопутствует береза пушистая B. pubescens Ehrh. В северо-западной части их ареалов произрастает также карельская береза Betula pendula Roth var. carelica (Mercklin) Hämet-Ahti, обладающая широкой известностью благодаря высокоценной узорчатой текстуре древесины, и которая, согласно существующей номенклатуре, считается разновидностью березы повислой. Ее появление исключительно на территории стран Балтийского региона, вероятно, явилось результатом особого направления в эволюции берез (Ветчинникова, Титов, 2016). Однако ее активная эксплуатация на протяжении ХХ века привела к резкому сокращению не только численности популяций карельской березы, но и занимаемой ею площади вплоть до полного исчезновения в ряде стран и регионов (Ветчинникова, Титов, 2018а,б, 2020а; Kosonen et al., 2004). Несмотря на повышенный к ней интерес, структура ее популяций по разным причинам остается до сих пор слабо изученной. Имеющиеся в этом отношении работы белорусских и российских исследователей, изучавших эколого-фитоценотические особенности произрастания карельской березы (Побирушко, 1992а), а также возрастную изменчивость в культурах и возможности селекционно-генетической оценки ее популяций (Курносов, 1993; Погиба и др., 2013), носят единичный характер.
Исходя из вышеизложенного, в данной работе нами предпринята попытка обобщения и систематизации собственных и литературных данных, касающихся структуры популяций карельской березы в местах естественного произрастания и выявления ее особенностей.
ПРОСТРАНСТВЕННАЯ СТРУКТУРА ПОПУЛЯЦИЙ КАРЕЛЬСКОЙ БЕРЕЗЫ
К настоящему времени в природных условиях карельская береза произрастает преимущественно на территории ряда стран Северной (Норвегия, Дания, Швеция, Финляндия) и Восточной Европы (Россия, Беларусь, Эстония, Украина) (Ветчинникова, Титов, 2019, 2020в). В середине ХХ века она также встречалась в Центральной Европе (Германия, Польша, Словакия и Чехия) (Lindquist, 1947, 1954; Hejtmánek, 1957; Vάclav, 1961, 1963; Scholz, 1963; Pagan, Paganová, 1994; Kosonen et al., 2004; Naujoks et al., 2017). В целом можно констатировать, что ареал карельской березы располагается исключительно на территории северо-западной части континентальной Европы (или стран Балтийского региона в широком смысле его понимания (Корнеевец, 2008)), но является дизъюнктивным. При этом на всем протяжении ареала лесов карельская береза не образует. Пространственное распределение деревьев носит случайный характер (по классификации: Грейг-Смит, 1967; Одум, 1986; Ипатов, Кирикова, 1997), причем в одних популяциях их количество исчисляется единицами, а в других – несколькими десятками и лишь изредка сотнями или тысячами (табл. 1). Не случайно, изучая распространение карельской березы на территории Словакии, Паган и Паганова (Pagan, Paganová, 1994) отмечают, что, как правило, обнаружив группу деревьев карельской березы, в дальнейшем при обследовании многих десятков гектаров трудно найти еще хотя бы одно дерево.
Таблица 1.
Численность популяций карельской березы в разных частях ее ареала (по: Ветчинникова, Титов, 2020а, с изменениями)
| Страна | Количество деревьев, площадь насаждения | Источник |
|---|---|---|
| Северная Европа | ||
| Дания | 2 | Rönne, 2005, устное сообщение; Ветчинникова и др., 2013 |
| Только в культуре | Наши данные, 2019 | |
| Норвегия | Единичные | Ruden, 1954; Hodnebrog, 1996а,b |
| Финляндия | Единичные ~100 шт. |
Heikinheimo, 1951; Kosonen, 2004 Hagqvist, Mikkola, 2008 |
| Швеция | Единичные | Johnsson, 1951; Lindquist, 1954 |
| 125 шт., 0.5 га | Ветчинникова и др., 1998; Martinsson, Vetchinnikova, 1999 | |
| 87 шт. | Emanuelsson, 1999; Побирушко и др., 1999 | |
| ~200 шт. | Ветчинникова и др., 2013, наши данные, 2018 |
|
| Центральная Европа | ||
| Германия | Единичные | Scholz, 1963 |
| Польша | 8.5 га | Scholz, 1963 |
| Единичные | Jakuszewski, 1970, 1973 | |
| ~100 шт. | Piroznikow, 2015, устное сообщение | |
| Словакия | Единичные | Scholz, 1963; Pagan, Paganová, 1994 |
| Чехия | Единичные | Hejtmanek, 1957; Vaclav, 1961 |
| Восточная Европа | ||
| Республика Беларусь | ~10 га | Соколов, 1959; Любавская, 1966 |
| ~40 тыс шт. | Побирушко, 1992а | |
| Латвия | ~400 шт. | Бандер, 1959; Сакс, Бандер, 1970 |
| Единичные | Наши данные, 2015 | |
| Литва | Единичные В культуре |
Vailionis, 1935; Сакс, Бандер, 1973, Backaitis, 2013, устное сообщение |
| Украина | Единичные >1000 шт. |
Соколов, 1950 Тарасевич, 2011 |
| Эстония | ~100 шт. | Sibul et al., 2011 |
| Россия | ||
| Республика Карелия |
> 5 тыс. шт. ~1700 шт. |
Соколов, 1950 Ветчинникова, Титов, 2018а,б, 2019 |
| Владимирская область | ~230 шт., 15 га | Шапкин и др., 1996 |
| Костромская область | 58 шт. | Багаев, 1963 |
| Ленинградская область | Единичные Только в культуре |
Соколов, 1950 Наши данные, 2017 |
| Псковская область | ~450 шт. | Николаева, Воробьев, 2016 |
| Смоленская область | Единичные > 900 шт. ~400 шт. 14 шт. |
Соколов, 1975 Евдокимов, 1978 Глушенко, 1988, устное сообщение Наши данные, 2018 |
| Ярославская область | Единичные | http://www.plantarium.ru/page/image/id/222946.html |
В настоящее время наибольшей численностью характеризуются популяции, находящиеся на территории Республики Беларусь, которые занимают площадь от нескольких сот квадратных метров до нескольких гектаров (от 0.2 до 30.0 га) (Сидор и др., 2016). Но и здесь, также как и в других странах, они распределены неравномерно (табл. 2). Согласно данным последних лет, наибольшая часть насаждений с участием карельской березы зафиксирована в Могилевской области (более 55%), наименьшая – в Брестской (менее 1%) (Сидор и др., 2016). Суммарный запас карельской березы в Республике Беларусь оценивается в 40 тыс. м3 , из которых для заготовки древесины рекомендовано около 15 тыс. м3 (табл. 2), причем в разных популяциях объем варьирует от 6 до 100 м3 /га (Пугачевский, 2008).
Таблица 2.
Распространение карельской березы на территории Республики Беларусь
| Область | Площадь насаждений с участием карельской березы | Запасы, тыс. м3 *** |
|
|---|---|---|---|
| в га* | в %** | ||
| Брестская | 42.3 | 0.8 | ~1.5 |
| Витебская | 563.9 | 24.7 | ~3.0 |
| Гомельская | 19.7 | 1.3 | < 0.1 |
| Гродненская | 9.8 | 4.6 | ~2.0 |
| Минская | 45.7 | 13.0 | ~6.0 |
| Могилевская | 13.7 | 55.6 | >2.5 |
| Всего | 695.1 | 100 | ~15.1 |
В связи с ограниченностью ресурсов и особенностями произрастания карельская береза в настоящее время считается редким растением. Более того, в соответствии с Красной книгой Российской Федерации и Красного списка МСОП (Международный союз охраны природы – International Union for Conservation of Nature and Natural Resources) (IUCN, 2012) она отнесена к категории 2/EN, то есть к числу исчезающих, находящихся в опасном состоянии видов (Красная книга Республики Карелия, 2007; Красная книга Владимирской области, 2010).
При характеристике популяций, как известно, ключевым показателем выступает величина их критической (эффективной) численности, ниже которой их длительное существование становится невозможным. К сожалению, точных данных о критических величинах численности популяций в литературе нет, а приводимые значения варьируют в широких пределах. Очевидно, это связано с тем, что при определении этой величины учитывают не все особи, а только активно участвующие в процессе воспроизводства. На основании анализа местоположения границ ареала карельской березы нами сделан вывод, что в условиях слабого влияния конкурентных отношений численность популяций карельской березы в 100–500 особей вполне способна обеспечить ее выживание и сохранность в течение нескольких поколений (Ветчинникова, Титов, 2020в). Но в случае более сильного влияния конкурентных отношений требуется существенно бóльшая численность популяции, возможно, от 1 тыс. деревьев и выше.
Важным показателем для характеристики популяций является также плотность, под которой обычно подразумевается среднее число особей на единицу занимаемой ими площади. Например, в 1980-е годы в Республике Карелия – на территории наибольшего распространения карельской березы в России – плотность естественных популяций карельской березы варьировала от 23.4 до 380 (в ботаническом заказнике “Анисимовщина”) деревьев на 1 га (Ветчинникова, Титов, 2019, 2020а), во Владимирской области – не более 15, в то время как в Беларуси – в среднем не менее 200.
Низкая плотность популяции, как известно, отрицательно сказывается на уровне воспроизводства особей, но способствует их выживанию, высокая, наоборот, благоприятна для репродукции, но усиливает внутривидовую конкуренцию. Исследования, проведенные на территории Республики Беларусь, показали, что численность взрослых особей карельской березы, соответствующих по густоте стояния деревьев среднеполнотному насаждению (0.6–0.7), способна обеспечивать относительно стабильное существование популяций, находящихся в условиях умеренной рекреационной нагрузки (Побирушко, 1992а,б; Барсукова, 1995; Сидор и др., 2016). При более высокой плотности лесного сообщества карельская береза из него выпадает.
ЭКОЛОГИЧЕСКАЯ СТРУКТУРА ПОПУЛЯЦИЙ КАРЕЛЬСКОЙ БЕРЕЗЫ
Как отмечено выше, карельская береза имеет локальный и довольно ограниченный ареал. При этом структура ее популяций формируется в разных природно-климатических условиях: на севере граница ареала доходит до 64° с.ш., на юге – до 48° с.ш. (в долготном направлении: от Финляндии–России (Республика Карелия) до Словакии – протяженность около 2.3 тыс. км), с запада – от 10° в.д. на восток – до 40° в.д. (в широтном направлении: от Дании до России (Костромская область) – протяженность около 2.5 тыс. км). Заметим, что ареал карельской березы в значительной степени перекрывается с ареалами березы повислой Betula pendula Roth и березы пушистой B. pubescens Ehrh., однако ее синэкологический оптимум отличается от последних, прежде всего отношением к уровню освещенности (Соколов, 1950; Евдокимов, 1989; Ветчинникова, 2005; Ветчинникова и др., 2013; Попов, 2017; Ветчинникова, Титов, 2020а; Mikkelä, 1992; Paganová, 2004). Карельская береза, как и береза повислая, является светолюбивой породой, однако в отличие от нее, не способна образовывать леса, расти в насаждениях с высокой плотностью и конкурировать с ней в местах их контакта (часто оказываясь в подчиненном ярусе).
Заметим, что карельская береза характеризуется высоким уровнем полиморфизма по целому ряду признаков, включая жизненную форму (или форму роста) (от одноствольного дерева до многоствольного дерева-куста и кустарника), тип поверхности ствола, насыщенность рисунка древесины и некоторые другие. Это прежде всего отражает биологические особенности данной древесной породы, придает ей определенную пластичность и расширяет возможности ее существования в различных экологических условиях. Так, во всех частях ареала карельской березы ведущая роль в формовом составе ее популяций принадлежит короткоствольной форме роста – в среднем около 60%, на долю высокоствольной приходится 10%, а кустообразная составляет 30% и выше, причем количество последних достоверно возрастает по направлению с севера на юг (Побирушко, 1992б; Пугачевский, 2008; Ветчинникова и др., 2013). В северной части ареала преобладают деревья с мелкобугорчатым типом поверхности ствола – до 70%, шаровидноутолщенный тип имеют около 25% особей, а ребристый – не более 5% (Евдокимов, 1989; Ветчинникова, Титов, 2019). В направлении к южной части ареала увеличивается количество деревьев с шаровидноутолщенным типом поверхности ствола и почти выравнивается с мелкобугорчатыми (50.2 и 49.2% соответственно), деревья с ребристой поверхностью ствола за редким исключением исчезают (0.6%) (Побирушко, 1992б). В целом, несмотря на территориальное разобщение популяций в долготном и широтном направлениях, зафиксировано значительное сходство деревьев по форме роста и типу поверхности ствола. Диаметр ствола, как правило, варьирует от 10 до 16 см, редко достигая 28–50 см и более при высоте до 18 м (Евдокимов, 1989; Ветчинникова и др., 2016; Pagan, Paganová, 1994). При этом радиальный прирост у нее не соотносится с возрастом (как, впрочем, и у других видов березы, но в меньшей степени). Вследствие этого даже в одновозрастном насаждении деревья по величине диаметра ствола могут различаться в несколько раз.
Деревья карельской березы, как правило, растут на значительном расстоянии друг от друга, хотя иногда произрастают относительно большими, но изолированными группами (Pagan, Paganová, 1994; Hodnebrog, 1996а,b). Она встречается в редко- и мелколесьях (~68–70%), обнаруживается на опушках леса и по берегам водоемов (~15%), а также на нелесных землях (заброшенных пастбищах или сенокосных угодьях) (~10%) и возле жилых строений. В Швеции и Беларуси отдельные деревья и небольшие группы до сих пор можно найти в придорожных полосах (~8%) (Побирушко и др., 1999; Ветчинникова и др., 2013; Martinsson, 1995; Emanuelsson, 1999). В Эстонии карельская береза отмечена на альварных почвах (безлесная поверхность, сложенная известняком), каменных грядах и других местах вдоль побережья Балтийского моря (Pork, Sander, 1973). В Латвии и в Беларуси ее изредка находили в дюнах и на откосах, где почвы преимущественно песчаные (Бандер, 1959; Побирушко, 1992а). На территории Польши и Беларуси участки с присутствием карельской березы ранее выделяли в так называемый “крестьянский лес”, который использовали в качестве лесного пастбища и для заготовки дров (Барсукова, 1987; Побирушко, 1992б; Пугачевский, 2008; Ветчинникова и др., 2013). В целом карельская береза обнаруживается в растительных сообществах с пониженной напряженностью конкурентных отношений и/или в той или иной мере подверженных антропогенному воздействию.
Накопленные к настоящему времени сведения об условиях произрастания карельской березы опровергают мнение о ее приуроченности к определенному типу почв. По мнению большинства специалистов, относительно невысокая требовательность карельской березы к почвенным условиям позволяет ей успешно расти на бедных песчаных и каменистых почвах (Соколов, 1950; Побирушко, 1992а; Scholz, 1963; Kosonen et al., 2004), поскольку почвенное питание у нее, как и у большинства других видов березы, осуществляется преимущественно за счет эктотрофной микоризы, которая не является видоспецифичной для отдельных древесных пород. Тот факт, что карельская береза часто произрастает в местах, менее благоприятных для других древесных пород, объясняется ее низкой конкурентоспособностью и необходимостью поиска незанятых ниш, причем с достаточно высокой освещенностью (Ветчинникова и др., 2013; Scholz, 1963; Hynynen et al., 2010).
На плодородных почвах она также хорошо растет, но при условии отсутствия конкуренции с другими, более быстрорастущими породами. Отсюда следует, что карельская береза имеет достаточно широкую эдафическую амплитуду, хотя и занимает вполне определенную экологическую нишу и характеризуется вполне определенными требованиями к среде обитания. Возможно, поэтому естественное возобновление карельской березы является в целом неудовлетворительным (Пугачевский, 2008; Ветчинникова и др., 2013).
К настоящему времени выявлены внутрипопуляционные взаимоотношения особей карельской березы между собой и другими древесными породами, и прежде всего, сопутствующими. Так, на основании изучения искусственно созданных насаждений показано, что в первое десятилетие растения карельской березы (особенно высокоствольной формы роста) развиваются довольно интенсивно и не уступают по высоте березе повислой (Ермаков, 1986; Евдокимов, 1989; Ветчинникова, 2004; Ветчинникова и др., 2013; Ветчинникова, Титов, 2018а). На данном этапе, по всей вероятности, это обусловлено активным развитием корневой системы и повышенным насыщением ризосферы микробным населением, продукты жизнедеятельности которых активно используются растениями. В дальнейшем по мере усиления затенения в результате смыкания крон рядом растущих безузорчатых особей или сопутствующих пород карельская береза снижает темпы роста и переходит в подчиненный ярус. Растения, имеющие короткоствольную и кустообразную формы роста, уже на этом этапе не выдерживают конкуренцию с другими более быстрорастущими лиственными породами и, как правило, отмирают. По-видимому, уступая сопутствующим породам в конкурентной борьбе, она теряет устойчивость к патогенной микофлоре (Пугачевский, 2008).
В условиях высокой плотности (обычно к 25–30-ти годам) сильное угнетение со стороны эдификаторов лесных сообществ сказывается и на развитии деревьев карельской березы высокоствольной формы роста. Степень их повреждения в насаждениях достигает 85% от общего числа деревьев, находящихся во втором ярусе (Пугачевский, 2008). Однако, замечено, что “пограничные” особи карельской березы, расположенные по периметру насаждений, отличаются более крупными размерами по сравнению с одновозрастными особями, расположенными внутри насаждений (рис. 1а). По всей вероятности, морфометрические характеристики растений карельской березы не слишком сильно зависят от площади почвенного питания, а более важным фактором, определяющим темпы их роста и накопление биомассы, выступает освещенность (Ветчинникова и др., 2013; Ветчинникова, Титов, 2018а; Hynynen et al., 2010). Подтверждением этому является, в частности, и тот факт, что у “пограничных” особей происходит изменение направления роста ствола и даже формы кроны: дерево становится наклонным и флагообразным в сторону открытых пространств и/или наибольшей освещенности, что не свойственно березе повислой (рис. 1а, б). В случае конкуренции за элементы минерального питания при высокой плотности в насаждении, очевидно, “включаются” механизмы самоизреживания, в результате которых наиболее сильные деревья занимают доминирующее положение. Особенностью карельской березы является то, что при отрицательном воздействии фитоценотической среды, небольшая часть ее популяции “уходит” из лесного сообщества в менее благоприятные для других древесных пород экологические ниши. Различия в скорости роста, наблюдаемые между растениями карельской березы и березы повислой, а также другими сопутствующими древесными видами, обусловлены, вероятно, воздействием разнонаправленных факторов и являются скорее следствием, чем причиной. Добавим, что у березы повислой с увеличением плотности насаждения усиливается скорость вертикального роста на фоне торможения радиального, при этом крона становится более компактной, что повышает ее конкурентоспособность в местах контакта с карельской березой и даже позволяет ей образовывать леса.

ВОЗРАСТНАЯ СТРУКТУРА ПОПУЛЯЦИЙ КАРЕЛЬСКОЙ БЕРЕЗЫ
К настоящему времени показано, что в популяциях карельской березы по возрастному составу преобладают средневозрастные (g2) (около 60%) и старые генеративные (g3) деревья (более 30%) (Побирушко, 1992; Пугачевский, 2008; Ветчинникова, Титов, 2018а). Высокая репродуктивная способность и большое количество семян могли бы способствовать активному семенному возобновлению карельской березы. Однако низкая выживаемость проростков (pl) и высокая гибель растений в ювенильном состоянии (j) (Бюллетень …, 2008) обусловливают почти полное отсутствие в популяциях виргинильных (v) и молодых генеративных особей (g1), количество которых на всем протяжении ареала составляет не более 5%. Вследствие этого у карельской березы к настоящему времени фактически отсутствуют популяции, включающие растения всех возрастных групп. Более того, в границах ее ареала количественное отношение прегенеративных растений к генеративным значительно меньше единицы, что говорит о том, что популяции карельской березы являются сокращающимися.
Наличие в благоприятных условиях довольно большого количества постгенеративных – субсенильных (ss) и сенильных (s) деревьев (около 10%) означает, что общий биологический цикл развития карельской березы (100 лет и более) примерно соответствует большинству основных видов рода Betula (100–140 лет) (Ветчиникова и др., 2013; Hynynen et al., 2010) и является не столь коротким, как считалось ранее (50–60 лет) (Ермаков, 1986; Raulo, Sirén, 1978 и др.).
Важно отметить, что образование узорчатой текстуры в древесине карельской березы также связано с возрастным состоянием растений. Так, начальные изменения в деятельности камбия у карельской березы могут происходить уже в первый год (Щетинкин, 1988; Щетинкин, Щетинкина, 2018), то есть в прегенеративный период на стадии имматурного (im) развития растений. На второй год, с началом ветвления, косвенные признаки узорчатой текстуры древесины визуально проявляются в виде утолщений (выпуклостей) в основании боковых побегов. В дальнейшем, на этапе виргинильного развития, в период активного роста, как правило, формирование узорчатой текстуры в древесине усиливается. Утолщения на поверхности ствола становятся особенно заметными у растений короткоствольной и кустообразной форм роста. У деревьев высокоствольной формы роста визуальное проявление косвенных признаков “узорчатости” наблюдается в возрасте 8–10 лет и часто совпадает с их переходом в генеративную фазу и/или с образованием бетулина в коре (Евдокимов, 1989; Курносов, 1998; Сидор и др., 2016).
Формирование узорчатой текстуры в древесине и ее внешнее проявление у карельской березы в значительной степени зависят также от плотности популяции. При благоприятных условиях ее косвенные признаки (в виде выпуклостей на поверхности ствола) становятся визуально заметными у одних растений уже в возрасте 2–5 лет (Соколов, 1970; Ветчинникова, 2005; Ryynänen, 1988; Hynynen et al., 2010 и др.), а у других – только в 20–25 (Сакс, Бандер, 1970, 1971(72); Соколов, 1970 и др.). При высокой плотности насаждения узорчатая текстура в древесине карельской березы может совсем отсутствовать.
Одной из важных биологических характеристик популяций древесных растений является их жизненное состояние (или виталитетная структура). К настоящему времени существует целый ряд классификаций, включающих разные диагностические признаки, и способы их расчетов (Алексеев, 1989; Санитарные …, 1998; Бебия, 2000 и др.). Для оценки жизненного состояния карельской березы, например, в Республике Беларусь – на территории наибольшего распространения карельской березы в мире – используется классификация, соответствующая “Санитарным правилам в лесах Российской Федерации” (1998). Показано, что в Беларуси доля нормально развитых здоровых особей в среднем составляет около 50%, причем среди одиночно растущих их количество может достигать 55% и выше, а в популяциях – только около 40% (Побирушко, 1992а). В Республике Карелия показатели жизненного состояния природных популяций оказались менее удовлетворительными. В частности, на территории ботанического заказника карельской березы “Каккоровский” к концу ХХ века здоровые деревья составляли не более 10% (рис. 2). Очевидно, это обусловлено преобладанием здесь субсенильных деревьев, которые вошли в группу “сильно ослабленных”.
Рис. 2.
Состояние деревьев карельской березы в ботаническом заказнике “Каккоровский” (Россия, Республика Карелия, Прионежский район, 1998 г.).
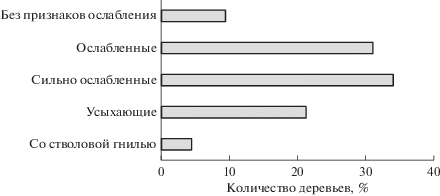
В целом, судя по возрастному и виталитетному состоянию, можно предположить, что помимо экологических особенностей местообитаний на современную структуру популяций карельской березы влияют также и генетические факторы.
ПОЛОВАЯ (ГЕНДЕРНАЯ) СТРУКТУРА ПОПУЛЯЦИЙ КАРЕЛЬСКОЙ БЕРЕЗЫ
Как известно, половое размножение у растений обеспечивается благодаря генеративным органам. Однако в отличие от животных они формируются многократно, как правило, с началом нового вегетационного сезона и с определенной периодичностью по годам. Это приводит к тому, что количество пестичных и тычиночных цветков у растений может варьировать не только от одного сезона к другому, но и среди цветков одной особи, между особями в одной популяции или разными популяциями вида. В.Н. Годин (Годин, 2007) выделяет два методологических подхода к изучению пола у растений, один из которых базируется на описании морфологических особенностей генеративных органов цветка, а другой – на оценке функционального состояния растений и включает не только производство гамет, но и вклад родителей в оплодотворение, образование семян, их рассеивание и жизнеспособность потомства следующего поколения. Тем не менее, до сих пор сведения о половой структуре популяций растений, включая древесные виды, остаются малочисленными и фрагментированными.
Карельская береза, подобно всем представителям рода Betula, относится к однодомным раздельнополым анемофильным перекрестноопыляемым растениям и размножается преимущественно семенами. Многочисленные наблюдения и экспериментальные данные, полученные в разные годы как в России, так и за рубежом, показывают, что при высокой концентрации деревьев (например, в специальных теплицах или при контролируемом опылении) доля растений с ярко выраженными признаками, характерными для карельской березы, составляет в потомстве 90% и более. Однако чаще всего места ее произрастания в естественных условиях находятся на значительном расстоянии друг от друга, поэтому обмен пыльцой между растениями не происходит или затруднен. По данным ряда авторов, пыльца у березы может распространяться в радиусе более чем на 103 км (Sofiev et al., 2006; Sofiev, 2017). Исходя из этого, можно было бы ожидать перемещения пыльцы (и/или семян) на значительные расстояния. Но реальная ситуация и особенности популяционной структуры древесных растений говорят о том, что даже при отсутствии каких-либо физико-географических преград этого не происходит (Динамика …, 2004; Kurganskiy et al., 2020). Существующий радиус распространения пыльцы (и семян), очевидно, значительно меньше, поскольку даже рядом растущие деревья являются преградой на ее пути. Более того, и жизнеспособность пыльцы карельской березы, как и других видов березы, резко падает с увеличением продолжительности и дальности ее переноса. Добавим, что далеко не все семена достигают подходящих для их прорастания микроклиматических условий, а их всхожесть заметно ослабевает уже на следующий год. Наконец, в небольших локальных и/или фрагментированных популяциях, которые характерны для карельской березы, наблюдается ограничение в количестве деревьев-опылителей. При совместном произрастании карельской березы с березой повислой, березой пушистой или другими видами березы между ними возможна гибридизация, но при устранении обычно существующей фенологической изоляции, что периодически происходит в отдельные годы (Ветчинникова, Титов, 2016). Подобные скрещивания, как известно, приводят к расщеплению признаков в потомстве. Поэтому в настоящее время, когда численность популяций карельской березы значительно сократилась, при свободном опылении вероятность появления растений с узорчатой древесиной в естественных условиях крайне невелика и может составлять всего 2–3%, в лучшем случае 25% или чуть больше.
В целом следует сказать, что изучение половой структуры популяций карельской березы требует более глубоких исследований, поскольку может дать более точное представление о родительском вкладе в следующие поколения.
ГЕНЕТИЧЕСКАЯ СТРУКТУРА ПОПУЛЯЦИЙ КАРЕЛЬСКОЙ БЕРЕЗЫ
Значительный прогресс, наблюдаемый в области молекулярной генетики, открывает новые возможности для изучения генетической структуры популяций и выявления генетических факторов, обусловливающих узорчатую текстуру древесины карельской березы. Однако, несмотря на то, что к настоящему времени в базе данных GenBank представлено более 25 тысяч нуклеотидных последовательностей, относящихся к экспрессируемым и не экспрессируемым областям ДНК разных видов березы (Баранов и др., 2018), информация по карельской березе пока крайне ограничена. Возможно, это связано с тем, что методические подходы к определению ДНК-маркеров представляют собой сборную группу методов, которые направлены на установление разных генетических параметров, например, локализацию генов в геноме (в ядерной, хлоропластной или митохондриальной ДНК), характер наследования (доминантный, кодоминантный), механизмы передачи (обоеполое наследование или только по линии одного из родителей) и уровень изменчивости (мономорфность, гипервариабельность). Как правило, изучение полиморфизма на уровне ДНК основано на выявлении разницы в цепях ДНК анализируемых объектов, которое может быть и количественным (варьирование длины ДНК-фрагментов образцов), и качественным (последовательность нуклеотидов в ДНК). Однако, следует иметь в виду, что при проведении молекулярно-генетических исследований полученные результаты во многом зависят не только от поставленных задач, но и от используемого оборудования (его аналитических возможностей), разрешающих способностей применяемых методов, квалификации персонала, биологических особенностей объекта изучения и др. (Матвеева и др., 2011; Ковалевич и др., 2015 и др.).
Возможно, в силу этих причин результаты секвенирования генома березы повислой (ближайшего сородича карельской березы), который включает от 30 до 40 тыс. генов (http://www.helsinki.fi/bio/faculty/news/2013/Genome_og_the_silver_ birch_mapped.httscience/metapop/EBFB/ru_index.html), впервые были опубликованы только в 2017 г., то есть через 4 года после осуществления секвенирования, когда была выявлена функциональная роль некоторых из генов (Salojarvi et al., 2017). Например, впервые обнаружены фитохромные гены PHYC и FRS10, которые являются основными медиаторами реакции растений на красный и дальний красный свет, определяющие в значительной степени рост и воспроизводство растений. Другие два гена, также впервые обнаруженные в геноме березы повислой, – KAK и MED5A – экспрессируются в клетках камбия и определяют соотношение ксилемы и флоэмы в ходе формирования сосудистой системы.
В настоящее время геномные исследования карельской березы ведутся в Финляндии, Польше, Республике Беларусь, России, но публикации пока единичны (Матвеева и др., 2008; Можаровская и др., 2018; Баранов и др., 2019). В частности, секвенирование хлоропластного генома карельской березы белорусскими учеными позволило выявить 134 кодирующих локуса, а их общий размер составил 161.1 тыс. пар нуклеотидов (Баранов и др., 2018; Кирьянов и др., 2018). Исследования показали, что по структурно-функциональной организации хлоропластного (а также и митохондриального) генома карельская береза очень схожа с березой повислой. Небольшие различия между ними выявлены исключительно в некодирующих областях, включая те, которые содержат короткие тандемно повторяющиеся последовательности ДНК (Кирьянов и др., 2018). Но учитывая особенности размножения карельской березы, для установления генетического контроля и характера наследования узорчатой текстуры в древесине, очевидно, более целесообразно изучение ядерной ДНК. К аналогичному выводу пришли, в частности, воронежские ученые, которые для молекулярно-генетической паспортизации и идентификации ценных для селекции генотипов березы (включая карельскую березу) предлагают применять ядерные микросателлитные локусы (Федулова и др., 2017; Исаков и др., 2017).
Проведенное нами сравнительное изучение генетического разнообразия внутри популяций карельской березы, расположенных на северной границе ее ареала, в условиях, сходных по природно-климатическим характеристикам на территории России (Республика Карелия, ботанические заказники “Анисимовщина” и “Каккоровский”) и Финляндии (район Пункахарью), позволило установить наибольшие и наименьшие значения по количеству аллелей (А) и гетерозиготности: наблюдаемой (HO) и ожидаемой (HE). Как оказалось, аллельное разнообразие по значениям ниже в российских популяциях (в среднем 7.2) по сравнению с финскими (7.6) (табл. 3). Вместе с тем, у карельской березы значения данного показателя значительно выше, чем у березы повислой (1–2) или березы пушистой (4 и ниже) (табл. 4) (Кирьянов и др., 2019).
Таблица 3.
Показатели генетического разнообразия популяций карельской березы в северной части ее ареала
| Страна (регион) | Популяция | Количество аллелей (А) |
Гетерозиготность | |
|---|---|---|---|---|
| наблюдаемая (HO) | ожидаемая (HE) | |||
| Россия (Республика Карелия) |
Анисимовщина-1 | 7.0 | 0.53 | 0.67 |
| Анисимовщина-2 | 8.2 | 0.47 | 0.70 | |
| Каккоровский-1 | 7.0 | 0.56 | 0.70 | |
| Каккоровский-2 | 6.5 | 0.46 | 0.67 | |
| В среднем | 7.2 | 0.50 | 0.69 | |
| Финляндия | Пункахарью | 7.6 | 0.59 | 0.75 |
Таблица 4.
Количество аллелей в SSR-локусах, выявленных у карельской березы, березы повислой, березы пушистой и гибрида между березой повислой (♀) и березой пушистой (♂)
| Локус | Карельская береза* | Береза повислая** | Береза пушистая** | Б. повислая (♀) х Б. пушистая (♂)** |
|---|---|---|---|---|
| L2.2 | – | 1–2 | 4 | 2–3 |
| L2.3 | 10 | – | – | – |
| L5.4 | 6 | 2 | 4 | 2–3 |
| L7.8 | – | 1–2 | 3–4 | 3 |
| L3.1 | – | 2 | 4 | 3 |
| L7.3 | 8 | 1–2 | 2–3 | 1 |
| L7.4 | – | – | – | – |
| L022 | 7 | 1–2 | 3–4 | 2–3 |
| L1.10 | – | 1–2 | 1–4 | 2–3 |
Примечание: * – по: Ветчинникова и др., 2012; ** – по: Кирьянов и др., 2019. Здесь и в табл. 5: – прочерк означает отсутствие данных.
Уровень гетерозиготности карельской березы в популяциях на северной границе ареала варьирует от 50 до 60% (НО) и/или от 69 до 75% (НЕ), а на южной – от 70 до 72% (Ветчинникова, Титов, 2018а; Кирьянов и др., 2019). Ранее с использованием изоферментного анализа трех ферментов (аланинаминопептидазы AAP, аспартатаминотрансферазы AAT и дегидрогеназы NADHdh-1) показано, что в условиях Южного Урала гетерозиготность березы повислой в локусе NADHdh-1 составляет 0.52 и 0.48 соответственно для НО и НЕ (Коновалов и др., 2001).
Анализ молекулярной изменчивости (AMOVA – Analysis of Molecular Variance) показал, что генное разнообразие внутри изученных популяций карельской березы составило 86%. Вместе с тем значения ожидаемой гетерозиготности (НЕ) превосходили значения наблюдаемой (НО) (табл. 3). Отрицательным последствием для перекрестноопыляемых видов в данном случае является то, что популяция становится все более гомозиготной (Ветчинникова и др., 2012; Ветчинникова, Титов, 2018а). При увеличении гомозиготности на 10% общая репродуктивная способность популяции может снизиться на 25% (Сулей, 1983). Более того, в небольших по численности популяциях существует достаточно высокая вероятность случайной утери редких аллелей (в том числе участвующих в формировании узорчатой текстуры древесины), которые в новом поколении могут отсутствовать уже у половины деревьев (Динамика …, 2004; Падутов и др., 2008).
Оценка генетической дифференциации популяций карельской березы, расположенных на территории Республики Карелия (Россия), проведена нами с помощью значений F-статистики Райта (FST), средняя величина которых оказалась равной 0.14. Полученные результаты выявили не только значительную их дифференциацию, но и наличие в них большого числа близкородственных скрещиваний (Ветчинникова и др., 2012). Обнаруженное уменьшение доли гетерозигот, по-видимому, свидетельствует о пониженной выживаемости растений в изученных популяциях (Pannell, Barrett, 2001). Выявленные популяционно-генетические особенности позволяют заключить, что зафиксированное снижение гетерозиготности при низкой эффективной численности популяций карельской березы ведет к обеднению генофонда, и в дальнейшем это может привести к его полной деградации.
Существенные различия в генетической структуре популяций карельской березы и березы повислой, произрастающих на территории Республики Беларусь, были также выявлены с помощью изоферментного анализа (13 ферментных систем, находящихся под контролем 20 генов) (Баранов, Марковская, 2003). В частности, с использованием метода UPGMA (Unweighted Pair Group Method with Arithmetic mean – метода оценки попарного внутригруппового невзвешенного среднего), показано, что в условиях Республики Беларусь карельская береза обособлена от березы повислой. Наибольшее расхождение было отмечено по аллозимному локусу Gpi-2 (глюкозофосфатизомераза), у которого в популяциях березы повислой частоты аллелей 1.00 и 1.30 составляли соответственно 77.2% и 18.6%, а у карельской березы – 47.9 и 51.1% (Баранов, Марковская, 2003). Добавим, что у большинства деревьев карельской березы (более 90%) ген Gpi-2 находится в гетерозиготном состоянии по аллелям 1.00 и 1.30 или гомозиготном – по аллелю 1.30, а для березы повислой более характерно присутствие аллеля 1.00. Отличия, выявленные у карельской березы по гену Gpi-2, авторы объясняют ее гибридным происхождением и обособленностью от березы повислой, а относительно высокий процент у последнего аллеля 1.30 (18.6%) – существованием генного потока между видами.
ЗАКЛЮЧЕНИЕ
Исследования последних десятилетий убедительно доказывают необходимость изучения структурной организации популяций не только основных видов лесной дендрофлоры (Семериков, Казанцев, 1979; Заугольнова и др., 1993; Динамика …, 2004; Ставрова, 2012; Санников и др., 2012, 2014; Фардеева и др., 2008 и др.), но и редких видов, находящихся на грани исчезновения в результате антропогенных воздействий и глобального изменения климата (Заугольнова и др., 1993; Злобин и др., 2013; Hamrick et al., 1992). Среди них важное место занимает карельская береза Betula pendula var. carelica, древесина которой, благодаря уникальным декоративным свойствам ценится особенно высоко. Исторически ареал карельской березы сформировался исключительно на территории стран Балтийского региона благодаря особому сочетанию климатических, геоморфологических и эдафических условий в данном макрорегионе (Ветчинникова, Титов, 2016). Возможно, изначально он занимал единое пространство, но затем в процессе эволюции приобрел явно выраженную дискретность.
Результаты собственных многолетних исследований и анализ литературы позволили представить в данной работе характеристику современной структуры природных популяций карельской березы, которая, как нам представляется, сложилась главным образом под влиянием антропогенных и фитоценотических факторов. Анализ структуры популяций карельской березы и ее сопоставление с ближайшими сородичами – березой повислой и березой пушистой, позволил также выявить у нее целый ряд важных особенностей (табл. 5). В частности, в отличие от них, у карельской березы в результате эволюционных изменений сформировался и закрепился качественно другой морфотип, обеспечивший ей собственные способы и механизмы приспособления к вполне определенным местообитаниям. Например, благодаря полиморфизму по форме роста (от деревьев до кустарников) у нее расширилась норма реакции и способность адаптироваться к разным экологическим условиям и местообитаниям, включая малопригодные для других древесных видов, а прочная узорчатая древесина способствовала усилению механической функции ствола деревьев (за счет разрастания клеток паренхимы и их склерификации) и появлению возможности депонирования (а при необходимости их быстрого расходования) повышенного количества запасных веществ. Однако, несмотря на это, в последние десятилетия во многих регионах наблюдаются резкое снижение численности карельской березы и занимаемой ею площади, причины которых нами подробно описаны (Ветчинникова, Титов, 2020а). Здесь же отметим лишь то, что из-за практически полного отсутствия естественного возобновления на всем протяжении ее ареала происходит снижение жизнеспособности популяций в целом. В возрастной структуре современных популяций карельской березы преобладают средневозрастные (g2) и старые генеративные (g3) особи. Проведенный нами микросателлитный анализ ДНК выявил сравнительно высокий уровень генетической гетерогенности в популяциях карельской березы, расположенных на северной границе ее ареала, что обусловлено в том числе и характером ее размножения как ветроопыляемого растения. В то же время, значительная межпопуляционная дифференциация (FST = 0.14), зафиксированная в условиях Карелии, по всей вероятности, связана с пространственной изолированностью популяций и усилением в них инбридинга (Ветчинникова и др., 2012; Ветчинникова, Титов, 2018а). Следовательно, несмотря на то, что карельская береза обладает достаточно высоким адаптивным потенциалом, продолжающееся в настоящее время сокращение ее численности и наличие близкородственных скрещиваний создают реальную угрозу не только в плане существенного снижения генетического разнообразия, но и деградации ее популяций.
Таблица 5.
Некоторые характерные особенности структуры популяций карельской березы, березы повислой и березы пушистой
| Структура популяций | Карельская береза | Береза повислая | Береза пушистая |
|---|---|---|---|
| Пространственная | |||
| Ареал | Дизьюнктивный, локальный | Сплошной, широкий | Сплошной, широкий |
| Пространственное размещение в фитоценозе | Лесов не образует | Лесообразующая порода, верхний ярус древостоя | Нижний полог древостоя |
| Плотность | Низкая | Не имеет значения | Средняя |
| Экологическая | |||
| Экологическая ниша | Открытые места, часто малоблагоприятные для других древесных видов | Тенистые леса. Сухие и бедные местообитания | Полуоткрытые места, влажные и средне богатые |
| Конкурентоспособность | |||
| внутри фитоценоза | Крайне низкая | Высокая | Средняя |
| на границе фитоценоза | Средняя | Высокая | Высокая |
| Жизненная форма | От дерева до кустарника | Дерево | Дерево, редко куст |
| Возрастная | |||
| Возрастные группы | Преобладают средневозрастные (g2) и старые генеративные (g3) деревья | От ювенильных (j) до сенильных (s) | От ювенильных (j) до сенильных (s) |
| Длительность жизненного цикла | 100 лет и более (35–50 лет – при высокой плотности) | 120–150 лет | 100–120 лет |
| Жизненное состояние (виталитет) | Депрессивное | Процветающее | Равновесное |
| Половая | |||
| Тип размножения | Семенной, анемофильное опыление |
Семенной, анемофильное опыление | Семенной, анемофильное опыление |
| Наследование отличительных признаков таксона | Устойчивое (при наличии опылителей) | Устойчивое | Устойчивое |
| Вероятность интрогрессии | Высокая | Низкая | Низкая |
| Генетическая | |||
| Степень инбридинга | Повышенная | Низкая | Низкая |
| Гетерозиготность | 0.70–0.72 | 0.45–0.50 | – |
| Число аллелей в изученных SSR-локусах |
6–10 | 1–2 | 2–4 |
Другой важный вывод, к которому мы приходим на основании проведенного анализа, заключается в том, что карельская береза, которая по-прежнему считается разновидностью березы повислой, по сути существенно от нее отличается по целому ряду важных биологических характеристик, и это служит еще одним аргументом в пользу ранее сделанного вывода (Ветчинникова, Титов, 2020б) о том, что ее вполне можно рассматривать в качестве самостоятельного биологического вида. На практике это означало бы расширение возможностей охраны этого уникального биологического объекта, так как по существующим правилам большинство ограничений, которые вводятся в отношении использования человеком редких и исчезающих растений касаются только тех из них, которые обладают видовым статусом.
Список литературы
Алексеев В.А. Диагностика жизненного состояния деревьев и древостоев // Лесоведение. 1989. № 4. С. 51–57.
Багаев С.Н. Карельская и капокорешковая береза в лесах Костромской области // Лесн. хоз-во. 1963. Вып. 6. С. 20–22.
Бандер В.Л. Карельская береза в Латвийской ССР // Тр. Латвийской с.-х. акад. 1959. Вып. 8. С. 353–365.
Баранов О.Ю., Марковская Ю.А. Особенности генетической структуры березы карельской по гену Gpi-2 // Проблемы лесоведения и лесоводства. Гомель: ИЛ НАН Беларуси, 2003. Вып. 50. С. 181–185.
Баранов О.Ю., Кирьянов П.С., Пантелеев С.В., Падутов В.Е. Высокопроизводительное секвенирование хпДНК карельской березы // Лесное хоз-во / Тез. науч.-техн. конф. Минск: БГТУ, 2018. С. 52–53. https://elib.belstu.by/handle/123456789/25625
Баранов О.Ю., Кирьянов П.С., Пантелеев С.В. и др. Анализ структурно-функциональной организации хлоропластного генома карельской березы на основании данных высокопроизводительного секвенирования // Докл. национальной академии наук Беларуси. 2019. Т. 63. № 3. С. 312–316. https://doi.org/10.29235/1561-8323-2019-63-3-312-316
Барсукова Т.Л. Береза карельская в Белоруссии // Интенсификация лесн. хоз-ва в Белорусской ССР. М.: ВНИИЛМ, 1987. С. 142–149.
Барсукова Т.Л. Изменчивость, отбор и разведение березы карельской в Беларуси: Автореф. дис. … канд. с.-х. наук. Гомель: Ин-т леса АН Беларуси, 1995. 21 с.
Бебия С.М. Дифференциация деревьев в лесу, их классификация и определение жизненного состояния древостоев // Лесоведение. 2000. № 4. С. 35–43.
Бюллетень экологических исследований на территории музея-заповедника “Кижи”, 2007 год: программа экологического мониторинга музея-заповедника “Кижи”. Петрозаводск: Издательский центр музея-заповедника “Кижи”, 2008. С. 25–29.
Ветчинникова Л.В. Береза: вопросы изменчивости. М.: Наука, 2004. 183 с.
Ветчинникова Л.В. Карельская береза и другие редкие представители рода Betula L. М: Наука, 2005. 269 с.
Ветчинникова Л.В., Титов А.Ф. Происхождение карельской березы: эколого-генетическая гипотеза // Экол. генетика. 2016. Т. 14. № 2. С. 3–18. https://doi.org/10.17816/ecogen1423-18
Ветчинникова Л.В., Титов А.Ф. Ботанические заказники карельской березы в Республике Карелия: история, современное состояние и проблемы // Ботан. журн. 2018а. Т. 103. № 2. С. 256–265. https://doi.org/10.1134/S0006813618020096
Ветчинникова Л.В., Титов А.Ф. Роль особо охраняемых природных территорий в сохранении генофонда карельской березы // Тр. КарНЦ РАН. Серия Экол. исслед. 2018б. Т. 10. С. 3–10. https://doi.org/10.17076/eco912
Ветчинникова Л.В., Титов А.Ф. Карельская береза – уникальный биологический объект // Успехи соврем. биол. 2019. Т. 139. № 5. С. 412–433. https://doi.org/10.1134/S0042132419050107
Ветчинникова Л.В., Титов А.Ф. Современное состояние ресурсов Betula pendula var. carelica (Betulaceae) // Раст. ресурсы. 2020а. Т. 56. Вып. 1. С. 16–33. https://doi.org/10.31857/S0033994620010082
Ветчинникова Л.В., Титов А.Ф. Карельская береза: разновидность или самостоятельный вид? // Изв. высш. уч. зав. / Лесн. журн. 2020б. № 1. С. 26–48. https://doi.org/10.37482/0536-1036-2020-1-26-48
Ветчинникова Л.В., Титов А.Ф. О границах ареала карельской березы // Изв. высш. уч. зав. / Лесн. журн. 2020в. № 6 (в печати).
Ветчинникова Л.В., Мартинссон У., Побирушко В.Ф. Результаты рекогносцировки популяций березы карельской в Центральной Швеции // Научные основы селекции древесных растений Севера. Петрозаводск: КарНЦ РАН, 1998. С. 137–142.
Ветчинникова Л.В., Титов А.Ф., Кузнецова Т.Ю. Карельская береза: биологические особенности, динамика ресурсов и воспроизводство. Петрозаводск: КарНЦ РАН, 2013. 312 с.
Ветчинникова Л.В., Титов А.Ф., Топчиева Л.В., Рендаков Н.Л. Оценка генетического разнообразия популяций карельской березы в Карелии с помощью микросателлитных маркеров // Экол. генетика. 2012. Т. 10. Вып. 1. С. 34–37.
Годин В.Н. Половая дифференциация у растений. Термины и понятия // Журн. общ. биол. 2007. Т. 68. № 2. С. 98–108.
Грейг-Смит П. Количественная экология растений. М.: Мир, 1967. 359 с.
Динамика популяционных генофондов при антропогенных воздействиях / Ред. Ю.П. Алтухов. М.: Наука, 2004. 619 с.
Евдокимов А.П. Эколого-биологические свойства и обоснование методов выращивания карельской березы: Автореф. дис. … канд. с.-х. наук. Л.: ЛТА, 1978. 20 с.
Евдокимов А.П. Биология и культура карельской березы. Л.: Изд-во ЛГУ, 1989. 228 с.
Ермаков В.И. Механизмы адаптации березы к условиям Севера. Л.: Наука, 1986. 144 с.
Заугольнова Л.Б., Денисова Л.В., Никитина С.В. Подходы к оценке состояния ценопопуляций растений // Бюл. Моск. общества испытателей природы. Отдел биол. 1993. Т. 98. № 5. С. 100–108.
Злобин Ю.А., Скляр В.Г., Клименко А.А. Популяции редких видов растений: теоретические основы и методика изучения. Сумы: Университетская книга, 2013. 439 с.
Ипатов В.С., Кирикова Л.А. Фитоценология. СПб.: СПбГУ, 1997. 316 с.
Исаков И.Ю., Федулова Т.П., Исаков Ю.Н. и др. Применение постгеномных технологий при идентификации жизненных форм карельской березы // Научно-практ. журн. 2017. Т. 20. № 4. С. 212–218.
Кирьянов П.С., Баранов О.Ю., Можаровская Л.В. и др. Анализ генетико-таксономических отношений карельской березы с представителями семейства Betulaceae на основании данных секвенирования хлоропластного генома // Перспективы развития и проблемы современной ботаники / Мат. Всерос. молод. конф. Новосибирск: Академиздат, 2018. С. 95–97.
Кирьянов П.С., Баранов О.Ю., Маслов А.А., Падутов А.В. Молекулярно-генетические подходы к идентификации межвидовых и внутривидовых гибридов берез Восточно-Европейского региона // Мол. и приклад. генетика. 2019. Т. 26. С. 45–55.
Ковалевич А.И., Падутов В.Е., Баранов О.Ю. Полногеномное секвенирование – новый этап генетических исследований // Наука и инновации. 2015. № 5 (147). С. 56–58.
Коновалов В.Ф., Галеев Э.И., Янбаев Ю.А. Генетическая дифференциация популяций березы повислой на Южном Урале // Вестн. МГУЛ. Лесной вестник. 2001. № 5. С. 62–67.
Корнеевец В.С. Понятия “страны Балтийского региона” и “Балтийский регион” // Космополис. 2008. № 2 (21). С. 68–77.
Красная книга Республики Карелия. Петрозаводск: Карелия, 2007. С. 45–46.
Красная книга Владимирской области. Владимир: Транзит-ИКС, 2010. С. 95.
Курносов Г.А. Возрастная изменчивость березы карельской в культурах центральной части зоны смешанных лесов: Автореф. дис. … канд. с.-х. наук. М.: МГУЛ, 1993. 18 с.
Курносов Г.А. Селекция и семеноводство березы карельской // Вестн. МГУЛ. Лесной вестник. 1998. № 1. С. 124–127.
Любавская А.Я. Селекция и разведение карельской березы. М.: Лесная промышленность, 1966. 124 с.
Матвеева Т.В., Машкина О.С., Исаков Ю.Н., Лутова Л.А. Молекулярная паспортизация клонов карельской березы при помощи ПЦР с полуслучайными праймерами // Экол. генетика. 2008. Т. 6. № 3. С. 18–23.
Матвеева Т.В., Павлова О.А., Богомаз Д.И. и др. Молекулярные маркеры для видоидентификации и филогенетики растений // Экол. генетика. 2011. Т. 9. № 1. С. 32–43.
Можаровская Л.В., Пантелеев С.В., Кирьянов П.С. и др. Структурно-функциональный анализ хлоропластного генома карельской березы // Научные стремления. 2018. Вып. 23. С. 9–12. https://doi.org/10.31882/2311-4711.2018.23.1
Николаева Н.Н., Воробъев В.В. Итоги инвентаризации генетического резервата карельской березы в НП “Себежский” // Научные исследования в заповедниках и национальных парках России / Тез. Всерос. науч.-практ. конф. Петрозаводск: КарНЦ РАН, 2016. С. 164.
Одум Ю. Экология. М.: Мир, 1986. Т. 2. 376 с.
Падутов В.Е., Хотылева Л.В., Баранов О. Ю., Ивановская С.И. Генетические эффекты трансформации лесных экосистем // Экол. генетика. 2008. Т. 6. № 1. С. 3–11.
Побирушко В.Ф. Распространение и изменчивость березы карельской в Беларуси // Ботаника. Минск: Навука i тэхнiка, 1992а. Вып. 31. С. 31–39.
Побирушко В.Ф. Эколого-биологические особенности и внутривидовая изменчивость некоторых видов рода Betula L. на границах ареалов (в условиях Беларуси): Автореф. дис. … канд. биол. наук. Минск: Ин-т экспер. ботаники АН Беларуси, 1992б. 25 с.
Побирушко В.Ф., Мартинссон У., Эмануэльссон Е. Распространение и изменчивость карельской березы в юго-восточной Швеции // Биологические основы изучения, освоения и охраны животного и растительного мира, почвенного покрова Восточной Фенноскандии / Тез. Междунар. конф. Петрозаводск: КарНЦ РАН, 1999. С. 43–44.
Погиба С.П., Рабцун А.С., Коровин В.В. Селекционно-генетический анализ лесных популяций // Вестн. МГУЛ. Лесной вестник. 2013. Т. 94. № 2. С. 4–7.
Попов С.Ю. Ценотическое распределение и экологические предпочтения Betula pendula и Betula pubescens в Центральной России // Журн. общ. биол. 2017. Т. 78. № 2. С. 61–73.
Пугачевский А.В. Карельская береза в Беларуси. Минск, 2008. 8 с.
Сакс К.А., Бандер В.Л. Опыт по выращиванию карельской березы в Латвийской ССР // Лесная генетика, селекция и семеноводство. Петрозаводск: Карелия, 1970. С. 294–300.
Сакс К.А., Бандер В.Л. Выращивание карельской березы в Латвийской ССР // Научные труды Украинской сельскохоз. академии. 1971 (1972). Вып. 65. С. 128–129.
Сакс К.А., Бандер В.Л. Новое в разведении березы карельской // Лесн. хоз-во. 1973. № 1. С. 40–41.
Санитарные правила в лесах РФ. М.: ВНИИЦлесресурс, 1998. 25 с.
Санников С.Н., Санникова Н.С., Петрова И.В. Очерки по теории лесной популяционной биологии. Екатеринбург: УрО РАН, 2012. 276 с.
Санников С.Н., Санникова Н.С., Петрова И.В., Шавнин С.А. Развитие лесной популяционной биологии и биогеоценологии на Урале // Сиб. лесн. журн. 2014. № 3. С. 9–24.
Семериков Л.Ф., Казанцев В.С. Популяционная структура дуба черешчатого (Quercus robur L.) в Поволжье и Предуралье // Экология. 1979. № 2. С. 12–21.
Сидор А.И., Ковалевич А.И., Луферова Н.С. и др. Карелка: Что имеем … // Лесное и охотничье хозяйство. Минск: Мин-во лесного хоз-ва Республики Беларусь, 2016. № 11. С. 18–23.
Соколов Н.О. Карельская береза. Петрозаводск: гос. изд-во Карело-Финской ССР, 1950. 116 с.
Соколов Н.О. Карельская береза. Л.: Изд. науч.-иссл. сектора ЛТА, 1959. 36 с.
Соколов Н.О. Отбор и выращивание березы карельской в Ленинградской области с использованием самосева // Лесная генетика, селекция и семеноводство. Петрозаводск: Карелия, 1970. С. 277–281.
Соколов Н.О. Отбор карельской березы в лесах и культурах Северо-Запада // Закономерности внутривидовой изменчивости лиственных древесных пород. Свердловск: УНЦ РАН, 1975. С. 111–114.
Ставрова Н.И. Структура популяций основных лесообразующих видов на Европейском севере России: Автореф. дис. … докт. биол. наук: СПб.: БИН РАН, 2012. 39 с.
Сулей М. Пороги для выживания: поддержание приспособленности и эволюционного потенциала // Биология охраны природы. М.: Мир, 1983. С. 177–196.
Тарасевич А.В. Популяционные разновидности рода Betula L. и их оценка для использования в Полесье Украины // Структурные и функциональные отклонения от нормального роста и развития растений под воздействием факторов среды / Мат. Междунар. конф. Петрозаводск: КарНЦ РАН, 2011. С. 337–346, 379–391.
Шапкин О.М., Погиба С.П., Казанцева Е.В. Популяционно-генетический анализ карельской березы и вегетативное размножение ее ценных форм // Лесохозяйственная информация Федеральной службы лесного хоз-ва. М.: ВНИИЦлесресурс, 1996. Вып. 9. С. 4–15.
Щетинкин С.В. Гистогенез узорчатой древесины березы (Betula pendula Roth var. carelica Mercl. и Betula pendula Roth): Автореф. дис. … канд. с.-х. наук. Воронеж: ЦНИИЛГиС, 1988. 24 с.
Щетинкин С.В., Щетинкина Н.А. К формированию узорчатой древесины карельской березы // Актуальные проблемы лесного комплекса. Вып. 51. Брянск: БГИТУ, 2018. С. 180–186.
Фардеева М.Б., Исламова Г.Р., Чижикова Н.А. Анализ пространственно-возрастной структуры растений на основе информационно-статистических подходов // Уч. записки Казанского гос. ун-та. 2008. Т. 150. Кн. 4. С. 226–240.
Федулова Т.П., Исаков Ю.Н., Корчагин О.М. и др. Молекулярно-генетическая дифференциация генотипов березы на основе полиморфизма SSR-маркеров // Лесотехн. журн. 2017. Т. 7. № 4. P. 6–16. https://doi.org/10.12737/article_5a3cef5a94cab2.12952001
Emanuelsson J. The natural distribution and variation of curly birch (Betula pendula Roth var. carelica (Merkl.) Sok.) (in Sweden: Examensarbete i ämnet skogsskötsel. Institutionen för skogsskötsel sveriges lantbruksuniversitet). Umeå, 1999. 54 p.
Hagqvist R., Mikkola A. Visakoivun kasvatus ja käyttö. Metsäkustannus Oy, 2008. 168 s.
Hamrick J.L., Godt M.J.W., Sherman-Broyles S.L. Factors influencing levels of genetic diversity in woody plant species // New Forests. 1992. V. 6. P. 95–124. https://doi.org/10.1007/BF00120641
Heikinheimo O. Kokemuksia visakoivun kasvatuksesta // Commun. Inst. Forest. Fenn. 1951. V. 39. № 5. P. 1–26.
Hejtmánek J. Betula pendula var. carelica Sokolov v Československu // Preslia. 1957. V. 29. P. 264–268.
Hodnebrog T. Valbjørk-produksjon på utvalgte kloner // Fagnytt, Plantedyrking. 1996a. № 3. 6 p.
Hodnebrog T. Utvalg av kloner valbiørk (Betula pendula f. carelica) // Norsk Landbruks forsking. 1996b. V. 10. № 2. P. 101–106.
Hynynen J., Niemistö P., Viherä-Aarnio A. et al. Silviculture of birch (Betula pendula Roth and Betula pubescens Ehrh.) in northern Europe // Forestry. 2010. V. 83. № 1. P. 103–119.
IUCN Red List Categories and Criteria. Version 3.1. IUCN, 2012. 32 p.
Jakuszewski T. Nowe stanowisko brzozy czeczotowatej Betula verrucosa Ehrh. var. carelica K. Merclin w Beskidzie Sadeckim // Rocz. Sek. Dendrol. PTB. 1970. V. 24. P. 31–33.
Jakuszewski T. Badania, zmienności i dziedziczności populacji brzozy karelskiej z Gorców // Arboretum Kórnickie. 1973. № 18. P. 35–81.
Johnsson H. Avkommor av masurbjork // Svenska Skogsvf. Tidskr. 1951. B. 49. № 1. S. 34–45.
Kosonen M., Leikola M., Hagqvist R. et al. Visakoivu. Curly Birch. Metsälehti Kustannus, 2004. 208 p.
Kurganskiy A., Skjøth C.A., Baklanov A. et al. Incorporation of pollen data in source maps is vital for pollen dispersion models //Atmos. Chem. Phys. 2020. V. 20. Is. 4. P. 2099–2121. https://doi.org/10.5194/acp-20-2099-2020
Lindquist B. On the variation in Scandinavian Betula verrucosa Ehrh. with some notes on the Betula series Verrucosae Sukacz // Bot. Tidskr. 1947. V. 41. № 1. P. 45–80.
Lindquist B. Forstgenetik in der Schwedischen Waldbaupraxis. Radebeul und Berlin: Neumann, 1954. 156 s.
Martinsson O. Odling av masurbjörk – en outvecklad nisch för svenskt skogsbruk // Fakta Skog. 1995. № 11. 4 s.
Martinsson O., Vetchinnikova L. Management, reproduction and protection of Karelian birch in Fennoscandia // Biological basis of the study, management and protection of flora, fauna and the cover in Eastern Fennoscandia // Intern. conf. Petrozavodsk: KarRC RAS, 1999. P. 64–65.
Mikkelä H. Guide to the montell trail in the Punkaharju experimental area. Helsinki: The Finnish Forest Research Institute, 1992. 27 p.
Naujoks G., Schneck V., Dietrich E. 30 Jahre in-vitro-vermehrung der Braunmaser-birke // AFZ-DerWald. 2017. B. 5. S. 32–35.
Pagan J., Paganová V. Breza biela svalcovita (Betula alba L. var. carelika Merk.) // Technicka univerzita vo Zvolen. 1994. № 10. 75 s.
Paganová V. Analysis of inheritance and growth of curly birch progenies from controlled hybridization and possibilities of their utilization for timber production in agricultural landscape // Czech. J. Genet. Plant Breed. 2004. V. 40. № 2. S. 51–62.
Pannell J.R., Barrett S.C.H. Effects of population size and metapopulation dynamics on a mating-system polymorphism // Theor. Popul. Biol. 2001. V. 59. Is. 2. P. 145–155. https://doi.org/10.1006/tpbi.2000.1496
Pork K., Sander R. Maarajkase levikust Lääne-Eestis // Eesti loodus. 1973. № 6. P. 332–335.
Raulo J., Sirén G. Neljän visakoivikon päätehakkuun tuotos ja tuotto // Silva Fennica. 1978. V. 12. № 4. P. 245–252.
Ruden T. Om valbjørk og endel andre unormale veddannelser hos bjørk // Medd. Foren. Det. Norske Skogforsøksv. 1954. B. 43. S. 451–505.
Ryynänen L. Cloning of Betula pendula and Betula pubescens by means of tissue culture // Bull. Finn. Forest Res. Inst. 1988. № 304. P. 24–30.
Salojärvi J., Smolander O.-P., Nieminen K. et al. Genome sequencing and population genomic analyses provide insights into the adaptive landscape of silver birch // Nature Genetics. 2017. V. 49. № 6. P. 904–912. https://doi.org/10.1038/ng.3862
Scholz E. Das Verbreitungsgebiet der Braunmaserbirke // Arch. Forstwesen. 1963. B. 12. № 12. S. 1243–1253.
Sibul I., Habicht K.-L., Ploomi A. Curly birch stands and cultivation results in Estonia // Structural and functional deviations from normal growth and development of plants under the influence of environmental factors / Mat. of intern. conf. Petrozavodsk: KarRC RAS, 2011. P. 310–313.
Sofiev M. On impact of transport conditions on variability of the seasonal pollen index // Aerobiologia. 2017. V. 33. Is. 3. P. 167–179. https://doi.org/10.1007/s10453-016-9459-x
Sofiev M., Siljamo P., Ranta H., Rantio-Lehtimäki A. Towards numerical forecasting of long-range air transport of birch pollen: theoretical considerations and a feasibility study // Int. J. Biometeorol. 2006. V. 50. Is. 6. P. 392–402.
Vailionis L. Lietuvos berzu reta (Die wisakrankheit in den wäldern Litauens) // Scripta Horti Bot. Univ. Bytauti Magni.1935. V. 3. P. 5–36.
Václav E. Rozšíření, vlastnosti a pěstování svalcovité břízy v ČSSR // Přírodovědecký časopis slezský. 1961. Sv. 22. №2. S. 151–171.
Vάclav E. Rozšíření, stanovištní podmínky a růst svalcovité břízy (karelské) v Evropě // Sborník lesnické fakultyVŠZ v Praze. 1963. № 6. S. 217–237.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии