

Успехи современной биологии, 2020, T. 140, № 6, стр. 574-583
Питание и онтогенез. Часть 2. Степенная энерговременная взаимосвязь между удельной интенсивностью метаболизма и продолжительностью жизни и онтогенетический (биологический) возраст видов плацентарных млекопитающих
В. М. Жминченко *
Федеральный исследовательский центр питания и биотехнологии
Москва, Россия
* E-mail: zhminchenko@ion.ru
Поступила в редакцию 31.05.2020
После доработки 31.05.2020
Принята к публикации 14.06.2020
Аннотация
Целью исследования было формализовать энерговременные закономерности постнатального онтогенетического развития видов плацентарных млекопитающих. Подтверждена обратная функциональная связь между максимальной продолжительностью жизни исследованных видов плацентарных млекопитающих и удельной суточной интенсивностью метаболизма (энергии), описываемая уравнением степенной регрессии. Обнаружена также обратная степенная закономерность между удельным суточным потреблением белка взрослыми млекопитающими и продолжительностью жизни. Функциональная энерговременная закономерность постнатального онтогенетического развития млекопитающих порождает единый энергетический эталон онтогенеза, который одновременно предстает пределом длительности жизни, что делает онтогенез видов энергетически равноценным с расходом общей удельной энергии. Распределение видов внутри класса в зависимости от диапазона удельных интенсивностей метаболизма белка и энергии и соответствующей продолжительности жизни описывается уравнением показательной функции с основанием 2 : Y = 2n, где каждый вид принадлежит только к одной из 9 групп, нумерация групп (n) начинается с нулевой. Значения скорости постнатального развития и продолжительности жизни видов в каждой группе можно определить из следующих уравнений: Т – максимальная продолжительность жизни видов = 2(7– n), в годах; Э – удельная интенсивность метаболизма = 100 × 2n, кДж/кг/сутки; Б – удельная интенсивность обмена белка = 2n, г/кг/сутки. Аллометрия обобщения и подобия взаимосвязи интенсивности метаболизма энергии (скорости онтогенетического развития) с максимальной продолжительностью жизни изучаемых видов млекопитающих позволила обосновать и оценить их онтогенетический (биологический) возраст.
ВВЕДЕНИЕ
Изучение онтогенетического развития (старения) человека и животных – прерогатива многих научных дисциплин. Каждая дисциплина избрала свою нишу в исследовании онтогенеза: так биология рассматривает закономерности и механизмы продолжительности жизни организмов; геронтология – процессы старения и механизмы ограничения продолжительности жизни; нутрициология – влияние пищи, ее количества или отдельных пищевых веществ на продолжительность жизни, на ее качество и проявления риска возрастных заболеваний; биохимия и физиология изучают молекулярно-генетические механизмы возможной запрограммированности старения и долголетия или их отсутствия.
Методологический подход к исследованию процессов онтогенеза, определяющих длительность жизни от действия внешних и внутренних факторов, должен основываться на изучении закономерностей перманентно происходящих процессов жизнедеятельности в течение онтогенетического развития, приводящих в конечном итоге организм к завершению онтогенеза.
В предыдущей статье (Жминченко, 2020) были проанализированы потенциальные энергетические возможности постнатального онтогенеза различных видов и классов. Было установлено, что общие удельные энергозатраты на постнатальный онтогенез плацентарных млекопитающих, кроме рукокрылых и видов, входящих в оцепенение или спячку, практически равны и составляют постоянную величину в 4926.7 ± 162.3 МДж/кг массы тела/максимальную продолжительность жизни (МПЖ). Равноценная удельная энергоемкость постнатального онтогенеза млекопитающих указывает на существование общих закономерностей их онтогенетического развития и на наличие клеточных структур и механизмов, которые определяют (лимитируют) энергоемкость онтогенеза и, соответственно, длительность жизни видов.
В настоящей работе была поставлена задача формализовать энерговременные закономерности онтогенетического развития видов плацентарных млекопитающих в зависимости от удельной интенсивности метаболизма (скорости онтогенетического развития) и продолжительности их жизни.
МАТЕРИАЛЫ ИССЛЕДОВАНИЯ
В классе плацентарных млекопитающих исследовали взаимосвязь между интенсивностью метаболизма и длительностью жизни видов. Объектом исследования были интегральные параметры постнатального онтогенеза: среднесуточные энергозатраты – кДж/кг массы тела/сутки и МПЖ – годы изучаемых видов.
Материалом исследования послужили данные литературы о среднесуточных удельных расходах энергии взрослых особей или нормы суточных потребностей в энергии и МПЖ 88 видов плацентарных млекопитающих из 8 отрядов: насекомоядные (Insectivora) – 2; грызуны (Rodentia) – 12, хищные (Carnivora) – 43, парнокопытные (Artiodactyla) – 8, непарнокопытные (Perissodactyla) – 2, зайцеобразные (Lagomorpha) – 1, китообразные (Cetacea) – 3 и приматы (Primates) – 17, значения которых представлены в работе (Жминченко, 2020). Данное разнообразие отрядов и видов обеспечивает филогенетическую независимость проведения сравнительных межвидовых исследований (Felsenstein, 1985) и дает правомочность для установления энерговременных закономерностей постнатального онтогенеза видов внутри класса.
Предполагая наличие обратной степенной взаимосвязи между интенсивностью метаболизма и продолжительностью жизни видов, и последующего математического описания процесса полученных зависимостей между интегральными атрибутами постнатального онтогенеза изучаемых видов, использовали алгоритмы и компьютерные программы. Вычисляли параметры линии регрессии функции методом наименьших квадратов, линейного, нелинейного и корреляционного анализов, при помощи которых вычисляли коэффициенты: масштабный множитель – k и показатель степени – b уравнений степенной или линейной регрессии между логарифмами изучаемых признаков и индексы: корреляции, детерминации, ошибки аппроксимации, критерии: F – Фишера и DW – Дарбина–Уотсона. Использован метод последовательного укомплектовывания экспериментальных групп (секвенциальный анализ) (Сепетлиев, 1968).
Обратная степенная закономерность взаимосвязи между интенсивностью метаболизма энергии и максимальной продолжительностью жизни 88 видов плацентарных млекопитающих.
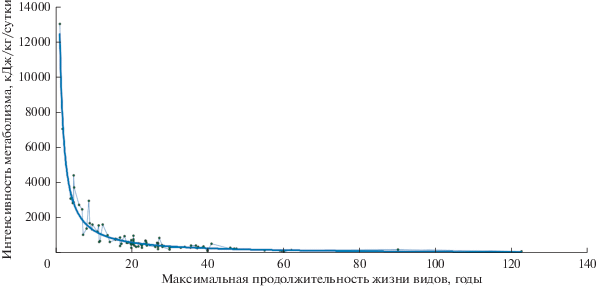
Аллометрия обобщения и подобия взаимосвязи между интенсивностью метаболизма энергии и максимальной продолжительностью жизни изучаемых видов млекопитающих позволила установить и оценить их онтогенетический (биологический) возраст.
Таблица.
Ранжирование видов плацентарных млекопитающих внутри класса в зависимости от скорости онтогенетического развития продолжительности жизни
| Нумерация групп Показатель | 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|---|---|---|---|---|---|---|---|---|
| МПЖ = 2(7 –n), годы | 128 | 64 | 32 | 16 | 8 | 4 | 2 | 1 | 0.5 |
| Уд. интенсивность метаболизма или скорость онтогенетического развития = 100 × 2n, кДж/кг/сутки | 100.0 | 200.0 | 400.0 | 800.0 | 1600.0 | 3200.0 | 6400.0 | 12800.0 | 25600.0 |
| Уд. интенсивность белка = 2n, г/кг/сутки | 1.0 | 2.0 | 4.0 | 8.0 | 16.0 | 32.0 | – | – | – |
| Коэффициент соизмеримости* | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | 256 |
| Виды млекопитающих | Человек, касатка | Лошадь, осел, верблюд, горилла, шимпанзе, орангутан, тюлень, дельфин, нерпа, верблюд, бонобо | Корова, свинья, собака, кошка, павиан, лисица, макак-резус, мартышка, ревун, гиена, енот, лев, овца и др. | Заяц, лисица, песец, игрунка, сурикат, волк, гепард, мунго, сервал, каракал, шакал, спрингбок, рысь, пума и др. | Прыгун, песчанка, ласка, белка, хорек, норка | Крыса, мышь, суслик, полевка | Землеройка | Бурозубка | Этрусская землеройка |
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Для обнаружения энерговременных закономерностей постнатального онтогенеза плацентарных млекопитающих величины ежедневных удельных расходов энергии и МПЖ 88 видов были внесены в матрицы компьютерных программ графического, регрессионного и корреляционного анализов. После обработки данных на равномасштабном графике была видна регрессионная кривая в виде гиперболы между изучаемыми показателями онтогенетического развития видов: интенсивностью метаболизма энергии (Y) и МПЖ видов (Х), которая может быть описана уравнением степенной функции типа:
где b – показатель степени, аллометрический показатель; k – масштабный множитель, коэффициент пропорциональности.После вычисления коэффициентов k = 15118.6 и b = –1.0517 уравнение степенной регрессии между удельными суточными затратами энергии и МПЖ 88 видов плацентарных млекопитающих приобрело вид:
где Э – энергия (кДж/кг/сут) и Т – МПЖ (годы).Для оценки значимости параметров регрессии и корреляции найдены: среднее значение энергии Y = 1051.2 кДж/кг/сут, индекс корреляции R = ~0.98 и индекс детерминации R2 = ~0.96 при средней ошибке аппроксимации A = ~25%. F-критерии Фишера: F-критический (табличный) F = (0.05, 1.86) ~3.95 и F-фактический = ~1973.8, DW-критерии Дарбина–Уотсона: DW-фактический d = ~1.56.
Уравнение (1) путем логарифмирования признаков превратилось в уравнение прямой линии:
После вычисления коэффициентов линейной регрессии уравнение приняло вид:
Определены коэффициенты линейной парной корреляции (rxy) и детерминации (R2), которые равны соответственно: rxy = –0.94 ± 0.037 и R2 = 0.88. Для оценки значимости параметров регрессии и корреляции вычислены: Х-средний = 1.306334, средняя ошибка аппроксимации А = ~3.83%, F-критерии Фишера: F-критический (табличный) = ~3.95 и F-фактический = ~620.4, случайные ошибки (m) параметров k и b и коэффициента корреляции rxy: mk = ~0.042; mb = ~0.056; mrxy = = ~0.037; t-критерий Стьюдента (табличный) 1.987, при df (количество степеней свободы) = n – 2 = = 88 – 2 = 86; α (уровень значимости) = 0.05; tk = = –24.9, tb = rxy ~73.4, trxy = –24.9 и DW-критерий Дарбина–Уотсона: DW-фактический d = ~1.65.
Секвенциальный анализ подтвердил, что необходимое число наблюдений уже давно превысило достаточное количество видов для выявления обратной степенной энерговременной зависимости развития постнатального онтогенеза плацентарных млекопитающих (Жминченко, 1988, 1991, 2004).
Анализируя уровень адекватности обнаруженных закономерностей в аллометрических уравнениях степенного подобия (Каркищенко, 2007) и оценивая при этом величину индекса корреляции R = ~0.98 в уравнении (2) и коэффициент линейной парной корреляции rxy = –0.94 в уравнении (4), у которых величины показателей соответствуют значениям сверхсильной и сильнейшей связи между изучаемыми факторами онтогенетического развития млекопитающих соответственно (Мазуркин, 2013), следует обратить внимание на величину показателя степени (наклона) b = –1.05 в этих аллометрических уравнениях.
На равномасштабном графике (рисунок) точки пересечения между величинами удельных суточных расходов энергии с МПЖ 88 исследованных видов взрослых млекопитающих расположены вдоль кривой линии, характеризующей обратную зависимость между изучаемыми признаками онтогенеза плацентарных млекопитающих. В данном случае на графике линия регрессии обозначает биологическую зависимость, показывающую, что продолжительность жизни млекопитающего уменьшается как величина интенсивности метаболизма энергии в степени –1.05.
Подтверждением реального существования степенной энерговременной закономерности онтогенетического развития видов млекопитающих служит зеркальная степенная зависимость (Atanasov, 2007), установленная на 95 млекопитающих, включая 3 – однопроходных, 16 – сумчатых и 76 видов плацентарных из 13 отрядов млекопитающих на основании изучения взаимосвязи между общей метаболической энергией (кДж/сут) и средней продолжительностью жизни Т (дни) и М – массой тела животных (кг), которая описывается уравнением степенной функции:
где Эоб – это общая энергия основного обмена; коэффициент пропорциональности А = 7158 × × 105 (кДж/кг).При сравнении уровней наклона (коэффициентов степени) уравнений (2), (4) и (5) видно, что они по величине практически равны, но только с противоположным знаком. Эти три уравнения содержат параметры одной зависимости продолжительности жизни видов от уровня интенсивности метаболизма: в первых двух случаях – от общего суточного удельного расхода энергии, а в третьем – только от суточного расхода энергии животными на основной обмен.
Таким образом, формализована обратная степенная взаимозависимость между удельной суточной интенсивностью метаболизма (энергии) и МПЖ исследованных видов млекопитающих, которая требует теоретического и экспериментального подтверждения для ее существования на основании характера подобия изменений других признаков онтогенетического развития видов от интенсивности метаболизма.
Связь между питанием и временным онтогенетическим развитием видов плацентарных млекопитающих должна быть также проанализирована по одному из ключевых субстратов питания – белку, который является, с одной стороны, универсальным пищевым веществом, идущим на обеспечение пластических, регуляторных, энергетических и других потребностей организма, а с другой – уникальным нутриентом, содержащим эссенциальные аминокислоты. Данная специфичность и несомненная ведущая роль белков в процессах онтогенеза служат основанием для выбора его в качестве главного атрибута измерения и оценки постнатального развития видов.
Раннее было установлено, что затраты белка на постнатальный онтогенез видов млекопитающих составляют 50.6 ± 0.21 кг белка/кг массы тела/МПЖ (Жминченко, 2004). Было проанализировано соотношение между уровнями содержания белка и энергии в суточных рационах и корме млекопитающих, что позволило обнаружить прямо пропорциональную зависимость между потребляемым количеством белка различными видами и необходимым уровнем энергии для оптимальной его утилизации (при R = 0.98 и при n = 27), которая описывается следующим уравнением:
где Эсут – энергия суточного рациона (кДж/кг массы тела); Б – белок, (г), то есть на каждый 1 грамм потребляемого белка приходится примерно 109 кДж общей энергии, включая энергию белка. Усредненный рацион питания млекопитающих содержит 15.5 ± 1.3% белка по калорийности (Жминченко, 2000).Полный корреляционный анализ зависимости между продолжительностью жизни видов млекопитающих и удельными интенсивностями обмена белка выявил степенную закономерность при показателе прямолинейной и недостоверной связи равной 0.07, показателем криволинейной связи – 0.7, при критерии криволинейности, равном 9.1, и его достоверности при р < 0.001 (Жминченко, 2004). Степенная закономерность описывается уравнением регрессии между продолжительностью жизни (T, годы) и удельным суточным потреблением белка (Б, г/кг/сут) взрослыми млекопитающими:
Обратная степенная зависимость подтверждает наличие взаимосвязи между увеличением интенсивностей обмена белка и энергии и одновременным уменьшением продолжительности жизни видов. Энерговременная закономерность постнатального онтогенетического развития млекопитающих порождает единый энергетический эталон для постнатального онтогенеза, который одновременно предстает пределом длительности жизни, что делает онтогенез видов энергетически равноценным по расходу энергии в 4926.7 ± 162.3 МДж/кг массы тела/МПЖ и затратами белка в 50.6 ± 0.21 кг/МПЖ.
Как видно на графике (рисунок) параметры изучаемых показателей онтогенеза видов находятся в пределах минимальных и максимальных значений по МПЖ от 1 года до 122 лет и в интервале максимальных и минимальных удельных суточных уровней энергозатрат от 134 00 до 100 кДж/кг/сут соответственно. Диапазоном границ распределения по энергетическим характеристикам могут служить минимальные и максимальные физиологические уровни интенсивности метаболизма видов, которые могут обеспечить жизнедеятельность гомойотермного организма. Для поддержания температуры тела требуется примерно 83.6 кДж/кг/сут (Galloway, 1974), что является нижним пределом интенсивности метаболизма и существования теплокровного животного. При экстраполяции линии регрессии уравнения (2) вычислили теоретически максимальную величину интенсивности метаболизма млекопитающего, которая составляет примерно 31 339.5 кДж/кг/сут при МПЖ животного 0.5 года. Такой возможной удельной интенсивностью метаболизма в 28 939.2 кДж/кг/сут обладает этрусская землеройка (Etruscan shrew) (Jurgens et al., 1996). Это животное – с минимально возможной массой тела в 1–2 грамма, c минимальной длительностью жизни, при которой обеспечивается жизнедеятельность всего тела эндотермного взрослого индивидуума пищевыми веществами и кислородом, с функционально возможной активностью сердечно-сосудистой, дыхательной систем и эффективностью окислительного фосфолирирования (Dobson, Headrick, 1995; West et al., 2002; West, Brown, 2005). В этих реальных диапазонах интенсивностей метаболизма и продолжительностей жизни плацентарных млекопитающих можно ранжировать изучаемые виды в 9 группах, отличающихся друг от друга скоростью онтогенетического развития, где каждая группа будет иметь собственные параметры, характеризующие данную группу в зависимости от удельной интенсивности метаболизма белка и энергии и продолжительности жизни видов.
Распределение видов в зависимости от интенсивности метаболизма и продолжительности жизни может быть описано уравнением показательной функции с основанием 2:
Каждый вид может принадлежать только одной из 9 групп, нумерация групп (n) начинается с нулевой. Значения удельной интенсивности метаболизма или энергетическая скорость онтогенетического развития и МПЖ каждой группы можно определить из следующих уравнений: где Т – МПЖ (годы); Эуд – удельная интенсивность метаболизма (кДж/кг/сут); Буд – удельная интенсивность обмена белка (г/кг/сутки).Аллометрическое энергетическое подобие онтогенеза видов внутри класса, возникающее при измерении общей удельной энергии в интервале времени онтогенеза от рождения до конца постнатального развития, и учитывая, что все удельные энергетические константы развития у видов аллометрического нелинейного подобия одинаковы, то любое аддитивно безразмерное отношение, найденное в “модели” онтогенеза, можно переносить на прототип (Каркищенко, 2007).
Перед биологами давно стояла задача найти единицу биологического времени, которая была бы сопоставима между индивидуумами одного вида животного при равных условиях, а также при сравнении разных видов. Отдельными исследователями было предложено несколько частных решений этой задачи. При этом во всех случаях время было определено не в единицах астрономического времени, а в долях (или числе) того или иного периода развития, продолжительность которого была принята за единицу времени. Это метод относительной или безразмерной характеристики продолжительности (времени) развития, то есть использование относительной единицы времени (relative hours) (Детлаф Т., Детлаф А., 1982). Здесь вполне оправдано использование скорости суточного метаболизма в качестве показателя физиологического времени, и это понятие можно применять ко всем типам процессов в организме, имеющих скорость (Шмидт-Ниельсон, 1987; Алимов, Казанцева, 2007; Lindstedt, Calder, 1981). “Удельная скорость метаболизма задает переменную единицу собственного, физиологического, времени, и физиологическая длительность любого периода развития или всей жизни имеет размерность физиологического времени и численно равна суммарному удельному метаболизму за этот период развития” (Алимов, Казанцева, 2007, с. 352).
Очевидно, что соизмеримость биологического возраста правомочна лишь при общности онтогенеза видов и в случае, если последние обладают единым механизмом, определяющим длительность жизни. Виды млекопитающих показывают общее сходство в фенотипе старения; это предполагает, что фундаментальная причина старения человека является общей для большинства, если не для всех млекопитающих (de Magalhães, Toussaint, 2002). Можно предположить, что отличия между видами проявляются лишь в скорости клеточного обновления, которая в свою очередь зависит от интенсивности метаболизма. Возможно, возраст органа соответствует определенному делению стволовой клетки и затраченному на данный момент удельному количеству энергии (Жминченко, 1992).
В связи с удельной энергетической тождественностью постнатального развития плацентарных млекопитающих можно заключить, что как человек, так и другие виды обладают одинаковым онтогенетическим (биологическим) возрастом (Жминченко, 1991), который условно можно принять за 1, при одинаковых затратах энергии у видов на онтогенез около 5000 МДж/кг/МПЖ. Исходя из одинаковых удельных затрат энергии на постнатальный онтогенез исследованных видов плацентарных млекопитающих и принимая эту величину за константу подобия, полученную при измерении их постнатального онтогенеза в единицах энергии, в кДж, можно прийти к пониманию существования онтогенетического (биологического) возраста этих видов.
В таблице изучаемые виды млекопитающих были сгруппированы в 9 группах в зависимости от реально возможных среднесуточных скоростей онтогенетического развития, то есть уровней удельного метаболизма энергии, белка и МПЖ, где каждая последующая группа видов отличается от предыдущей в 2 раза по величинам соответствующих признаков постнатального развития, а максимальные значения признака первой близки к среднему значению показателя последующей группы. Рассматривая таблицу, становится понятно, что виды ранжированы в определенном порядке в зависимости от возрастания удельных интенсивностей энергии и белка, следуя показательной закономерности увеличения интенсивности метаболизма и одновременного уменьшения МПЖ.
На основании установленной энерговременной закономерности онтогенеза млекопитающих можно вычислить безразмерные коэффициенты соизмеримости онтогенетического (биологического) возраста в зависимости от скоростей онтогенетического развития видов (Жминченко, 1992). Их реальная величина равна значениям соотношений видовых удельных суточных затрат энергии или суточного метаболизма белка сравниваемых видов. Подобным коэффициентом соизмеримости служит величина отношения МПЖ сравниваемых видов, так как длительность жизни находится в обратно пропорциональной зависимости от скорости онтогенеза. Таблица дает возможность соизмерить скорости онтогенетического развития видов на основании того, что энергетическая тождественность онтогенеза млекопитающих приравнивает их онтогенетический (биологический) возраст к одной единице. При сравнении видов из разных групп, значения коэффициентов соизмеримости скоростей онтогенетического развития можно получить при делении друг на друга их видовых МПЖ или уровней интенсивностей метаболизма энергии или белка, соответственно.
ЗАКЛЮЧЕНИЕ
Можно предположить, что феномен равноценной удельной энергоемкости постнатального онтогенеза видов плацентарных млекопитающих есть прямой результат единых механизмов жизнедеятельности и онтогенетического развития видов. В таком случае, расход метаболизируемой энергии (пищи) в единицу времени будет определять скорость онтогенетического развития организма.
Подтверждением данному заключению служат следующие факты изменения скорости онтогенетического развития видов в зависимости от расхода ими метаболизируемой энергии. Повышенное поступление пищи (энергии) по сравнению с обычным потреблением количества пищи в раннем постнатальном периоде у людей (Kirkwood et al., 1987; Cameron, 1996; Stipp, 2011), самок кроликов (Fodor et al., 2001), телок (Barcellos et al., 2014) и у птиц: кур (Katanbaf et al., 1989), индюшек (Hester, Stevens, 1990) приводит к преждевременному половому созреванию. Ограничение потребления пищи в этом периоде онтогенеза задерживает половое развитие: у девочек (Kirkwood et al., 1987; Cameron, 1996; Stipp, 2011), самок кроликов (Fodor et al., 2001), свинок (Kirkwood et al., 1987; Cameron, 1996), самок крыс (Holehan, Merry, 1985). Для достижения времени половой зрелости самкам крыс требуется определенное количество кумулятивной энергии, приблизительно 2300 кДж/крысу (Odum et al., 2004). Линейная зависимость наблюдается между длительностью ограничения времени кормления экспериментальных животных и увеличением МПЖ видов грызунов (данные взяты из 36 исследований, рассматривающих эксперименты по ограничению потребления калорий в диапазоне 40–50% (Merry, 2002)). У некоторых штаммов мышей снижение потребления калорий до 60%, ad libitum животных, вызывает периодическое оцепенение и увеличивает продолжительность жизни до 50% (Koizumi et al., 1992). При этом одним из надежных эффектов ограничения калорий является снижение скорости пролиферации клеток в митотических тканях (Bruss et al., 2011). Предыдущие работы с использованием 3Н-тимидина показали снижение темпов клеточной пролиферации в двенадцатиперстной, тонкой и толстой кишках, мочевом пузыре, коже, тканях пищевода и молочной железы в ответ на 25%-ное ограничение калорий в диете швейцарских мышей Вебстер (Lok et al., 1988; Hsieh et al., 2005) и снижение пролиферации клеток в первичных культурах гепатоцитов с низкокалорийной диетой F344 крыс (Shaddock et al., 1996). Интенсивность внутриклеточного и клеточного обновления жизненно важных органов: сердца, печени, почек зависит прямо пропорционально от потребляемого количества пищи лабораторными животными (Жминченко, Соколов, 1990).
За последние 100 лет в экспериментальных исследованиях было обнаружено увеличение длительности жизни у представителей различных классов: дрожжей, нематод, насекомых, паукообразных, рыб, млекопитающих: мышей, крыс, хомяков, собак, нечеловекообразных обезьян (Masoro, 2002, 2006; Spindler, 2010; Arslan-Ergul et al., 2013; Colman et al., 2014; Mattison et al., 2017), достигнутое путем уменьшения потребления пищи.
На основании этих эмпирических фактов ускорения или замедления онтогенетического развития индивидуумов различных видов млекопитающих, происходящих в зависимости от потребляемого ими количества пищи и расходов энергии, можно прийти к следующему выводу: скорость постнатального онтогенеза плацентарных видов млекопитающих есть результат проявления единого процесса интенсивностей метаболизма и клеточного обновления органов и тканей (Жминченко и др., 2008). Расходуемая энергия (пища) и продолжительность жизни организма видов предстают мерилом для их онтогенетического развития.
Энерговременное измерение постнатального онтогенеза видов млекопитающих в зависимости от их МПЖ позволило обнаружить сверхсильную корреляционную связь, как при исследовании закономерности между средней продолжительностью жизни млекопитающих и суммарной энергией основного обмена массы тела (Atanasov, 2007), так и при изучении связи между МПЖ видов и общими удельными расходами энергии и белка за постнатальный онтогенез. Установление одинаковой энерговременной закономерности постнатального онтогенеза млекопитающих исследователями, которые использовали при этом различные признаки онтогенетического развития млекопитающих, показали несостоятельность возражений оппонентов (Speakman, 2005; Hulbert et al., 2007) против сравнительного изучения постнатального развития или старения видов внутри этого класса с помощью вышеназванных критериев постнатального онтогенеза.
Явление равноценности удельных энергозатрат за постнатальный онтогенез и функциональная взаимосвязь между продолжительностью жизни и интенсивностями обмена белка и энергии у видов данного класса дают основание прийти к следующему заключению:
1) продолжительность жизни исследованных видов млекопитающих регулируется одними и теми же механизмами, 2) пища для плацентарных млекопитающих имеет одинаковую удельную энергетическую ценность для их постнатального развития.
Следовательно, вывод об удельной энергетической равноценности онтогенеза плацентарных млекопитающих достоверен и данные показатели онтогенеза, в виде белковой и энергетической удельной постоянной, могут служить таксономическими признаками для плацентарных млекопитающих.
Список литературы
Алимов А.Ф., Казанцева Т.И. Определение единицы внутреннего (физиологического) времени организма // Изв. РАН. Серия биол. 2007. № 3. С. 347–353.
Детлаф Т.А., Детлаф А.А. Безразмерные критерии как метод количественной характеристики длительности развития животных. Математическая биология развития. М.: Наука, 1982. С. 25–39.
Жминченко В.М. Энерговременная организация постнатального онтогенеза млекопитающих и продолжительность жизни / Тез. докл. V Всесоюз. съезда геронтологов и гериаторов. Тбилиси, 22–25 ноября, 1988 г. Киев, 1988. С. 233–234.
Жминченко В.М. Продолжительность жизни, биологический возраст и энерговременная организация онтогенеза млекопитающих и птиц // Продолжительность жизни: механизмы, прогнозы, пути увеличения / Тез. докл. Всесоюз. конф. 15–17 октября 1991 г. Киев, 1991. С. 41–42.
Жминченко В.М. О соизмеримости биологического возраста млекопитающих // Ускоренное старение, связь с возрастной патологией / Тез. докл. науч. конф. 13–15 октября 1992 г. Киев, 1992. С. 64.
Жминченко В. М. Концепция оценки потребностей млекопитающих в белке, энергии и их оптимального соотношения в рационе / Мат. междунар. науч.-техн. конф. “Пищевой белок и экология”. М.: МГУПБ, 2000. С. 199.
Жминченко В.М. Питание, энергия онтогенеза и время жизни человека, млекопитающих и птиц // Национальная политика здорового питания Республики Казахстан / Мат. междунар. науч.-практ. конф., г. Алматы, 2004. С. 95–99.
Жминченко В.М. Питание и онтогенез. Часть 1. Энергетическая удельная равноценность постнатального онтогенеза плацентарных млекопитающих // Успехи соврем. биол. 2020. Т. 140. № 5. С. 507–520.
Жминченко В.М., Соколов А.И. Питание (количество пищи) и скорость онтогенетического развития млекопитающих // Питание: здоровье и болезнь / Тез. науч. конф. с международным участием. М., 1990 г. С. 77.
Жминченко В.М., Соколов А.И., Тарасова И.Б., Сафронова А.М. Питание и взаимосвязь между интенсивностями клеточного обновления и метаболизма внутренних органов крыс // Питание и здоровье / Мат. Х Всерос. конгресса диетологов и нутрициологов. Москва, 1–3 декабря 2008 г. С. 34.
Каркищенко Н.Н. Через критерии подобия и аллометрии к валидации и экстраполяции в биомедицине // Биомедицина. 2007. № 6. С. 5–28.
Мазуркин П.М. Идентификация статистических устойчивых закономерностей // Наука и мир. 2013. № 3 (3). С. 28–33.
Сепетлиев Д. Статистические методы в научных медицинских исследованиях / Ред. А.М. Мерков. М.: Медицина, 1968. 419 с.
Шмидт-Ниельсен К. Размеры животных: почему они так важны? Пер. с англ. М.: Мир, 1987. 259 с.
Arslan-Ergul A., Ozdemir A.T., Adams M.M. Aging, neurogenesis, and caloric restriction in different model organisms // Aging Dis. 2013. V. 4. № 4. P. 221–232.
Atanasov A.T. The linear allometric relationship between total metabolic energy per life span and body mass of mammals // BioSystems. 2007. V. 90. P. 224–233.
Barcellos J.O., Pereira G.R., Dias E.A. et al. Higher feeding diets effects on age and liveweight gain at puberty in crossbred Nelore × Hereford heifers // Trop. Anim. Health Prod. 2014. V. 46 (6). P. 953–960.
Bruss M.D., Thompson A.C., Aggarwal I. et al. The effects of physiological adaptations to calorie restriction on global cell proliferation rates // Am. J. Physiol. Endocrinol. Metab. 2011. V. 300. № 4. P. E735–E745.
Cameron J.L. Nutritional determinants of puberty // Nutr. Rev. 1996. V. 54. № 2. P. S17–S22.
Colman R.J., Beasley T.M., Kemnitz J.W. et al. Caloric restriction reduces age-related and all-cause mortality in rhesus monkeys // Nat. Commun. 2014. V. 5. P. 3557.
de Magalhães J.P., Toussaint O. The evolution of mammalian aging // Exp. Gerontol. 2002. V. 37. № 6. P. 769–775.
Dobson G.P., Headrick J.P. Bioenergetic scaling: metabolic design and body-size constraints in mammals // PNAS USA. 1995. V. 92. № 8. P. 7317–7321.
Felsenstein J. Phylogenies and the comparative method // Am. Nat. 1985.V. 125. P. 1–15.
Fodor K., Fekete S.G., Zoldag L. et al. Influence of feeding intensity on corporeal development, body composition and sexual maturity in female rabbits // Acta Vet. Hung. 2001. V. 49. № 4. P. 399–411.
Galloway N.O. Heat production and senescence // J. Am. Geriatr. Soc. 1974. V. 22. № 4. P. 149–150.
Hester P.Y., Stevens R.W. Feed restriction of turkey breeder hens – a review // Poult. Sci. 1990. V. 69. № 9. P. 1439–1446.
Holehan A.M., Merry B.J. The control of puberty in the dietary restricted female rat // Mech. Age. Dev. 1985. V. 32. № 2–3. P. 179–191.
Hsieh E.A., Chai C.M., Hellerstein M.K. Effects of caloric restriction on cell proliferation in several tissues in mice: role of intermittent feeding // Am. J. Physiol. Endocrinol. Metab. 2005. V. 288. № 5. P. E965–E972.
Hulbert A. J., Pamplona R., Buffenstein R., Buttemer W.A. Life and death: metabolic rate, membrane composition, and life span of animals // Physiol. Rev. 2007. V. 87. P. 1175–1213.
Jurgens K.D., Fons R., Peters T., Sender S. Heart and respiratory rates and their significance for convective oxygen transport rates in the smallest mammal, the Etruscan shrew Suncus etruscus // J. Exp. Biol. 1996. V. 199. Pt 12. P. 2579–2584.
Katanbaf M.N., Dunnington E.A., Siegel P.B. Restricted feeding in early and late-feathering chickens. 2. Reproductive responses // Poult. Sci. 1989. V. 68. № 3. P. 352–358.
Kirkwood R.N., Cumming D.C., Aherne F.X. Nutrition and puberty in the female // Proc. Nutr. Soc. 1987. V. 46. № 2. P. 177–192.
Koizumi A., Tsukada M., Wada Y. et al. Mitotic activity in mice is suppressed by energy-restriction induced torpor // J. Nutr. 1992. V. 122. P. 1446–1453.
Lindstedt S.L., Calder W.A. Body size, physiological tame, and longevity of homeothermic animals // Quart. Rev. Biol. 1981. V. 56. № 1. P. 1–16.
Lok E., Nera E.A., Iverson F. et al. Dietary restriction, cell proliferation and carcinogenesis: a preliminary study // Cancer Lett. 1988. V. 38. № 3. P. 249–255.
Masoro E.J. Research profiles in aging. Caloric restriction: a key to understanding and modulating aging. Charleston, South Carolina USA: Elsevier Science, 2002. V. 1. 192. p.
Masoro E.J. Dietary restriction-induced life extension: a broadly based biological phenomenon // Biogerontology. 2006. V. 7. № 3. P. 153–155.
Mattison J.A., Colman R.J., Beasley T.M. et al. Caloric restriction improves health and survival of rhesus monkeys // Nat. Commun. 2017. V. 8. P. 14063.
Merry B.J. Molecular mechanisms linking calorie restriction and longevity // Internat. J. Biochem. Cell Biol. 2002. V. 34. № 11. P. 1340–1354.
Odum J., Tinwell H., Tobin G., Ashby J. Cumulative dietary energy intake determines the onset of puberty in female rats // Environ. Health Perspect. 2004. V. 112. №15. P. 1472–1480.
Shaddock J.G., Chou M.W., Casciano D.A. Effects of age and caloric restriction on cell proliferation in hepatocyte cultures from control and hepatectomized Fischer 344 rats // Mutagenesis. 1996. V. 11. №3. P. 281–284.
Speakman J.R. Correlations between physiology and lifespan – two widely ignored problems with comparative studies // Aging Cell. 2005. V. 4. № 4. P. 167–175.
Spindler S.R. Caloric restriction: from soup to nuts // Age. Res. Rev. 2010. V. 9. № 3. P. 324 –353.
Stipp D. Linking nutrition, maturation and aging: from thrifty genes to the spendthrift phenotype // Aging (Albany NY). 2011. V. 3. № 2. P. 85–93.
West G.B., Woodruff W.H., Brown J.H. Allometric scaling of metabolic rate from molecules and mitochondria to cells and mammals // PNAS USA. 2002. V. 99. P. 2473–2478.
West G.B., Brown J.H. The origin of allometric scaling laws in biology from genomes to ecosystems: towards a quantitative unifying theory of biological structure and organization // J. Exp. Biol. 2005. V. 208. P. 1575–1592.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии