

Успехи современной биологии, 2022, T. 142, № 1, стр. 90-104
Изменчивость гена цитохрома b мтДНК полевки-экономки (Alexandromys oeconomus Pallas, 1776) Северного Охотоморья
В. В. Переверзева 1, *, Н. Е. Докучаев 1, А. А. Примак 1, Е. А. Дубинин 1, С. В. Киселев 1
1 Институт биологических проблем Севера Дальневосточного отделения Российской академии наук
Магадан, Россия
* E-mail: vvpereverzeva@mail.ru
Поступила в редакцию 21.07.2021
После доработки 23.08.2021
Принята к публикации 13.09.2021
- EDN: XQXFBC
- DOI: 10.31857/S0042132422010057
Аннотация
Исследован полиморфизм нуклеотидной последовательности и гаплотипическое разнообразие гена цитохрома b в выборках полевки-экономки (Alexandromys oeconomus) из 6 материковых и 4 островных популяций Северного Охотоморья. Найдено 58 гаплотипов гена цитохрома b (32 центрально-азиатской и 26 берингийской филогрупп), различающихся между собой 96-ю нуклеотидными заменами. Генофонды полевок-экономок о. Парамушир и о. Талан представлены уникальными для каждого из этих островов гаплотипами гена цитохрома b, не обнаруженными в других популяциях этого вида. Установлена идентичность части нуклеотидных замен в последовательностях гена cytb особей, населяющих острова и сопредельные им материковые территории. Это указывает на общность их происхождения. Значения индексов молекулярного разнообразия в большинстве исследованных выборок свидетельствуют о стабильности популяций, имеющих в течение длительного времени высокое значение эффективной численности. Генетические различия между большинством исследованных выборок статистически достоверны. Своеобразие генетической структуры популяций A. oeconomus Северного Охотоморья сформировалось в результате заселения данной территории на западе представителями центрально-азиатской, а на востоке – берингийской гаплогруппы и образованием в бассейнах рек Дукча и Ола смешанных популяций. В настоящее время линия разграничения этих клад расположена несколько восточнее г. Магадан.
ВВЕДЕНИЕ
Полевка-экономка (Alexandromys (=Microtus) oeconomus Pallas, 1776) имеет обширный голарктический ареал, охватывающий преимущественно северные территории Евразии и Северной Америки (Воронцов и др., 1986; Павлинов, Лисовский, 2012; Shenbrot, Krasnov, 2005). На севере Дальнего Востока России это один из фоновых видов мышевидных грызунов, занимающий сырые участки по долинам рек, берегам озер, болот и луговин (Юдин и др., 1976; Чернявский, 1984). Вдоль западного побережья Охотского моря ареал полевки-экономки простирается на юг до бассейна р. Уда (Костенко, 2000). Населяет она также северные и центральные острова Курильской гряды (Костенко и др., 2004) и некоторые острова Тауйской губы Охотского моря (Талан, Недоразумения, Вдовушка), включая мелкие острова Ольской лагуны (Сикулун и Уратамлян). Митохондриальный ген цитохрома b (сytb) кодирует одноименный трансмембранный фермент, который является ключевым звеном дыхательной цепи переноса электронов. Уровень функциональной активности этого энзима играет важнейшую роль в жизнеобеспечении и возможности адаптации организма к различным экологическим условиям. Полиморфизм нуклеотидной последовательности гена сytb широко используется в популяционной генетике и молекулярной филогеографии различных видов млекопитающих (Переверзева, Павленко, 2014; Григорьева и др., 2015; Малярчук и др., 2015; Переверзева и др., 2018; Esteva et al., 2010; Petrova et al., 2014; Kohli et al., 2015), в том числе и полевки-экономки (Abramson, Tikhonova, 2005; Brunhoff et al., 2006; Fink et al., 2010; Dabrowski et al., 2013; Jancewicz et al., 2015). У этого вида на основании разнообразия нуклеотидной последовательности cytb были выделены четыре гаплогруппы – североевропейская, центрально-европейская, центрально-азиатская и берингийская (Brunhoff et al., 2003). На территории Северо-Восточной Азии обитают A. oeconomus только центрально-азиатской и берингийской филогрупп (Brunhoff et al., 2003; Galbreath, Cook, 2004; Abramson, Tikhonova, 2005; Iwasa et al., 2009; Haring et al., 2011; Lissovsky et al., 2018). Ранее было установлено, что в верховьях р. Омолон области распространения полевок-экономок данных гаплогрупп перекрываются с образованием смешанных популяций (Galbreath, Cook, 2004). При этом внутрипопуляционная генетическая изменчивость нуклеотидной последовательности гена сytb у A. oeconomus Северного Охотоморья оставалась малоизученной. Следует особо отметить важность этой территории в зоогеографическом отношении, служившей своеобразным “экологическим коридором”, по которому проходили основные миграционные пути расселения растений и животных, в том числе мелких млекопитающих (Докучаев, 2012, 2019). В настоящее время исследование структуры генофондов популяций является одной из важнейших частей комплексного анализа видовой изменчивости. Особое значение имеет изучение изолированных внутривидовых группировок. Идеальными объектами таких исследований являются островные популяции. Сравнительный анализ генетической структуры континентальных и островных популяций позволяет реконструировать историю формирования генофонда видов на различных участках ареала. Такие задачи успешно решаются с применением методов молекулярно-генетического анализа мтДНК (Джикия и др., 2007; Переверзева и др., 2013а,б, 2018; Hinten et al., 2003; Suzuki et al., 2004, 2015; Fulgione et al., 2008; Hanazaki et al., 2017). Цель проведенного исследования – сравнительный анализ генетической структуры и филогенетических связей островных и континентальных популяций A. oeconomus Северного Охотоморья по данным полиморфизма гена cytb мтДНК.
МАТЕРИАЛЫ И МЕТОДЫ
Были исследованы десять выборок полевки-экономки из материковых и островных охотоморских популяций (рис. 1). Характеристики выборок представлены в табл. 1. Выделение, очистку тотальной ДНК, амплификацию нуклеотидной последовательности гена cytb, очистку продукта полимеразной цепной реакции (ПЦР) и секвенирование полной нуклеотидной последовательности гена cytb мтДНК проводили ранее описанными методами (Переверзева и др., 2018). Ген cytb картировали относительно полной нуклеотидной последовательности мтДНК Alexandromys (=Microtus) fortis fortis (GenBank № JF261174). Для определения филогенетических отношений гаплотипов полевок-экономок Северного Охотоморья из GenBank были взяты сведения о строении полной нуклеотидной последовательности гена cytb у образцов AY305201, AY305205 центрально-азиатской AY305090, AY305091 (Brunhoff et al., 2003) и AB372197–AB372200 (Iwasa et al., 2009) берингийской филогрупп. Гаплотипы особей AB372197–AB372200, отловленных на п-ове Камчатка, использованы в сопоставлении строения нуклеотидных последовательностей берингийской клады и филогенетических отношений популяций. Гаплотипам гена cytb A. oeconomus Северного Охотоморья центрально-азиатской генетической линии нами присвоена аббревиатура EcbСА, берингийской – EcbBr. Ниже приведены номера выявленных гаплотипов гена cytb в электронной базе данных GenBank: MT081259–MT081261, MT085481–MT085490, MT093801–MT093817, MT109502–MT109508, MT125678, MT125679, MT127790–MT127792, MT953663, MT953664, MT953667, MT953668, MT936870–MT936874, MW598279–MW598284, MZ020156. В качестве внешней группы использовали последовательность нуклеотидов гена cytb Craseomys (=Clethrionomys) rufocanus, GenBank № KR059876 (Переверзева и др., 2018). Статистическая обработка и анализ генетических данных проводились с помощью пакетов программ MEGA 10.0.2.74 (Tamura et al., 2013) и ARLEQUIN ver. 3.5 (Excoffier et al., 2005). Детекция филогенетических связей гаплотипов гена cytb проводилась с использованием пакета программ MEGA. Дендрограммы филогенетических отношений построены на основании бипараметрической модели дистанций Кимуры, учитывающей разную вероятность транзиций и трансверсий и выбранной с помощью байесовского информационного критерия. Дендрограмма гаплотипов гена cytb строилась по методу максимального правдоподобия (ML); популяций – по методу ближайшего связывания (NJ). Оценка узлов ветвления осуществлялась бутстреп-методом (1000 итераций).
Рис. 1.
Карта-схема мест сбора проб полевки-экономки центрально-азиатской и берингийской филогрупп (показаны соответственно кружками с черной и белой заливкой); в случае присутствия обеих филогрупп их доля соответствует размеру выделенных секторов.
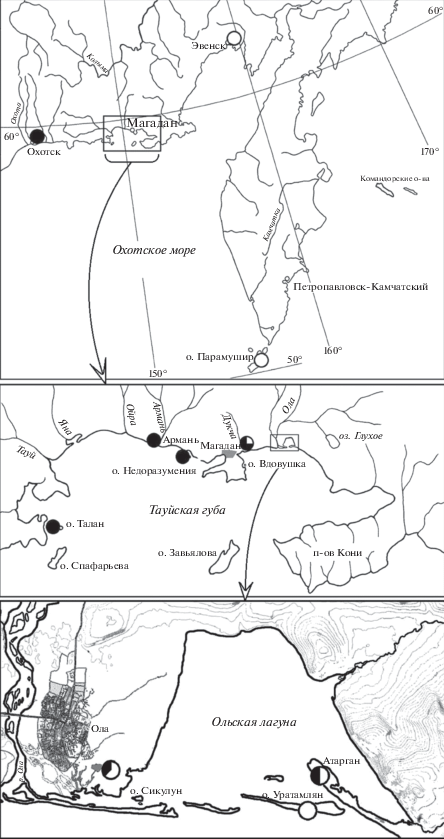
Таблица 1.
Характеристика выборок полевки-экономки из популяций Северного Охотоморья
| Место сбора материала | N | n | Доля гаплотипов филогрупп | Индекс молекулярного разнообразия | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| EcbСА | EcbBr | V | Pi ± sd | π ± sd | h ± sd | ||||||
| Континентальная часть (окрестности населенных пунктов) | |||||||||||
| Магадан | 60 | 20 | 0.7500 | 0.2500 | 71 | 18.7763 ± 8.4363 | 0.0164 ± 0.0082 | 0.8904 ± 0.0209 | |||
| Ола | 36 | 13 | 0.3056 | 0.6944 | 49 | 17.7317 ± 8.0609 | 0.0155 ± 0.0078 | 0.8825 ± 0.0340 | |||
| Армань | 17 | 6 | 1.00 | 0.00 | 7 | 2.9559 ± 1.6267 | 0.0026 ± 0.0016 | 0.7647 ± 0.0929 | |||
| Охотск | 7 | 5 | 1.00 | 0.00 | 9 | 4.0000 ± 2.2718 | 0.0035 ± 0.0023 | 0.9048 ± 0.1033 | |||
| Эвенск | 17 | 6 | 0.00 | 1.00 | 16 | 6.6912 ± 3.3208 | 0.0059 ± 0.0033 | 0.8603 ± 0.0415 | |||
| Атарган | 2 | 2 | 0.50 | 0.50 | 39 | 39.0000 ± 27.9285 | 0.0341 ± 0.0346 | 1.0000 ± 0.5000 | |||
| Острова | L | S | t | Острова Тауйской губы | |||||||
| Недоразумения* | 2.2 | 4.5 | 7 | 16 | 2 | 1.00 | 0.00 | 2 | 0.2500 ± 0.2966 | 0.0002 ± 0.0003 | 0.1250 ± 0.1064 |
| Талан*** | 7 | 1.6 | 10 | 7 | 2 | 1.00 | 0.00 | 2 | 0.5714 ± 0.5208 | 0.0005 ± 0.0005 | 0.2857 ± 0.1964 |
| Уратамлян** | 0.2 | 0.33 | нд | 11 | 1 | 0.00 | 1.00 | 0 | 0.0000 ± 0.0000 | 0.0000 ± 0.0000 | 0.0000 ± 0.0000 |
| Курильские острова | |||||||||||
| Парамушир*** | 11 | 2053 | 10 | 29 | 6 | 0.00 | 1.00 | 8 | 2.5813 ± 1.4244 | 0.0023 ± 0.0014 | 0.6626 ± 0.0820 |
| Характеристика молекулярного разнообразия филогрупп | |||||||||||
| Центрально-азиатская | 104 | 32 | 1.00 | 0.00 | 42 | 5.7526 ± 2.7762 | 0.0050 ± 0.0027 | 0.9175 ± 0.0140 | |||
| Берингийская | 98 | 26 | 0.00 | 1.00 | 46 | 7.7494 ± 3.6406 | 0.0068 ± 0.0035 | 0.9106 ± 0.0161 | |||
| Общая выборка | 202 | 58 | 0.5149 | 0.4851 | 96 | 22.056 ± 9.7591 | 0.0193 ± 0.0095 | 0.9573 ± 0.0053 | |||
Примечание: L – расстояние от острова до материка (км); S – площадь острова (км2); t – время отделения острова от материка (тыс. л. н.); N – объем выборок из популяций; n – число гаплотипов; EcbСА – центрально-азиатская филогруппа; EcbBr – берингийская филогруппа; V – число вариабельных сайтов в гене cytb мтДНК; Pi – среднее число попарных различий между гаплотипами; π – нуклеотидное разнообразие; h – гаплотипическое разнообразие; sd – стандартное отклонение, нд – нет данных. * – https://ru.wikipedia.org/wiki/Остров_Недоразумения; ** – по (Хорева и др., 2016); *** – по (Велижанин, 1976); https://ru.wikipedia.org/wiki/Парамушир.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Характеристика выборок полевки-экономки из популяций Северного Охотоморья и нуклеотидных замен в кодонах гена цитохрома b
Исследована полная нуклеотидная последовательность гена cytb двухсот двух экземпляров А. oeconomus из шести материковых и четырех островных выборок (табл. 1). Ген cytb расположен с 14118-ой по 15260-ю пару нуклеотидов (п.н.) мтДНК и состоит из 1143 п.н. У изученных полевок-экономок определено 58 гаплотипов гена cytb, различающихся между собой 96-ю нуклеотидными заменами. Доля замен в нуклеотидной последовательности cytb полевок клады EcbCA – 0.0367, филогруппы EcbBr – 0.0402. Суммарная доля вариабельных сайтов составляет 0.0840 от общей длины гена cytb. Замены относительно нуклеотидной последовательности EcbCA1 и номера выявленных гаплотипов гена cytb в электронной базе данных GenBank представлены на рис. 2. Характеристика обнаруженных нуклеотидных замен показана в табл. 2.
Рис. 2.
Гаплотипы гена цитохрома b полевки-экономки из популяций Северного Охотоморья. Нуклеотидные замены представлены относительно последовательности варианта EcbCA1. Сайты замен показаны от начала гена цитохрома b.

Таблица 2.
Локализация нуклеотидных замен в кодонах гена цитохрома b у полевки-экономки Северного Охотоморья
| Филогруппа | N | n | k | Доля замен | |||||
|---|---|---|---|---|---|---|---|---|---|
| транзиция | трансверсия | ||||||||
| позиция замены в кодоне | позиция замены в кодоне | ||||||||
| 1 | 2 | 3 | 1 | 2 | 3 | ||||
| Центрально-азиатская | 104 | 42 | 32 | 0.2143 | 0.0476 | 0.6190 | 0.0238 | 0.0238 | 0.0714 |
| Берингийская | 98 | 46 | 26 | 0.1087 | 0.0870 | 0.7174 | 0.00 | 0.00 | 0.0870 |
| Между EcbCA1и EcbBr1 | – | 38 | – | 0.1316 | 0.00 | 0.7105 | 0.00 | 0.00 | 0.1579 |
| Общая выборка | |||||||||
| Между центрально-азиатской и берингийской | 202 | 52 | 32/26 | 0.1154 | 0.0577 | 0.7115 | 0.00 | 0.00 | 0.1154 |
| Суммарно в центрально- азиатской и берингийской | 202 | 96 | 58 | 0.1458 | 0.0521 | 0.6771 | 0.0104 | 0.0104 | 0.1042 |
В Охотоморье у полевки-экономки клады EcbСА и EcbBr найдены примерно в равных долях. Общее количество нуклеотидных замен и гаплотипов в генетических линиях так же сопоставимо. Транзиций в нуклеотидной последовательности cytb A. oeconomus обеих клад обнаружено на порядок больше, чем трансверсий, что соответствует литературным данным (Nei, 1987; Nei, Kumar, 2000). Соотношение транзиций/трансверсий в генетической линии EcbСА составляет 7.4, EcbBr – 10.5. Полученный результат согласуется с данными Брунхофф с соавт. (Brunhoff et al., 2003). Однако, между гаплотипами EcbCA1 и EcbBr1, а также между кладами EcbСА и EcbBr и в общей выборке А. oeconomus доля трансверсии в третьей позиции кодона одного порядка с транзициями в первой и третьей позициях кодона и сопоставима с долями транзиций в первой позиции (табл. 2).
Вырожденность кода определяет наибольшую вариабельность третьего нуклеотида у большинства кодонов в транслируемых участках гена (Zardoya, Meyer, 1996). Полученные нами результаты отличаются от обычного распределения нуклеотидных замен в триплетах (2 : 1 : 9 в первой, второй и третьей позициях соответственно (Hassanin et al., 1998)). У полевки-экономки линии EcbСА в гене cytb доля транзиций в первой, второй и третьей позициях кодона составляет 4.5 : 1 : 13. У экземпляров EcbBr-клады – 1.2 : 1 : 8.2. Транзиции в триплетах, различающие у полевки-экономки эти генетические группы, соотносятся как 2 : 1 : 12.3. В суммарной выборке – 2.8 : 1 : 13. Трансверсии в нуклеотидной последовательности cytb у особей EcbСА клады соответствуют 1 : 1 : 3. У представителей филогруппы EcbBr найдены трансверсии в последовательности нуклеотидов этого гена только в третьей позиции кодона (табл. 2). Можно предположить, что такое распределение замен в триплетах гена cytb является видовой особенностью A. oeconomus.
Характеристика гаплотипов гена цитохрома b в выборках из популяций полевки-экономки материковой части Северного Охотоморья
На рис. 3 показаны доли вариантов cytb-гаплотипов в выборках A. oeconomus из исследуемых материковых популяций. Выборки полевок-экономок из окрестностей г. Магадан, пос. Ола и пос. Атарган состоят из особей EcbСА- и EcbBr-генетических линий.
Рис. 3.
Доли гаплотипов гена цитохрома b в выборках полевки-экономки из материковых популяций Северного Охотоморья (гаплотипы центрально-азиатской и берингийской филогрупп показаны соответственно столбцами с черной и белой заливкой, здесь и на рис. 4).
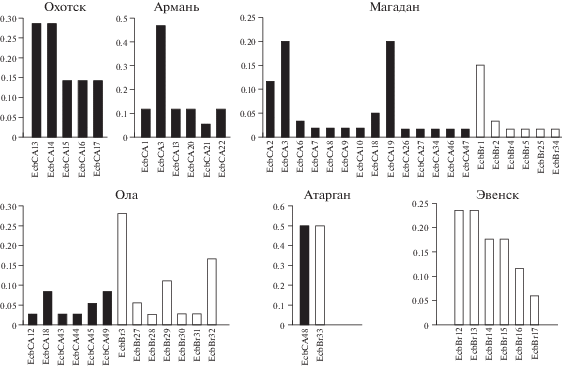
В окрестностях г. Магадан обитают преимущественно EcbСА-особи (табл. 1). Следует отметить, что в данную наиболее полиморфную выборку вошли экземпляры, отловленные в разные годы и в разных местах, что могло отразиться на уровне полиморфизма. Возле пос. Ола пойманы в основном EcbBr-особи. Это вторая по величине индексов молекулярного разнообразия выборка. В окрестностях пос. Атарган отловлено всего две полевки-экономки, относящиеся к EcbСА- и EcbBr-кладам. Выборки из популяций A. oeconomus окрестностей пос. Армань и Охотск состоят из EcbСА-особей. Все полевки-экономки из популяции окрестностей пос. Эвенск относятся к филогруппе EcbBr. Закономерно, что нуклеотидное разнообразие (π) и среднее число попарных различий между гаплотипами (Pi) в выборках A. oeconomus, состоящих из особей обеих филогрупп (окрестности г. Магадан, пос. Ола и пос. Атарган), на порядок превышают таковые в выборках, включающих особей одной генетической клады (окрестности пос. Армань, Охотск и Эвенск) (табл. 1).
Характеристика гаплотипов гена цитохрома b в выборках из популяций полевки-экономки о-вов Тауйской губы и о. Парамушир Курильской гряды
Формирование генофондов островных популяций происходит в результате действия стохастических процессов на частоты аллелей, количество которых изначально ограниченно эффектом основателя. Уникальность генетической структуры таких сообществ зависит также от времени и степени изоляции. На о. Недоразумения (рис. 1) большинство популяции составляют полевки-экономки генотипа EcbСА12 (рис. 4). Этот гаплотип также был найден только у одного экземпляра, отловленного вблизи пос. Ола (рис. 3). Гаплотип EcbСА11, отличающийся от EcbСА12 двумя транзициями (рис. 2) в третьей позиции триплета, обнаружен у одной особи с о. Недоразумения. Время изоляции этого острова (7 тыс. л. н., табл. 1) достаточно для формирования уникального генофонда популяции полевок-экономок. По-видимому, при отделении острова на нем случайно оказались особи генотипа EcbСА12, однако нельзя исключить присутствие полевок-экономок и других гаплотипов. В дальнейшем в результате стохастических процессов частота гаплотипа EcbСА12 стала преобладать. Возможно, вследствие мутаций появился EcbСА11. Низкая доля этого гаплотипа объясняется либо его недавним возникновением, либо дрейфом генов, если EcbСА11-особи оказались на отделившейся от материка территории.
Рис. 4.
Доли гаплотипов гена цитохрома b в выборках из популяций полевки-экономки о-вов Тауйской губы и о. Парамушир Курильской гряды.
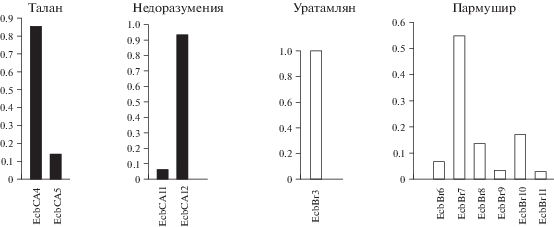
На о. Талан преобладают полевки-экономки генотипа EcbСА4. Особи с гаплотипом EcbСА5, который отличается от нуклеотидной последовательности EcbСА4 двумя транзициями в третьей позиции триплета (С444T, G303A (рис. 2)), встречаются реже (рис. 4), причем оба гаплотипа уникальные. Подобный генофонд A. oeconomus о. Талан сформировался в результате тех же микроэволюционных процессов, что и в популяции этого вида на о. Недоразумения. Однако обращает на себя внимание сходство нуклеотидных последовательностей EcbCA4 с EcbCA11 и EcbCA5 с EcbCA12 (рис. 2). По-видимому, транзиции в третьей позиции нуклеотидов A303G и C444T, относительно последовательности EcbCA1, возникли в генотипе A. oeconomus до отделения этих островов от материка, то есть ранее 7–10 тыс. л. н.
Небольшой о. Уратамлян (табл. 1), распложенный в Ольской лагуне (рис. 1), заселен полевками-экономками с гаплотипом EcbBr3 (рис. 4), который наиболее распространен у особей, обитающих вблизи пос. Ола (рис. 3). По-видимому, действие дрейфа генов на популяцию этого вида, ограниченную размерами территории островка, оказывает достаточно сильный прессинг, не позволяя закрепиться другим вариантам cytb-гаплотипов в генофонде полевок-экономок этого острова.
Генофонд A. oeconomus крупного острова Парамушир (табл. 1) Курильской гряды включает гаплотипы EcbBr6–EcbBr11, не найденные у особей других исследованных нами популяций. Транзиции в третьей позиции триплета Т165С, A816G и C837T (рис. 2 и 5) являются уникальными и присутствуют у всех нуклеотидных последовательностей cytb полевок-экономок этого острова. Следует отметить, что cytb-гаплотипы A. oeconomus, населяющих о. Парамушир и окрестности пос. Эвенск, не только значительно отличаются от остальных нуклеотидных последовательностей субкластера EcbBr (рис. 5), но и имеют определенное сходство между собой и с вариантами cytb AB372197–AB372200-экземпляров (рис. 2 и 5), отловленных на п-ове Камчатка. Все эти гаплотипы относительно нуклеотидной последовательности EcbBr1 имеют в третьей позиции кодона трансверсию С1137А и транзиции G882A, G912A. Кроме того, гаплотипы EcbBr13, EcbBr15–EcbBr17 (пос. Эвенск), EcbBr7–EcbBr11 (о. Парамушир) и AB372197–AB372200 содержат транзицию в третьей позиции кодона G297A. Гаплотипы EcbBr12, EcbBr14 (пос. Эвенск), EcbBr7–EcbBr11 и AB372197–AB372200 включают транзиции в третьей позиции кодона Т1050С и С1116Т (рис. 5). Полученные результаты свидетельствуют о возникновении этих нуклеотидных замен до отделения территории о. Парамушир от континентальной части, то есть ранее 10 тыс. л. н. Было обнаружено, что индексы молекулярного разнообразия нуклеотидных последовательностей cytb полевок-экономок, населяющих небольшие острова – Недоразумения, Талан и Уратамлян, на порядок меньше, чем аналогичные показатели открытых материковых популяций, состоящих из особей одной генетической клады – EcbCA или EcbBr. В тоже время данные параметры полевок-экономок о. Парамушир сопоставимы со значениями этих индексов в выборке особей из окрестностей пос. Армань (табл. 1). По-видимому, обширная площадь о. Парамушир препятствует сильному действию стохастических процессов в популяции мелких млекопитающих. Это позволило сформироваться в изолированной группе A. oeconomus генофонду, достаточно полиморфному по нуклеотидной последовательности cytb.
Рис. 5.
Гаплотипы гена цитохрома b полевки-экономки берингийской клады из популяций Северного Охотоморья. Нуклеотидные замены представлены относительно последовательности варианта EcbBr1. Сайты замен показаны от начала гена цитохрома b.

В целом было найдено, что, за исключением островных популяций Тауйской губы, индексы молекулярного разнообразия имеют высокие значения для всех исследованных выборок полевок, а также объединенной группы А. oeconomus (табл. 1). Ранее было показано (Avise, 2000), что большие значения индексов молекулярного разнообразия означают стабильность популяции, имеющей в течение длительного времени высокое значение эффективной численности. По-видимому, это соответствует всем исследованным сообществам А. oeconomus, возможно, за исключением населения крайне небольшого о. Уратамлян.
Филогенетические связи гаплотипов гена цитохрома b полевок-экономок Северного Охотоморья
Результаты детекции филогенетических связей гаплотипов гена cytb представлены на дендрограмме (рис. 6). Наличие двух крупных кластеров с высокими значениями бутстреп-индексов обусловлено генетической обособленностью центрально-азиатской и берингийской линий A. oeconomus. Кластеры полиморфны по структуре и подразделяются на субкластеры, часть из которых имеет значимые бутстреп-индексы в узлах ветвей. В субкластере EcbCA-филогруппы вариант EcbCA1, имеющий наименьшее количество замен с нуклеотидными последовательностями других гаплотипов этой клады, возможно, является предковым. Однако данный вариант обнаружен только у особей, населяющих окрестности пос. Армань, причем он не превалирует у полевок-экономок этой выборки (рис. 3). В субкластере центрально-азиатской генетической линии наибольшее значение бутстреп-индекса (94%) у субклады гаплотипов EcbCA7, EcbCA10, EcbCA19. Полевки-экономки с этими гаплотипами были отловлены в окрестностях г. Магадан. По-видимому, в данной субкладе родоначальным является EcbCA19, имеющий минимальное количество нуклеотидных замен в этой группе по отношению к последовательности нуклеотидов EcbCA1 (рис. 2). Значимые бутстреп-индексы в узлах ветвления имеют субклады EcbCA14, EcbCA16 (84%) и EcbCA13, EcbCA17 (73%). Вероятно, данную генетическую обособленность можно объяснить географической отдаленностью, так как полевки-экономки этих гаплотипов отловлены в окрестностях пос. Охотск (рис. 1, 3).
Рис. 6.
ML-филогенетическое дерево, построенное по данным об изменчивости нуклеотидной последовательности гена цитохрома b мтДНК полевки-экономки из популяций Северного Охотоморья. В узлах ветвления указаны бутстреп-индексы (> 50%).

В субкластере берингийской генетической линии наибольшее значение бутстреп-индекса (97%) у субклады гаплотипов EcbBr6–EcbBr11. Эти варианты нуклеотидных последовательностей гена cytb найдены у A. oeconomus о. Парамушир (рис. 4), крупного острова, который отделился от континента около 10 тыс. л. н. (табл. 1). Длительный срок изоляции позволил микроэволюционным процессам сформировать уникальный генофонд популяции. Гаплотипы полевок-экономок, отловленных в окрестностях пос. Эвенск (рис. 3) образуют два субкластера со значимыми бутстреп-индексами – EcbBr12, EcbBr14 (96%) и EcbBr13, EcbBr15–EcbBr17 (71%).
Филогенетические отношения исследованных популяций полевок-экономок Северного Охотоморья
На рис. 7 показаны филогенетические отношения между популяциями полевки-экономки. Дендрограмма содержит два крупных кластера, подразделенных на компактно сгруппировавшиеся субкластеры, включающие в себя нуклеотидные последовательности центрально-азиатской и берингийской генетической линий и более обособленные ветви нуклеотидных последовательностей, смешанных по этому признаку популяций. Полевки-экономки о. Парамушир и п‑ова Камчатка образуют отдельный субкластер, что отражает генетическое родство этих групп.
Рис. 7.
Филогенетические отношения между некоторыми популяциями полевки-экономки Северного Охотоморья (NJ-дендрограмма). Масштабный отрезок соответствует числу нуклеотидных замен на один сайт.

В табл. 3 представлены генетические дистанции (популяционные попарные Fst) между исследованными выборками A. oeconomus.
Таблица 3.
Генетические дистанции между выборками из популяций полевок-экономок Северного Охотоморья
| Локальность | Окрестности населенного пункта | Острова | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Магадан | Ола | Атарган | Армань | Охотск | Эвенск | Недоразумения | Талан | Уратамлян | Парамушир | ||
| Окрестности населенных пунктов | Магадан | – | 0.00 ± 0.00 | 0.7387 ± 0.0403 | 0.0090 ± 0.0091 | 0.0270 ± 0.0139 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.0541 ± 0.0201 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Ола | 0.2709 | – | 0.2432 ± 0.0385 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.0180 ± 0.0121 | 0.00 ± 0.00 | |
| Атарган | –0.0573 н.д. |
–0.0327 н.д. |
– | 0.0090 ± 0.0091 | 0.0721 ± 0.0227 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.0451 ± 0.0203 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Армань | 0.1562 | 0.5763 | 0.6517 | – | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Охотск | 0.1438 | 0.5033 | 0.4546 н.д. | 0.3986 | – | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Эвенск | 0.4818 | 0.2499 | 0.5187 | 0.8594 | 0.8236 | – | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Острова | Недоразумения | 0.2056 | 0.5818 | 0.7953 | 0.5187 | 0.5885 | 0.8924 | – | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Талан | 0.1473 н.д. |
0.5311 | 0.5991 | 0.5057 | 0.5429 | 0.8497 | 0.9042 | – | 0.00 ± 0.00 | 0.00 ± 0.00 | |
| Уратамлян | 0.5377 | 0.1638 | 0.7675 | 0.9523 | 0.9577 | 0.5576 | 0.9958 | 0.9940 | – | 0.00 ± 0.00 | |
| Парамушир | 0.6099 | 0.4696 | 0.7999 | 0.9297 | 0.9239 | 0.6653 | 0.9525 | 0.9405 | 0.8331 | – | |
Статистически значимых различий между нуклеотидными последовательностями гена cytb полевок-экономок, отловленных в окрестностях г. Магадан, пос. Атарган и о. Талан, а также пос. Атарган и поселков Ола и Охотск не найдено. По-видимому, ограниченный объем выборок из окрестностей пос. Атарган (2 экз.), Охотска и о. Талан (по 7 экз.) (табл. 1) не позволил выявить реальные генетические отличия между этими популяциями. Кроме того, следует учитывать, что выборки полевок-экономок из окрестностей г. Магадан, поселков Ола и Атарган состоят из особей обеих генетических линий. Остальные выборки A. oeconomus имеют статистически достоверные различия (табл. 3). Наибольшие значения Fst найдены при сопоставлении островных выборок (табл. 3). Максимальное значение (Fst = = 0.9958) обнаружено между популяциями о-вов Недоразумения и Уратамлян, заселенными полевками-экономками разных генетических линий (табл. 1). Большие генетические дистанции и высокая степень статистической достоверности для островных популяций закономерны и отражают уровень генетической обособленности генофондов изолированных популяций, сформировавшихся в результате процессов микроэволюции. Наименьшее достоверное генетическое различие обнаружено между популяциями полевок-экономок, отловленных в окрестностях г. Магадан и пос. Охотск (Fst = 0.1438, Р < 0.05). Низкий уровень генетических различий выявлен также между выборками из популяций A. oeconomus в окрестностях г. Магадан и пос. Армань (Fst = 0.1562, Р < 0.05) и окрестностями пос. Ола и о. Уратамлян (Fst = = 0.1638, Р < 0.05). Остальные выборки из популяций генетически различаются с высокой достоверностью (Р = 0.00 ± 0.00) (табл. 3), что свидетельствует об изоляции исследованных континентальных популяций полевок-экономок.
Методом попарной дифференциации в пакете программ AMOVA рассчитана генетическая изменчивость групп популяций A. oeconomus, составленных по географическому критерию (рис. 1). В первую группу вошли выборки из популяций полевок-экономок, добытых в окрестностях г. Магадан, поселков Ола, Атарган, Охотск и Армань; во вторую – отловленных возле пос. Эвенск; каждая островная популяция представляет отдельную группу. При таком объединении доля межгрупповой генетической изменчивости составила 33.14%, внутригрупповая доля – 27.51% и внутрипопуляционная – 39.35%. Полученные результаты отражают высокий уровень изменчивости нуклеотидной последовательности гена cytb у представителей вида A. oeconomus, населяющих Северное Охотоморье.
ЗАКЛЮЧЕНИЕ
В результате исследования полиморфизма нуклеотидной последовательности и гаплотипического разнообразия гена цитохрома b шести материковых и четырех островных популяций A. oeconomus Северного Охотоморья было установлено, что полевки-экономки из окрестностей пос. Охотск и Армань, а также о-вов Талан и Недоразумения относятся к центрально-азиатской гаплогруппе. Населяющие о. Парамушир и окрестности пос. Эвенск – к берингийской. Вблизи г. Магадан, поселков Ола и Атарган обитают полевки-экономки обеих филогрупп. Все особи с небольшого о. Уратамлян Ольской лагуны принадлежат к берингийской гаплогруппе. Генофонды A. oeconomus о. Парамушир и о. Талан представлены уникальными для каждого из этих островов гаплотипами гена цитохрома b, не обнаруженными в других популяциях этого вида. Формирование генетической структуры генофондов изолированных групп полевок-экономок островных популяций является следствием синергии эффекта основателя и микроэволюционных процессов. Интенсивность дрейфа генов зависит от численности популяции, способной обитать на территории конкретного острова. Обнаружена идентичность части нуклеотидных замен в последовательностях гена cytb у полевок-экономок, населяющих острова и сопредельные им материковые территории, что свидетельствует об общности их происхождения.
Полученные результаты показывают, что A. oeconomus, заселяющие охотоморское побережье западнее г. Магадан, представлены центрально-азиатской филогруппой. В окрестностях Магадана большая часть популяции также состоит из полевок-экономок этой генетической линии, однако треть выборки составляют особи берингийской филогруппы. Восточнее Магадана в исследованных выборках доля полевок-экономок берингийской линии увеличивается, а возле пос. Эвенск и на о. Парамушир отловлены особи только этой генетической клады. Своеобразие выявленной генетической структуры популяций A. oeconomus Северного Охотоморья обусловлено ее формированием в результате постледникового заселения данной территории носителями центрально-азиатской и берингийской гаплогрупп. Разделение этих генетических линий произошло в среднем плейстоцене – 0.21–0.394 млн л. н. (Brunhoff et al., 2003; Galbreath, Cook, 2004). В позднем плейстоцене на Северо-Востоке Азии уже были представлены полевки-экономки обеих филогрупп. Носители центрально-азиатской линии занимали западную часть охотоморского побережья. Полевки-экономки берингийской линии населяли Камчатку и Чукотку, откуда в период низкого стояния уровня моря проникли на Аляску. Часть этой популяции осталась на о. Парамушир после его отделения от Камчатки. По-видимому, с окончанием последнего оледенения A. oeconomus берингийской клады с камчатского рефугиума расселились на запад по прибрежной части Охотского моря. Полевки-экономки центрально-азиатской линии в это же время расселялись на восток. В результате встречной экспансии двух филогрупп на северном побережье Тауйской губы в бассейнах рек Дукча и Ола образовались смешанные популяции A. oeconomus. В настоящее время линия разграничения ареалов этих клад расположена несколько восточнее г. Магадан.
Список литературы
Велижанин А.Г. Время изоляции материковых островов северной части Тихого океана // ДАН СССР. 1976. Т. 231. № 1. С. 205–207.
Воронцов Н.Н., Ляпунова Е.А., Боескоров Г.Г., Ревин Ю.В. Стабильность кариотипа полевки-экономки (Microtus oeconomus) в центральной части ареала и история становления современного ареала вида // Зоол. журн. 1986. Т. 65. Вып. 11. С. 1705–1715.
Григорьева О.О., Борисов Ю.М., Стахеев В.В. и др. Генетическая структура популяций обыкновенной бурозубки Sorex araneus L., 1758 (Mammalia, Lipotyphla) на сплошных и фрагментированных участках ареала // Генетика. 2015. Т. 51. № 6. С. 711–723.
Джикия Е.Л., Колесников А.А., Чудакова Д.А. и др. Генетический полиморфизм командорских популяций песцов // Генетика. 2007. Т. 43. № 9. С. 1239–1245.
Докучаев Н.Е. Особенности распространения землероек (Soricomorpha) и грызунов (Rodentia) на материковом побережье Охотского моря // Вестн. СВНЦ ДВО РАН. 2012. № 3. С. 118–123.
Докучаев Н.Е. Хребет Джугджур как географическая преграда для распространения животных (на примере землероек-бурозубок) // Вестн. СВНЦ ДВО РАН. 2019. № 4. С. 66–72.
Костенко В.А. Грызуны (Rodentia) Дальнего Востока России. Владивосток: Дальнаука, 2000. 210 с.
Костенко В.А., Нестеренко В.А., Трухин А.М. Млекопитающие Курильского архипелага. Владивосток: Дальнаука, 2004. 186 с.
Малярчук Б.А., Деренко М.В., Денисова Г.А. Изменчивость митохондриального генома росомахи (Gulo gulo) // Генетика. 2015. Т. 51. № 11. С. 1291–1296.
Павлинов И.Я., Лисовский А.А. Млекопитающие России: систематико-географический справочник. М.: Тов-во науч. изд. КМК, 2012. 604 с.
Переверзева В.В., Павленко М.В. Разнообразие строения гена цитохрома b митохондриальной ДНК полевой мыши Apodemus agrarius Pallas, 1771 из популяций юга Дальнего Востока России // Изв. РАН. Серия Биол. 2014. № 1. С. 5–16.
Переверзева В.В., Примак А.А., Дубинин Е.А. Генетическая структура популяций красной полевки Myodes (= Clethrionomys) rutilus Pallas, 1779 Северного Приохотья по данным об изменчивости нуклеотидных последовательностей гена цитохрома b митохондриальной ДНК // Вавил. журн. генетики и селекции. 2013а. Т. 17. № 3. С. 435–443.
Переверзева В.В., Примак А.А., Дубинин Е.А. Филогенетические отношения популяций красной полевки Myodes (=Clethrionomys) rutilus Pallas, 1779 Северного Приохотья и Колымского региона // Вавил. журн. генетики и селекции. 2013б. Т. 17. № 3. С. 444–451.
Переверзева В.В., Примак А.А., Докучаев Н.Е. и др. Изменчивость гена цитохрома b мтДНК красно-серой полевки (Craseomys rufocanus Sundevall, 1846) Северного Приохотья и бассейна р. Колыма // Вестн. СВНЦ. 2018. № 1. С. 101–112.
Хорева М.Г., Зеленская Л.А., Андриянова Е.А. Формирование растительного покрова на островных барах Ольской лагуны (Охотское море) в условиях быстрорастущей численности морских птиц // Сиб. экол. журн. 2016. № 3. С. 299–312.
Чернявский Ф.Б. Млекопитающие крайнего северо–востока Сибири. М.: Наука, 1984. 389 с.
Юдин Б.С., Кривошеев В.Г., Беляев В.Г. Мелкие млекопитающие Севера Дальнего Востока. Новосибирск: Наука, 1976. 270 с.
Abramson N.I., Tikhonova E.P. Reevaluation of taxonomic structure of the root vole (Microtus oeconomus Pallas, 1776, Rodentia, Arvicolidae) from the territory of the former USSR based on evidence from craniometric and molecular data // Russ. J. Theriol. 2005. V. 4. № 1. P. 63–73.
Avise J.C. Phylogeography: the history and formation of species. Cambridge, Massachusetts: Harvard Univ. Press, 2000. 447 p.
Brunhoff C., Galbreath K.E., Fedorov V.B. et al. Holarctic phylogeography of the root vole (Microtus oeconomus): implications for late Quaternary biogeography of high latitudes // Mol. Ecol. 2003. V. 12. № 4. P. 957–968.
Brunhoff C., Yoccoz N.G., Ims R.A., Jaarola M. Glacial survival or late glacial colonization? Phylogeography of the root vole (Microtus oeconomus) in north-west Norway // J. Biogeogr. 2006. V. 33. P. 2136–2144.
Dabrowski M.J., Pomorski J.J., Gliwicz J. Cytochrome b gene (cytb) sequence diversity in a Microtus oeconomus population from Bialowieza Primeval Forest // Acta Theriol. 2013. V. 58. P. 119–126.
Esteva M., Cervantes F. A., Brant S. V., Cook J. A. Molecular phylogeny of long-tailed shrews (genus Sorex) from México and Guatemala // Zootaxa. 2010. V. 2615. P. 47–65.
Excoffier L., Laval G., Schneider S. Arlequin ver. 3.0: an integrated software package for population genetics data analysis // Evol. Bioinform. Online. 2005. V. 1. P. 47–50.
Fink S., Fischer M.C., Excoffier L., Heckel G. Genomic scans support repetitive continental colonization events during the rapid radiation of voles (Rodentia: Microtus): the utility of AFLPs versus mitochondrial and nuclear sequence markers // Syst. Biol. 2010. V. 59. № 5. P. 548–572.
Fulgione D., Guglielmi S., Odierna G. et al. Morphological differentiation and genetic structure in island lizard populations // Zool. Sci. 2008. V. 25. P. 465–474.
Galbreath K.E., Cook J.A. Genetic consequences of Pleistocene glaciations for the tundra vole (Microtus oeconomus) in Beringia // Mol. Ecol. 2004. V. 13. P. 135–148.
Hanazaki K., Tomozawa M., Suzuki Y. et al. Estimation of evolutionary rates of mitochondrial DNA in two Japanese wood mouse species based on calibrations with Quaternary environmental changes // Zool. Sci. 2017. V. 34. № 3. P. 201–210.
Hassanin A., Lecointre G., Tiller S. Related articles, links abstract. The “evolutionary signal” of homoplasy in protein-coding gene sequences and its consequences for a priori weighting in phylogeny // C.R. Acad. Sci. 1998. V. 321. № 7. P. 611–620.
Haring E., Sheremetyeva I., Kryukov A. Phylogeny of Palearctic vole species (genus Microtus, Rodentia) based on mitochondrial sequences // Mamm. Biol. – Zeitschrift für Säugetierkunde. 2011. V. 76. № 3. P. 258–267.
Hinten G., Harriss F., Rossetto M., Braverstock P.R. Genetic variation and island biogeography: microsatellite and mitochondrial DNA variation in island populations of the Australian bush rat, Rattus fuscipes greyii // Conserv. Genet. 2003. V. 4. P. 759–778.
Jancewicz E., Falkowska E., Ratkiewicz M. mtDNA evidence for a local northern latitude Pleistocene refugium for the root vole (Microtus oeconomus, Arvicolinae, Rodentia) from Eastern Poland // J. Zool. Syst. Evol. Res. 2015. V. 53. № 4. P. 331–339.
Iwasa M.A., KostenkoV.A., Frisman L.V., Kartavtseva I.V. Phylogeography of the root vole Microtus oeconomus in Russian Far East: a special reference to comparison between Holarctic and Palaearctic voles // Mamm. Study. 2009. V. 34. P. 123–130.
Kohli B.A., Fedorov V.B., Waltari E., Cook J.A. Phylogeography of a Holarctic rodent (Myodes rutilus): testing high-latitude biogeographical hypotheses and the dynamics of range shifts // J. Biogeogr. 2015. V. 42. № 2. P. 377–389.
Lissovsky A.A., Petrova T.V., Yatsentyuk S.P. et al. Multilocus phylogeny and taxonomy of East Asian voles Alexandromys (Rodentia, Arvicolinae) // Zool. Scripta. 2018. V. 47. Iss. 1. P. 9–20.
Nei M. Molecular evolutionary genetics. N.Y.: Columbia Univ. Press, 1987. 495 p.
Nei M., Kumar S. Molecular evolution and phylogenetic. N.Y.: Oxford Univ. Press, 2000. 333 p.
Petrova T.V., Zakharov E.S., Samiya R., Abramson N.I. Phylogeography of the narrow-headed vole Lasiopodomys (Stenocranius) gregalis (Cricetidae, Rodentia) inferred from mitochondrial cytochrome b sequences: an echo of Pleistocene prosperity // J. Zool. Syst. Evol. Res. 2015. V. 53. № 2. P. 97–108.
Shenbrot G.I, Krasnov B.R. An atlas of the geographic distribution of the arvicoline rodents of the world (Rodentia, Muridae: Arvicolinae). Sofia: Pensoft Publishing, 2005. 336 p.
Suzuki H., Yasuda S.P., Sakaizumi M. et al. Differential geographic patterns of mitochondrial DNA variation in two sympatric species of Japanese wood mice, Apodemus speciosus and A. argenteus // Gen. Genet. Syst. 2004. V. 79. Iss. 3. P. 165–176.
Suzuki Y., Tomozawa M., Koizumi Y. et al. Estimating the molecular evolutionary rates of mitochondrial genes referring to Quaternary Ice Age events with inferred population expansions and dispersals in Japanese Apodemus // BMC Evol. Biol. 2015. V. 15. P. 187.
Tamura K., Stecher G., Peterson D. et al. MEGA-6: molecular evolutionary genetics analysis version 6.0. // Mol. Biol. Evol. 2013. V. 30. P. 2725–2729.
Zardoya R., Meyer A. Phylogenetic performance of mitochondrial protein-coding genes in resolving relationships among vertebrates // Mol. Biol. Evol. 1996. V. 13. № 7. P. 933–942.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии