

Успехи современной биологии, 2022, T. 142, № 1, стр. 70-89
Биоклиматическая система Волжского бассейна и ее ресурсный потенциал в регуляции углеродного цикла при глобальных изменениях климата (прогнозное эмпирико-статистическое моделирование)
Э. Г. Коломыц 1, *, Л. С. Шарая 1
1 Самарский исследовательский центр, Институт экологии Волжского бассейна РАН
Тольятти, Россия
* E-mail: egk2000@mail.ru
Поступила в редакцию 17.08.2021
После доработки 28.10.2021
Принята к публикации 28.10.2021
- EDN: OMUSKP
- DOI: 10.31857/S0042132422010045
Аннотация
По коренной лесной растительности Волжского бассейна описана региональная биоклиматическая система и проведен опыт ее прогнозного аналитического и картографического моделирования в свете известных сценариев глобальных изменений климата. Проведено эмпирическое обоснование выдвинутой В.Г. Горшковым и А.И. Уткиным концепции экологических ресурсов лесного покрова как его способности поглощать парниковые газы с помощью механизмов регуляции углеродного цикла и тем самым смягчать климатические колебания. Описана структура углеродно-балансовых ниш коренных лесов, раскрывающая различные зонально-региональные типы регуляции лесным покровом содержания СО2 в атмосфере при потеплении и похолодании. Прогнозируется, что при одном и том же термоаридном сигнале бореальные коренные леса будут преимущественно поглощать дополнительно парниковые газы, а суббореальные – выбрасывать их в атмосферу, оказывая соответствующее влияние на состояние наземных экосистем. На примере Окского бассейна проведено виртуальное прогнозное картографирование углеродного баланса по отдельным категориям коренных лесов, что важно для стратегии лесного хозяйства в лесодефицитных регионах.
ВВЕДЕНИЕ
Под региональной биоклиматической системой понимается упорядоченная совокупность пространственных связей почвенно-растительного покрова с ландшафтно-геофизическими факторами (Базилевич и др., 1986; Коломыц, 2003), то есть взаимодействие макро- и микросубстратных природных компонентов, по А.А. Григорьеву (1965), в организации эколого-географического пространства. Элементами системы являются климатические ниши фитоценологических или почвенных объектов, распределенные по гидротермическим градиентам и представляющие собой гиперпространственный, согласно Ю. Одуму (1975), а также ресурсный, по Ю.М. Свирежеву (1982), аспекты экологических ниш. В отношении растительного покрова региональная биоклиматическая система определяет граничные условия формирования фитоценотических единиц топологического и субрегионального уровней (ассоциаций, их групп и классов) под преломляющим воздействием местных геоморфологических и эдафических факторов.
Операционными единицами лесной растительности как одного из представителей макросубстрата в биоклиматической системе служат коренные (первичные, или девственные) леса, которые наиболее чувствительны к изменениям климата (Сукачев, 1975) и отображают зонально-провинциальные закономерности организации природной среды. Коренные леса Земли сохранились на площади около 13.5 млн кв. км, что составляет 40% территории суши. Из них почти половину площади составляют бореальные леса, среди которых более 51% приходится на коренные леса России (Bryant et al., 1997). Доля российских лесов достигает почти трети всех девственных лесов мира (Лосев, 2001), и российские леса обеспечивают свыше 90% стока углерода всех мировых бореальных лесов (Швиденко и др., 2014). В биоклиматической интерпретации – это климаксные и близкие к ним квазиклимаксные лесные сообщества (хвойные, смешанные и широколиственные), знаменующие собой заключительный этап эндоэкогенетических (восстановительных) сукцессий, по В.Н. Сукачеву (1972).
Современное глобальное потепление, вызванное увеличением выбросов парниковых газов в атмосферу, является свершившимся фактом. Климатические прогнозы, основанные на сценариях техногенных выбросов парниковых газов в атмосферу, предполагают повышение за период с 1990 г. по 2100 г. средней глобальной температуры поверхности Земли в пределах 1.4–5.8°С, что в 2–10 раз больше величины потепления, произошедшего в ХХ в. (Албриттон и др., 2003). Согласно Парижскому (2015) Соглашению по изменению климата (Paris Agreement, 2015), во избежание глобальной экологической катастрофы необходимо принимать меры, чтобы потепление к 2050 г. не превысило 1.5−2°С. Одной из этих мер является достижение баланса между возникающими в результате деятельности человека парниковыми газами и их поглощением морями и лесами (Ст. 4 Парижского Соглашения). Через процессы поглощения парниковых газов необходимо оценить роль естественных экосистем в регуляции углеродного цикла и стабилизации окружающей среды (Кондратьев и др., 2003).
Оценка пулов углеродного баланса пока не привела к формированию общей точки зрения на пространственное распределение наземных стоков и источников углерода, в том числе и в лесах России, где даже в недавних публикациях (Замолодчиков и др., 2007, 2011; Моисеев, Филипчук, 2009; Shvidenko et al., 2010) величины суммарного стока углерода варьируют от 100 до 700 Мт С в год. Возможные причины этого состоят в различии методов как процедур измерения стока углерода, так и интерполяции данных между точками измерений в условиях резко выраженных контрастов лесных экосистем, какими они нередко бывают.
Оценка потенциальных возможностей биоклиматической системы Волжского бассейна в смягчении климатических изменений, в том числе глобального потепления, является одной из актуальных экологических проблем этого обширного региона – индустриального и демографического “ядра” нашей страны. Опыт решения некоторых задач этой проблемы методами ландшафтной экологии представлен в настоящем сообщении.
Идеологической основой научного поиска стала выдвинутая В.Г. Горшковым (1995) и А.И. Уткиным (1995) и обобщенная К.С. Лосевым (2001) новая экологически ориентированная парадигма в учении о лесе. Речь идет об экологических ресурсах лесного покрова как его способности поглощать парниковые газы с помощью механизмов регуляции углеродного цикла при изменениях климата. Эта регуляция направлена на возвращение среды в оптимальное для лесных экосистем состояние и способствует сохранению относительной стабильности их продукционного процесса в меняющемся климате, что обеспечивает и устойчивость механизмов самой регуляции углеродного цикла как ведущего звена биологического круговорота.
Таковы основные экологические биосферно-стабилизирующие функции леса (Уткин, 1995; Исаченко и др., 2000), то есть ведущие “экосистемные услуги” лесного покрова (Тишков, 2005). Задача “поддерживать … возобновительную способность лесов … выполнять защитные функции лесных ресурсов … на местном, национальном и глобальном уровнях” входит составной частью стратегии устойчивого управления лесами (Швиденко и др., 2014, с. 14). Одним из важнейших направлений этой стратегии является использование лесов в качестве средства смягчения изменений климата (Швиденко и др., 2017) с помощью механизмов биотической регуляции углеродного цикла.
МАТЕРИАЛЫ И МЕТОДЫ
Исходными материалами послужили карты групп лесных формаций (ГЛФ) Европейской России (Растительность европейской …, 1980; Растительность европейской … и Кавказа, 1987), а также результаты наших крупномасштабных ландшафтных съемок, проведенных в лесной зоне Волжского бассейна (Коломыц, 2008) примерно в те же годы, когда создавались эти карты. Согласно В.Б. Сочаве (1979), данные фитоценологические единства относятся к классам (и подклассам) растительных формаций, которые являются региональными вариантами (например, восточноевропейскими или камско-печерско-предуральскими) типов и подтипов растительности (среднетаежного, подтаежного и т.д.). На территорию Волжского бассейна приходится 13 групп коренных растительных формаций (табл. 1).
Таблица 1.
Классификационная схема коренных растительных формаций основного водосбора Волжского бассейна и его окружения
| Фитоценологические единицы (по Грибова и др., 1980) |
Группы растительных формаций | |||
|---|---|---|---|---|
| зональные типы и классы |
региональные варианты | подзональные подтипы |
краткая характеристика | номер и символ |
| I. Темнохвойные и широколиственно-темнохвойные леса (производные осиново-березовые) | Восточноевропейские (верхневолжские) | Среднетаежные | Еловые кустарничково-зеленомошные | 1  |
| Южнотаежные | Еловые кустарничково-травяные | 2  |
||
| Подтаежные | Широколиственно-еловые сложные неморальнотравяные | 3  |
||
| Камско-Печерско-Пред- уральские |
Средне- и южнотаежные | Пихтово-еловые и елово-пихтовые травяно-кустарничковые зеленомошные и травяные | 4  |
|
| Подтаежные | Пихтово-еловые сложные неморальнотравяные |
5  |
||
| Широколиственно-пихтово-еловые неморальнотравяные | 6  |
|||
| II. Сосновые и широколиственно-сосновые леса (производные осиново-березовые) | Восточноевропейские (верхневолжские) | Средне- и южнотаежные | Сосновые с елью кустарничково-зеленомошные | 7  |
| Подтаежные | Сосновые (с дубом в подлеске) кустарничково-травяные | 8  |
||
| Широколиственно-сосновые и сосновые сложные с елью | 9  |
|||
| Лесостепные и степные | Сосновые и широколиственно-сосновые со степными кустарниками и разнотравно-злаковые | 10  |
||
| III. Широколиственные леса | Восточноевропейские | Северо-лесостепные | Липово-дубовые и дубовые | 11а  |
| Липовые с примесью других широколиственных пород | 11б 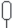 |
|||
| IV. Типичная и южная лесостепь |
Причерноморского типа | Типично-лесостепные | Луговые степи в сочетании с лесами из дуба | 12 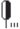 |
| Южно-лесостепные | Богато-разнотравно-типчаково-ковыльные степи | 13 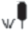 |
||
Новейшая “Карта лесных экосистем Северной Евразии”, составленная по спутниковым данным SPOT-Vegetation (Барталев и др., 2010; Разнообразие и динамика …, 2012) не могла быть использована. Выделенные на ней типы растительности (например, хвойные вечнозеленые леса, хвойные листопадные, лиственные леса вообще, смешанные с преобладанием хвойных и т.п.) имеют скорее лесопромышленное, нежели лесоведческое значение, что затрудняет интерпретацию этой карты в свете классических законов лесной биогеоценологии, согласно (Сукачев, 1972). Такие категории лесов совершенно не соответствуют принятому в традиционной геоботанике содержательному смыслу понятий “классы растительных формаций” и “типы растительности” (Работнов, 1978; Сочава, 1979), на основе которых и построена классификация растительного покрова Волжского бассейна.
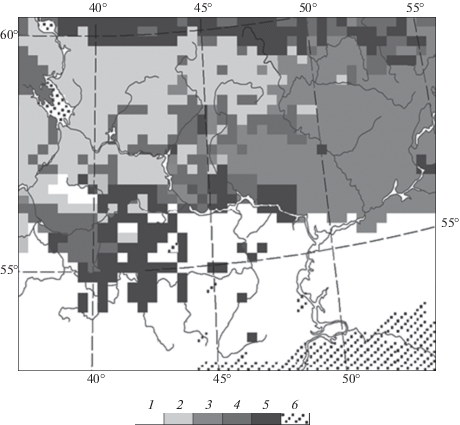
На рис. 1. представлена базовая растровая карта зонально-провинциальных групп растительных формаций Волжского бассейна 1 : 8 000 000. Карта построена на основе Геоботанической карты Европейской России (Растительность европейской … и Кавказа, 1987). В ареалы каждой из фитоценологических групп вошли коренные лесные сообщества (темно- и светлохвойные, смешанные, широколиственные) – как современные, так и восстановленные на месте длительно производных (мелколиственных) лесов, а также агроландшафтных комплексов. Таким образом, эта карта представляет собой модель биоклиматической системы лесов Волжского бассейна. Она отображает зонально-провинциальную фитоклиматическую структуру этой территории. На ее основе предстояло дать прогнозные оценки углеродного баланса лесного покрова, гипотетически представленного только коренными формациями, что весьма важно для оценок потенциальных возможностей регуляции лесным покровом Волжского бассейна углеродного цикла при климатических изменениях.
Рис. 1.
Растровая базовая карта зонально-провинциальных групп коренных растительных формаций (современных + + восстановленных) и распределение экспериментальных полигонов на территории основного водосбора Волжского бассейна. Экспериментальные полигоны (по: Коломыц, 2008): 1 – Жигули; 2 – Присурье; 3 – Зеленый Город; 4 – Щелоковский Хутор; 5 – Выкса; 6 – Керженец; 7 – Приокско-Террасный заповедник; 8 – Кудьма. Обозначения групп 1–13 растительных формаций см. в табл. 1.
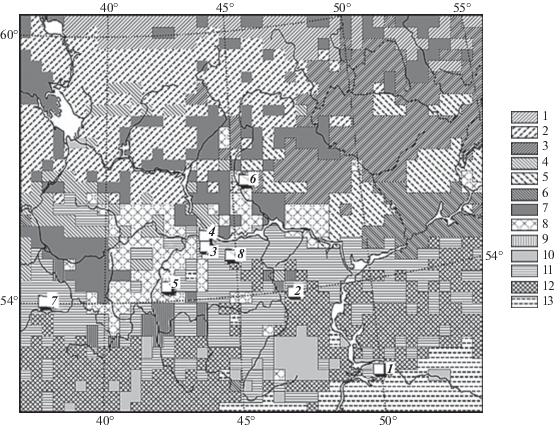
На карте показаны также места расположения 8 экспериментальных полигонов, на которых была проведена одноразовая крупномасштабная (1 : 50 000) ландшафтная съемка по специально разработанной программе (Коломыц, 2008). Каждый полигон включал 40–50 пробных площадей (лесных биогеоценозов), которые в совокупности характеризовали данную региональную экосистему. Среди 70 полученных параметров структурно-функционального состояния биогеоценозов были измерены, либо рассчитаны следующие фитомассы: BS – живая скелетная древесно-кустарниковая; BV – общая зеленая масса всех растительных ярусов; BR – общая масса корней; WD – мертвая древесно-кустарниковая фитомасса (дебрис); ML – масса лесной подстилки (опад + + отпад + мор); HU – масса лабильного гумуса в горизонте А1.
Для оценок изменений содержания углерода в различных биотических компонентах был применен известный лесоводственный метод (Морозов, 1949; Цельникер, 2006), основанный на динамике живых и мертвых фитомасс и дающий наиболее эффективные результаты за большие промежутки времени. Изменение (∆С) массы потока углерода в системе почва–растение–атмосфера представляется в наиболее общем виде (Кобак, 1988) как
где ∆С(Fa) – поток СО2 над растительным покровом; Rm – выделение СО2 в результате жизнедеятельности почвенных и наземных сапротрофов (преимущественно бактерий и грибов), разлагающих гумус, лесную подстилку и скелетный детрит; NPP – чистая первичная продукция биогеоценоза (надземная + корневая). Используя дискретные показатели малого биологического круговорота, приведенное выше уравнение было развернуто в следующей форме:(2)
$\begin{gathered} \Delta {\text{С}}(Fa) = \Delta {\text{С}}(WD) + \Delta {\text{С}}(ML) + \Delta {\text{С}}(HU)-- \\ - \,\,\Delta {\text{С}}(BS)--\Delta {\text{С}}(BV)--\Delta {\text{С}}(BR).~ \\ \end{gathered} $Здесь представлены углеродные пулы, содержащие упомянутые выше фитомассы.
Следует подчеркнуть, что рассчитываемые по уравнению (2) значения параметра ∆С(Fa) характеризуют динамический (так будем его называть) углеродный баланс лесных экосистем, обусловленный изменениями баланса депозита и эмиссии углерода в почвенно-растительном покрове под воздействием устойчивой смены климатических условий. Это, так сказать, баланс балансов – как соотношение прогнозных балансов и баланса конца базового периода. Динамический баланс коренным образом отличается от традиционного статического углеродного баланса, который рассчитывается для данных почвенно-геоморфологических и фитоценотических условий при стационарном климате. Как правило, статический углеродный баланс по своим абсолютным значениям на порядок больше баланса динамического.
По балансовому уравнению (2) рассчитывались возможные изменения потоков углерода между почвенно-растительным покровом и атмосферой в различные прогнозные сроки (по заданным климатическим моделям) для каждой группы коренных лесных сообществ во всех рассматриваемых региональных экосистемах. Каждый член правой части уравнения (2) может иметь как положительные, так и отрицательные значения. При положительных значениях (потепление) первые три слагаемые дают прибавку эмиссии СО2 из почвенно-растительного покрова в атмосферу, а вторые – убыль этого потока. При отрицательных же значениях указанных параметров (похолодание) – картина обратная. В итоге складывается общий баланс изменений углеродного обмена между биогеоценозами и атмосферой [±∆С(Fa)], который должен показать, поглощает ли данная группа лесных топоэкосистем дополнительное количество СО2 из атмосферы в результате обусловленных устойчивым климатическим сигналом сдвигов в биологическом круговороте или, наоборот, становится источником его дополнительных выбросов.
В результате расчетов были получены парциальные (по отдельным пулам-фитомассам) и суммарные значения базового содержания углерода и прогнозирумые углеродные балансы для каждой группы лесных биогеоценозов данного экспериментального полигона. Биогеоценозы группировались по элементам системы локальных ландшафтных сопряжений (Глазовская, 1988), а также по основным типам леса (Сукачев, 1972). По полученным формулам был произведен расчет величин всех шести метаболических параметров при прогнозируемых значениях температур воздуха и атмосферных осадков, которые трансформируются в гидротермические характеристики почвы.
Термо- и гидроэдафическая ординация метаболических характеристик лесных топоэкосистем проведена по двум геофизическим параметрам: температуре почвы на глубине 50 см (t50) и запасам летней продуктивной влаги в слое почвы 0–50 см. С этими параметрами функциональные характеристики лесных сообществ обнаруживают наиболее тесные связи. В табл. 2 приведен небольшой фрагмент расчетных формул, полный набор которых представлен в работе (Коломыц, 2008). По этим формулам определялись величины всех шести метаболических параметров при прогнозируемых значениях среднеянварских и среднеиюльских температур воздуха, а также годового количества осадков.
Таблица 2.
Расчетные модели, описывающие связи параметров биологического круговорота с температурой и влагосодержанием почвы в вегетационный период в лесных биогеоценозах экорегионов Среднего и Верхнего Поволжья
| Вид модели (расчетная формула)* | Экспериментальные полигоны | Параметры (см. в тексте) | Коэффициенты | Статистические характеристики** |
||||
|---|---|---|---|---|---|---|---|---|
| b0 | b1 | b2 | R2 | P | Sy | |||
| y = b0 + b1x1 + b2x2 | Присурье | BV | 21.93 | –0.7415 | –0.7135 | 0.441 | 0 | 3.09 |
| HU | 5.99 | 0 | 55.75 | 0.484 | 0 | 20.29 | ||
| Зеленый Город | BR | 8.712 | 5.838 | –0.0678 | 0.339 | 0.1 × 10–3 | 17.92 | |
| HU | 16.34 | –0.2402 | 0.07426 | 0.508 | 0 | |||
| $y = {{b}_{0}} + {{b}_{1}}{{x}_{1}} + {{b}_{2}}x_{1}^{2}$ | Щелоковский Хутор |
WD | 642.1 | –114.7 | 5.142 | 0.876 | 0 | 2.21 |
| $y = {{b}_{0}} + {{b}_{1}}{{x}_{{2{\text{ }}}}} + {{b}_{2}}x_{2}^{2}$ | Жигули | ML | 44.08 | –0.3611 | 0.00091 | 0.489 | 0 | 17.75 |
| Щелоковский Хутор |
BS | –34.93 | 0.9472 | 0.00403 | 0.591 | 0 | 90.49 | |
| HU | 12.5 | 0.1712 | 0 | 0.449 | 0 | 9.31 | ||
| Керженец | WD | 22.85 | –0.2588 | 0.00149 | 0.518 | 0 | 12.78 | |
| HU | 8.84 | 0.0091 | 0.0006 | 0.724 | 0 | 8.03 | ||
| ПТЗ | BS | 244.6 | 0.3956 | –0.0031 | 0.310 | 0.7 × 10–3 | 71.4 | |
| HU | 25.31 | 0.0159 | 0.00042 | 0.333 | 0.4 × 10–3 | 16.32 | ||
| y = exp(b0 + b1/x1) | Жигули | BS | 3.706 | 15.42 | – | 0.373 | 0 | 39.3 |
| BR | 1.768 | 26.26 | – | 0.377 | 0 | 12.25 | ||
| Зеленый Город |
ML | 0.0845 | 35.08 | – | 0.256 | 0 | 16.0 | |
| WD | –4.389 | 64.63 | – | 0.412 | 0 | 7.0 | ||
| Выкса | ML | 0.3274 | 46.81 | – | 0.637 | 0 | 0.34 | |
Корреляционные связи не всегда достаточно высоки (коэффициент детерминации меняется от 0.25–0.30 до 0.65–0.70), хотя и вполне значимы (критерий Пирсона Р = 1.0–2.5 × 10–4). При слабой связи последнюю можно интерпретировать как некоторую общую тенденцию изменений данного метаболического параметра под влиянием геофизического тренда на фоне “шумового” воздействия других факторов локального порядка (прежде всего, факторов литогенной основы ландшафтов).
Как известно, природные комплексы ландшафтного уровня отличаются сложной зарегулированной системой прямых и обратных экологических связей, которые имеют поэтому принципиально вероятностный характер (Арманд и др., 1969; Зейдис и др., 2001). При описании сложных многокомпонентных биологических систем могут нарушаться базовые принципы статистического анализа. В частности, может не выполняться требование линейной независимости предикторов, а коэффициенты корреляции и детерминации не могут быть высокими (Montgomery, Peck, 1982). Диссимметризация и дробление гидротермических ниш, с преимущественно пуассоновским характером распределения биогеоценотических единиц, указывают на сложный, дифференцированный в пространстве процесс их климатогенных преобразований (Коломыц, 2018). Далее, “… численные предсказания прогнозных моделей … должны рассматриваться …скорее как информация к размышлению о вероятных будущих траекториях лесов …, принимая во внимание значительную неопределенность прогнозов” (Швиденко, 2012, с. 55).
Основная задача ординационного анализа состояла в том, чтобы отделить изучаемый сигнал от “шума” и оценить количественно величину этого сигнала. Данный сигнал нередко оказывается гораздо менее мощным, нежели множество “шумовых” воздействий, однако важно то, что он постоянно действующий и однонаправленный, и в этом его эффективность. Локальный разброс метаболических характеристик, обусловленный информационным “шумом”, в значительной степени элиминировался при зонально-региональном обобщении результатов топологической ординации (см. ниже).
Для получения базовых и прогнозных углеродных характеристик лесных формаций был осуществлен переход с локального уровня операционных единиц на региональный с помощью разработанного нами метода индукционно-иерархической экстраполяции, подробно описанного в работе (Коломыц и др., 2009). Метод основан на эмпирически установленном явлении полизональности локальных экосистем как формы их реакции на глобальные изменения климата (Коломыц, 2008).
Каждый тип/подтип растительной формации, представленный на мелкомасштабной геоботанической карте, идентифицировался определенной группой конформных фитоценозов, взятых из факторально-динамических рядов биогеоценозов по всем полигонам. Пример такой идентификации приведен в табл. 2. В свою очередь, каждый экорегион, описываемый определенным экспериментальным полигоном, содержал целый спектр локальных представителей различных зонально-региональных фитоценологических систем ранга растительных формаций.
Одновременно создавалась карта мезоместоположений, для чего была проведена экологизация ландшафтной карты (Коломыц, 2005). Каждый ареал данной формации, представленный как поливекторное множество мезокатен, дробился на региональные типы местоположений – от элювиального и трансэлювиального до аккумулятивного и супераквального (Глазовская, 1964). Принимая далее мезокатену в качестве гомоморфного образа микрокатены, согласно концепции ландшафтных сопряжений Б.Б. Полынова (1952), разносили имеющиеся биогеоценозы всех выделенных групп из каждого экспериментального полигона по мезоместоположениям соответствующего ему геоботанического ареала (табл. 3). Полученная таким образом региональная фитокатенарная мозаика насыщалась затем метаболическими параметрами биогеоценозов, взятых уже в качестве локальных представителей соответствующих зонально-региональных типов/подтипов географической среды. Это были базовое содержание углерода и углеродный баланс к данному прогнозному сроку в отдельных пулах и в целом по лесному биогеоценозу. Наконец, на основе информационного анализа по каждой группе растительных формаций бассейна рассчитывались средневзвешенные значения ΣС(баз) и ∆С(Fa) в соответствии с площадной долей ее участия в данном зональном типе формаций.
Таблица 3.
Подтаежный елово-широколиственный биом (полигон № 4 “Зеленый город”, см. рис. 1). Пример индукционно-иерархической экстраполяции групп лесных фитоценозов на зонально-региональные группы растительных формаций
| № пробной площади | Экспозиция | Тип местоположения* | Зональная группа фитоценоза |
Группа растительных формаций, коррелятивная растительному сообществу данного биогеоценоза** |
|---|---|---|---|---|
| 33 | Горизонтальная | ТА | Бореальная таежная |
Североевропейские среднетаежные еловые кустарничково-зеленомошные леса |
| 1 | С | Э | Бореальная таежная |
Североевропейские южнотаежные еловые леса кустарничково-травяные с участием неморальных элементов |
| 39 | ЮЮВ | Т | Неморально-бореальная | Предуральские южнотаежные пихтово-еловые леса, неморальнотравяно-кисличные с участием липы в подлеске |
| 20 | ЮВ | ТЭ | Бореально-неморальная | Подтаежные широколиственно-еловые неморальнотравяные леса |
| 47 | Горизонтальная | ТЭ | Бореальная боровая |
Североевропейские средне- и южнотаежные сосновые (с примесью ели и березы) зеленомошные и кустарничковые леса |
| 11 | ЮЗ | Т | Неморально-бореальная | Североевропейские подтаежные широколиственно-сосновые леса с елью, лещиновые, разнотравно-злаковые |
| 19 | Горизонтальная | Э | Неморальная европейская |
Среднерусско-приволжские северные (с примесью ели) дубовые и липово-дубовые неморальнотравяные леса |
| 14 | Горизонтальная | Э | Неморальная евразиатская |
Уральские горные и предгорные кленово-липово-дубовые злаково-высокотравно-папоротниковые леса |
| 8 | Горизонтальная | ТА | Неморальная евроазиатская | Среднерусско-приволжские липовые,
с примесью дуба, клена, ильма, злаково-разнотравные леса |
В каждую группу формаций входят лесные сообщества разного возраста (от молодого до перестойного), что создает определенное многообразие содержания углерода в различных пулах. Считается, что наиболее высокие значения депонирования СО2 свойственны молодым и средневозрастным насаждениям (Замолодчиков и др., 2005; Разнообразие и динамика …, 2012 и др.). Однако влияние фактора возраста леса удалось обойти. В процессе обобщения по каждой лесной формации, с одной стороны, интегрировались локальные геоморфологические и гидротермоэдафические условия ее формирования, а с другой, – осреднялись возрастные показатели лесообразующих пород в каждом множестве пробных площадей (табл. 3), что “стирало” соответствующий разброс продуктивности и позволяло получить некоторые средние для каждой группы формаций значения как базового содержания углерода в различных пулах, так и ее прогнозируемого углеродного баланса. Таким образом, каждая лесная формация описывалась осредненными зонально-региональными ресурсами регуляции углеродного цикла. Эти ресурсы складываются из топологического разнообразия абиотической среды и уровней функционирования биогеоценозов, входящих в данную формацию.
Оценка потенциальных возможностей биоклиматической системы Волжского бассейна в регуляции углеродного цикла для различных сценариев глобальных изменений климата (см. ниже) проведена по углеродным балансам лесных формаций (табл. 1) трех иерархических уровней: высшего (зональных типов и классов формаций), среднего (подзональных подтипов) и низшего (групп формаций). По каждому иерархическому уровню вся совокупность углеродных балансов фитоценологических единиц была представлена в виде единой экологической ниши их функционирования, включающей как положительные, так и отрицательные значения параметра ∆С(Fa). Назовем ее балансовой экологической нишей. По удельным углеродным балансам формационных единиц данного уровня и занимаемых ими площадям были рассчитаны парциальные суммарные балансы ∑∆С(Fa), а затем – доля каждой формации в интегральном суммарном балансе [∑∑∆С(Fa)]. В итоге балансовая экологическая ниша лесного покрова Волжского бассейна для каждого прогнозного климатического сценария была представлены в виде распределения параметра ∑∆С(Fa), выраженного в долях от ∑∑∆С(Fa). Распределение прогнозируемого баланса углерода получено для низшего фитоценологического таксона (рис. 2) и для двух высших таксонов (рис. 3).
Рис. 2.
Распределение долей суммарного углеродного баланса y групп растительных формаций Волжского бассейна x (нижний иерархический уровень) по модели E GISS: (а) – на 2200 г. (термоаридный тренд); (б) – на 2050 г. (холодно-гумидный тренд).

Рис. 3.
Распределение долей прогнозного баланса углерода в зональных типах и классах коренных растительных формаций Волжского бассейна (верхний и средний иерархические уровни) по модели E GISS: (а) – на 2200 г. (термоаридный тренд); (б) – на 2050 г. (холодно-гумидный тренд).
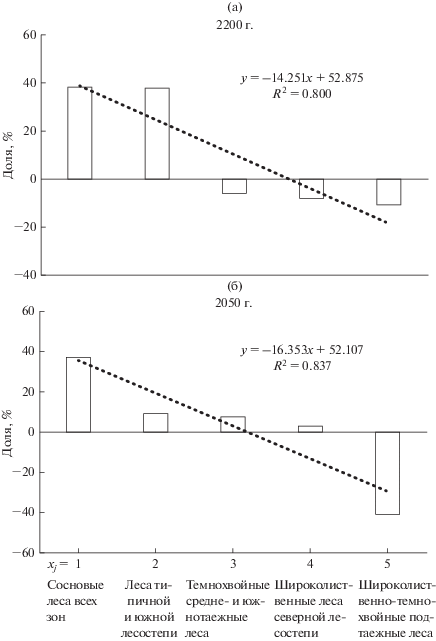
Если расположить ГЛФ по нише в порядке убывания параметра ∑∆С(Fa), то их распределение описывается закономерностью, выраженной арифметической прогрессией с коэффициентом детерминации выше 0.84 (рис. 2). На графиках можно видеть доминирующие формационные единицы, формации-субдоминанты с положительной и отрицательной регуляцией углеродного цикла.
Ряд авторов, в том числе Р. Уиттекер (1980) при анализе распределения видового богатства различных жизненных форм растений описали явление захвата экологической ниши. Захват описывался геометрической прогрессией, определяющей закономерности распределения долей жизненных форм в системе, объем занимаемых ниш доминантами и субдоминантами (Шарый и др., 2019). Рассмотрим подобным образом нишу углеродного баланса. Наличие дискретных компонентов с разными знаками, отвечающих процессам депонирования и эмиссии, не позволяет использовать геометрическую прогрессию для описания ниши. Однако, если принять весь объем ниши, состоящей из двух частей, – депонирующей (положительные значения) и эмиссионной (отрицательные значения) за 100%, можно провести расчет долей для каждой ГЛФ в процентах без учета знака баланса. Распределение балансов можно описать арифметической прогрессией, отвечающей закону линейного убывания дискретных компонентов. Захват ниши в этом случае состоит в следующем. Если доминирующая единица растительного покрова занимает долю k от объема ниши, то следующая по рангу единица занимает такую же долю от оставшейся части ниши (1 – k), следующая – от (1 – 2k) и т.д. Это соотношение описывается уравнением:
(3)
${{p}_{i}} = {{p}_{1}}--[{{(i--1)} \mathord{\left/ {\vphantom {{(i--1)} {(n--1)}}} \right. \kern-0em} {(n--1)}}]({{p}_{1}}--{{p}_{n}}),~$Степень доминирования ГЛФ оценивается долей k относительного объема ее балансовой ниши. Эта доля пропорциональна значениям y1 или |yn| в зависимости от того, какое из них больше по абсолютной величине. Показатель доминирования d может быть также оценен по углу наклона тренда балансовой ниши, то есть как величина, пропорциональная |а| в тренде. Более строгое определение k и d для арифметической прогрессии пока не найдено. Однако сравнение распределений, например, для разных климатических сценариев можно проводить с помощью описанных оценок.
Показатель k, рассчитанный с помощью d, характеризует спектр занимаемых лесными формациями объемов регуляции углеродного цикла и их пропорции: чем больше значение k, тем сильнее доминирование, и – наоборот. Таким образом, по значению параметра k дается оценка доминирования рангов в системе, а значение коэффициента детерминации линейного тренда отвечает согласованности дискретных компонентов: чем он больше, тем более согласованы в общей системе балансовой ниши компоненты положительной и отрицательной регуляции углеродного цикла.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Прогнозные климатические сценарии
Количественная оценка экологических ресурсов биоклиматической системы Волжского бассейна проведена по прогнозным климатическим сценариям двух глобальных моделей из семейства моделей общей циркуляции атмосферы AOGCMs: 1) модели E GISS (Hansen et al., 2007) и 2) модели HadCM3, версия А-2 (Pope et al., 2000). Первая модель дает пределы климатических изменений, соответствующие целям Парижского (2015) Соглашения – не допустить среднегодовое потепление более чем на 1.5–2.0°С к середине нашего столетия, то есть к сроку удвоения концентрации СО2 в атмосфере. В бореальном поясе Волжского бассейна к 2050 г. ожидается понижение средней июльской температуры (tиюля) на 0.5–1.9°С, с ростом коэффициента атмосферного увлажнения Высоцкого–Иванова (Кувл) с 1.15–1.52 до 1.27–1.79 (холодно-гумидный сценарий), а в 2200 г. – ее повышение на 0.5–0.7°С при уменьшении Кувл до 1.00–1.36 (умеренный термоаридный сценарий). Оба прогнозных срока нами рассматривались безотносительно к их временнóй привязке, то есть они использованы как модели для ответа на вопрос “что будет, если …”. Аналогичный подход принят, как известно, в ретроспективном прогнозе по палеогеографическим данным.
По экстремальной модели HadCM3 повышение tиюля в Средневолжском регионе к 2050 г. составит 2–3°С, а в конце XXI в. достигнет 5.5–7.0°С. Годовое количество осадков при этом изменится незначительно, поэтому в лесостепных районах вековое падение Кувл составит с 0.95–1.07 до 0.36–0.45, а в подтаежных – с 1.37 до 0.65. По данному термоаридному сценарию весь юг бореального пояса на Русской равнине окажется в суббореальных климатических условиях (Волобуев, 1963). Они уже могут превысить порог выносливости основных лесообразующих пород и вызвать распад на обширных пространствах не только неморальных, но и бореальных лесов. Следует отметить, что климатический прогноз по хэдловской модели адекватен существующему в настоящее время тренду потепления, который способен привести к повышению средней глобальной температуры к 2100 г. на 4°С (Le Quere et al., 2015), а на территории России – на 6–11°С (Швиденко и др., 2017).
Экологические ресурсы лесных формаций при умеренных климатических колебаниях
Базовое суммарное содержание углерода, а также удельные и суммарные углеродные балансы восстановленных коренных лесов в меняющемся климате (табл. 4; рис. 3 и 4) раскрывают потенциальную регуляцию углеродного цикла зонально-региональной структурой биоклиматической системы Волжского бассейна для трех описанных климатических сценариев.
Таблица 4.
Суммарные базовые запасы углерода, суммарный и удельный углеродный баланс в восстановленных коренных (зонально-климаксных) лесах Волжского бассейна (см. рис. 1) для различных климатических сценариев, согласно глобальным климатическим моделям E GISS и HadCM3
| Группы формаций (см. табл. 1) |
Общая площадь, кв. км |
Суммарные базовые запасы углерода, млн т |
Суммарный (удельный) углеродный баланс, в млн т (т/га), по климатическим сценариям |
||
|---|---|---|---|---|---|
| модель E GISS | модель HadCM3, экстремальному термоаридному, 2100 г. |
||||
| холодно- гумидному, 2050 г. |
умеренному термоаридному, 2200 г. | ||||
1  |
44 167 | 508.596 | –70.932 [+] (–16.06) |
+187.489 [+] (+42.45) |
+91.205 [+] (+20.65) |
2  |
123 429 | 2307.238 | –122.565 [+] (–9.93) |
–51.100 [–] (–4.14) |
+2.222 [+] (+0.18) |
3  |
92 997 | 1975.984 | +36.362 [–] (+3.91) |
+137.822 [+] (+14.82) |
+103.785 [+] (+11.16) |
4  |
68 015 | 579.350 | –64.274 [+] (–9.45) |
–26.800 [–] (–3.94) |
+ 221.865 [+] (+32.62) |
5, 6  |
25 896 | 1663.224 | +9.426 [–] (+3.64) |
+35.685 [+] (+13.78) |
+40.372 [+] (+15.59) |
7  |
89 595 | 2445.425 | +111.725 [–] (+12.47) |
+189.404 [+] (+21.14) |
+147.563 [+] (+16.47) |
8, 9  |
63 479 | 1122.653 | +117.881 [–] (+18.57) |
+173.537 [+] (+27.33) |
+55.448 [+] +(9.05) |
11  |
132 245 | 2486.206 | +18.118 [–] (+1.37) |
–102.622 [–] (–7.76) |
+110.689 [+] +(8.37) |
10, 12  |
39 344 | 703.510 | +58.859 [–] (+14.96) |
–56.891 [–] (–14.46) |
– 13.613 [–] –(3.46) |
13  |
3181 | 4.372 | +4.165 [–] (+13.09) |
+2.416 [+] (+7.60) |
+0.767 [+] +(2.41) |
| Заболоченные леса | 5026 | 70.721 | +8.463 [–] (+16.84) |
+7.172 [+] (+14.27) |
+20.370 [+] +(40.53) |
| Лесные болота | 2062 | 283.649 | +2.142 [–] (+10.39) |
+5.798 [+] (+28.12) |
+4.182 [+] +(20.28) |
| Неморальные поймы | 14 915 | 158.680 | +0.223 [–] (+0.15) |
+11.798 [+] (+7.91) |
+12.469 [+] +(8.36) |
| Сумма (среднее) |
704 351 | 14 309.61 | +109.586 [–] (+4.61) |
+513.708 [+] (+11.31) |
+797.319 [+] (+14.02) |
Рис. 4.
Распределение удельного углеродного баланса в восстановленных коренных лесных формациях Волжского бассейна для сценария умеренного регионального потепления (2200 г.), согласно модели E GISS. Углеродный баланс (т/га): 1 – (–15.0... –8.0); 2 – (–8.0... 0); 3 – 0–15.0; 4 – 15.0–25.0; 5 – 25.0–43.0; 6 – озера, водохранилища и территории с отсутствием лесной растительности.
Анализ балансовой ниши по холодно-гумидному сценарию на 2050 г. показывает, что распределение углерода в низшем таксоне лесных формаций характеризуется заметно большей согласованностью, чем в зональных типах и классах формаций (рис. 2б и 3б). Депонирующая часть балансовой ниши несколько преобладает над эмиccионной и составляет 57.8% от общего объема. Доминантом депонирующей части являются сосновые подтаежные леса (их доля составляет 19.0%), субдоминантом (18.0%) – средне- и южнотаежные сосновые леса. Доминант и субдоминант эмиccионной части баланса – южнотаежные и среднетаежные темнохвойные леса, с их долей соответственно 19.7 и 11.4%.
Для высших фитоценологических таксонов в депонирующей части баланса место доминанта заняли сосновые и широколиственно-сосновые леса (37.2% от объема ниши), в эмиccионной части – средне- и южнотаежные темнохвойные леса (41.5%). В условиях холодно-гумидного климатического тренда доминантная в бассейне по площади и базовым запасам углерода формация темнохвойных лесов в бассейне “раскололась” на две части, противоположные по типу регуляции углеродного цикла.
Балансовая ниша 2200 г., сформированная по условиям мягкого термоаридного тренда, также характеризуется большей согласованностью прогнозируемого углерода для групп лесных формаций по сравнению с их зональными типами и классами (рис. 2а и 3а). По этому климатическому сценарию депонирующая часть балансовой ниши значительно превышает эмиссионную (доля 75.3% от всей ниши против 24.7%). В первом случае на низшем иерархическом уровне доминируют верхневолжские средне- и южнотаежные сосновые леса. Субдоминанты депонирования – восточноевропейские среднетаежные темнохвойно-лесные. Доминантом же эмиccионной части являются широколиственные леса. По всем фитоценологическим таксонам при термоаридном тренде взаимозависимость между компонентами углеродного баланса выражена слабее, чем при тренде холодно-гумидном.
В целом коренные бореальные и суббореальные леса бассейна могут оказывать два взаимно противоположных воздействия на углеродный обмен земной поверхности с атмосферой: 1) сравнительно слабое негативное – при холодно-гумидном тренде (дополнительный депозит СО2 и соответствующее усиление похолодания); 2) гораздо более мощное позитивное – при тренде термоаридном, с трехкратно возрастающим значением ∆С(Fa), которое снижает концентрацию парниковых газов в атмосфере и тем самым ослабляет антропогенное потепление. В первом случае общая адсорбция СО2 из атмосферы составляет около 1% суммарного базового содержания углерода в бореальных и суббореальных лесах, а во втором – 3.5%. Для сравнения отметим, что для хвойных, смешанных и других лесов США по расчетам, основанным на климатической модели UKMO-1987 (предшественнице модели HadCM3) получено, что удвоение содержания СО2 в атмосфере приводит к изменениям содержания углерода в диапазоне от –(1.5–1.8) до +(7.8–12.5)% (Aber et al., 2001).
По сценарию умеренного потепления (на 2200 г., рис. 2а) депонирующая часть углеродно-балансовой ниши лесов Волжского бассейна значительно превышает эмиccионную (доля 75.3% от всей ниши, против 24.7%). Наиболее значительное сокращение парниковых газов в атмосфере и, следовательно, максимальное ослабление регионального потепления ожидается со стороны двух доминирующих групп коренных лесных формаций, занимающих заметную часть площади бассейна (около 20%) – средне- и южнотаежных сосняков в бассейнах Унжи, Ветлуги и Средней Вятки по Низменному Заволжью и среднетаежных пихто-ельников на северных водоразделах Среднего Поволжья (рис. 3). В каждой из них суммарный углеродный баланс ∑∆С(Fa) приближается к +190 млн т, что составляет 19.5–19.7% от всего объема балансовой ниши. Почти столь же высоким экологическим ресурсом обладают подтаежные елово- и сосново-широколиственные леса в западном секторе бассейна, где [∑∆С(Fa)] ≈ ≈ +140–170 млн т). В восточном же секторе (Камско-Печерско-Западноуральском) суммарный углеродный баланс снижается почти в четыре раза (табл. 4; рис. 3).
Разительным контрастом по отношению к упомянутым лесным формациям при термоаридном тренде выступают восточноевропейские широколиственные леса, которые как коренные сообщества должны занимать значительную часть бассейна (19.5%). Они должны оказывать существенное негативное воздействие на углеродный обмен земной поверхности с атмосферой: здесь [∑∆С(Fa)] > –100 млн т. В два с лишним раза возрастет отрицательный баланс в маргинальных типично-лесостепных сосновых и широколиственно-сосновых лесах (рис. 2а). Последние, таким образом, оказываются не только наиболее неустойчивыми к глобальному потеплению (Коломыц, 2008), но и как фитоценологические системы с положительной обратной связью сами способствуют своей деградации, усиливая данный климатический тренд.
Интересными оказались отрицательные значения суммарного углеродного баланса у всех южнотаежных пихто-ельников. Причины этого феномена остаются неясными, и здесь можно высказать следующее. Полог темнохвойных лесов оказывает наиболее мощное эдификаторное воздействие на радиационный и тепловой балансы поверхности почвы и приземного слоя воздуха (Морозов, 1926). На открытых площадках метеостанций эти балансы определяют распределение зональных типов и подтипов растительного покрова и почв (Волобуев, 1963; Базилевич и др., 1986). Простой расчет показывает (Коломыц, 2003), что если бы не было снижения под пологом южнотаежных пихто-ельников годовой суммарной радиации и коэффициента увлажнения на 10–15% по сравнению, например, с сосняками того же возраста (Работнов, 1978), то на месте темнохвойных формаций здесь должны были бы произрастать широколиственно-сосновые либо чисто широколиственные леса. Находясь по-существу в “не своей” фоновой климатической зоне, южная тайга реагирует на слабый климатический сигнал (по сценарию E GISS на 2200 г.) с отрицательным углеродным балансом. Лишь при гораздо более сильном потеплении (по эстремальной модели HadCM3) их баланс становится хотя и незначительно, но положительным (табл. 4).
Для сценария холодно-гумидного тренда имеют место аналогичные соотношения абсолютных значений [∑∆С(Fa)] между группами коренных лесных формаций, однако знак их углеродного баланса меняется на противоположный (рис. 2 б): положительный углеродный баланс коренных лесов способствует дополнительному похолоданию. Данный сценарий свойствен всему левобережью Средней Волги, занятому средне- и южнотаежными и подтаежными лесами. Наибольшее смягчение похолодания следует ожидать со стороны средне- и южнотаежных лесных формаций Ветлужско-Камcкого региона (∑∆С(Fa) ≈ –(128–220) т/га). Леса же северной и средней лесостепи должны оказывать максимальное усиление холодного климатического тренда (∑∆С(Fa) ≈ +(74–243) т/га).
В целом группы коренных лесов Волжского бассейна, при заполнении каждой из них своего зонально-регионального экологического пространства, должны оказывать общее положительное регулирующее влияние на углеродный обмен земной поверхности с атмосферой при потеплении и отрицательное влияние – при похолодании, причем в первом случае в три раза более мощное, нежели во втором (табл. 4, нижняя строка). Обладая достаточно высокими экологическими ресурсами, коренные бореальные леса восточноевропейского субконтинента могут служить эффективным поглотителем парниковых газов из атмосферы, снижая тем самым грядущее глобальное потепление.
Ресурсный потенциал лесных формаций в условиях экстремального потепления
Не менее важно оценить регуляцию углеродного цикла лесными экосистемами при экстремальном термоаридом сигнале, согласно модели HadCM3, климатические сценарии которой могут оказаться, как уже говорилось, вполне реальными при сохранении текущей тенденции глобального потепления. По сравнению с е-гиссовским термоаридным сценарием, хэдловская модель дает почти трехкратную прибавку не только летних, но и зимних температур. Соответственно должна возрасти продолжительность вегетационного периода, что даже при небольшом увеличении годовых атмосферных осадков неизбежно вызовет рост продуктивности лесных сообществ, с соответствующим усилением ими поглощения парниковых газов.
В результате при экстремальном потеплении коренной лесной покров Волжского бассейна может приобрести преимущественно положительный углеродный баланс (табл. 4, рис. 5). Так, восточноевропейские подтаежные широколиственно-еловые леса, оказывающие по е-гиссовскому сценарию потепления слабо выраженную отрицательную регуляцию углеродного цикла (∆С(Fa) = –7.12 т/га), при более мощном термоаридном сигнале, согласно модели HadCM3, становятся субъектами существенной положительной регуляции (∆С(Fa) = +20.01 т/га). Аналогично меняется углеродный баланс камско-печерско-предуральских коренных подтаежных лесов – с –18.24 до +15.59 т/га. Восточноевропейские широколиственные леса оказываются уже не источником углерода, как это было при слабом термоаридном сигнале, а его стоком. Совместно с бореальными (подтаежными и таежными) лесами они способны выполнять общую положительную регуляцию углеродного цикла. В целом можно констатировать общее смягчающее воздействие лесного покрова Волжского бассейна на климатические изменения по мере усиления самого гидротермического сигнала.
Итак, регуляция углеродного цикла со стороны коренных бореальных и неморальных лесов Русской равнины должна быть достаточно пестрой не только по знаку, но и по абсолютной величине. Такая пестрота между группами лесных формаций будет проявляться в пределах каждого типа (подтипа) растительности, поэтому итоговый эффект регуляции углеродного цикла для всей рассматриваемой территории Волжского бассейна будет существенно зависеть от соотношения площадей ГЛФ, то есть от структуры самой биоклиматической системы.
Высокие удельные средневзвешенные значения углеродного баланса той или иной ГЛФ нередко не соответствуют занимаемой им площади. Так, сосновые средне- и южнотаежные леса имеют не столь значительные площади, однако благодаря высоким положительным значениям ∆С(Fa) их итоговый вклад в региональный сток углерода окажется весьма существенным. При экстремальном термоаридном тренде их суммарный углеродный баланс будет равен +51.725 млн т, что составит около 38% общего положительного баланса.
Незначительной будет негативная регуляция углеродного цикла со стороны наиболее распространенных (занимающих более 30% площади региона) темнохвойных южнотаежных и смешанных лесов – по причине низких отрицательных значений ∆С(Fa). И наоборот, менее распространенные широколиственно-сосновые подтаежные леса станут объектами наиболее мощного стока СО2 при термоаридном климатическом тренде (в сумме около 50.0 млн т), что и предопределит их доминирующую позитивную роль в региональной биотической регуляции углеродного цикла.
Виртуальные прогнозно-экологические сценарии углеродного баланса
В связи с ожидаемыми изменениями лесного покрова из-за потепления климата проводятся экономические оценки осуществления лесопосадок в масштабе планеты для снижения СО2 в атмосфере. При осуществлении новых лесопосадок сокращение стоимости связывания углерода для уменьшения СО2 в атмосфере может составить 5–25% (Benitez et al., 2007). Даются также оценки масс депонированного углерода на планете в зависимости от планируемых затрат и времени. Однако нерешенными остаются вопросы о том, какие и где лесообразующие породы или растительные ассоциации эффективнее использовать.
Очевидно, при подборе ассортимента лесопосадочных пород предпочтение следует отдавать тем из них, которые будут обеспечивать в лесном насаждении положительный углеродный баланс и смогут создавать, таким образом, лесные сообщества, наиболее устойчивые к изменениям климата. Тем самым можно достигнуть наиболее высокого уровня “… потенциала смягчения изменений климата средствами лесного хозяйства” (Швиденко и др., 2017, с. 15). При этом “более перспективно … выравнивание возрастной структуры насаждений в лесном фонде” (Замолодчиков и др., 2005, с. 328).
Для глобального потепления на период 2075–2100 гг. по экстремальной модели HadCM3 нами был проведен опыт построения регрессионных карт углеродного баланса лесных формаций Окского бассейна (рис. 6). Карты созданы на основе выявленных статистических связей параметра ∆С(Fa)2100 различных лесных формаций бассейна с наиболее значимыми предикторами (табл. 5). Были использованы картографически привязанные данные по балансу углерода к 2075–2100 гг. с учетом типов мезоместоположений, полученные в ходе описанных выше исследований. Далее отдельно для каждой формации рассчитывали карту баланса для всей площади бассейна, включив в регрессионный анализ матрицы рельефа и прогнозную температуру июля.
Рис. 5.
Распределение удельного углеродного баланса в восстановленных коренных лесных формациях Волжского бассейна для сценария экстремального регионального потепления (2100 г.), согласно модели HadCM3. Углеродный баланс (т/га): 1 – 32.5–28.5; 2 – 28.5–17.5; 3 – 17.5–5.5; 4 – 5.5–1.0; 5 – 1.0... –3.5; 6 – озера, водохранилища и территории с отсутствием лесной растительности.

Таблица 5.
Уравнения связей изменения суммарного содержания углерода в лесных формациях Окского бассейна к 2100 г. (∆С2150), по модели HadCM3, с наиболее значимыми факторами среды
| Зональные типы и классы растительных формаций | Уравнения регрессии |
|---|---|
| Еловые и широколиственно-еловые леса | ∆С2150 г. = 76.7Z + 46.7T + 24.0F – 63.8 |
| Сосновые и широколиственно-сосновые леса | ∆С2150 г. = –24.0Z + 21.0MCA – 7.1F + 12.1 |
| Широколиственные леса | ∆С2150 г. = –76.2T + 42.9MCA + 12.1GA + 15.1 |
| Коренные и вторичные леса бассейна (все леса бассейна в их современных ареалах) | ∆С2150 г. = 28.8MCA + 23.9kmax – 7.2T – 7.8 |
Сами карты носят условный характер. Каждая из них описывает виртуальную реальность – картину изменений содержания углерода в данной группе формаций при допущении, что она покрывает всю площадь бассейна. Однако такие карты позволяют более четко представить парциальный вклад лесных сообществ в биотическую регуляцию углеродного цикла на территориях с различными природными условиями. Это особенно важно для стратегии лесоразведения в ныне обезлесенных районах при ожидаемых изменениях климата, а также для лесовосстановления после пожаров и вырубок. На территории Российской Федерации площадь фонда лесоразведения составляет от 10.8 до 20.3 млн га, между тем с учетом лесовосстановления она может достичь 35–40 млн га (Исаев, Коровин, 2006).
Мозаика депонирования и эмиссии углерода в лесных формациях при опробованных сценариях посадки лесов заметно различается. В восточной половине территории Окского бассейна целесообразно высаживать сосну в смеси с широколиственными породами, а в крайней северо-западной полосе – чистые липово-дубовые древостои (рис. 6б, 6в); они дадут максимум поглощения парниковых газов. В то же время при посадках широколиственных пород на юго-востоке бассейна, где прогнозируемая температура имеет наибольшие значения, ожидается активная эмиссия СО2. Положительная, хотя и слабо выраженная, регуляция углеродного цикла проявится на всей территории бассейна при лесопосадках с сохранением современного площадного соотношения коренных и вторичных лесов. Это, по-видимому, самый “позитивный” сценарий, который характеризуется общим наибольшим депонированием углерода.
Рис. 6.
Регрессионные карты углеродного баланса в растительных формациях бассейна р. Ока на прогнозный период 2075–2100 гг. глобального потепления, согласно модели HadCM3. Допущение: территория бассейна р. Ока полностью покрыта лесами: (а) – еловыми и широколиственно-еловыми; (б) – сосновыми и широколиственно-сосновыми; (в) – широколиственными; (г) – коренными и вторичными в пропорциях их современного распределения. Изменения суммарного углерода, т/га: 1 – (–136... 0); 2 – 0–15; 3 – 15–30; 4 – 30–45; 5 – 45–115.
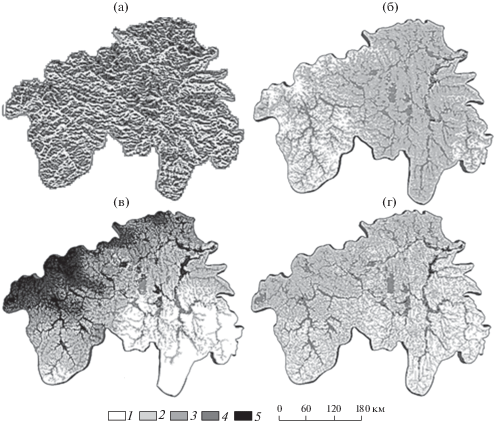
Для большинства лесных формаций (табл. 5) депозит углерода возрастает с увеличением площади водосбора МСА, то есть с переходом от элювиальных (Э) и трансэлювиальных (ТЭ) местоположений к трансаккумулятивным (ТА) и аккумулятивным (А). Особенно это характерно для сосновых и широколиственно-сосновых лесов Окско-Цнинской низменной равнины, где на обширных площадях дополнительное поглощение углерода достигает 15 т/га. В то же время в верховьях Оки на Среднерусской возвышенности положительные значения ∆C(Fa) свойственны только крупным речным долинам, а на местных водоразделах преобладает уже эмиссия СО2. В противоположность светлохвойным формациям, в еловых и широколиственно-еловых лесах депонирование углерода усиливается с абс. высотой Z, между тем как в речных долинах ельники дают его слабо выраженную эмиссию. При повышении освещенности склонов F (фактора, влияющего на локальную гидротермику) возрастают депозит углерода в еловых лесах и его эмиссия – в лесах сосновых и широколиственно-сосновых.
Крутизна склонов GA и максимальная кривизна kmax, определяющая “гребневые формы” в рельефе, оказывают положительное действие на аккумуляцию углерода. Среднеиюльская температура воздуха как один их основных климатических факторов влияет на углеродный баланс лесов по-разному. В относительно сложных по генезису группах лесных формаций повышение температуры вызывает тенденцию к преобладающей эмиссии углерода, а в сравнительно простых группах – к его депонированию.
Предикторы уравнений регрессии, описывающих пространственную вариабельность изменений суммарного углерода, могут действовать разнонаправленно, поэтому необходим результирующий учет их интерференции. В целом можно сказать, что на местном уровне более высокую эмиссию СО2, то есть отрицательную регуляцию углеродного цикла при потеплении, следует ожидать на приводораздельных местоположениях, особенно занятых хвойно-лесными сообществами. Широтно-зональные контрасты параметра ∆С(Fa) наиболее резко выражены в случае покрытия территории широколиственными лесами. В северо-западной полосе Окского бассейна, относящейся к подтаежной зоне, дубравы и липняки станут активно поглощать СО2 из атмосферы (∆С(Fa) достигает 50–100 т/га), ослабляя тем самым ее парниковый эффект. В юго-восточных же районах типичной и южной лесостепи широколиственные леса будут осуществлять, наоборот, отрицательную биотическую регуляцию углеродного цикла.
ЗАКЛЮЧЕНИЕ
Изложены представления о региональной биоклиматической системе лесного покрова и проведен опыт ее прогнозного пространственного анализа по коренной растительности как репрезентативному представителю этой системы.
По удельным и суммарным значениям углеродного баланса восстановленных коренных лесных формаций Волжского бассейна проведена количественная оценка эколого-ресурсного потенциала данной региональной биоклиматической системы в обеспечении положительной регуляции углеродного цикла и соответствующего смягчения прогнозируемого глобального потепления.
Балансовые экологические ниши лесного покрова Волжского бассейна для каждого прогнозного климатического сценария представлены в виде распределения долей суммарного углеродного баланса с закономерностью, выраженной арифметической прогрессией с заметными коэффициентами детерминации. На этих аналитических моделях выделены доминирующие формационные единицы, формации-субдоминанты и второстепенные формации с положительной и отрицательной регуляцией углеродного цикла. Растровые карты удельных углеродных балансов лесных формаций характеризуют региональные структуры депонирования и эмиссии парниковых газов для прогнозируемых сценариев потепления и похолодания.
Установлено, что при одном и том же климатическом сигнале различные лесные формации Волжского бассейна будут как поглощать дополнительно парниковые газы (положительный углеродный баланс лесных сообществ), так и выбрасывать их в атмосферу (отрицательный баланс), оказывая соответствующее влияние на состояние наземных экосистем. В каждом экорегионе выявляются определенные зонально-региональные типы регуляции коренным лесным покровом содержания СО2 в атмосфере при развитии климатической термоаридизации.
На примере Окского бассейна созданы прогнозно-экологические карты, которые описывают виртуальную реальность – картину изменений содержания углерода в данном зональном типе и классе формаций при допущении, что она покрывает всю площадь бассейна. Оценка парциального вклада коренных лесных формаций в биотическую регуляцию углеродного цикла особенно важна для стратегии лесоразведения в ныне обезлесенных районах при ожидаемых изменениях климата, а также для лесовосстановления после пожаров и вырубок.
Полученные обобщающие материалы по углеродным балансам крупного региона Европейской России могут быть использованы для количественных оценок потенциальных возможностей влияния бореальных и неморальных лесов на потоки парниковых газов в системе земная поверхность–атмосфера. Результаты проведенных исследований раскрывают некоторые механизмы регуляции углеродного цикла лесным покровом при климатических сценариях регионального потепления и похолодания и призваны внести определенный вклад в эмпирическое обоснование новой экологически ориентированной парадигмы В.Г. Горшкова и А.И. Уткина в учении о лесе.
Список литературы
Албриттон Д.Л., Баркер Т., Башмаков И. Изменение климата. 2001 г. Обобщенный доклад МГЭИК / Ред. Р.Т. Уотсон. Geneva: World Meteorological Organization, 2003. 220 p.
Арманд Д.Л., Преображенский В.С., Арманд А.Д. Природные комплексы и современные методы их изучения // Изв. АН СССР. Сер. геогр. 1969. № 5. С. 5–16.
Базилевич Н.И., Гребенщиков О.С., Тишков А.А. Географические закономерности структуры и функционирования экосистем. М.: Наука, 1986. 297 с.
Барталев С.А., Ершов Д.В., Исаев А.С. и др. Карта растительного покрова России. М.: ИКИ РАН и ЦЭПЛ РАН, 2010.
Волобуев В.Р. Экология почв (Очерки). Баку: Изд-во АН Азерб. ССР, 1963. 308 с.
Глазовская М.А. Геохимические основы типологии и методики исследований природных ландшафтов. М.: Изд-во МГУ, 1964. 230 с.
Глазовская М.А. Геохимия природных и антропогенных ландшафтов СССР. М.: Высшая школа, 1988. 328 с.
Горшков В.Г. Физические и биологические основы устойчивости жизни / Ред. К.С. Лосев. М.: б.и., 1995. 470 с.
Григорьев А.А. Развитие теоретических проблем советской теоретической географии. М.: Наука, 1965. 246 с.
Замолодчиков Д.Г., Грабовский В.И., Коровин Г.Н., Курц В.А. Динамика бюджета углерода лесов России за два последних десятилетия // Лесоведение. 2011. № 6. С. 16–28.
Замолодчиков Д.Г., Коровин Г.Н., Гитарский М.Л. Углеродный бюджет управляемых лесов Российской Федерации // Лесоведение. 2007. № 6. С. 23–34.
Замолодчиков Д.Г., Уткин А.И., Коровин Г.Н., Честных О.В. Динамика пулов и потоков углерода на территории лесного фонда России // Экология. 2005. № 5. С. 323–333.
Зейдис И.М., Кружалин В.И., Симонов Ю.Г. Общие свойства динамики геосистем // Вестн. МГУ. Сер. 5. География. 2001. № 4. С. 3–8.
Исаев А.С., Коровин Г.Н. Леса России и Киотский протокол // Возможности предотвращения изменений климата и его негативных последствий. Проблема Киотского протокола. М.: Наука, 2006. С. 287–305.
Исаченко Г.А., Книзе А.А., Романюк Б.Д. Экологические функции леса в южнотаежных ландшафтах Северо-Запада // Изв. ВГО. 2000. Вып. 1. С. 3–12.
Кобак К.И. Биотические компоненты углеродного цикла. Л.: Гидрометеоиздат, 1988. 248 с.
Коломыц Э.Г. Региональная модель глобальных изменений природной среды. М.: Наука, 2003. 371 с.
Коломыц Э.Г. Бореальный экотон и географическая зональность: Атлас-монография. М.: Наука, 2005. 390 с.
Коломыц Э.Г. Локальные механизмы глобальных изменений природных экосистем. М.: Наука, 2008. 427 с.
Коломыц Э.Г. Экспериментальная географическая экология. Записки географа-натуралиста. М.: КМК, 2018. 716 с.
Коломыц Э.Г., Розенберг Г.С., Шарая Л.С. Методы ландшафтной экологии в прогнозных оценках биотической регуляции углеродного цикла при глобальном потеплении // Экология. 2009. № 6. С. 1–8.
Кондратьев К.Я., Лосев К.С., Ананичева М.Д., Чеснокова И.В. Естественно-научные основы устойчивости жизни. М.: ВИНИТИ, Ин-т географии РАН, 2003. 239 с.
Лосев К.С. Экологические проблемы и перспективы устойчивого развития России в XXI веке. М.: Космосинформ, 2001. 399 с.
Моисеев Б.Н., Филипчук А.Н. Методика МГЭИК для расчета годичного депонирования углерода и оценка ее применимости для лесов России // Лесн. хозяйство. 2009. № 4. С. 11–13.
Морозов Г.Ф. Учение о лесе. Изд. третье. Л.: Гос. изд-во, 1926. 368 с.
Одум Ю. Основы экологии. Пер. с англ. М.: Мир, 1975. 740 с.
Полынов Б.Б. Географические работы. М.: Географгиз, 1952. 399 с.
Работнов Т.А. Фитоценология. М.: Изд-во МГУ, 1978. 384 с.
Разнообразие и динамика лесных экосистем России. Книга 1 / Ред. А.С. Исаев. М.: КМК, 2012. 460 с.
Растительность европейской части СССР / Ред. С.А. Грибова, Т.И. Исаченко, Е.М Лавренко. Л.: Наука, 1980. 429 с.
Растительность европейской части СССР и Кавказа [Карты]: для высших учебных заведений / Ред. Т.В. Котова. М.: ГУГК, 1987. 4 л.
Свирежев Ю.М. Математические модели в экологии // Число и мысль. М.: Знание, 1982. Вып. 5. С. 16–55.
Сочава В.Б. Растительный покров на тематических картах. Новосибирск: Наука, 1979. 189 с.
Сукачев В.Н. Избранные труды. Т. 1. Основы лесной типологии и биогеоценологии. Л.: Наука, 1972. 418 с.
Сукачев В.Н. Избранные труды. Том 3. Проблемы фитоценологии. Л.: Наука, 1975. 543 с.
Тишков А.А. Биосферные функции природных экосистем России. М.: Наука, 2005. 310 с.
Уиттекер Р. Сообщества и экосистемы. М.: Прогресс, 1980. 327 с.
Уткин А.И. Углеродный цикл и лесоводство // Лесоведение. 1995. № 5. С. 3–20.
Цельникер Ю.Л. Газообмен СО2 в лесных биогеоценозах / Идеи биогеоценологии в лесоведении и лесоразведении. М.: Наука, 2006. С. 213–229.
Шарый П.А., Шарая Л.С., Иванова А.В. и др. Сравнительный анализ видового богатства жизненных форм сосудистых растений в Среднем Поволжье // Сиб. экол. журн. 2019. № 5. С. 182–196.
Швиденко А.З. Глобальные изменения и российская лесная таксация // Лесн. таксац. и лесоустр. 2012. Т. 47. № 1. С. 52–75.
Швиденко Ф.З., Кракснер Ф., Оберштайер М., Щепаченко Д.Г. Проблема перехода к устойчивому управлению лесами России; потенциал и риски // Мат. Всерос. конф. с Междунар. участием “Лесные биогеоценозы бореальной зоны: география, структура, функции, динамика”, 16–19 сентября 2014 г., Красноярск. Новосибирск: Изд-во СО РАН, 2014. С. 14–19.
Швиденко А.З., Щепаченко Д.Г., Кракснер Ф., Онучин А.А. Переход к устойчивому управлению лесами России: теоретико-методические предпосылки // Сиб. лесн. журн. 2017. № 6. С. 3–25.
Aber J., Nelson R.P., Mcnulty St. et al. Forest processes and global environmental change: predicting the effects of individual and multiple stressors // BioScience. 2001. V. 51. № 9. P. 735–751.
Benitez P.C., McCallum I., Obersteiner M., Yamagata Y. The economics of tree-planting for carbon mitigation: a global assessment // Reg. Externat. Berlin, Heidelberg: Springer, 2007. P. 307–321.
Bryant L.R., Nielsen d., Tangley L. The last frontier forests. Washington, DC: World Resources Institute, 1997. 42 p.
Hansen J., Sato M., Ruedy R. Climate simulations for 1880–2003 with GISS model E // Climat. Dynam. 2007. V. 29. P. 661–696.
Le Quere C., Moriary R., Andrew R.M. Global carbon budget 2014 // Earth Syst. Sci. Data. 2015. V. 7. Iss. I. P. 47–85.
Montgomery D.C., Peck E.A. Introduction to linear regression analysis. New York: Wiley, 1982.
Paris Agreement // Conf. of the Parties. Twenty-first session. Paris, 30 Nov. to 11. Dec., 2015. 19 p.
Pope V.D., Gallani M.L., Rowntree P.R., Stratton R.A. The impact of new physical parametrizations in Hadley Centre climate model – HadCM3 // Climat. Dynam. 2000. V. 16. P. 123–146.
Shvidenko A., Schepaschenko D., McCallum I., Nilsson S. Can the uncertainty of full carbon accounting of forest ecosystems be made acceptable to policymakers? // Climat. Change. 2010. V. 103. P. 137–157.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии