

Успехи современной биологии, 2022, T. 142, № 3, стр. 213-222
Систематика наших дней: путь к новому синтезу или бесконечный тупик?
1 Санкт-Петербургский государственный университет
Санкт-Петербург, Россия
2 Санкт-Петербургский филиал Института истории естествознания и техники им. С.И. Вавилова РАН
Санкт-Петербург, Россия
* E-mail: radix.vinarski@gmail.com
Поступила в редакцию 09.01.2022
После доработки 16.02.2022
Принята к публикации 21.02.2022
- EDN: EZBUFQ
- DOI: 10.31857/S0042132422030097
Аннотация
Выражение “современный” (или “новый”) синтез давно укоренилось в лексиконе исследователей, пишущих об истории эволюционной биологии в ХХ в. Однако развитие биологической систематики за последние 100 лет обычно изображается как дарвиновская по духу борьба трех идеологически и методологически различных течений – нумерической систематики (фенетики), кладистики и эволюционной систематики. В то же время в теоретических рассуждениях и в нашей стране, и за рубежом признавалась необходимость и даже неизбежность будущего синтеза в области биологической систематики. В статье рассматривается вопрос о современном состоянии практической систематики в контексте проблемы возможного синтеза конкурирующих направлений. Показано, что фактически подобный синтез уже достигнут, но – в отличие от эволюционного синтеза середины прошлого века – не путем теоретических усилий группы энергичных авторов, а за счет естественной – и частично неосознанной – “конвергенции” путей развития соперничающих таксономических школ, стремившихся достичь максимальной операциональности и объективности получаемых ими результатов. Показано, что, несмотря на доминирование филогенетического подхода к построению системы, практическая систематика наших дней является в основе своей псевдокладистикой, т.к. в основу принятия таксономических решений кладутся статистические процедуры реконструкции филогении и выделения видов, а не анализ гомологий (синапоморфий). Эту ситуацию можно назвать “операциональным синтезом”, или конвенцией, сложившейся в первые десятилетия текущего столетия. С одной стороны, это можно рассматривать как безусловный успех, который, вероятно, позволит систематике окончательно избавиться от клейма “субъективной” и “чисто описательной” науки. С другой, – эта ситуация несет угрозу дегуманизации таксономической практики, передачи значительной части традиционных таксономических задач в ведение компьютеров.
“To hope for the new systematics is to imply no disrespect for the old”
ВВЕДЕНИЕ
Выражение “новый” (или “современный”) синтез давно и прочно укоренилось в лексиконе исследователей, пишущих об истории эволюционной биологии в ХХ в. В специальной литературе оно встречается уже с первой половины 1940-х гг. и было введено не историками, а самими действующими лицами научного движения, приведшего к созданию синтетической теории эволюции (СТЭ) (Huxley, 1942; The evolutionary…, 1980). Под “новым” эволюционным синтезом принято понимать неодарвинистскую эволюционную теорию, возникшую путем “объединения некоторых данных и концепций генетики, экологии, биогеографии, систематики, морфологии и палеонтологии с теорией естественного отбора” (Колчинский, 2015, с. 25). В соответствии с этим пониманием, историки науки ищут и находят свидетельства плодотворного сотрудничества специалистов в разных областях биологии, приведших к “новому синтезу” (см., например, Mayr, 1982; Smokovitis, 1992). Несмотря на наличие эволюционных теорий, альтернативных по отношению к СТЭ, последняя абсолютно доминировала на протяжении всей второй половины прошлого столетия, что создает впечатление практически полного единомыслия среди эволюционистов того времени.
Совсем иначе принято представлять историю развития биологической систематики за последние 50–80 лет. Если историки эволюционной теории склонны подчеркивать тенденцию к примирению сторонников противоположных позиций (Mayr, 1982), то историки систематики не жалеют красок для описания конфликтов и противоречий в этой области биологии (Расницын, 2002; Hull, 1970, 1988; Cracraft, 2000; Stuessy, 2013; Sterner, Lidgard, 2018), причем порой в ход идут такие метафоры, как “войны систематиков” (Felsenstein, 2001; Rieppel, 2008), которые, по мнению ряда авторов, наиболее точно описывают состояние дел в этой области в 1960–1980-е гг.
Известный сборник статей “Новая систематика”, вышедший в 1940 г. под редакцией Хаксли (Huxley, 1940), открывался его предисловием, в котором намечались пути к созданию новой синтетической таксономии, что должно было стать параллелью к закладывавшимся в те же самые годы концептуальным основам эволюционного синтеза (Галл, 2004; Vernon, 1993). Систематика, по мысли Хаксли, это центральная дисциплина наук о жизни, в основе которой должен лежать синтез биологических знаний при безусловном главенстве эволюционистского мышления. Основной задачей систематики как науки автор считал “detecting evolution at work” (Huxley, 1940, p. 2). О желательности и даже неизбежности синтеза в этой области писали многие авторы, как в нашей стране (Любищев, 1971; Боркин, 1989), так и за рубежом (Майр, 1971; Mayr, 1981; Hall, 1988). Однако этого не случилось, и развитие систематики пошло в совсем ином направлении.
Так, в интерпретации одного из наиболее влиятельных специалистов по данному вопросу, Дэвида Халла (Hull, 1988), во второй половине ХХ в. в биологической систематике соперничали три основных школы, или направления, то есть нумерическая систематика (фенетика), представленная Р. Сокалом, П. Снитом и их последователями, эволюционная систематика, развивавшаяся Э. Майром и Дж.Г. Симпсоном, а также филогенетическая систематика, или кладистика, начавшаяся с работ В. Хеннига и в 1970–1980-е гг. получившая большую популярность, особенно среди североамериканских зоологов-систематиков нового поколения. Взаимоотношения между этими школами часто изображают как проявление дарвиновской по духу борьбы за существование (в этом суть “эволюционного” подхода к истории науки, сторонником которого был Халл; см. Hull, 1988), а позиции сторон – как полностью взаимоисключающие. Подобный взгляд проник и в научно-популярную литературу (Yoon, 2009).
Несмотря на то, что степень противоречий между соперничавшими направлениями может оказаться преувеличенной (Sterner, Lidgard, 2018), вряд ли можно отрицать, что борьба идей в области теории систематики не привела к подобию “нового синтеза” в эволюционной теории. Вместо этого, как принято считать, филогенетическая систематика постепенно взяла верх над другими направлениями и в “конкурентной борьбе” вытеснила их из научного мейнстрима (Hull, 1988), что нередко описывается как “кладистическая революция” в систематике (Rieppel, 2016). Взгляды Хеннига “часто расцениваются как принципиально новая парадигма, решившая чуть ли не все проблемы классификации” (Васильева, 1999, с. 134). При этом, кладистика была скорее “антисинтетической” по своему духу, о чем будет сказано ниже.
Впрочем, существуют основания полагать, что к настоящему моменту искомый синтез в области биологической систематики достигнут или почти достигнут, но произошло это не путем развития теоретической мысли, а, как я попробую показать, стихийным образом, и было обусловлено самой логикой развития систематики как особой биологической дисциплины и ее попытками сохранить свое место под солнцем в эпоху тотального доминирования экспериментальных и высокотехнологичных разделов наук о жизни, таких как молекулярная биология и ей подобные. Достижение подобного синтеза в области практической систематики ставит со всей определенностью вопрос о будущем этой научной дисциплины, как оно видится сейчас, когда все большее значение для классификации живых организмов приобретают компьютерные технологии, включая интенсивно развивающиеся технологии искусственного интеллекта.
ТРИ ШКОЛЫ СИСТЕМАТИКИ: КРАТКОЕ СОПОСТАВЛЕНИЕ
Центральным вопросом, по поводу которого разошлись три упомянутые выше направления систематики, был вопрос о принципах построения естественной системы организмов.
В додарвиновский период истории биологии единственным базисом для классификации было морфологическое (внешнее, гораздо реже внутреннее) сходство между сравниваемыми организмами. Дарвин внес в систематику принципиально новое измерение – генеалогическое, и в “Происхождении видов” прямо указал, что “любая настоящая классификация [должна быть] генеалогической” (Darwin, 1859, p. 420). С другой стороны, в его частной переписке встречается мнение, что “генеалогия сама по себе не дает классификации” (письмо Дж. Хукеру от 23.12.1859).
Кажущееся противоречие между этими утверждениями объясняется просто. Последовательное применение генеалогического принципа классификации сталкивается с недостатком эмпирических знаний о филогенезе. Дарвин отлично понимал, что полного изоморфизма филогении и системы достичь невозможно уже в силу недостаточности наших сведений о родстве между организмами. В те годы отсутствовали алгоритмизованные методы трансляции морфологической информации в филогенетическую. Поэтому морфологическое сходство, наряду с родством, продолжало оставаться базисом для построения системы и в постдарвиновскую эпоху. Практикующие систематики и после 1859 г. долго пользовались традиционной методологией, в основе которой лежали интуитивные практики классифицирования организмов, слабо поддающиеся верификации и объективации (Винарский, 2019).
Неодарвинистский синтез середины ХХ в. благоприятствовал синтезирующим устремлениям в сообществе систематиков. Сам Майр (Mayr, 1982, p. 9) писал о том, что ему симпатична гегелевская триада “тезис–антитезис–синтез”, и что он “всегда, когда это было возможно, пытался синтезировать противоположные точки зрения (в том случае, если одна из них не была явно ошибочной)”. “Гегельянство” Майра заслужило у некоего “весьма известного советского теоретика марксизма” оценку, что его труды – “это в чистом виде диалектический материализм” (там же), хотя сам Майр марксистом себя не считал.
“Синтетический” характер эволюционной систематики в понимании Дж. Хаксли, Э. Майра и Дж.Г. Симпсона состоял не только в том, что она предполагала интеграцию данных самых разных биологических наук для целей классификации, но и в попытке объединить в рамках единой методологии сведения о родстве и сходстве как равноправных источниках таксономической информации. Как неоднократно отмечалось (см., например, Vernon, 1993), на практике это вело к неизбежности субъективных решений по ряду важнейших вопросов, включая процедуру “взвешивания” признаков, определение линнеевского ранга рассматриваемых таксономических групп и т.п. Выяснилось, что взаимоотношения между родством и сходством хорошо описываются с помощью принципа дополнительности Н. Бора, известного из квантовой механики. Чем точнее мы определяем степень сходства между организмами, тем меньше филогенетической информации это нам дает (хотя бы потому, что сходство может возникать как продукт конвергенции или параллельной эволюции, что затемняет наши представления о реальном родстве). С другой стороны, выявлять родственные отношения между организмами без оценки степени сходства между ними просто невозможно, даже если это сходство не по морфологическим, а по молекулярным признакам. Суждения о родстве почти всегда деривативны по отношению к суждениям о сходстве.
Майр не отрицал целиком и традиционный “чисто интуитивный подход” к выделению таксонов, отмечая, что в некоторых ситуациях он работает эффективнее, чем по-видимости более “объективные” методы, такие как биометрический анализ (Майр, 1971, с. 213).
В 1960-е гг. критика эволюционной систематики велась с двух флангов одновременно. В качестве альтернатив выступили две школы, фенетика и кладистика, не соглашавшиеся с эволюционными систематиками и еше менее друг с другом (Sterner, Lidgard, 2018). Их объединяло стремление к подлинному объективизму, сторонники этих школ декларировали намерения сделать систематику по-настоящему точной наукой за счет устранения произвольных таксономических решений и алгоритмизации процедуры классификации (Винарский, 2019, 2020; Hull, 1988). “Интуиция” стала фактически обсценным понятием. Достигнуть этого было возможно лишь путем редукционизма, построения системы на основе только одного из источников данных – либо сходства (программа нумерической систематики) либо родства (программа кладистики). Оба подхода позволяли разработать относительно непротиворечивую систему стандартных приемов, которые, при условии их адекватного применения, должны были вести к однозначным и воспроизводимым заключениям. Таков был идеал обеих соперничающих школ, завоевавший симпатии большого числа практикующих таксономистов, нуждавшихся в положительной переоценке их деятельности, избавлении систематики от клейма “субъективной”, “неточной” и “старомодной” научной дисциплины (Vernon, 1993).
Нумерическая систематика, в основе которой лежал “продвинутый” (advanced) статистический анализ больших массивов данных о фенотипических признаках, уже в середине 1980-х гг. почти целиком утратила свою популярность, пик которой пришелся на вторую половину 1960-х–начало 1970-х гг. Сверхформализированный, чисто количественный и откровенно антитеоретический (Hull, 1988) характер фенетической программы оказался не вполне адекватным сложности проблем, которые ставит перед исследователями задача классификации организмов во всем их многообразии. Во многих группах (например бактерии) количественный анализ морфологических признаков оказывается совершенно непригодным. В середине 1980-х гг. было признано, что фенетика “просто не порождает эффективную (workable) систематику, которая бы соответствовала ожиданиям биологов. Материальные явления попросту не могут быть уложены и упорядочены в соответствии с надеждами нумерических систематиков” (Rosenberg, 1985, p. 186). Показательна неудача чисто фенетического подхода к разработке системы покрытосеменных, однодольных и некоторых других макротаксонов растений (Stuessy, 2013).
Тем не менее, нумерическое направление в систематике оставило богатое наследство в виде большого числа статистических алгоритмов и компьютерных программ, нацеленных на определение количественной степени сходства и построение иерархических кладограмм, например, на основе кластерного анализа (Stuessy, 2013). Были разработаны методы вероятностной оценки достоверности филогенетических гипотез, позволившие уйти от казавшейся неустранимой субъективности при выборе между возможными альтернативами (Павлинов, 2005).
К концу прошлого века безусловную победу в завоевании симпатий систематиков одержал подход Вилли Хеннига, в соответствии с которым родство, определяемое путем выявления гомологий (синапоморфий), является единственным объективным основанием классификации. Степень эволюционных расхождений между дивергировавшими линиями, какой бы значительной она ни была, не принимается в расчет. По словам отца-основателя кладистики, “истинный метод филогенетической систематики – это не определение степени морфологического соответствия (correspondence) и не разграничение между “сущностными” и “несущностными” признаками, но поиск синапоморфных соответствий” (Hennig, 1966, p. 146). Признак интересует систематика лишь настолько, насколько он может выступать показателем генеалогических отношений. Вся остальная информация, которую он несет, сознательно игнорируется. Естественная система по Хеннигу представляет собой “обезьяну филогенеза”, так как иерархическая классификация в идеале должна быть максимально полным отражением паттерна ветвления филогенетического дерева.
Огромная популярность столь откровенно одностороннего, антисинтетического и редукционистского подхода обусловливается отмеченным выше стремлением сообщества систематиков объективизировать и сциентизировать свою науку, добиваясь тем самым респектабельности, позволяющей ей не чувствовать себя “золушкой” (Padial, De La Riva, 2017) в сравнении с технологичными биологическими дисциплинами – генетикой, эмбриологией, молекулярной биологией. Отсутствие такой респектабельности имело негативные последствия не только для чувства собственного достоинства систематиков, но и для финансирования их исследований (Vernon, 1993). Стоит отметить, однако, что даже почти единодушный переход таксономистов западных стран к кладистической методологии не привел к ощутимому прорыву в переоценке биологической систематики, и жалобы на ее удручающее состояние, отток денег и умов из этой дисциплины наполняют текущую литературу (Agnarsson, Kuntner, 2007; Ebach et al., 2011; Britz et al., 2020).
Тем не менее, в последние 20–30 лет систематике удалось добиться определенных успехов в улучшении своего имиджа, что было достигнуто за счет освоения новых высокотехнологичных подходов, особенно в области молекулярной генетики и искусственного интеллекта. Молекулярная революция в систематике, произошедшая на рубеже столетий, весьма серьезно повлияла как на практику классификации организмов, так и на суждения таксономистов о многих вечных вопросах, таких как сущность биологического вида.
МОЛЕКУЛЯРНАЯ РЕВОЛЮЦИЯ И “ОПЕРАЦИОНАЛЬНЫЙ СИНТЕЗ” НАЧАЛА XXI ВЕКА
Несмотря на то, что молекулярная филогенетика и основанная на ней процедура классификации подвергаются порой резкой критике [так, Н.Ю. Клюге (2020, с. 9–10) квалифицирует современные компьютерные методы реконструкции филогении как “заведомую фальсификацию” и столь же критически высказывается о “мейнстримной” молекулярной филогенетике], невозможно отрицать, что в наши дни молекулярно-генетические методы абсолютно доминируют в систематике ныне живущих организмов и, через расшифровку “древней ДНК”, проникают даже в систематику палеонтологическую. Классические морфологические подходы были вытеснены на задний план и даже в исследованиях, позиционируемых себя как “интегративно-таксономические”, нередко играют лишь вспомогательную роль (Vinarski, 2020). Эта новая реальность требует своего осмысления.
Разработка методов быстрого и относительно дешевого прочтения первичной структуры ДНК вызвала лавинообразный рост информации о геномах живых организмов, а использование современных компьютерных программ привело к тому, что задачи реконструкции филогении и, отчасти, построения системы почти целиком перешли в ведение молекулярных систематиков и биоинформатиков. Молекулярная филогенетика стала настолько самодостаточной, что уже давно никого не шокируют работы, в которых реконструкция филогенеза проводится без даже минимального обсуждения морфологических данных. Явно или неявно, генеалогия организмов отождествляется с генеалогией отдельных генов (или митохондриальных геномов), что, безусловно, очень операционально, но едва ли правомерно с биологической точки зрения. Не только рутинная таксономическая идентификация (например, путем штрихкодирования ДНК), но и такие традиционно человеческие задачи, как разделение изучаемого множества организмов на таксоны разного ранга, все больше переходят в ведение компьютерных программ. Наиболее показательна здесь эволюция подходов к выделению видов (species delimitation), в течение веков служившего источником самых резких разногласий и споров между систематиками, о чем свидетельствуют огромные списки синонимов, которые можно найти в любой таксономической монографии о хорошо изученной группе.
Ещe в конце ХХ в. наиболее жаркие дебаты велись о наиболее предпочтительной “концепции вида”, то есть о выборе частной научной теории, объясняющей, что такое вид и предлагающей критерии для распознавания видов в природе. По этому вопросу накопилась обширная литература (см.The units of …, 1992; Wilkins, 2009), но никакого консенсуса между теоретиками достичь не удалось. Казавшаяся в середине прошлого столетия абсолютно доминирующей “биологическая концепция вида” (Майр, 1971), к концу столетия стала всего лишь одним из нескольких конкурирующих подходов.
Сегодня большинство практикующих систематиков, занимающихся видовой таксономией на молекулярной основе, гораздо больше заинтересованы не в выборе правильной концепции вида, а в выборе наиболее адекватного компьютерного алгоритма для species delimitation из числа имеющихся. Из числа наиболее популярных подходов, применимых в том числе для однолокусных массивов данных, можно назвать ABGD (Automated Barcode Gap Discovery), GMYC (General Mixed Yule Coalescent), PTP (Poisson Tree Process), а также алгоритмы, основанные на байесовом методе (Rannala, 2015). В теоретическом отношении эти подходы основаны на том, что именовалось “эволюционной концепцией вида”, в соответствии с которой виды – это независимо развивающиеся филогенетические линии, распознаваемые путем анализа популяционно-генетической истории отдельных аллелей. В этом суть предложенной сравнительно недавно “унифицированной концепции” (De Queiroz, 2007), определяющей вид как самостоятельно эволюционирующую группу метапопуляций (separately evolving metapopulation lineage). Доказательство такого самостоятельного развития считается достаточным для обоснования видового статуса, даже если критерии, предлагаемые другими концепциями, не удовлетворяются. Именно такому пониманию вида явно или неявно следуют многие молекулярные таксономисты наших дней, хотя далеко не во всех случаях виды, выделяемые на основе унифицированного подхода, обладают свойством репродуктивной изоляции (Campillo et al., 2020), что считается определяющим признаком “хорошего вида” в рамках биологической концепции (Майр, 1971).
Сравнительный анализ различных статистических алгоритмов показывает, что они не всегда дают вполне сходящиеся результаты (Carstens et al., 2013), а некоторые из них ведут к таксономической инфляции, то есть явному “передроблению” видов (Chambers, Hillis, 2020; Hillis et al., 2021). Бывают ситуации, когда в работе применяются сразу несколько альтернативных алгоритмов, и авторы, столкнувшись с разнобоем в полученных результатах, вынуждены принимать решение о том, сколько же видов в исследуемой ими выборке представлено на самом деле. Здесь открывается поле для субъективных решений, от которых молекулярная систематика и статистика предполагали избавить таксономистов. Несмотря на это, популярность алгоритмического способа выделения видов постоянно возрастает (рис. 1). Налицо явный сдвиг от теоретических дебатов о сущности вида к гораздо более техническому обсуждению достоинств и недостатков отдельных статистических подходов к species delimitation. Эта “тихая революция” имеет, однако, ряд очевидных теневых сторон.
Рис. 1.
Динамика числа статей в рецензируемых журналах, посвященных статистическому подходу к выделению видов (на основе поиска по соответствующим ключевым словам в базе данных Scopus). Приведены эмпирические значения и линия тренда (пунктир).
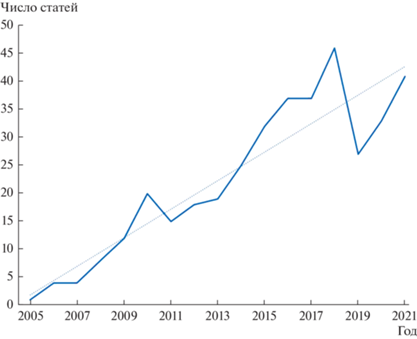
Практически во всех подобных исследованиях первым шагом является формирование библиотеки сиквенсов (оригинальных или почерпнутых из Генбанка), которые затем закладываются в компьютер, выдающий некую раскладку, представляющую собой гипотезу о числе представленных в обработанном массиве данных видов. Очень часто исследователи, работающие в таком ключе, не только не обращаются к изучению морфологических, экологических, физиологических и иных признаков изучаемых ими видов, но даже воздерживаются от привязки выделенных в ходе работы клад предположительно видового ранга к имеющимся в наличии номинальным видам, описанным на домолекулярном этапе развития систематики. Нередки и такие ситуации, когда компьютерный анализ выявляет один или несколько явно неизвестных науке видов, но и тогда авторы исследования не дают их формального описания (Vinarski, 2020). Причина проста – во многих научных коллективах, выполняющих исследования такого рода, участвуют высококвалифицированные генетики и биоинформатики, но отсутствуют специалисты с подготовкой в области систематики, знакомые с правилами таксономической номенклатуры и с процедурой описания новых таксонов.
Получается, что выделяемые в ходе работы молекулярных систематиков предполагаемые виды (putative species) или же виды-кандидаты (candidate species), практически ничего не добавляют к нашим знаниям о биологическом разнообразии, поскольку такими безымянными сущностями неясного таксономического статуса очень сложно оперировать, в том числе и для планирования и проведения природоохранных мероприятий. Эти сущности пополняют собой и без того многочисленную область, охватываемую современной “серой” номенклатурой (Minelli, 2017, 2019).
Виды, выделяемые с помощью чисто статистических приемов, и зачастую не охарактеризованные уникальными диагностическими признаками, фактически являют собой математические конструкции, которые, используя термин, предложенный А.Н. Горбанем и Р.Г. Хлебопросом (1988, с. 11), можно назвать “матемазаврами”. Согласно этим авторам, матемазавры населяют “особое “царство” формальных и математических моделей” и, несомненно, имеют определенную эвристическую ценность. Однако с точки зрения конечных целей биологической систематики – описания и упорядочивания биологического разнообразия Земли – они представляют собой лишь первый шаг исследования. Их полезность не следует преувеличивать, тем более что, как предупреждают сами авторы термина, существует реальная опасность размножения “примитивных и паразитических форм” матемазавров (там же).
Однако, несмотря на все перечисленные недостатки, очерченный выше подход к выделению видов имеет все признаки “синтетического”, являясь гибридным наследником некогда противостоящих друг другу школ – нумерической систематики и кладистики. От первой он получил статистический инструментарий и чисто алгоритмический характер, основанный на оценке сходства между нуклеотидными последовательностями. От кладистики – убежденность в том, что в основу системы должен быть положен паттерн ветвления родословного древа, выявляемый с помощью молекулярно-генетических методов, и некоторые другие конвенции, например острая неприязнь к парафилетическим таксонам (сейчас господствует хеннигово понимание монофилии как голофилии; см. Павлинов, 2005; Hennig, 1966).
Таким образом, сбылось давнее предсказание Майра, писавшего о том, что вероятностные соображения, “которые следует внести в кладистический подход, чтобы сделать его более надежным, неизбежно приблизят его к фенетическому подходу, и, возможно, лучшие элементы обоих подходов в конце концов можно будет объединить в один “синтетический” метод” (Майр, 1971, с. 249).
Было бы ошибкой, однако, рассматривать алгоритмический подход к выделению видов как осовремененную версию кладистики. При всем внешнем сходстве, в нем отсутствует ключевой для парадигмы Хеннига элемент – анализ синапоморфий, который подменен статистическим анализом генов, избранных по принципу их доступности (Клюге, 2020; Mooi, Gill, 2010). Это характерно не только для построения системы на уровне видов, но и для макротаксономии. Например, в основу наиболее популярной сегодня системы цветковых растений (так называемая система APG, Angiosperm Phylogeny Group) сознательно положены исключительно данные сравнительной геномики, а морфологические данные используются только для подтверждения полученных таким способом решений (Шипунов, 2003; Гельтман, 2019). Тем самым авторы системы APG стремились изгнать из нее субъективные экспертные суждения (Шипунов, 2003).
Вслед за А.Б. Шипуновым (2003) можно сказать, что при таком подходе единственным обоснованием для таксонов является кладограмма, паттерн ее ветвления, а не признаки самих организмов. Конечно, можно заменить одно- и даже мультилокусные филогении сравнением целых геномов, что на наших глазах становится рутинной исследовательской задачей (филогеномика), особенно применительно к вирусам, бактериям и протистам. Но даже в этом случае классическая хеннигова процедура поиска гомологий (= синапоморфий) сохранит права гражданства только там, где генетические данные недоступны, то есть в систематике большинства вымерших организмов.
Таким образом, можно говорить о том, что за последние десятилетия был реализован своего рода “операциональный синтез” в биологической систематике, достигнутый путем стихийной конвергенции некогда противостоящих друг другу таксономических методологий (см. Stuessy, 2013). Подчеркну, речь идет в первую очередь о практической систематике, в то время как теоретики продолжают говорить о плюрализме исследовательских программ и классификационных подходов в этой дисциплине (Павлинов, 2003; Pavlinov, 2020). Движущим фактором этого синтеза безусловно стало стремление сообщества систематиков к восстановлению утраченного престижа своей дисциплины, необходимость обновить ее методологию, приведя в соответствие с современными стандартами точного и доказательного научного знания (Vernon, 1993). Именно этот внешний по отношению к биологической систематике стимул, а не внутренняя логика развития теоретической мысли в этой дисциплине, приводит к унификации методов и подходов к классификации организмов, по крайней мере на видовом уровне. Материальной предпосылкой синтеза стали технологические достижения второй половины XX в., включая прогресс вычислительной техники и методов молекулярной биологии, которыми систематики успешно воспользовались для достижения своих целей.
ИСКУССТВЕННЫЙ ИНТЕЛЛЕКТ И ДЕГУМАНИЗАЦИЯ СИСТЕМАТИКИ?
Уже первые успехи нумерической таксономии вызвали опасения, что систематики будут заменены компьютерами (Hull, 1988). Пока этого не произошло, но продвинутые статистические алгоритмы и модели, методы искусственного интеллекта и машинного обучения уже становятся частью инструментария таксономистов, и остановить этот процесс едва ли возможно. Широкие возможности, которые открывает применение искусственного интеллекта (ИИ) в биологии рассмотрены в недавней статье в журнале “Природа” (Таскина и др., 2020). Систематика в этом отношении никоим образом не представляет исключения, что вызывает закономерную реакцию против возможных эксцессов автоматизации (de Carvalho et al., 2008; Sterner, Franz, 2017).
Идентификация видов животных и растений с помощью ИИ уже не является фантастикой, и, по-видимому, недалек тот день, когда не только идентификация, но и классификация живых организмов, в частности, разделение их на биологические виды, будут переданы на откуп искусственного интеллекта. Не об этом ли в свое время мечтал сэр Рональд Фишер, разрабатывая свой “дискриминантный анализ” (Fisher, 1936) для точного и однозначного распределения биологических объектов по группам? Нужно ли опасаться, что дальнейшее углубление тренда на математизацию завершится полной дегуманизацией систематики, превращением ее из области научного творчества в сверхалгоритмизированную процедуру, почти не требующую участия человека? С эмоциональной точки зрения это был бы крайне неинтересный финал многовековой истории древнейшей из биологических дисциплин. Хотя бы потому, что он означал бы полное выхолащивание биологической сущности таксонов, их окончательную редукцию к абстрактным моделям (“матемазаврам”), с которыми может иметь дело лишь компьютер, а не живой человек. Исчезло бы и эстетическое содержание систематики как науки, в том числе, и о форме живых существ, далеко не всегда поддающейся тотальной квантификации (Беклемишев, 1994) и уж тем более не сводимой нацело к нуклеотидным последовательностям. По словам А. А. Любищева (1971, с. 23), “понятию красоты нет места в биохимической систематике и морфологии, хотя неявным образом красота действует как объективный фактор в одном из разделов классического дарвинизма – гипотезе полового отбора”. Б.С. Кузин (1987, с. 137) сравнивал работу систематика с экскурсией по “великолепной картинной галерее”, доставляющей “непосредственное наслаждение”, состоящее в “любовании многообразием форм и в уловлении закономерностей, которым оно подчинено”. “Биохимия”, разлагающая целое организма на множество элементов, определенным образом взаимодействующих между собой, не видит ни смысла в изучении формы, ни ее эстетического значения, потому хотя бы, что “путь от биохимии к морфологии очень далек” (Любищев, 1971, с. 23). ИИ (как и чисто молекулярная систематика), очевидно, должен быть совершенно равнодушен к эстетической стороне вопроса, как, например, и к понятиям биостилистики, развиваемой некоторыми из современных теоретиков (Любарский, 1992, 1996). При замене целостных живых организмов математическими абстракциями или генотипами, ни о каком “любовании формами” (то есть фенотипами, данными нам в чувственных ощущениях) не может идти речи.
Можно утверждать определенно, что тренд на автоматизацию систематики, доведенный до логического предела, приведет не к смерти этой научной дисциплины, но к драматическому обеднению ее содержания, примитивизации, а также полной утрате творческого, “персоналистского” начала в ней.
Повод для сдержанного оптимизма дает то обстоятельство, что тренд на математизацию был не единственным из стимулов, управлявшим развитием биологической систематики за последнее столетие. Параллельно шло движение в сторону “интегративности” этой науки, выстраиванию ее междисциплинарных связей с другими областями биологии. Систематика наших дней утратила привычный образ науки одиночек, уединенного занятия музейных кураторов, “накалывающих букашек на булавки” и дающих им латинские названия. Очень ярко это проявляется в возрастании числа соавторов таксономических публикаций, для которых уже не редкость коллективы, включающие по десять и более участников (Гаврилов-Зимин, 2021; Poulin, Preswell, 2016; Vinarski, 2020).
По моему мнению, стратегией для сохранения биологической систематики как “человеческой науки” может стать ее постепенная интеграция (вместе с биогеографией, экологией, палеобиологией) в некую метанауку “о биоразнообразии”, рассматривающую его как сложный, целостный объект, имеющий исторический, таксономический, функциональный и другие аспекты. В ходе таких “метатаксономических” исследований систематик будет работать как часть команды, наряду с представителями других биологических дисциплин, создавая многомерный образ изучаемого таксона в различных его проявлениях. Возможно, это будет дисциплина, целиком ориентированная на реконструкцию эволюционных “историй”, использующая стилистику нарратива для всестороннего описания развивающейся во времени группы организмов, включая описание ее таксономического состава и структуры. В таком подходе нет никакой новизны. Насколько я могу судить, именно к этому призывал Джулиан Хаксли (Huxley, 1940) и другие архитекторы “новой систематики”. Вызовы наших дней, к числу которых относятся и современный кризис биоразнообразия (Myers, 1996; Pereira et al., 2012; Neubauer et al., 2021), и постоянно оплакиваемый кризис традиционной систематики (Agnarsson, Kuntner, 2007; Britz et al., 2020; Vinarski, 2020; Bond et al., 2022), а также реальная угроза ее дегуманизации, должны, как кажется, способствовать ускоренной трансформации этой классической дисциплины в новое, “постклассическое” состояние, одним из ключевых понятий в описании которого будет “синтез”.
Однако сам по себе “стихийный” синтез, основанный исключительно на прогрессе технологии, а не методологии, скорее всего будет бесплодным, так как заведет биологическую систематику в бесконечный тупик совершенствования компьютерных алгоритмов. Необходимо появление ярких новых идей в области теории (или воскрешение идей, давно высказанных, но крепко забытых).
Список литературы
Беклемишев В.Н. Методология систематики. М.: Товарищество научных изданий КМК, 1994. 250 с.
Боркин Л.Я. Через кладизм – к новой систематике? // Тр. Зоол. ин-та АН СССР. 1989. Т. 206. С. 3–7.
Васильева Л.Н. Кладистика – глазами типолога. 1. Парадигма Хеннига // Журн. общ. биол. 1999. Т. 60. № 2. С. 133–149.
Винарский М.В. Интуитивизм в биологической систематике: из прошлого в будущее // Философия науки. 2019. № 2 (81). С. 135–151.
Винарский М.В. Утопические проекты в отечественной зоологической систематике 1920-х гг.: Е.С. Смирнов и В.Н. Беклемишев // Историко-биол. исслед. 2020. Т. 12. № 2. С. 25–43.
Гаврилов-Зимин И.А. Коллективизация науки на примере систематики живых организмов // Социол. науки и технол. 2021. Т. 12. № 2. С. 90–111.
Галл Я.М. Джулиан Сорелл Хаксли. 1887–1975. СПб.: Наука, 2004. 294 с.
Гельтман Д.В. Современные системы цветковых растений // Бот. журн. 2019. Т. 104. № 4. С. 503–527.
Горбань А.Н., Хлебопрос Р.Г. Демон Дарвина: идея оптимальности и естественный отбор. М.: Наука, 1988. 208 с.
Клюге Н.Ю. Систематика насекомых и принципы кладэндезиса. М.: Товарищество научных изданий КМК, 2020. Т. 1. I–IV + 1–509 + I–V с.
Колчинский Э.И. Единство эволюционной теории в разделенном мире XX века. СПб: Нестор-История, 2015. 824 с.
Кузин Б.С. Принципы систематики // Вопр. истории естествознания и техники. 1987. № 4. С. 134–143.
Любарский Г.Ю. Биостилистика и проблема классификации жизненных форм // Журн. общ. биол. 1992. Т. 53. № 5. С. 649–661.
Любарский Г.Ю. Архетип, стиль и ранг в биологической систематике. М.: Товарищество научных изданий КМК, 1996. 434 с.
Любищев А.А. Значение и будущее систематики // Природа. 1971. № 2. С. 15–23.
Майр Э. Принципы зоологической систематики. М.: Мир, 1971. 456 с.
Павлинов И.Я. Разнообразие классификационных подходов – это нормально // Журн. общ. биол. 2003. Т. 64. № 4. С. 275–291.
Павлинов И.Я. Введение в современную филогенетику. М.: Товарищество научных изданий КМК, 2005. 391 с.
Расницын А.П. Процесс эволюции и методология систематики // Тр. Рус. энтомол. общества. 2002. Т. 73. С. 1–107.
Таскина А.К., Муравьева А.А., Ельсукова А.С., Фишман В.С. Методы машинного обучения в биологии // Природа. 2020. № 9. С. 3–17.
Шипунов А.Б. Система цветковых растений: синтез традиционных и молекулярно-генетических подходов // Журн. общ. биол. Т. 64. № 6. С. 499–507.
Agnarsson I., Kuntner M. Taxonomy in a changing world: seeking solutions for a science in crisis // Syst. Biol. 2007. V. 56. P. 531–539.
Bond J.E., Godwin R.L., Colby J.D. et al. Improving taxonomic practices and enhancing its extensibility – an example from araneology // Diversity. 2022. V. 14. № 5. https://doi.org/10.3390/d14010005
Britz R., Hundsdörfer A., Fritz U. Funding, training, permits – the three big challenges of taxonomy // Megataxa. 2020. V. 1. P. 49‒52.
Campillo L.C., Barley A.J., Thomson R.C. Model-based species delimitation: are coalescent species reproductively isolated? // Syst. Biol. 2020. V. 69. № 4. P. 708–721.
Carstens B.C., Pelletier T.A., Reid N.M., Satler J.D. How to fail at species delimitation // Mol. Ecol. 2013. V. 22. № 17. P. 4369–4383.
Chambers E.A., Hillis D.M. The multispecies coalescent over-splits species in the case of geographically widespread taxa // Syst. Biol. 2020. V. 69. P. 184–193.
Cracraft J. Species concepts in theoretical and applied biology: a systematic debate with consequences // Species concepts and phylogenetic theory: a debate / Eds Q.D. Wheeler, R. Meier. New York: Columbia Univ. Press, 2000. P. 3–14.
Darwin Ch. On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. London: J. Murray, 1859. 502 p.
de Carvalho M.R., Bockmann F.A., Amorim D.S., Brandão C.R.F. Systematics must embrace comparative biology and evolution, not speed and automation // Evol. Biol. 2008. V. 35. P. 150–157.
De Queiroz K. Species concepts and species delimitation // Syst. Biol. 2007. V. 56. № 6. P. 879–886.
Ebach M.C., Valdecasas A.G., Wheeler Q.D. Impediments to taxonomy and users of taxonomy: accessibility and impact evaluation // Cladistics. 2011. V. 27. P. 550–557.
Felsenstein J. The troubled growth of statistical phylogenetics // Syst. Biol. 2001. V. 50. № 4. P. 465–467.
Fisher R.A. The use of multiple measurements in taxonomic problems // Annals of Eugenics. 1936. V. 7. № 2. P. 179–188.
Hall A.V. A joint phenetic and cladistic approach for systematics // Biol. J. Linn. Soc. 1988. V. 33. № 4. P. 367–382.
Hennig W. Phylogenetic systematics. Urbana: The University of Illinois Press, 1966. 263 p.
Hillis D.M., Chambers E.A., Devitt T.J. Contemporary methods and evidence for species delimitation // Ichthyol. Herpetol. 2021. V. 109. № 3. P. 895–903.
Hull D.L. Contemporary systematic philosophies // Annu. Rev. Ecol. Syst. 1970. V. 1. P. 19–54.
Hull D.L. Science as a process: an evolutionary account of the social and conceptual development of science. Chicago–London: The University of Chicago Press, 1988. 586 p.
Huxley J. (ed.) The new systematics. Oxford: Clarendon Press, 1940. 583 p.
Huxley J. Evolution. The modern synthesis. London: Allen & Unwin, 1942. 652 p.
Mayr E. Biological classification: toward a synthesis of opposing methodologies // Science. 1981. V. 214. P. 510–516.
Mayr E. The growth of biological thought. Cambridge (MA): The Belknap Press, 1982. 974 p.
Minelli A. Grey nomenclature needs rules // Ecologica Montenegrina. 2017. V. 7. P. 654–666.
Minelli A. The galaxy of the non‑Linnaean nomenclature // Hist. Philos. Life Sci. 2019. V. 41. № 31. https://doi.org/10.1007/s40656-019-0271-0
Mooi R.D., Gill A.C. Phylogenies without synapomorphies – a crisis in fish systematics: time to show some character // Zootaxa. 2010. V. 2450. P. 26–40.
Myers N. The biodiversity crisis and the future of evolution // The Environmentalist. 1996. V. 16. P. 37–47.
Neubauer T.A., Hauffe T., Silvestro D. et al. Current extinction rate in European freshwater gastropods greatly exceeds that of the late Cretaceous mass extinction // Commun. Earth Environ. 2021. V. 2. № 97. https://doi.org/10.1038/s43247-021-00167-x
Padial J.M., De La Riva I. Taxonomy, the cinderella of science, hidden by its evolutionary stepsister // Zootaxa. 2007. V. 1577. P. 1–2.
Pavlinov I.Ya. Multiplicity of research programs in the biological systematics: a case for scientific pluralism // Philosophies. 2020. V. 5. № 7. https://doi.org/10.3390/philosophies5020007
Pereira H.M., Navarro L.M., Martins I.S. Global biodiversity change: the bad, the good, and the unknown // Ann. Rev. Environ. Res. 2012. V. 37. P. 25–50.
Poulin R., Presswell B. Taxonomic quality of species descriptions varies over time and with the number of authors, but unevenly among parasite taxa // Syst. Biol. 2016. V. 65. № 6. P. 1107–1116.
Rannala B. The art and science of species delimitation // Curr. Zool. 2015. V. 61. № 5. P. 846–853.
Rieppel O. Re-writing Popper’s philosophy of science for systematics // Hist. Philos. Life Sci. 2008. V. 30. P. 293–316.
Rieppel O. Phylogenetic systematics: Haeckel to Hennig. Boca Raton: CRC Press, 2016. 402 p.
Rosenberg A. The structure of biological science. Cambridge: Cambridge Univ. Press, 1985. 281 p.
Smokovitis V.B. Unifying biology: the evolutionary synthesis and evolutionary biology // J. Hist. Biol. 1992. V. 25. P. 1–65.
Sterner B., Franz N.M. Taxonomy for humans or computers? Cognitive pragmatics for big data // Biol. Theory. 2017. V. 12. P. 99–111.
Sterner B., Lidgard S. Moving past the systematics wars // J. Hist. Biol. 2018. V. 51. P. 51–67.
Stuessy T.F. Schools of data analysis in systematics are converging, but differences remain with formal classification // Taxon. 2013. V. 62. № 5. P. 876–885.
The evolutionary synthesis / Eds E. Mayr, W. Provine. Cambridge (MA): Harvard Univ. Press, 1980. 487 p.
The units of evolution: essays on the nature of species / Ed. M. Ereshefsky. Cambridge (MA)–London: Bradford Book; The MIT Press, 1992. 404 p.
Vernon K. Desperately seeking status: evolutionary systematics and the taxonomists' search for respectability 1940–60 // Brit. J. Hist. Sci. 1993. V. 26. № 2. P. 207–227.
Vinarski M.V. The roots of the taxonomic impediment: is the ‘integrativeness’ a remedy? // Integr. Zool. 2020. V. 15. № 1. P. 2–15.
Wilkins J.S. Species: A history of the idea. Berkeley etc: Univ. of California Press, 2009. 305 p.
Yoon C.K. Naming nature: the clash between instinct and science. New York–London: W.W. Norton & Co, 2009. 344 p.
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии