

Успехи современной биологии, 2022, T. 142, № 3, стр. 223-252
Развитие теоретических представлений о живорождении
1 Санкт-Петербургский филиал Института истории естествознания
и техники им. С.И. Вавилова РАН
Санкт-Петербург, Россия
2 Зоологический институт РАН
Санкт-Петербург, Россия
* E-mail: coccids@gmail.com
Поступила в редакцию 17.11.2021
После доработки 25.02.2022
Принята к публикации 27.02.2022
- EDN: UEMXKL
- DOI: 10.31857/S0042132422030036
Аннотация
Обсуждаются основные теоретические аспекты изучения живородящих групп организмов, а также предлагаются и аргументируются новые подходы к пониманию эволюции живорождения. Само это явление рассматривается, вопреки широко распространенному мнению, не как “большое биологическое преимущество”, а как заведомо проигрышный в отношении биологического разнообразия, простейший способ рождения потомства, актуальный лишь в тех случаях, когда по разным причинам откладка яиц оказывается невозможной или плохо совместимой с новоприобретенными морфоанатомическими и/или физиологическими особенностями. Аберрации репродуктивной сферы, изменяющие ее предковое состояние и приводящие к живорождению, можно разделить на три основные группы: 1) возникновение оогамии у древних многоклеточных организмов при отсутствии каких-либо механизмов выведения неподвижной зиготы во внешнюю среду; 2) переходы от наружного оплодотворения к внутреннему в ситуации, когда у организма еще отсутствуют специализированные половые протоки (яйцеводы) с придаточными железами и сперматеками; 3) нарушения в хорошо развитой половой яйцекладной системе: утрата имагинальных половых органов (вследствие педогенеза, неотении, педоморфоза), ларвальный мейоз, изменение места оплодотворения с эктодермальных частей половой системы на внутригонадное. Перечисленные явления обсуждаются в статье на основе обзора-анализа живорождения в крупнейших таксонах живых организмов, от примитивных многоклеточных до наиболее высокоразвитых позвоночных и беспозвоночных животных.
ВВЕДЕНИЕ
Живорождение, как природное явление, привлекало к себе пристальное внимание исследователей с очень давних времен в связи с тем, что сам биологический вид Homo sapiens Linnaeus, 1758 принадлежит к одной из живородящих групп животных. Разнообразные примитивные рисунки, изображающие материнский организм с развивающимся внутри него эмбрионом, известны даже во вполне первобытных обществах. В государствах древней Индии, Китая, Ассирии, Египта были в той или иной степени разработаны основы акушерства и гинекологии, сопровождавшиеся полуфантастическими представлениями о том, откуда появляется ребенок в организме матери (Нидхэм, 1947). Научный подход к осмыслению живорождения (как и естествознание в целом) впервые появляется в работах древнегреческих мыслителей, достигая своего пика на следующие две тысячи лет в знаменитых книгах Аристотеля (384–322 гг. до н. э.): “История животных”, “О возникновении животных”, “О частях животных”, написанных в IV веке до н.э. В этих работах впервые формулируется ясное понимание того, что человек – это лишь одно из множества живородящих существ на нашей планете. По-видимому, Аристотель лично производил вскрытия и изучение репродуктивной системы многих из описываемых им организмов. Он впервые открыл плаценту у акул и китообразных, обнаружил яйцеживорождение у пресмыкающихся и скорпионов, впервые разделил основные группы животных на “живородящих”, “яйцеродящих” и “черверодящих”, что отчасти (для позвоночных) совпадает и с современной классификацией (Плавильщиков, 1941; Нидхэм, 1947; Blackburn, Stewant, 2021). Рисунков, выполненных Аристотелем, к сожалению, не сохранилось. Лишь спустя два с лишним тысячелетия, в работах итальянских и немецких естествоиспытателей появятся иллюстрации, сделанные на основе аристотелевских описаний и впоследствии часто использовавшиеся многими другими, в том числе русскоязычными авторами, пишущими об Аристотеле (см., например, рис. на стр. 18 в книге: Плавильщиков, 1941). У Аристотеля же, по-видимому, впервые, появляется идея о “совершенных и несовершенных животных” (Аристотель, 1940, с. 93–94). К “самым совершенным”, в силу характерного для людей, в том числе и для великих философов, антропоцентрического мышления, отнесены млекопитающие, включая человека, и их “совершенность” обусловлена у Аристотеля именно тем, что они “рождают сразу живых себе подобных существ”, якобы минуя стадию яйца. Это, во всех отношениях ошибочное мнение (см. подробнее ниже), на тысячелетия предопределило ход теоретических рассуждений в области биологии живородящих организмов.
Эмбриологические достижения последующих античных авторов, прежде всего Галена (129–216 гг. н. э.), сводились, главным образом, к анатомическим исследованиям развивающихся зародышей человека и некоторых других позвоночных животных, особенно птиц (см. подробнее в книге: Нидхэм, 1947) и, в целом, добавляют мало нового к прояснению общей картины распространения живорождения.
Дальнейшее 1000-летнее тотальное господство христианской идеологии во всех сферах деятельности европейского общества в корне подрывает научный подход к познанию природы и не привносит в развитие эмбриологии (и биологии в целом) почти ничего нового. Кочуют из одной рукописной книги в другую цитаты из Аристотеля, добавляются новые ошибки и фантастические представления о возникновении организмов. Например, абсолютно ошибочный аристотелевский тезис о том, что личинки различных беспозвоночных животных, особенно насекомых – это, якобы, недозревшие яйца (см., например, Аристотель, 1940, с. 94) повторяется из книги в книгу на протяжении столетий. Некоторые новые фактические данные, касающиеся развития рыб, добавляются лишь у Альберта Больштедского (известного как Альберт Великий), жившего в XIII в. в Германии. Персидские и арабские ученые, много сделавшие в Средние Века для развития медицины, математики, астрономии и некоторых других наук, в области эмбриологии не добавляют ничего принципиально нового в сравнении с античными авторами. Лишь спустя почти 2000 лет после Аристотеля, в Западной Европе, вступившей в эпоху Возрождения античных наук и искусств, наконец, начинается прогрессивное развитие эмбриологических исследований. Появляются детальные анатомические рисунки, иллюстрирующие развитие эмбриона внутри материнского организма. Одни из первых таких рисунков содержатся в анатомических дневниках Леонардо да Винчи “Quaderni d’Anatomia”, датируемых приблизительно 1490-м годом. К середине XVI в. публикуются иллюстрированные книги Пьера Белона (1518–1564) и Гийома Ронделе (1507–1566), содержащие, среди прочего, рисунки репродуктивной системы живородящих рыб и китообразных. Особенно значительный вклад в сравнительную эмбриологию того периода внес итальянский врач и анатом Джироламо Фабрициус Аквапендентский (1533–1619), который, помимо человеческого эмбриона, изучал эмбрионы кроликов, мышей, собак, ряда других млекопитающих, а также гадюк и акул.
Ко второй половине XVII в., трудами таких знаменитых анатомов как Вильям Гарвей (1578–1657), Ренье де Грааф (1641–1673) и Николай Стенон (1638–1686), было впервые показано, что эмбрионы живородящих организмов точно так же, как и яйцекладущих, образуются исходно из яйцевых клеток, а не из сгустков крови или спермы, как считалось до этого на протяжении тысячелетий и кочевало из книг античных авторов по различным средневековым рукописям. Сравнительно малоизвестный английский естествоиспытатель Мартин Листер (1639–1712) одним из первых сообщает научной общественности о наличии живорождения у насекомых, а именно у одного из видов мух (Diptera) (Lister, 1671). Кроме того, в XVII веке вопросы эмбриологии живородящих организмов так или иначе затрагивают Теодор Краан (1633–1688), Вальтер Нидхэм (1631–1691), Антони ван Левенгук (1632–1723), Франческо Реди (1626–1698), Ян Сваммердам (1637–1680), то есть крупнейшие естествоиспытатели того времени.
XVIII в. – это период оформления биологии современного типа и, в частности, время появления многочисленных детальных исследований по эмбриологии различных групп растений и животных. Наиболее яркие фигуры этого периода, имеющие отношение к обсуждаемой теме – французский естествоиспытатель Рене Антуан Реомюр (1683–1757) и швейцарский натуралист и философ Шарль Бонне (1720–1793). Оба они много работали с беспозвоночными животными, прежде всего с насекомыми, которые демонстрируют наиболее сложные формы живорождения. Ш. Бонне, кроме того, известен своей знаменитой лестницей природы, на которой он изобразил живые и неживые объекты в иерархическом порядке, поместив млекопитающих и человека выше всех остальных существ, но ниже ангелов, серафимов и херувимов. Позднее эти представления об иерархичности природы трансформировались в гораздо более содержательную в научном плане лестницу природы Жана-Батиста Ламарка (1744–1829), хотя антропоцентричное представление о том, что живородящие млекопитающие являются вершиной эволюции, в полной мере сохраняется и у него.
В XIX–XX вв. вырисовывается уже относительно полная картина распространенности живородящих организмов среди основных таксонов животных и растений. Выясняется, что в огромном разнообразии организмов на нашей планете такие группы составляют скорее исключение, тогда как обычным способом отрождения потомства следует считать откладку яиц. Появляются обобщающие исследования, среди которых наиболее значительными можно признать монографию Гарольда Хейгана (Hagan, 1951), посвященную живорождению у насекомых – крупнейшей группы организмов, и выдающуюся серию монографий О.М. Ивановой-Казас (1975, 1977, 1978а, б, 1979, 1981, 1995) по эволюционной эмбриологии всех животных, где теме живорождения отведено существенное место. Интересный (и, кажется, единственный в своем жанре) обзор живородящих организмов сделан в учебном пособии, подготовленном группой петербургских ботаников и зоологов Т.Б. Батыгиной, Е.А. Брагиной, А.В. Ересковским и А.Н. Островским (Батыгина и др., 2006). Два последних автора в дальнейшем, совместно с зарубежными коллегами, опубликовали на английском языке обзорную работу по матротрофии (то есть живорождению и различным вариантам заботы о потомстве у беспозвоночных животных (Ostrovsky et al., 2016)). Аналитические обзоры по живорождению у позвоночных животных разной степени детализации также неоднократно публиковались в последние десятилетия (см., например, Wourms, 1994; Blackburn, 2015; Blackburn, Starck, 2015 и др.).
Несмотря на обилие современных исследовательских и обобщающих работ по живорождению, в теоретическом и терминологическом аспектах эта тема во многом остается в плену у очень старых традиций и представлений, сложившихся на заре естествознания, а обсуждение различий между многочисленными вариантами живорождения и яйцеживорождения зачастую сводится к “спору из-за слов”, нежели к оценке смыслового, в частности эволюционного, наполнения этих слов. Многие из вводившихся когда-либо терминов носят явно избыточный характер, перекрываются по смыслу с другими широко используемыми терминами, или же вовсе не соответствуют биологическим фактам.
ОСНОВНАЯ ТЕРМИНОЛОГИЯ
Живорождение (= вивипария; viviparity) – способ отрождения потомства многоклеточных организмов, при котором развитие эмбриона (а иногда и ранних личиночных стадий) проходит внутри тела матери и сопровождается получением от нее питательных веществ. Получение питания от материнского организма – принципиальный момент, отличающий живорождение от яйцеживорождения (см. Hagan, 1951). В связи с тем, что переход к такому питанию связан с теми или иными структурно-физиологическими модификациями половой системы, живорождение не бывает факультативным или характерным лишь для отдельных особей (в отличие от яйцеживорождения), но всегда является таксономическим признаком, характеризующим целиком некий вид или таксоны более высокого ранга.
Относительно четкую терминологическую границу можно провести между следующими вариантами живорождения (по: Hagan, 1951, с моими небольшими изменениями, дополнениями и комментариями – И. Г.-З.).
Аденотрофное живорождение (adenotrophic viviparity) представляет собой вариант эмбриогенеза, при котором зародыш развивается преимущественно за счет запаса питательных веществ, имеющихся в желтке, но на поздних этапах развития или уже на стадии личинки первого возраста питается выделениями половых путей матери. Такой тип развития характерен для некоторых групп беспозвоночных животных: некоторых брюхоногих моллюсков, онихофор, ряда семейств кровососущих мух (в частности, для мух це-це) и комаров-звонцов (Hagan, 1951; Meier et al., 1999; Иванова-Казас, 1977).
Внутриполостное (целомическое или гемоцельное) живорождение (coelomic or haemocoelous viviparity) – вариант эмбриогенеза, при котором развитие и внешнее питание зародыша происходит не в половых путях матери, а в ее целоме (гемоцеле), реже в иных по онтогенетическому происхождению внутренних полостях тела (схизоцель, миксоцель и др.). Встречается, как минимум, у некоторых мшанок, скребней и ряда насекомых из отрядов Diptera и Strepsiptera (см. подробнее: Иванова-Казас, 1975, 1978а; Батыгина и др., 2006; Hagan, 1951).
Плацентарное живорождение (placental viviparity) – вариант эмбриогенеза, при котором питание развивающегося зародыша осуществляется через плаценту, образуемую разрастающимися тканями материнского и эмбрионального происхождения либо только эмбрионального. В отношении беспозвоночных животных часто употребляется словосочетание “псевдоплацентарное живорождение”. Однако, как уже отмечалось ранее (Иванова-Казас, 1995, с. 266; Gavrilov-Zimin, 2018; Gavrilov-Zimin et al., 2021), для использования такого термина нет ни морфофункциональных, ни эволюционных оснований. Как среди позвоночных, так тем более среди беспозвоночных животных, плаценты существенно различаются по строению и возникали многократно независимо в разных филогенетических линиях. Более того, даже у млекопитающих имеются разные по строению и гистологическому происхождению типы плацент (Kardong, 2012, с. 193). Таким образом, нужно либо для каждого случая придумывать отдельный термин, либо все аналогичные структуры называть плацентой.
Живородящие растения – широко распространенное в научной и научно-популярной литературе словосочетание, впервые использованное, вероятно, Карлом Линнеем и с тех пор некорректно применяемое в большинстве случаев к различным растениям с вегетативным (клональным) размножением и отпочкованием новых клонов от листьев, стеблей, корней, луковиц, вегетативных тканей соцветий и т.д. исходного растения. Доказанные случаи облигатного прорастания семян внутри плодов материнского растения известны лишь у представителей отдельных, не родственных родов растений (из сем. Avicenniaceae, Arecaceae, Cymodoceaceae, Myrsinaceae, Pellicieraceae, Plumbaginaceae, Rhizophoraceae), образующих экологические сообщества мангровых зарослей (Батыгина и др., 2006). Однако применение термина “живорождение” даже по отношению к этим случаям сопряжено с некоторой двусмысленностью, поскольку у высших растений, а также водорослей и грибов отсутствует само явление яйцекладки (выведение яйцеклеток во внешнюю среду), которому можно было бы противопоставить живорождение. Нет у растений и такого этапа онтогенеза, как развитие эмбриона внутри яйца (оболочки яйцеклетки). Более того, гаметы многоклеточных растений (а также грибов) образуются в результате митоза, а не мейоза (как у животных). При этом у водорослей, грибов и высших растений оогамета (яйцеклетка) прорастает в спорофит, не отделяясь от гаметофита. Семена голосеменных и покрытосеменных растений – это уже сложные многоклеточные структуры, сочетающие в себе ткани разного онтогенетического происхождения, в то время как яйцо животных исходно представляет собой гамету, образовавшуюся в результате мейоза, то есть гаплоидное одноклеточное образование, которое выводится во внешнюю среду или начинает развиваться на месте происхождения. Учитывая такую ботаническую специфику полового размножения, правильнее называть растительные организмы, обладающие оогамией, спорородящими и/или семяродящими, а термин “живорождение” употреблять только по отношению к животным, как, собственно, и делал сам автор термина, то есть Аристотель, понимавший живорождение как противоположность яйцекладке.
Вегетативное живорождение или вегетативная вивипария – парадоксальный термин, применяемый рядом ботаников в отношении отдельных, произвольно выбранных случаев вегетативного размножения растений (Батыгина и др., 2006). По смыслу этот термин можно было бы уподобить выражениям “немасляное масло” или “твердая жидкость”, поскольку при вегетативном размножении отсутствует сам факт рождения нового организма, то есть возникновение его из яйцеклетки, зиготы или хотя бы из споры. Следуя этому вольному подходу к терминологии, вивипарией можно было бы называть по желанию также и любые многочисленные и разнообразные случаи бесполого размножения у животных, грибов и даже одноклеточных организмов. Почему, например, не назвать “живорождением” появление из старого мицелия новых тел грибов, образование дочерних колоний вольвоксов, “почкование” колониальных полипов, бесполое размножение плоских червей и т.д.? В настоящей статье термин не используется, как биологически необоснованный.
Ларвальный мейоз (larval meiosis) – мейотическое деление генеративных клеток, приводящее к образованию гамет на личиночной стадии. Термин был предложен мною (Gavrilov-Zimin et al., 2015) для описания цитогенетических механизмов размножения кокцид и тлей, но может применяться для всех педогенетических и неотенических организмов, а также для любых животных, у которых образование гамет происходит до, предусмотренного в онтогенезе, появления половых протоков и копулятивных органов. В то же время, ларвальный мейоз не во всех случаях приводит к утрате имагинальной стадии; эта стадия может сохраняться и оставаться стадией размножения.
Марзупиальность (marsupiality) – вынашивание потомства в различных полостях, образуемых наружными (эктодермальными) покровами тела. Разные по строению и гистологическому происхождению марзупии известны у ракообразных, некоторых насекомых (тараканов, кокцид), голотурий, сумчатых млекопитающих и некоторых других животных.
Матротрофия (matrotrophy) – термин, широко применяемый в отношении разнообразных случаев выкармливания родительскими особями своего потомства, как на этапе эмбрионального развития, так и в постэмбриональный период (см. Blackburn, 2015; Ostrovsky et al., 2016 и др.). Живорождение при таком подходе рассматривается как частный случай матротрофии, наряду с марзупиальностью и другими способами выкармливания несамостоятельных детенышей (личинок). При этом эволюционное происхождение живорождения и других вариантов матротрофии может быть различным.
Лецитотрофное живорождение (lecithotrophic viviparity) – часто используемый, особенно в англоязычной литературе, неудачный термин, обозначающий питание зародыша исключительно за счет желтка. По смыслу этот термин перекрывается с яйцеживорождением и в настоящей статье не используется, как избыточный.
Неотения (neoteny) – эволюционный переход к половому размножению на личиночных стадиях развития. Термин был исходно введен в научный оборот Кольманном (Kollmann, 1884) для описания размножения личинок саламандр, но в дальнейшем стал очень широко применяться в литературе по самым разным группам позвоночных и беспозвоночных животных, а также растений.
Педогенез (paedogenesis) – эволюционный переход к партеногенетическому размножению на личиночных стадиях развития. Термин был введен К. Бэром (1866) для описания репродуктивной стратегии галлиц (Diptera: Cecidomyiidae). В настоящее время широко используется в зоологической литературе.
Педоморфоз (paedomorphosis) – сохранение личиночных признаков у взрослого организма. Термин был предложен Гарстангом (Garstang, 1922, p. 97) и впоследствии сильно вульгаризировался, в результате чего это понятие часто ошибочно смешивается с педогенезом и неотенией.
Прогенез (progenesis) – младший синоним неотении. Введен в научный оборот в статье (Giard, 1887). Часто используется в литературе без ясного указания отличий от неотении (см. обсуждение этой проблемы у Ивановой-Казас, 1997, с. 1245).
Псевдоплацентарное живорождение (pseudoplacental viviparity) – см. выше комментарии к термину “плацентарное живорождение”.
Псевдоживорождение (pseudoviviparity) – под этим термином предлагалось (см. Blackburn, 2015 и др.) понимать разнообразные случаи вынашивания потомства в полостях тела, не связанных с репродуктивной системой: в ротовой или брюшной полостях, в специальной кожаной или кутикулярной сумке (марзупии). На мой взгляд, термин является неудачным и избыточным, поскольку перекрывается с термином “матротрофия” и объединяет разные по своей биологической сути и эволюционному происхождению явления.
Яйцеживорождение (ovoviviparity) – способ отрождения потомства, при котором развитие зародыша полностью или частично проходит внутри тела матери, но не сопровождается получением от материнского организма питательных веществ, а осуществляется за счет имеющихся в яйце запасов желтка. При полном яйцеживорождении (complete ovoviviparity) эмбриогенез завершается внутри материнского организма, тогда, как при неполном яйцеживорождении (incomplete ovoviviparity) яйцо откладывается во внешнюю среду на разных промежуточных стадиях эмбрионального развития (см., например, Gavrilov-Zimin, 2018, 2021). Факультативное яйцеживорождение характеризуется необязательностью задержки дробящегося яйца в половых путях самки; такая задержка может возникать в связи с разнообразными случайными физиологическими и климатическими обстоятельствами у отдельных особей или отдельных популяций яйцекладущего вида (см., например, Захваткин, 1966; Трапезникова, Гаврилов, 2008). Напротив, облигатное яйцеживорождение обычно является таксономическим признаком, характеризующим в целом некий вид или таксоны более высокого ранга (Gavrilov-Zimin, 2018, 2021). В ряде случаев провести границу между яйцеживорождением и живорождением бывает затруднительно в силу необходимости проведения сложных эмбриологических и анатомических исследований или по причине сочетания у одного и того же вида разных вариантов задержки потомства внутри тела материнского организма. Ниже, когда конкретный вариант живорождения или яйцеживорождения не установлен или же не имеет принципиального значения для дискуссии я буду употреблять термин “(яйце)живорождение” с соответствующими скобками.
Яйцерождение или яйцекладка (oviparity or oviposition) – откладка яйцеклеток (гамет) или покрытых дополнительными оболочками яиц во внешнюю, по отношению к организму, среду на стадии, предшествующей эмбриональному развитию. Такая репродуктивная стратегия характерна для подавляющего большинства групп животных (Иванова-Казас, 1975, 1977, 1978а, б, 1979, 1981, 1995).
В научной литературе фигурирует также ряд других терминов, прямо или косвенно относящихся к теме живорождения, но не имеющих значения для обсуждения общетеоретических проблем. Все эти термины с легкостью могут быть найдены заинтересованным читателем в основных обзорных работах: (Батыгина и др., 2006; Wourms, 1994; Meier et al., 1999; Blackburn, 2015; Ostrovsky et al., 2016 и др.).
ТЕОРИЯ ЖИВОРОЖДЕНИЯ
В отличие от многих других общебиологических проблем, теоретическая база представлений о живорождении и сходных вариантах репродуктивной стратегии остается разработанной весьма слабо. До сих пор нет общей теории и однозначных ответов на фундаментальные вопросы об эволюционном значении живорождения, причинах его спорадического появления у отдельных таксономических групп, обратимости/необратимости в ходе филогенеза, таксономическом значении и т.д. В приведенном ниже обсуждении я попытаюсь предложить небанальные ответы на эти вопросы и представить имеющуюся в моем распоряжении аргументацию.
Столетиями из статьи в статью в литературе по живородящим организмам повторялись и продолжаются по сей день попытки напрямую увязать возникновение живорождения с теми или иными экологическими причинами: холодным или жарким климатом, хорошим или плохим питанием, мелководным или глубоководным образом жизни и т.д. (см. обзоры таких представлений у: Батыгина и др., 2006; Hagan, 1951; Sellier, 1955; Wourms, 1994; Pyron, Burbrink, 2013). Однако такому подходу противоречит тот очевидный и общеизвестный факт, что в любой из живородящих групп организмов есть виды, как близкие по своим экологическим особенностям, так и весьма далекие. В то же время не вызывают сомнений многочисленные факты непродолжительной факультативной задержки яиц в половых путях самки в результате воздействия разнообразных случайных причин, таких, как резкое похолодание, затопление, отсутствие подходящего места для яйцекладки, болезнь или преждевременная смерть и т.п. (см. выше раздел Терминология – факультативное яйцеживорождение).
Столь же удивительны непрекращающиеся многие десятилетия попытки объяснять живорождение в рамках абстрактных экологических понятий о r- и K-стратегиях (см., например, Wourms, 1994, p. 553; Vreysen et al., 2013), хотя живорождение само по себе никак не влияет ни на скорость размножения, ни на скорость освоения среды обитания, ни на продолжительность жизни. Бессмысленность дискуссии на эту тему можно проиллюстрировать простым вопросом: к какой стратегии в рамках r- и K-отбора нужно отнести живородящих слонов, мышей, тлей, мух или яйцекладущих черепах и пингвинов?
Со второй половины XX в. стали популярными количественные методы изучения репродуктивной биологии, выражающиеся в подсчетах “энергетики размножения” и “инвестирования в потомство” (см., например, обзор: Касьянов, 1989; а также: Wourms, 1994; Webb et al., 2006). Разумеется, такие расчеты возможны лишь для единичных, наиболее массовых и хорошо изученных видов, но никак не для высших таксонов и крупных филогенетических линий. На мой взгляд, совокупная ценность таких изысканий для понимания эволюции репродуктивной сферы крайне незначительна, поскольку они никак не объясняют (и не могут в принципе объяснить), почему одному виду (роду, семейству и т.д.) нужно больше “инвестировать” в потомство, а другому, близкородственному и живущему в таких же экологических условиях, – меньше. Кроме того, “энергетические затраты” на размножение сильнейшим образом варьируют индивидуально, в зависимости от возраста, пола, занимаемой части ареала и т.д. (см. Касьянов, 1989, с. 107–114).
Доминирующей по сей день в научной, учебной и популярной литературе остается идея о том, что живорождение эволюционно выигрышно, высоко адаптивно и достигает наивысшего своего развития у плацентарных млекопитающих (см., например, Иванова-Казас, 1995, с. 294; Батыгина и др., 2006; Wourms, 1994; Ostrovsky et al., 2016). По сути дела, это все та же, 2400-летней давности идея Аристотеля о “совершенных” животных, рождающих себе подобных. Но Аристотель не мог знать того, что прекрасно известно современным биологам, а именно то, что млекопитающие – это очень уязвимая по отношению к внешним факторам среды и очень скромная по числу видов группа (всего лишь около 6000 в современной мировой фауне и около 15 000 уже вымерших за предыдущие эпохи), небольшая даже в сравнении с другими позвоночными, коих известно многие десятки тысяч современных видов, и подавляющее их большинство – яйцекладущие. Если же приблизительно оценить общее число живородящих (и яйцеживородящих) видов животных, то это составит, в лучшем случае, лишь несколько процентов от более чем полутора миллиона всех описанных на планете видов Animalia (см., например, табл. 1 в статье Ostrovsky et al., 2016 и мой обзор ниже). При этом ежегодный существенный прирост числа открываемых в природе новых для науки видов происходит в основном за счет яйцекладущих беспозвоночных (прежде всего, насекомых), а небольшое увеличение числа новых видов позвоночных – за счет преимущественно яйцекладущих костных рыб. Общеизвестно и показано на множестве примеров, что целые научные направления тысячелетиями находились “в плену” у Аристотеля, не смея преодолеть его даже явно ошибочных умозаключений. Но все же такое положение дел было характерно, в основном, для средневековой и ренессансной науки, точечно сохранялось кое-где в науке XVIII–XIX вв. Кажется, что в современной литературе никто не обращал внимания на то, что широко распространенные ныне и противоречащие фактам взгляды на значение живорождения – это своего рода “ментальный реликт”, возможно, одно из последних заблуждений великого античного философа, дожившее в науке до наших дней.
Столь затянувшееся отсутствие прогресса в понимании эволюции живорождения, на мой взгляд, объясняется тем, что ее постоянно пытались изучать и объяснять не с начала филогенетического древа (с низших беспозвоночных), а с конца (с высших позвоночных). При этом обычно руководствовались данными, полученными на какой-либо одной из живородящих групп, без понимания того, что происходит во всех остальных группах. В этой связи очень важную позитивную роль в прояснении общей картины живорождения в природе сыграли многократно цитируемые в настоящей статье фундаментальные обобщения российских зоологов и эмбриологов – специалистов по беспозвоночным животным (в том числе самым архаичным) – О.М. Ивановой-Казас, А.Н. Островского и А.В. Ересковского.
Однако принципиально новый подход к пониманию эволюционного значения живорождения обозначился в работах французского энтомолога Жака Карайона (Jacques Carayon, 1916–1997), который впервые стал говорить о живорождении не как об эволюционном успехе, а как о побочном следствии партеногенеза и/или изменении места оплодотворения – с эктодермальных частей половой системы на глубинные, мезодермальные, вплоть до внутривителлярного оплодотворения (Carayon, 1961), что происходит, например, при так называемом травматическом оплодотворении (см. ниже). Действительно, яйцо может начать развиваться внутри тела матери лишь после того, как оно будет оплодотворено, или в том случае, если оплодотворение не требуется вовсе. И если это развитие начинается уже в гонадах, то, даже при обычной скорости прохождения яйца по половым путям, оно до откладки успеет пройти ряд стадий эмбрионального развития. Именно такая ситуация имеет место в случае многочисленных вариаций неполного яйцеживорождения. Некоторые другие авторы (см., например, Островский, 2009; Willey, 1898; Meier et al., 1999; Köhler et al., 2004; Blackburn, 1998, 2015 и др.) отмечали ту или иную корреляцию между живорождением и аберрациями в репродуктивной сфере отдельных групп животных, не развивая, однако, этого направления и не анализируя аналогичные связи в других живородящих группах.
Мною в ряде публикаций (Гаврилов, 2009; Gavrilov-Zimin, 2018, 2021) и в докторской диссертации (Гаврилов-Зимин, 2017), посвященных паранеоптерным насекомым (Paraneoptera), был предложен подход к живорождению как к реализации запасного пути эволюционного развития. Этот путь становится актуальным лишь в тех случаях, когда по разным причинам откладка яиц становится невозможной или плохо совместимой с новоприобретенными в ходе эволюции морфоанатомическими и/или физиологическими особенностями. Такие новоприобретения в разных группах могут быть самыми разными; общее у них лишь то, что они побочно нарушают нормальное функционирование доставшейся от предков репродуктивной системы. Наиболее частыми причинами перехода к живорождению являются различные варианты педогенеза и неотении, при которых размножающаяся личинка лишена имагинальных структур, ответственных за быстрое прохождение яиц по половым путям и их откладку во внешнюю среду (рис. 1). Цитогенетически педогенез и неотения обусловлены ларвальным мейозом (см. раздел Терминология), то есть образованием гамет до того, как сформировались имагинальные половые органы. Если при таком мейозе имагинальная стадия сохраняется и остается стадией размножения, а образовавшиеся в личинке яйцеклетки, не дождавшись оплодотворения, начинают развиваться партеногенетически, то яйцеживорождение становится неизбежным даже при наличии полностью развитых половых органов. С другой стороны, важно отметить, что педогенез и неотения не во всех случаях связаны с недоразвитием половых протоков, копулятивных и яйцекладных органов. Если эти протоки и органы имеются уже на личиночных стадиях (как, например, у аксолотлей, Trauth et al., 1994), то выпадение из онтогенеза взрослой стадии не препятствует нормальной яйцекладке. Во многих случаях причинами изменения репродуктивной стратегии оказываются также и не связанные с педогенезом простые утраты и/или резкие структурно-функциональные изменения в репродуктивной системе, побочно исключающие ее дальнейшее нормальное функционирование в режиме яйцекладки. Например, такая ситуация неизбежно возникает в связи с переходом от внешнего оплодотворения к внутреннему, в частности, при переходе первичноводных организмов к наземному образу жизни. Наружное оплодотворение и откладка незащищенных от высыхания и лишенных желтка (алецитальных) яйцеклеток во внешнюю среду становятся при этом невозможными (см. подробнее ниже) до тех пор, пока не появятся специализированные железы и половые протоки, обеспечивающие откладку яйца, наполнение его желтком и покрытие защитными оболочками.
Рис. 1.
Схема основных вариантов эволюционного перехода к живорождению у билатеральных животных (Bilateria).

Живорождение по очевидным объективным причинам сопряжено с сокращением числа потомков, соответствующим уменьшением интервала наследственной изменчивости и возможностей для естественного отбора. Для сравнения рассмотрим банальную эволюционную ситуацию, при которой из тысяч яиц, отложенных самкой, выживают лишь небольшое количество этих яиц или последующих стадий развития. В противоположность этому представим себе ситуацию, в которой из всего лишь двух–трех рожденных детенышей выживает один или даже все. Понятно, что в последнем случае изменчивость и отбор сведены к минимуму и неизбежная расплата за это – сниженное биологическое разнообразие, которое наблюдается во всех таксонах, “скатившихся на рельсы” живорождения (см. обзор ниже). Отдельные, легко объяснимые исключения, в частности, случаи, связанные с производством большого количества потомков некоторыми живородящими группами беспозвоночных животных, только подтверждают это правило и будут обсуждаться в конце статьи. Кроме того, само по себе обычное сокращение числа потомков при живорождении (без последующей родительской заботы) отнюдь не подразумевает однозначно бóльшую выживаемость этого потомства ни в относительном, ни, тем более, в абсолютном выражении в сравнении с родственными яйцекладущими группами. Живородящие же группы (а это, как правило, отдельные рода, реже семейства и очень редко целиком таксоны более высоких рангов) характеризуются относительно небольшим числом видов (максимум 6000) и, как правило (за редчайшим исключением), однообразной морфологией и внутренним строением. Чрезвычайно важным обстоятельством мне представляется также и то, что при живорождении действию естественного отбора по отношению к факторам внешней среды подвержены только постэмбриональные или даже только имагинальные стадии, а не целиком все стадии онтогенеза, как при нормальной яйцекладке. Таким образом, переход к живорождению сам по себе явно невыгоден с эволюционной точки зрения. При этом, разумеется, в силу мозаичности эволюции (гетеробатмии), регрессивные и неадаптивные признаки вполне могут сочетаться в организме с прогрессивными, обеспечивающими выживание группы. В настоящей статье нет возможности перечислять и обсуждать многочисленные случаи появления и сохранения неадаптивных признаков в эволюции организмов. Ограничусь лишь наиболее известными и очевидными примерами, непосредственно относящимися к обсуждаемой теме. Вряд ли кто-то станет настаивать на адаптивности таких свойств, как чрезвычайно болезненный родовой процесс у Homo sapiens, сопровождающийся частой смертью матери и ребенка, а также болезненные менструации у приматов и некоторых других млекопитающих. По крайней мере в отношении первого свойства хорошо известно, что оно является всего лишь побочным следствием перехода к прямохождению, изменения в строении таза и одновременного увеличения объема головного мозга (Washburn, 1960).
Попробуем в рамках обозначенного подхода проанализировать возникновение живорождения (и яйцеживорождения) в крупнейших филогенетических линиях многоклеточных организмов. При этом рассмотрении я намеренно буду избегать наиболее дискуссионных вопросов, касающихся филогенетического положения ряда мелких примитивных групп беспозвоночных животных, поскольку филогения этих групп и особенности их репродуктивной биологии не предоставляют каких-либо дополнительных аргументов или контраргументов для обсуждаемых теоретических закономерностей. В целом, я следую ниже классификационной схеме, используемой в одном из самых известных современных руководств по зоологии беспозвоночных – двухтомном издании под редакцией Вестхайде и Ригера (немецкий оригинал: Westheide, Rieger, 2004 и русский перевод: Зоология …, 2008). В отличие от более поздних статей (например, Dunn et al., 2014), так или иначе рассматривающих общий филогенез животных, это фундаментальное руководство отличается опорой преимущественно на легко проверяемые и хорошо изученные фенотипические признаки организмов. В то же время многочисленные “молекулярные” кладограммы представляют собой схемы, которые не могут быть мною проверены. Более того, “молекулярный” кладизм как методический подход базируется на недоказуемом и антиэволюционном догмате об одинаковой скорости накопления мутаций в одинаковых генах различных организмов (см., например, дискуссию у Клюге, 2020, с. 38–40). Для структурирования текста выделены названия параграфов, соответствующие основным этапам и направлениям эволюции животных.
Древние многоклеточные организмы. “Растительный” и “животный” пути эволюции способов размножения
Как хорошо известно, половой процесс исходно не был связан с размножением, а представлял собой просто обмен молекулами ДНК между клетками прокариот или же слияние двух гаплоидных клеток у одноклеточных эукариот. Для восстановления диплоидности возникает мейотическое деление, в результате которого образуются четыре дочерние клетки. Мейоз может происходить как непосредственно после образования диплоидной клетки (зиготы), так и существенно позднее, после многократных митотических делений. На основе исходной гаплоидной или диплоидной клетки могут возникать соответственно гаплоидные или диплоидные колонии одноклеточных организмов. Например, колонии вольвоксов (Volvox Linnaeus, 1758) состоят из гаплоидных клеток, а единственной диплоидной стадией жизненного цикла является зигота (Десницкий, 2021). Колониальные хоанофлагелляты (Choanoflagellata), которые обычно рассматриваются в качестве наиболее вероятных предков многоклеточных животных (Зоология …, 2008, с. 73), к сожалению, очень плохо изучены цитогенетически, однако смена гаплоидных и диплоидных стадий известна и у них (Levin, King, 2013). У настоящих многоклеточных организмов в жизненном цикле могут преобладать гаплоидная (гаметофит) или диплоидная (спорофит) стадия или же эти стадии могут равномерно чередоваться. Уже у колониальных одноклеточных существ помимо изогамии (равенства гамет) возникает гетерогамия (различие гамет по размеру) и оогамия, при которой крупная неподвижная женская гамета (яйцеклетка) сливается с мелкой подвижной мужской гаметой (сперматозоидом) (см., например, Levin, King, 2013; Nozaki et al., 2014) и, таким образом, появляется двуполое (бисексуальное) размножение. Оогамия характерна для многих групп водорослей, некоторых грибов, всех высших растений и всех многоклеточных животных (хотя в ряде групп животных мужские гаметы также утрачивают подвижность). Неподвижность яйцеклетки с самого начала эволюции многоклеточных организмов предопределяет “живорождение” как самый примитивный из возможных способов размножения (рис. 2). У водорослей и грибов, обладающих оогамией, а также у всех мхов, хвощей, плаунов и папоротников оплодотворенная яйцеклетка начинает делиться митотически и прорастает в спорофит непосредственно на материнском организме (гаметофите), а у семенных растений стадия гаметофита предельно редуцирована, и спорофит следующего поколения (семя) образуется на теле спорофита предыдущего поколения. Таков, в самом общем виде, “растительный” путь эволюции репродуктивной сферы, путь, на котором не возникло способов выведения яиц (яйцеклеток) во внешнюю среду. Увеличение числа потомков на этом пути достигается за счет производства либо многочисленных спор (споророждение), либо многочисленных семян (семярождение). При этом ни спора, ни семя растений не являются аналогами яйца животных, в связи с чем возникает терминологическая проблема применимости самого термина “живорождение” по отношению к растительным организмам (см. выше раздел Терминология).
Рис. 2.
Схема возникновения различных вариантов развития потомства на базе оогамии многоклеточных организмов.

В эволюции животных стадия гаметофита изначально отсутствует и продукты мейоза являются не спорами, а непосредственно гаметами (по типу оогамии). Выведение неподвижных оплодотворенных яйцеклеток во внешнюю, по отношению к организму, среду, для колониальных одноклеточных предков животных возможно только при распаде колонии (см., например, Беклемишев, 1964, с. 328). Если отказаться от распада колонии (что необходимо для возникновения многоклеточности), то нужно “научиться” выводить тем или иным способом половые продукты наружу, причем массово и синхронно сперматозоиды и яйцеклетки. Очевидно на начальных этапах эволюции Metazoa добиться такой синхронности (например, за счет появления феромонов и мускулатуры) удалось отнюдь не всем примитивным группам организмов. Так, среди губок (Porifera) большинство изученных видов характеризуются внутренним оплодотворением с выходом спермиев в воду и их последующим проникновением внутрь тела соседних губок, где тем или иным способом происходит слияние мужских и женских гамет. Дальнейшее развитие яиц, в силу отсутствия каких-либо половых органов или половых протоков, неизбежно осуществляется либо по типу яйцеживорождения или даже по типу примитивного плацентарного живорождения с образованием специальных эмбриональных капсул (Иванова-Казас, 1975; Ересковский, 2005; Батыгина и др., 2006; Riesgo et al., 2014; Ostrovsky et al., 2016). При этом, согласно недавним филогенетическим реконструкциям (Riesgo et al., 2014), наиболее древними из ныне живущих губок являются именно живородящие и яйцеживородящие, а все известные виды “яйцекладущих” губок произошли позднее от живородящих предков, когда, наконец, “научились” синхронно выбрасывать во внешнюю среду как мужские, так и женские гаметы и кроме того стали вырабатывать защитную коллагеновую оболочку яйца. Такая гипотеза в целом согласуется с гораздо более ранними идеями о том, что яйцерождение (яйцекладка) возникало в разных группах эволюционно “продвинутых” Demospongiae многократно и независимо (см. дискуссию у Ересковского, 2005, с. 55–59). В этих случаях выведение неподвижных яйцеклеток (часто вместе с окружающем слоем материнских клеток) технически обеспечивается за счет совершенствования регуляции потока воды, проходящего через тело, организованное по прогрессивному типу “лейкон”. Подобные переходы (или возвраты) к нормальному, яйцекладному способу отрождения потомства внутри живородящих таксонов разных рангов будут в дальнейшем повторяться в истории Metazoa неоднократно (см. ниже).
Сходная эволюционная судьба постигла представителей и других примитивно организованных многоклеточных – ортонектид (Orthonectida), дициемид (Dicyemidae) и пластинчатых (Placozoa), не имеющих специализированных гонад и половых протоков. Оплодотворенные яйцеклетки этих существ развиваются непосредственно внутри материнского организма (Иванова-Казас, 1975; Зоология …, 2008, с. 131–139; Eitel et al., 2011). Однако не вполне ясное до сих пор филогенетическое положение всех этих групп не позволяет судить о том, является ли их крайне примитивное строение (в том числе строение репродуктивной системы) исходной характеристикой или же вторичным упрощением. В любом случае, первичное отсутствие половых органов или же их вторичная утрата у указанных групп полностью укладываются в предлагаемую здесь теорию возникновения живорождения.
Иной путь эволюции репродуктивной сферы был реализован кишечнополостными (Coelenterata), составляющими парафилетический таксон, от которого так или иначе произошли все остальные более высокоорганизованные, билатеральные животные (Bilateria) (см. Малахов, 2004; Зоология …, 2008, с. 141). У кишечнополостных появляются примитивные гонады и мускулатура, в частности мускульный кишечник/желудок, через который выводятся наружу половые продукты. У некоторых гребневиков (Ctenophora) появляются даже специализированные половые протоки и семяприемники (Беклемишев, 1964, с. 334). В результате, как среди стрекающих кишечнополостных (Cnidaria), так и среди гребневиков (Ctenophora) у абсолютного большинства видов преобладает наружное оплодотворение с развитием яиц во внешней среде (в воде), и лишь для немногих видов характерно живорождение или яйцеживорождение в специальных выводковых сумках (Беклемишев, 1964; Иванова-Казас, 1975; Ostrovsky et al., 2016).
Билатеральные животные (Bilateria), кроме членистоногих и вторичноротых
Самыми примитивными билатеральными животными считаются плоские черви (Plathelminthes), большинство которых составляют паразитические организмы и лишь около четверти видов относятся к парафилетической группе свободноживущих “ресничных” червей (Turbellaria), которые, несомненно, более архаичны (Зоология …, 2008, с. 230–231). Женская половая система у различных Turbellaria проделывает очень значительную эволюцию. У примитивных ксенотурбеллярий (Xenoturbellida) нет еще никаких половых органов, ни мужских, ни женских. Оогонии и сперматогонии диффузно расположены в периферической паренхиме, а оплодотворенные яйца начинают развиваться внутри тела материнской особи и затем выводятся наружу через рот (Беклeмишев, 1964, с. 334; Westblad, 1949; Israelsson, Budd, 2005). У бескишечных турбеллярий (Acoela) наблюдаются все этапы эволюционного перехода от диффузно расположенных оогониев до парных компактных яичников, но специализированные женские половые протоки еще отсутствуют и яйца выводятся либо через рот, либо через разрыв наружного эпителия. У более высокоразвитых турбеллярий (Rhabditophora) развиваются сложные специализированные протоки для выведения яиц, и появляется разделение гонады на гермарий и вителлярий (Беклемишев, 1964, с. 335–343). В результате такой эволюции большинство свободноживущих плоских червей являются яйцекладущими организмами. Живорождение и яйцеживорождение встречается, главным образом, у паразитических плоских червей Digenea, Monogenea и Cestoda (Иванова-Казас, 1975; Ostrovsky et al., 2016). У дигенетических сосальщиков (Digenea) в сложном жизненном цикле чередуются яйцекладущее поколение (мариты), достигающее полного морфологического развития, и педогенетические живородящие поколения (спороцисты и редии), у которых отсутствуют органы имагинальной половой системы (Иванова-Казас, 1975, 1997). У моногеней (Monogenea) и цестод (Cestoda) спорадические случаи (яйце)живорождения, очевидно, связаны с существенными редукциями и аберрациями женской половой системы, как, например, у хорошо изученных моногенетических сосальщиков рода Gyrodactylus Nordmann, 1832 (см. обзор у Ивановой-Казас, 1975, с. 249–252).
У немертин (Nemertea) примеры живородящих и яйцеживородящих видов единичны (Ostrov-sky et al., 2016) и связаны с переходами к внутреннему оплодотворению, а среди гнатостомулид (Gnathostomulida), сипункулид (Sipunculida) и эхиурид (Echiurida), насколько мне известно, таких видов нет вовсе.
В огромном по числу современных видов (более 80 000) и чрезвычайно разнообразном во всех отношениях типе моллюсков (Mollusca) отдельные яйцеживородящие (облигатно и факультативно) рода и виды встречаются лишь как исключения (Tompa, 1979; Köhler et al., 2004). Важно отметить, что последние цитируемые авторы связывают возникновение яйцеживорождения (по крайней мере у специально изученных ими пресноводных гастропод) с утратой личиночной стадии развития, соответствующим увеличением размера яиц и неизбежным уменьшением их количества. Эта корреляция, как будет продемонстрировано ниже, прослеживается и во многих других группах животных со сложно устроенной половой системой. Подавляющее же большинство моллюсков являются яйцекладущими организмами, причем производство выметываемых в воду яйцеклеток достигает у многих видов гигантских значений – десятков и даже сотен миллионов яиц (Касьянов, 1989, с. 106).
Среди первичнополостных червей (Nemathelminthes), возможно представляющих собой полифилетическую группу и традиционно объединяющих гастротрих (Gastrotricha), нематод (Nematoda), волосатиков (Nematomorpha), коловраток (Rotatoria), скребней (Acanthocephala), приапулид (Priapulida), лорицифер (Loriccifera) и киноринх (Kinorhyncha), примеры живорождения встречаются как редкие исключения у отдельных родов и видов нематод, лорицифер, гастротрих и коловраток (Иванова-Казас, 1975; Ostrovsky et al., 2016), тогда как у скребней, наоборот, живорождение является общим правилом. Эта последняя небольшая (около 1400 видов), морфологически однообразная, группа полностью паразитических организмов характеризуется уникальной аномалией женской репродуктивной системы. Яичники еще на личиночной стадии распадаются на “яйцевые комки”, которые затем плавают в брюшном мешке или в полости тела. Вход в матку начинается так называемым маточным колоколом, который, ритмично сокращаясь, пропускает через узкий просвет только яйца веретеновидной формы, содержащие полностью развитых личинок, тогда как яйцевые комки и яйца, находящиеся на более ранних стадиях развития, обладают округлой формой и не могут пройти через отверстие маточного колокола (Зоология …, 2008, с. 750–751). Ясно, что при столь аберрантном строении и функционировании половой системы нормальная откладка яиц полностью исключена.
Среди тентакулят (Tentaculata) живорождение и яйцеживорождение обнаружено у многих видов мшанок (Bryozoa), тогда как форониды (Phoronida) и плеченогие (Brachiopoda) характеризуются откладкой неэмбрионизированных яиц во внешнюю среду, хотя встречаются и отдельные случаи вынашивания эмбрионов в мантийной полости или в лофофоре (Батыгина и др., 2006; Ostrovsky et al., 2016). Живорождение (и яйцеживорождение) у мшанок довольно хорошо изучено и известно во всех трех классах (Островский, 2009; Ostrovsky et al., 2016). Базовая причина такой “склонности” мшанок к живорождению, по моему мнению, заключается в их радикальном метаморфозе, “самом разрушительном, какой только известен у животных, так как от личинки (оозооида) остаются лишь стенки тела, на основе которых путем почкования развивается новое поколение” (Иванова-Казас, 1977, с. 239). У Вестхайде и Ригера (Зоология…, 2008, с. 356) это почкование даже названо “неотеническим”, несмотря на всю спорность применения этого термина к бесполому размножению. Вследствие такого метаморфоза у взрослых мшанок отсутствуют целые системы органов, которые имеются у взрослых особей более архаичных тентакулят – форонид и плеченогих. В частности, обе эти группы характеризуются наличием метанефридиев, через которые осуществляется вывод гамет в наружную среду. У мшанок нефридии утрачены, в результате чего они по характеру организации своей репродуктивной сферы фактически возвращаются на уровень примитивных Metazoa, со всеми вытекающими отсюда последствиями (см. выше). При этом важно отметить, что более архаичные классы современных мшанок, Phylactolaemata и Stenolaemata (см., например, обсуждение филогении у Taylor, Waeschenbach, 2015, а также Ostrovsky et al., 2016), представлен исключительно живородящими организмами, тогда как яйцекладущие виды встречаются в более “продвинутом” классе Gymnolaemata; выход женских гамет при этом происходит через “целомическую пору”, которая, вероятно, гомологична утраченному нефридию.
Напротив, у камптозой (Kamptozoa или Entoprocta) – небольшой группы, сходной с мшанками по своей морфологии и образу жизни, гонады обладают половыми протоками, через которые яйца выводятся во внешнюю среду – в полость атриума и там прикрепляются к его стенке или даже развиваются в специальных “выводковых карманах” (Зоология …, 2008, с. 354; Nielsen, 1971). Таким образом, у большинства камптозой речь идет о вынашивании потомства в сумках, сходных с марзупиями некоторых других животных (см. ниже), а не о живорождении. Единичные сообщения о развитии яиц непосредственно внутри яичника нескольких видов камптозой (Иванова-Казас, 1977, с. 270) нуждаются в дополнительных исследованиях.
Кольчатые черви (Annelida) начали свой эволюционный путь с выбрасывания гамет в воду и наружного оплодотворения; это способ размножения сохраняется по сей день у большинства многощетинковых червей (Polychaeta), причем самки некоторых видов выметывают сотни тысяч яйцеклеток (Иванова-Казас, 1977, с. 23; Зоология …, 2008, с. 387). Малощетинковые или поясковые черви (Clitellata) характеризуются существенно более сложной половой системой и относительно небольшим количеством откладываемых крупных яиц, что вероятно связано с утратой личиночной стадии в онтогенезе. Примеры живородящих и яйцеживородящих видов среди Polychaeta и Clitellata единичны и встречаются спорадически в разных семействах (Иванова-Казас, 1977; Ostrovsky et al., 2016).
Членистоногие (Arthropoda), кроме насекомых
Крупнейшая группа живых организмов на нашей планете – членистоногие (Arthropoda s.l.), насчитывающая более 1 000 000 видов и составляющая более ¾ всего разнообразия известных животных (а по многим оценкам до 10 млн и 90%), убедительнейшим образом демонстрирует огромное эволюционное преимущество яйцекладного способа размножения над всеми иными. Живородящие (яйцеживородящие) группы среди членистоногих составляют редкие исключения и суммарно объединяют не более 1% их видового разнообразия.
Так, маленькая (около 200 видов) архаичная группа членистоногих – онихофоры (Onychophora) объединяет преимущественно яйцеживородящие и живородящие виды, тогда как немногие примеры современных яйцекладущих видов рассматриваются как возникшие вторично от живородящих предков (Mayer et al., 2015; Treffkorn et al., 2019). Этот современный подход полностью согласуется со взглядами эмбриологов XIX в. (см. дискуссию у Willey, 1898, p. 34–35), которые предполагали, что живорождение у онихофор возникло в связи с переходом их предков к наземному образу жизни, при котором наружное оплодотворение и откладка незащищенных от высыхания алецитальных яйцеклеток в внешнюю среду оказались невозможны, а личиночная стадия утрачена. В результате эмбриональное развитие стало вынужденно проходить с задержкой внутри половых путей материнского организма. По мере адаптации к сухопутному образу жизни в ряде филогенетических линий онихофор произошел переход к откладке богатых желтком яиц, покрытых прочной скорлупой, защищающей от высыхания, и даже появился яйцеклад. Со своей стороны, я могу добавить, что, вероятно, переход к наземному образу жизни случился у онихофор относительно недавно, поскольку почти все известные вымершие онихофоры – морские животные, а все современные онихофоры обитают исключительно во влажных тропических/субтропических условиях и ведут ночной образ жизни. О все еще слабой приспособленности к жизни на суше свидетельствуют также чрезвычайно тонкая кутикула и открытые отверстия трахей. В свете всего сказанного логично предположить, что прогрессивная эволюция наземных онихофор идет сейчас и будет идти в дальнейшем именно по пути отказа от живорождения в пользу полноценной “сухопутной” яйцекладки.
У тихоходок (Tardigrada), еще одной небольшой группы примитивных членистоногих, живородящих или яйцеживородящих видов не известно вовсе.
Среди хелицеровых (Chelicerata), насчитывающих более 100 000 современных видов, подавляющее большинство (98%) являются яйцекладущими. Так, мечехвосты (Xiphosura), морские пауки (Pantopoda), сухопутные пауки (Araneae), телифоны (Uropygi), фрины (Amblypygi), сенокосцы (Opiliones), сольпуги (Solifugae) и рицинулеи (Ricinulei) – полностью яйцекладущие группы. Единичные примеры яйцеживорождения встречаются у клещей (Acari) (Иванова-Казас, 1979; Ostrovsky et al., 2016). Псевдоскорпионы (Pseudo-scorpiones) вынашивают отложенные яйца в наружной выводковой сумке, секретируемой железами половой камеры, то есть не являются живородящими, вопреки расхожему мнению. Настоящее живорождение и яйцеживорождение характерны лишь для настоящих скорпионов (Scorpiones) (см., например, Warburg, 2012), которые насчитывают около 2400 современных видов и считаются самыми архаичными из всех наземных хелицеровых (Зоология …, 2008, с. 475). Вероятно, (яйце)живорождение скорпионов, как и ряда других групп исходно водных животных, возникло при выходе на сушу, вследствие невозможности откладки алецитальных яйцеклеток во внешнюю среду, перехода к внутреннему оплодотворению и утраты личиночной стадии.
Подавляющее большинство ракообразных (Crustacea, более 70 000 современных видов) относятся к яйцекладущим животным, хотя нередко встречается вынашивание потомства в выводковых сумках различного строения и иные формы заботы о потомстве. Примеры (яйце)живородящих ракоообразных единичны (Klapow, 1970). В то же время, яйцеживорождение характерно для пятиусток (Pentastomida) – маленькой (около 100 видов) группы паразитических организмов, гипотетически относящихся к ракообразным (Зоология …, 2008, с. 548). Однако сильнейшая редукция систем органов у этих животных не позволяет в настоящее время однозначно судить об их происхождении и родственных связях, а соответственно и о путях возникновения их яйцеживорождения.
У многоножек (Myriapoda) каких-либо примеров живорождения или яйцеживорождения неизвестно.
Насекомые (Insecta)
Среди насекомых (Insecta sensu Hexapoda), как и среди беспозвоночных в целом, общее число живородящих и яйцеживородящих видов не превышает 1%. При этом первичнобескрылые насекомые (Apterygota) – предковая (парафилетическая) группа по отношению ко всем остальным (крылатым) насекомым, демонстрируют исключительно яйцекладущий способ размножения. Это важное обстоятельство, а также и то, что гипотетические предки насекомых – ракообразные и многоножки, в ходе своей адаптации к жизни на суше, сумели избежать живородящего пути развития, позволяет уверенно говорить о первичности именно яйцекладности у всего класса Insecta. Очевидно, такое благоприятное развитие событий было связано с тем, что внутреннее оплодотворение и развитые половые протоки появились уже у полностью водных ракообразных, многие из которых к тому же откладывают устойчивые к высыханию яйца, покрытые плотной оболочкой (Зоология …, 2008, с. 516–599). Дальнейшая эволюция репродуктивной сферы насекомых проходила по пути постоянного усложнения внешних и внутренних половых органов, препятствующих преждевременному оплодотворению яиц и их развитию внутри половых путей самки. Разделение овариол на гермарий и вителлярий, появление амниона и сложных наружных оболочек яйца, развитие разнообразных придаточных желез в яйцеводах, сложно устроенные сперматеки, чрезвычайно разнообразные и затейливо устроенные копулятивные аппараты, а также яйцеклады – все эти структурные новообразования необходимы для того, чтобы откладывать во внешнюю среду большое количество защищенных, богатых желтком яиц до начала их эмбрионального развития. Все изученные случаи живорождения (яйцеживорождения) насекомых сопряжены со значительными аберрациями половой системы, как правило, обусловленными ларвальным мейозом, педогенезом, неотенией, травматическим оплодотворением и другими отклонениями.
Так, среди древнекрылых насекомых (Palaeoptera) живорождение неизвестно, а яйцеживорождение достоверно обнаружено лишь у некоторых видов поденок (Ephemeroptera) из трех родов сем. Baetidae: Callibaetis Eaton, 1881, Cloeon Leach, 1815, Procloeon Matsumura, 1931 (Kluge, 2016) и одного вида из сем. Leptophlebiidae – Thraulodes viviparus Kluge, 2020. Примечательно, что специальные (в том числе экспериментальные) исследования репродуктивной биологии отдельных видов яйцеживородящих поденок показали факультативный характер их яйцеживорождения, коррелирующий с аномальным соотношением полов (8 самок на 1 самца) в популяциях (Edmunds, 1945) и факультативным партеногенезом (Harker, 1997). Более ранние работы, так же указывающие на факультативный характер яйцеживорождения поденок, анализируются у Хейгана (Hagan, 1951, p. 69–71).
У полинеоптерных насекомых (Polyneoptera) яйцеживородящие рода известны у тараканообразных (Pandictyoptera), а живородящие – у уховерток (Dermaptera). Современные Pandictyoptera демонстрируют уникальную апоморфию – их яйцеклад сильно модифицирован и скрыт во внешней генитальной камере, образованной увеличенным VII стернитом брюшка, вследствие чего откладываемые яйца оказываются заключенными в специальную белковую оболочку – оотеку (Клюге, 2020, с. 435, 438). Однако у древних вымерших тараканов (Palaeoblattariae) имелся нормальный яйцеклад и соответственно нормальная яйцекладка, без оотеки (Клюге, 2020, с. 435, 437). Вероятно, радикальная перестройка яйцекладного аппарата на определенном этапе эволюции тараканов привела к тому, что они не смогли откладывать яйца поштучно, и последние стали задерживаться в половых протоках самки. Выходом из этой ситуации были либо 1) переход к яйцеживорождению, реализовавшийся в ряде родов тараканов, у которых оотека остается внутри генитальной камеры, либо 2) откладка целой оотеки во внешнюю среду, как это и происходит у большинства современных тараканов (Neoblattariae), у всех богомолов (Raptoriae) и архаичных термитов (Hemiclidoptera). Весьма примечательно, что у многих тараканов оотека выводится наружу, но еще остается прикрепленной к гениталиям самки или же вынашивается в специальной выводковой сумке (марзупии) (Roth, Willis, 1957), а у большинства современных термитов (Cryptoclidoptera) оотека вовсе исчезает, и происходит восстановление нормальной яйцекладки (Клюге, 2020, с. 455–456).
У большинства современных уховерток (Dermaptera или Dermatoptera) яйцеклад частично или полностью утрачен, а число откладываемых яиц сравнительно невелико (20–80 штук). Вероятно, с этим связана хорошо известная забота уховерток о потомстве и переход в нескольких родах (Arixenia Jordan, 1909, Xeniaria Maar, 1974, Hemimerus Walker, 1871 и Araeomerus Maar, 1974) к плацентарному живорождению. При этом все четыре упомянутых рода демонстрируют явно педоморфные черты строения имаго, которое является пятой постэмбриональной стадией (Nakata, Maa, 1974), в отличие от остальных уховерток, у которых имаго соответствует шестой или седьмой стадии (то есть с 5–6 предшествующими личиночными стадиями). Малоизученные пока случаи яйцеживорождения сообщались также для двух видов уховерток из родов Marava Burr, 1911 и Chaetospania Karsch, 1886 (см. Kocarek, 2009).
Относительно большое число живородящих (и яйцеживородящих) видов насекомых известно среди паранеоптерных отрядов (Paraneoptera) (см. Gavrilov-Zimin, 2021). Так, различные варианты яйцеживорождения и плацентарного живорождения наблюдаются у самых архаичных паранеоптерных насекомых – некоторых сеноедов (Copeognatha) из сем. Archipsocidae, Trogiidae и Pseudocaeciliidae (Fernando, 1934; Jentsch, 1936; Mockford, 1957; Wong, Thornton, 1968). По меньшей мере некоторые из паразитических потомков сеноедов – пухоеды (Mallophaga) из рода Meinertzhageniella Eichler, 1940 и вши (Siphunculata) Polyplax serrata (Burmeister, 1839) и Hoplopleura sp. демонстрируют яйцеживорождение (Eichler 1946; Golub, Nokkala, 2004). Однако общая картина живорождения/яйцеживорождения у архаичных Paraneoptera остается пока не ясной из-за слабой изученности репродуктивной биологии большинства видов.
Среди трипсов (Thysanoptera) яйцеживородящие виды известны в подотряде Tubulifera, характеризующемся утратой яйцеклада (Bagnell, 1921; John, 1923; Hood, 1934, p. 71; Hathaway, 1938; Bournier, 1966). У некоторых видов доказано факультативное и неполное яйцеживорождение с откладкой яиц на разных стадиях эмбриогенеза (Viswanathan, Ananthakrishnan, 1973; Ananthakrisnan, Dhileepan, 1984; Dhileepan, Ananthakrisnan, 1987; Nagrale, 2012).
Ни одного живородящего или яйцеживородящего вида не выявлено до сих пор у трех из пяти подотрядов равнокрылых хоботных насекомых (Homoptera) – цикадовых (Cicadinea), псиллид (Psyllinea) и алейродид (Aleyrodinea), которые суммарно насчитывают более 50 000 рецентных видов. Два других подотряда, тли (Aphidinea) и кокциды (Coccinea), наоборот, демонстрируют многочисленные и разнообразные примеры задержки развивающихся яиц внутри половых путей самки. Архаичные тли – адельгиды и филлоксеры (Adelgidae и Phylloxeridae), насчитывающие всего лишь 140 современных видов, характеризуются наличием яйцеклада и облигатной яйцекладкой. Все остальные тли (около 5000 видов) утратили яйцеклад и производят потомство либо исключительно путем плацентарного партеногенетического живорождения (реже яйцеживорождения), либо циклически чередуя такое размножение с обоеполым размножением и яйцекладкой (Hille Ris Lambers, 1950; Blackman, 1987). Уникальность живорождения у тлей обусловлена ларвальным мейозом и чрезвычайно ранним педогенетическим развитием яйца в теле еще не рожденной личинки, находящейся в половых путях самки предыдущего поколения. Такие яйца имеют очень мелкие размеры, лишены желтка и хориона и проходят полное эмбриональное развитие внутри вителлярия, получая питание от клеток фолликулярного эпителия (Uichanco, 1924; Hagan, 1951; Blackman, 1987).
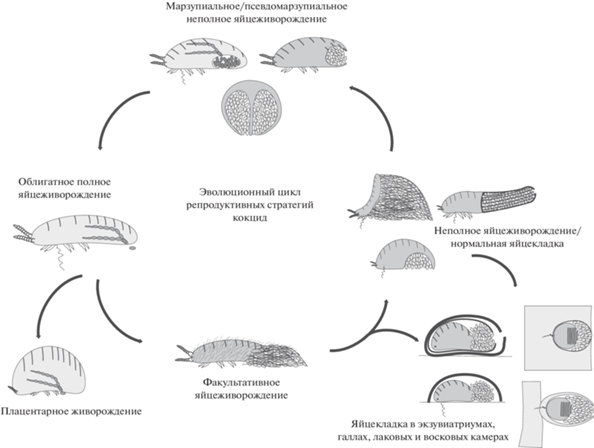
Наибольшее разнообразие вариантов яйцеживорождения и живорождения среди паранеоптерных насекомых демонстрируют кокциды (Coccinea) (Gavrilov-Zimin 2018, 2021) (рис. 3). Яйцеживорождение, по-видимому, является апоморфией этой группы, напрямую связанной с ларвальным мейозом и неотенией, имеющей место у всех без исключения самок современных кокцид. Среди наиболее архаичных современных кокцид (надсемейство Orthezioidea) имеются рода и целые трибы, характеризующиеся облигатным яйцеживорождением, у других родов яйцеживорождение носит факультативный характер, у третьих на базе неполного яйцеживорождения возникает марзупиальность, у четвертых происходит возврат к более или менее полноценной яйцекладке. Вторичность такой откладки яиц подтверждается тем, что она сопряжена с появлением оригинальных новообразований – специальных восковых или лаковых яйцевых “мешков” и камер, вырабатываемых многочисленными и чрезвычайно разнообразными воскоотделяющими и лаковыми железами, а также марзупиальных полостей (рис. 4). Возникновение всей неококцидной филогенетической линии (надсемейство Coccoidea) связано с облигатным полным яйцеживорождением (Gavrilov-Zimin, Danzig, 2012; Danzig, Gavrilov-Zimin, 2014; Gavrilov-Zimin, 2018). Эта особенность была, вероятно, унаследована мучнистыми червецами (сем. Pseudococcidae), наиболее архаичной группой среди неококцид, от облигатно яйцеживородящих вымерших предков из сем. Phenacoleachiidae, представленного в современной фауне единственным видом. К настоящему времени облигатное яйцеживорождение обнаружено у более 500 видов из более, чем 60 родов мучнистых червецов, что составляет около 25% их мирового разнообразия. Это число, несомненно, значительно увеличится в дальнейшем, поскольку у подавляющего большинства видов псевдококцид (и кокцид в целом) репродуктивная биология до сих пор не изучена. Многочисленные яйцеживородящие виды известны также и в других неококцидных семействах: Eriococcidae, Micrococcidae, Coccidae, Aclerdidae, Dactylopiidae, Keriidae, Stictococcidae, Asterolecaniidae s.l., Beesoniidae и Diaspididae (см. списки у: Gavrilov-Zimin 2018, p. 216–244). Плацентарное живорождение известно пока только в трех родах неококцид, Apiomorpha Rübsaamen, 1894 (сем. Eriococcidae), Stictococcus Cockerell, 1903 и Parastictococcus Richard, 1971 (оба из сем. Stictococcidae) (Buchner, 1957, 1963, 1965).
Рис. 3.
Живорождение, яйцеживорождение и яйцекладка у кокцид (Insecta: Coccinea) (по: Gavrilov-Zimin, 2018).
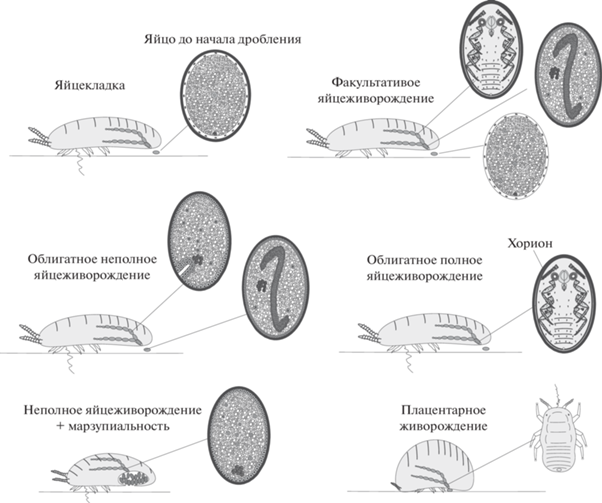
Рис. 4.
Эволюционный цикл репродуктивных стратегий кокцид (Insecta: Coccinea) (по: Gavrilov-Zimin, 2018).
В крупном (около 45 000 видов) отряде клопов (Heteroptera) немногочисленные примеры живорождения и яйцеживорождения известны в семействах Polyctenidae, Cimicidae, Anthocoridae, Plokiophilidae, Microphysidae, Aradidae и Lygaeidae (Hagan, 1931, 1951; Carayon, 1956, 1961; Cobben, 1968). У наиболее изученного из живородящих клопов, поликтениды Hesperoctenes fumarius (Westwood, 1874), овуляция, оплодотворение и начальные этапы развития эмбриона проходят еще на личиночной стадии (Hagan, 1931, p. 38, 1951, p. 396), то есть являются следствием ларвального мейоза. Согласно исследованиям Карайона (Carayon, 1961, 1966, p. 179), для многих живородящих и яйцеживородящих клопов характерно своеобразное травматическое оплодотворение – самец протыкает копулятивным органом покровы тела самки и впрыскивает сперму за пределы женских половых протоков. Последующее оплодотворение связано со структурами своеобразной “парагенитальной системы”, в результате работы которой оплодотворение происходит в овариолах, а не в эктодермальных частях половой системы, как у яйцекладущих клопов.
Олигонеоптерные насекомые (Oligoneoptera) объединяют крупнейшие по числу видов отряды живых организмов: перепончатокрылые (Hymenoptera, более 150 000 видов), жесткокрылые (Coleoptera, более 400 000 видов), чешуекрылые (Lepidoptera, более 180 000 видов), двукрылые (Diptera, более 150 000 видов), а также несколько малочисленных отрядов. Единственными указаниями на яйцеживорождение у перепончатокрылых остаются старые сообщения о таком способе отрождения потомства у двух родов наездников из сем. Ichneumonidae (см. обзор: Hagan, 1951, p. 109–110). Среди жуков отдельные яйцеживородящие виды и рода встречаются в сем. Micromalthidae, Staphylinidae, Chrysomelidae, Tenebrionidae, Carabidae, Cerambycidae (Захваткин, 1966, 1970; Hagan, 1951; Bontems, 1988; Iwan, 2000; Zilberman et al., 2019), у чешуекрылых – в сем. Psychidae, Tineidae, Glyphipterigidae, Oecophoridae, Coleophoridae, Cosmopterigidae, Gelechiidae, Pterophoridae, Papilionidae, Pieridae, Nymphalidae и Geometridae (см. обзор у: Lee, Li, 2018), у ручейников (Trichoptera) – в роде Triplectides Kolenati, 1859 (см. Morse, Neboiss, 1982). В настоящей статье нет возможности подробно останавливаться на рассмотрении репродуктивной специфики всех этих разрозненных и независимо возникших случаев отклонения от нормальной яйцекладки. Более пристального внимания заслуживают двукрылые насекомые (Diptera), у которых многочисленные живородящие и яйцеживородящие виды/рода известны в 22 семействах (Meier et al., 1999), а общая картина распространения и возникновения (яйце)живорождения, на первый взгляд, выглядит довольно запутанной. Во многом эта путаница связана с тем, что указанные авторы, как и многие другие, цитируемые ими диптерологи, не отличают живорождение от яйцеживорождения, отвергают терминологические подходы, разработанные на гораздо более широком сравнительном материале (например, Hagan, 1951; Sellier, 1955) и вводят собственные самобытные термины. В результате, Мейер с соавт. (Meier et al., 1999) указывают на 61 случай независимого возникновения живорождения у двукрылых, объединяя в этом числе как облигатно живородящие и яйцеживородящие линии, так и примеры факультативного яйцеживорождения. Однако почти все эти случаи (57 из 61) относятся к филогенетической линии Schizophora (так называемые щеленосные мухи), в которой произошло структурное изменение половых путей самки – удлинение и расширение непарного яйцевода. Это преобразование по непонятным причинам случилось еще у яйцекладущих предков Schizophora и, по мнению Мейера с соавт. (Meier et al., 1999), явилось “экзаптацией (exaptation)” к живорождению. У некоторых яйцеживородящих Schizophora в таком расширенном яйцеводе (матке) вынашиваются сотни и даже тысячи микроскопических яиц, и за счет этого сохраняется возможность для широкого естественного отбора потомства, в отличие от большинства других (яйце)живородящих групп организмов, но никакого питания от материнского организма такие яйца, естественно, не получают. Напротив, у самых известных живородящих мух, относящихся к роду Glossina Wiedemann, 1830 (мухи це-це), имеет место редукция яичника до единственной овариолы. Попеременно в левом и правом яичнике созревает по одному яйцу, которое оплодотворяется и проходит полное эмбриональное развитие внутри аномального расширенного непарного яйцевода. Выходящая из яйца личинка питается выделениями придаточных желез, протоки которых открываются в полость яйцевода (Hagan, 1951, p. 111–158). Только после второй линьки, личинка третьего возраста откладывается мухой в почву. В результате всех этих сложных репродуктивных “достижений”, за всю жизнь каждая муха це-це рождает не более 10 личинок (Vreysen et al., 2013), а все монотипное сем. Glossinidae отличается ничтожным биологическим разнообразием – всего лишь около 20 видов.
Среди прочих мух, не относящихся к Schizophora, примеры (яйце)живорождения единичны и обусловлены иными причинами. Например, гемоцельное живорождение личинок некоторых Cecidomyiidae напрямую связано с педогенезом (Hagan, 1951, с. 206–225); более того, педогенетическое живорождение было впервые открыто именно у этой группы животных Вагнером (1862).
С педогенезом или с неотенией связано также и гемоцельное живорождение у небольшой, родственной жукам, паразитической группы веерокрылых насекомых Strepsiptera (Hagan, 1951, p. 226–260), у которых яичники диссоциированы на свободно плавающие в гемоцеле яйцеклетки. Кроме того, для ряда представителей этой аберрантной группы насекомых характерно травматическое оплодотворение (Kathirithamby et al., 2015; Peinert at al., 2016), сходное с тем, которое известно для (яйце)живородящих клопов (см. выше).
Вторичноротые (Deuterostomia), кроме позвоночных
Большинство иглокожих (Echinodermata) относятся к яйцекладущим организмам, причем у некоторых видов число выметываемых самкой яиц достигает сотен миллионов (Зоология …, 2008, с. 837). Отдельные случаи (яйце)живорождения встречаются у морских звезд (Asteroida) и офиур (Ophiuroida) (Иванова-Казас, 1978а; Hendler, 1975; Ostrovsky et al., 2016), но изученность репродуктивной сферы соответствующих видов пока что крайне недостаточна. Более многочисленные и лучше изученные примеры живорождения и яйцеживорождения демонстрируют голотурии (Holothuroida) из разных семейств (Иванова-Казас, 1978а, с. 77; Hansen, 1968; Ostrovsky et al., 2016). Педоморфозу при этом отводится важнейшая роль в эволюции всего класса голотурий (Смирнов, 2014). Помимо этого общего обстоятельства, наиболее изученный из живородящих видов, Oneirophanta mutabilis Théel, 1879, характеризуется отсутствием самцов и облигатным партеногенезом (Hansen, 1968; Ramirez-Llodra et al., 2005), то есть обладает дополнительным отклонением от обычной бисексуальной репродуктивной стратегии.
Морские стрелки (Chaetognatha), бесчерепные (Acrania) и полухордовые (Hemichordata) откладывают яйца во внешнюю среду. Однако для колониальных крыложаберных (Pterobranchia), которые остаются очень слабо изученными в отношении репродуктивной биологии, характерно развитие эмбрионов внутри коллагеновых трубок, представляющих собой жилые домики (ценоции) родительских особей (Stebbing, 1970; Lester, 1988).
Среди низших хордовых (Chordata) яйцеживорождение и живорождение известно у ряда асцидий (Ascidiacea), огнетелок (Pyrosomida) и сальп (Salpida). Примечательно, что у большинства одиночных асцидий имеется наружное оплодотворение и, соответственно, нормальная яйцекладка с большим количеством выбрасываемых в воду яйцеклеток, хотя у некоторых видов встречается задержка яиц в клоаке или околожаберной полости, тогда как яйцеживорождение характерно для колониальных видов. Подобно колониальным тентакулятам (Tentaculata) (см. выше), колонии асцидий и огнетелок образуются путем своеобразного почкования, нередко с утратой половых протоков. В результате, яйца остаются внутри фолликула или же попадают в полость тела матери, где и проходит эмбриональное развитие; у сальп редукция яйцевода происходит уже после оплодотворения, и яйца развиваются внутри фолликулов по типу плацентарного живорождения (Иванова-Казас, 1978б, с. 24–26, 116).
Позвоночные (Vertebrata)
Наиболее архаичные позвоночные животные – бесчелюстные (Agnatha) характеризуются исключительно яйцекладущим способом размножения и внешним оплодотворением. Среди хрящевых рыб (Chondrichthyes), насчитывающих примерно 1100 современных видов, около 55% считаются живородящими или яйцеживородящими, тогда как среди костных рыб (Osteichthyes), составляющих подавляющее большинство современных рыб (около 28 000 видов), живорождение известно лишь у 2–3% видов (Wourms, 1994). Эти различия напрямую коррелируют с переходом от внешнего оплодотворения к внутреннему, который в эволюции рыб происходил неоднократно. Однако, все хрящевые рыбы характеризуются внутренним оплодотворением (Wourms, 1977, 1994), что, вероятно, свидетельствует о появлении этой особенности (и, соответственно, копулятивных органов самцов) у их общего предка. Как и в случае аналогичных переходов к внутреннему оплодотворению у беспозвоночных животных (см. выше), оплодотворение яйцеклетки внутри гонады не означает синхронного появления структур, ответственных за формирование оболочек яйца и последовательное выведение яиц во внешнюю среду. Пока и если таких структур нет, развивающиеся яйца неизбежно задерживаются внутри тела самки, что и наблюдается у 55% доживших до наших дней видов хрящевых рыб, часть из которых демонстрирует яйцеживорождение, а часть переходит к плацентарному живорождению. У остальных же 45% видов развиваются приспособления для откладывания защищенных яиц во внешнюю среду (в частности скорлуповая железа). То, что эти приспособления вторичны, доказывается их отсутствием у предкового таксона – бесчелюстных рыб, которые не обладают специализированными половыми протоками и характеризуются внешним оплодотворением. У костных рыб переходы к внутреннему оплодотворению происходили многократно (Wourms, 1977, 1994), чем, вероятно, и объясняется хаотичность появления среди них яйце(живородящих) видов и родов.
Среди ныне живущих амфибий (Lissamphibia) живородящие (и яйцеживородящие) виды составляют не более 1% и встречаются как отдельные исключения среди саламандр и лягушек, но доминируют среди червяг (Blackburn, 2015). Представляется вполне очевидным, что яйцеживорождение амфибий возникает на базе многократных эволюционных переходов от наружного оплодотворения к внутреннему и утраты личиночной стадии (Wake, 1993), а у саламандр дополнительно осложняется еще и факультативным партеногенезом (Buckley, 2012).
Возникновение наземных позвоночных – амниот (Amniota), как хорошо известно, связано с переходом к внутреннему оплодотворению, утратой самостоятельно питающейся личиночной стадии и появлением дополнительной эмбриональной оболочки (амниона). При этом живорождение или яйцеживорождение в современной фауне характерно для примерно 20% скваматных (Squamata) пресмыкающихся (Blackburn, 2015) и всех млекопитающих, но полностью отсутствует у черепах, гаттерий, крокодилов и птиц. Ранее неоднократно высказывались предположения о происхождении всех амниот непосредственно от неких яйцеживородящих амфибий (см. обзор у Ивановой-Казас, 1995, с. 293–294). Допуская такой ход событий и сообразуясь с современными представлениями о филогенезе позвоночных (например, Kardong, 2012), эволюцию репродуктивных стратегий амниот можно представить в виде следующей схемы (рис. 5). Эта схема предполагает многократный возврат от яйцеживорождения к яйцекладке в противовес гипотезе о многократном независимом возникновении (яйце)живорождения в разных группах амниот (см. об этом также в разделе “Простота, обратимость и продуктивность живорождения”). Независимо от того, какая из этих двух противоположных гипотез верна, отсутствие самостоятельно питающейся личиночной стадии диктует необходимость снабжения потомства питанием вплоть до достижения последним размеров, близких к размерам взрослого организма. Это означает либо длительное вынашивание потомства в теле матери (с последующим дополнительным выкармливанием), либо откладку крупных, богатых желтком яиц, часто тоже с необходимостью дополнительного выкармливания. Оба этих варианта означают резкое уменьшение количества потомков в сравнении с яйцекладущими первичноводными предками, и соответственно, оба эволюционно невыгодны в смысле биологического разнообразия, принятого в настоящей статье в качестве основного критерия оценки эволюционной “успешности” таксона. Относительное внешне-морфологическое разнообразие современных плацентарных амниот (млекопитающих) оказалось возможным только в уникальных условиях отсутствия конкуренции с крупными яйцекладущими амфибиями, рептилиями и крупными беспозвоночными, вымершими при изменении климата и снижении количества кислорода в атмосфере нашей планеты. Приведу здесь цитату из классического учебника по зоологии беспозвоночных (Наумов, Карташев, 1979, с. 165): “На протяжении двух третей своей геологической истории млекопитающие оставались мелкими, внешне напоминающими крыс существами и не играли заметной роли в природе”. По сей день “крысообразные” существа – грызуны (Rodentia) составляют около 40% видового разнообразия всех млекопитающих. Общеизвестно, что живорождение у млекопитающих сочетается с появлением волосяного покрова, теплокровностью, разделением кругов кровообращения, развитием коры головного мозга и др. ароморфозами, которые и обеспечивают существенные конкурентные преимущества. Однако даже при всех этих колоссальных эволюционных достижениях, млекопитающие почти в четыре раза проигрывают в биологическом разнообразии ближайшим к ним современным группам – рептилиям (которые преимущественно яйцекладущи), а также птицам, которые обладают сходными ароморфозами, но полностью яйцекладущи. По поводу тотального отсутствия живорождения (и яйцеживорождения) у птиц мне здесь хотелось бы обратить внимание читателя на то, что причиной такого отсутствия никак нельзя считать адаптации к полету, поскольку, во-первых, существует достаточно примеров нелетающих птиц, а во-вторых, живорождение (или яйцеживорождение) имеется у летучих мышей и некоторых прекрасно летающих насекомых, заметно превосходящих птиц по маневренности полета (cм. подробнее об этом у Blackburn, Evans, 1986). Более актуальным в рамках настоящей дискуссии мне представляется вопрос о том, почему живорождение сохраняется во всех таксонах плацентарных млекопитающих, если они в большинстве случаев все равно рождают “недоделанных” детенышей, которых необходимо дополнительно выкармливать длительное время? Казалось бы, в этой ситуации нет принципиальной разницы – вылупится ли детеныш из отложенного яйца или появится непосредственно из половых путей самки. Вероятно, объяснение заключается в том, что млекопитающие в силу своей физиологии оказались неспособны откладывать полноценные, обильные желтком “сухопутные” яйца, защищенные достаточно плотной оболочкой, в отличие от других групп сухопутных животных, таких, например, как птицы и членистоногие. В этой связи, интересными представляются обобщенные биохимические данные по эмбрионам амниот: у скваматных рептилий кальций, необходимый для построения эмбрионального скелета, локализуется в желтке, у черепах, крокодилов и птиц – в скорлупе, а у живородящих амниот резервуар для накопления кальция отсутствует вовсе (Kardong, 2012, с. 589). Яйца немногочисленных, доживших до наших дней яйцекладущих млекопитающих – однопроходных (Monotremata), как известно, не имеют кальцинированной оболочки и проходят часть своего эмбрионального развития в половых путях матери (Иванова-Казас, 1995, с. 289; Kardong, 2012, p. 173), что вполне соответствует неполному яйцеживорождению, аналогичному таковому у многих прочих обсуждавшихся выше групп животных. Примечательно также, что новорожденные детеныши однопроходных и сумчатых млекопитающих еще сохраняют очень мелкие размеры в сравнении с размерами взрослых особей, то есть близки по этому признаку к личинкам амфибий, но нуждаются в длительном выкармливании и защите со стороны матери. Напротив, водные рептилии (крокодилы и морские черепахи), вторично переходят к откладке большего количества более мелких яиц в сравнении со своими сухопутными предками, вероятно, по той причине, что в водной среде (в отличие от суши) их относительно мелкие личиночные стадии способны самостоятельно найти пропитание, как и личинки амфибий. Кроме того, даже если бы мелкие личинки амниот (длина тела новорожденных детенышей однопроходных и сумчатых составляет всего лишь несколько сантиметров) способны были самостоятельно питаться на суше, они не смогли бы конкурировать с наземными членистоногими, особенно насекомыми, которыми переполнены все сухопутные экологические ниши, пригодные для жизни животных в размерном классе от долей миллиметра до нескольких сантиметров (очень редко до 10–30 см) длины тела.

ПРОСТОТА, ОБРАТИМОСТЬ И ПРОДУКТИВНОСТЬ ЖИВОРОЖДЕНИЯ
Из приведенного выше обзора ясно, что различные аберрации репродуктивной сферы, изменяющие ее предковое состояние и приводящие к (яйце)живорождению можно разделить на три основные группы: 1) возникновение оогамии у древних многоклеточных организмов, при отсутствии каких-либо механизмов выведения неподвижной зиготы во внешнюю среду; 2) переходы от наружного оплодотворения к внутреннему, когда у организма еще отсутствуют специализированные половые протоки (яйцеводы) с придаточными железами и сперматеками; зачастую такой переход осложняется утратой самостоятельно питающейся личиночной стадии; 3) нарушения в хорошо развитой половой яйцекладной системе: утрата взрослых половых органов (вследствие педогенеза, неотении, педоморфоза), ларвальный мейоз, изменение места оплодотворения с эктодермальных частей половой системы на внутригонадное (например, при травматическом оплодотворении). Во всех случаях живорождение вовсе не является результатом какой-то сложной и длительной эволюции половой системы животных, а представляет собой всего лишь результат задержки и развития яйца внутри тела материнского организма с получением дополнительного питания или без такового. Для обеспечения зародыша питанием, даже в самых сложных случаях, не возникает каких-либо принципиально новых органов, а происходит лишь разрастание и модификация уже имеющихся структур зародыша и/или структур тела самки. Более того, пример низших многоклеточных животных доказывает, что для возникновения живорождения не нужно даже обладать самими половыми органами, ни внешними, ни внутренними. Напротив, все сложнейшие структуры женской половой системы высокоразвитых животных (обособленные и структурированные гонады, специализированные половые протоки, сперматеки и семяприемники, скорлуповые и прочие придаточные железы, яйцеклады), эмбриональные оболочки и накопление желтка в яйце необходимы для того, чтобы избежать живорождения и откладывать во внешнюю среду многочисленные и хорошо защищенные яйца. С эволюционной выигрышностью откладки мелких многочисленных яиц, на мой взгляд, связано возникновение и сохранение в большинстве групп животных такой личиночной стадии развития, которая еще не имеет всех систем органов, присущих имаго, но способна самостоятельно питаться и постепенно достигать размеров тела взрослого организма. Первые личиночные стадии губок и кишечнополостных (бластулы, паренхимулы, плакулы и т.п.), как известно, питаться не умеют: они плавают, затем прикрепляются, претерпевают существенный метаморфоз, и только после этого начинают самостоятельное питание (Захваткин, 1949; Иванова-Казас, 1975; Ересковский, 2005). Для примитивных прикрепленных животных сохранение такой стадии актуально, поскольку она обеспечивает расселение, а для активно перемещающихся организмов такие личинки бесполезны, что обуслoвливает процессы эмбрионизации онтогенеза и развитие непитающихся стадий внутри яйцевых оболочек (Ежиков, 1940; Иванова-Казас, 1975, с. 71). Эволюция онтогенеза более высокоразвитых животных сопровождается процессами дальнейшей эмбрионизации или, наоборот, дезэмбрионизации, то есть преждевременным выходом недоразвитой личиночной стадии из яйца (Ежиков, 1940, 1953; Иванова-Казас, 1975, с. 66–75; Шатров, 2003), но ни в каких случаях не приводит к восстановлению непитающейся бластулоподобной личинки низших многоклеточных. С дезэмбрионизацией обычно связывают возникновение вторичного метаморфоза в ряде групп животных (как, например, у насекомых Holometabola) (см. обзоры: Иванова-Казас, 1981, с. 159–174; Шатров, 2003). Однако ясных объяснений того, зачем нужна сама дезэмбрионизация, кажется, никто до сих пор не обнародовал. В рамках предлагаемого в настоящей статье подхода логично предположить, что дезэмбрионизованная личинка, способная к самостоятельному питанию, позволяет уменьшить размер откладываемых яиц и за счет этого увеличить их количество, что в свою очередь увеличивает размах изменчивости и возможности для естественного отбора. Таким способом животные со сложно организованной половой системой, внутренним оплодотворением и сложными оболочками яйца получают возможность, не утрачивая всех этих эволюционных преимуществ, вернуться к производству почти такого же огромного количества потомков (исчисляемого тысячами у многих Holometabola), какое было у первичноводных предков, выметывавших в воду многочисленные неоплодотворенные яйцеклетки.
К сожалению, в рамках настоящей статьи нет возможности обсуждать саму возможность дезэмбрионизации, как биологического явления, и разнообразные аргументы “за” и “против”, предъявлявшиеся разными авторами (см. цитированные выше обзоры). Более актуальным в рамках основной дискуссии является вопрос о возможной обратимости живорождения. В большинстве как старых, так и современных обзоров на эту тему возможность эволюционного возврата живородящих организмов к нормальной яйцекладке категорически отрицалась. Так, например, Иванова-Казас (1995, с. 294) пишет: “Возможность перехода от высокоспециализиро-ванного плацентарного живорождения к откладке богатых желтком яиц совершенно неправдоподобна, так как означала бы отказ от больших биологических преимуществ”. Примерно такие же соображения можно найти у Вурмса (Wourms, 1994, p. 552), Мейера (Meier et al., 1999, p. 207) и во многих других работах. Оставляя в стороне бездоказательные утверждения о “больших биологических преимуществах”, опровергаемые детальной аргументацией, представленной в настоящей статье, имеет смысл сосредоточиться на относительно хорошо изученных примерах возврата живородящих групп к яйцекладке. Такие примеры уже рассматривались мною выше для губок, среди которых наиболее древними являются именно живородящие и яйцеживородящие группы (Ересковский, 2005; Riesgo et al., 2014) и для онихофор, у которых немногие примеры современных яйцекладущих видов рассматриваются как возникшие вторично от живородящих предков (Mayer et al., 2015). Наиболее архаичные из современных мшанок, относимые к классу Phylactolaemata, представлены исключительно живородящими организмами, тогда как яйцекладущие виды встречаются в более “продвинутых” классах Stenolaemata и Gymnolaemata (Taylor, Waeschenbach, 2015; Ostrovsky et al., 2016). Однако самое важное и неоспоримое доказательство онтогенетической легкости возврата от плацентарного живорождения к обычной яйцекладке демонстрируют тли (Homoptera: Aphidinea) надсемейства Aphidoidea, у которых чередование живородящих и яйцекладущих поколений в жизненном цикле является общим правилом для большинства видов. При этом тли обладают одним из самых сложных и аберрантных вариантов живорождения (Hagan, 1951; Gavrilov-Zimin, 2021 и мн. др. авторы). Эмбрионы живородящих поколений тлей начинают развиваться в организме матери задолго до того, как она сама достигает взрослого состояния. В результате в теле тли, как в матрешке, оказываются “вложены” следующие поколения. В частности, по этой причине, упоминавшиеся во введении к настоящей статье “старые” естествоиспытатели – Антони Левенгук и Шарль Бонне, изучавшие тлей, были ярыми сторонниками преформизма, то есть теории вложенности поколений, вплоть до половых клеток. У тлей из сем. Eriosomatidae (= Pemphigidae) (рис. 6) зависимость яйцекладущего поколения от предыдущего живородящего поколения доходит до крайней, почти абсурдной ситуации, при которой появившийся на свет путем плацентарного живорождения организм живет только за счет того запаса питательных веществ, который он получил от матери в ходе эмбрионального развития. Говоря иными словами, новое существо рождается, линяет несколько раз, превращается во взрослое насекомое, размножается, и при этом никогда за свою самостоятельную жизнь не ест и не пьет. Похожая ситуация, но без чередования поколений, наблюдается у некоторых кокцид (Homoptera: Coccinea) сем. Stictococcidae (Richard, 1971). У некоторых живородящих мух (Diptera: Schizophora) не только эмбриональное развитие, но и все этапы личиночного онтогенеза проходят внутри расширенных яйцеводов самки. Развивающиеся личинки при этом получают питание от матери за счет выделений модифицированных придаточных желез яйцевода (называемых даже “молочными железами” в англоязычной литературе – Vreysen et al., 2013, p. 17) или сперматеки, и в этом случае число рождаемых личинок резко ограничено (вплоть до единственного эмбриона). Яйцеживородящие шизофорные мухи, особенно из сем. Tachinidae, наоборот, не предоставляют никакого питания развивающимся внутри их половых путей миниатюрным эмбрионам, но число этих эмбрионов может достигать нескольких тысяч, а иногда даже 20 000 в яйцеводе одной самки (Meier et al., 1999).

В этой связи уместно поставить вопрос о сравнительной степени сложности, продуктивности и завершенности живорождения в разных группах животных. Как видно из перечисленных примеров, вопреки широко распространенному заблуждению, наиболее сложное и “совершенное” живорождение характерно отнюдь не для позвоночных, а для насекомых. Плацентарные млекопитающие с их малочисленными недоразвитыми детенышами, нуждающимися в родительской заботе и длительном постэмбриональном выкармливании, даже близко не подошли к такому уровню независимости потомства от внешнего питания, какое демонстрируют живородящие насекомые. Самым же заметным эволюционным достижением последних оказывается возможность сочетать живорождение (или яйцеживорождение) с производством большого количества потомков, то есть разрешить, насколько возможно, то базовое противоречие, которое возникло на ранних этапах эволюции многоклеточных животных и не было преодолено никем, кроме некоторых облигатно-паразитических групп и тлей. У тлей это было достигнуто за счет педогенетического партеногенеза и “вложенности” поколений, а у паразитических организмов – за счет предельной миниатюризации яиц и личинок. Так у паразитических мух-тахин (Tachinidae), производящих большое количество яиц с полностью или почти полностью развитыми личинками, длина яйца составляет около 170–310 мкм (см., например, Marini, Campadelli, 1994). Такой же большой плодовитостью (тысячи личинок) обладают и гемоцельно живородящие веерокрылые насекомые (Strepsiptera), у которых миниатюрные личинки имеют длину тела всего лишь 80–300 мкм при размерах неотенических самок 2000–6000 мкм (Kathirithamby, 1989). Уменьшенные почти до самых нижних возможных размеров многоклеточных животных, личинки яйцеживородящих мух и живородящих стрепсиптер не способны к свободному образу жизни, а являются облигатными внутренними паразитами других насекомых. Большое количество чрезвычайно мелких яиц с развитыми эмбрионами внутри образуется также у некоторых живородящих паразитических червей, в частности у скребней (Acanthocephala) (см., например, Wahl, Sparkes, 2012).
Все эти примеры демонстрируют, что даже большое количество потомков, произведенных на базе (яйце)живорождения, не приводит к какому-либо заметному росту биоразнообразия, по сравнению с ближайшими яйцекладущими таксонами. Это объяснимо не только облигатным внутренним паразитизмом, при котором среда обитания организма становится несравнимо менее разнообразной, нежели у свободноживущих организмов, но также частой подменой полового процесса партеногенезом. Кроме того, естественный отбор живородящих организмов по отношению к факторам внешней среды воздействует только на постэмбриональные или даже только на имагинальные стадии. Из упомянутых выше групп, стрепсиптеры насчитывают всего лишь около 600 современных видов, тогда как анцестральные по отношению к ним и почти тотально яйцекладущие жуки (отряд Coleoptera) – 400 000 видов. Точное число видов мух, которые производят большое количество (больше 100 на самку) личинок путем яйцеживорождения, указать затруднительно, но из имеющегося обзора (Meier et al., 1999) создается впечатление, что такие примеры единичны на фоне примерно 150 000 преимущественно яйцекладущих Diptera. Живородящие и яйцеживородящие тли насчитывают около 5000 номинальных видов в современной фауне, но большинство этих видов различается между собой лишь по метрическим (и часто перекрывающимся) признакам, что, вероятно, свидетельствует о сильно переоцененном количестве реально существующих в природе видов тлей, особенно в типовом семействе Aphididae s.s. Кроме того, в отличие от большинства других групп насекомых, почти все разнообразие тлей сосредоточено в хорошо изученной голарктической зоне, что исключает возможность какого-либо существенного прироста числа реальных новых видов, не обнаруженных до сих пор в природе. В любом случае общее морфоанатомическое, экологическое, цитогенетическое разнообразие тлей невелико в сравнении со сходными по формальному числу видов другими таксонами паранеоптерных насекомых. Особенно наглядно это проявляется при сравнении тлей с сестринской к ним группой – кокцидами, среди которых есть как яйцекладущие, так и яйцеживородящие виды (Gavrilov-Zimin, 2018). Кроме того, как уже было сказано выше, живорождение у самих тлей циклически чередуется с яйцекладкой. Скребни (Acanthocephala) насчитывают около 1400 видов и, как и большинство внутренних паразитов, обладают сильно упрощенным и однообразным строением. Не вполне ясное филогенетическое положение скребней не позволяет корректно сравнивать их c другими группами животных. Однако сходные по уровню организации круглые черви (Nematoda), среди которых немало высокоспециализированных паразитов, характеризуются преимущественно яйцекладным способом размножения и насчитывают около 24 000 современных видов. Яйцекладущие коловратки (Rotifera), считающиеся возможными предками скребней, насчитывают около 2200 видов, но их предельно малые для многоклеточных животных размеры тела (от 40 мкм длиной) и часто встречающийся партеногенез не могут способствовать биологическому разнообразию, независимо от яйцекладности.
Возвращаясь к проблеме эволюционной обратимости задержки яиц в материнском организме, необходимо отметить, что такая реверсия представляется более вероятной и более частой в случаях яйцеживорождения, нежели плацентарного, аденотрофного или внутриполостного живорождения. Например, у кокцид известны многочисленные примеры циклических переходов от полного яйцеживорождения к нормальной или почти нормальной откладке яиц на самых ранних этапах эмбрионального развития с попутным возникновением специфических приспособлений для защиты яиц (рис. 4). Так, у археококцид (Oerthezioidea) факультативное неполное яйцеживорождение (в “примитивных” родах Xylococcinae s.l. и у большинства Callipappinnae s.l.) эволюционирует в нормальную яйцекладку у их предполагаемых потомков (Margarodinae s.s., некоторые Monophlebinae и Ortheziidae); при этом у них появляются новые адаптации, такие как откладывание яиц в специальную полость под телом или в плотный восковой мешок позади тела. Наоборот, “продвинутые” Monophlebinae и их потомки (Phenacoleachiidae, Carayonemidae и первые неококциды) снова характеризуются неполным или полным яйцеживорождением с откладкой эмбрионизированных яиц в марзупий или прямо во внешнюю среду. Среди неококцид (Coccoidea) полное яйцеживорождение, свойственное “примитивным” родам мучнистых червецов (Pseudococcidae), эволюционирует в неполное яйцеживорождение или почти нормальную яйцекладку (на самых начальных этапах дробления) у ряда сильно дивергировавших родов. Некоторые ложнощитовки (Coccidae: Pulvinariini, Eriopeltinae, Filippiinae) образуют рыхлый яйцевой мешок, подобно их далеким предкам из Monophlebinae и Xylococcinae, но, в отличие от последних, используют для его построения не многоячеистые восковые железы, а более сложные трубчатые железы различной структуры. Многие сильно дивергировавшие ложнощитовки (Coccidae) и кермесы (Kermesidae) откладывают более или менее эмбрионизированные яйца в полость под телом или же во внутреннюю полость, подобную марзупию монофлебин. В наиболее аберрантных кокцидных семействах Asterolecaniidae s.l., Diaspididae, Phoenicoccoccidae s.l., демонстрирующих дальнейшую редукцию стадий онтогенеза и утрату подвижности неотенической самки, вторично происходит возврат к полному яйцеживорождению во многих родах (Gavrilov-Zimin, 2018).
Примеры вероятного возврата некоторых яйцеживородящих мух к яйцекладке кратко обсуждались Мейером с соавт. (Meier et al., 1999, p. 207–208).
В недавнее время разгорелась дискуссия вокруг проблемы возможной первичности живорождения у скваматных (чешуйчатых) рептилий и вторичного возврата к яйцекладке в большинстве современных их родов. Одна группа специалистов (Pyron, Burbrink, 2013) обосновывает первичность живорождения в эволюции Squamata, тогда как другая (Griffith et al., 2015) – это отрицает. Примечательно, что и те, и другие являются поклонниками статистического кладизма и настаивают на объективности своего анализа филогенеза. В рамках общей теории живорождения, предлагаемой в настоящей статье, более логичным представляется исходное яйцеживорождение всех амниот в связи с переходом к внутреннему оплодотворению и утрате самостоятельно питающейся личиночной стадии, дальнейшее сохранение этого свойства (с переходами к плацентарному живорождению) у синапсид (Synapsida) и древних скваматных (Squamata) рептилий, но однократный возврат к нормальной яйцекладке у архозавров (крокодилов, черепах и птиц) и аналогичные многократные возвраты в разных линиях Squamata (рис. 5). Такой подход позволяет объяснить возникновение (яйце)живорождения амниот той же самой, обозначенной выше, простой причиной, по которой оно возникало и во многих других группах животных.
ЗАКЛЮЧЕНИЕ
Подводя итог обсуждению, мне хотелось бы выделить следующие тезисы, представляющиеся наиболее важными для понимания эволюции живорождения (яйцеживорождения).
1) Живорождение не является “венцом” эволюции репродуктивной сферы организмов, а возникает на самых ранних этапах филогенеза многоклеточных животных и, в дальнейшем, реверсируется многократно в качестве “резервного”, самого простого способа размножения.
2) Для живорождения не требуется сложной и длительной эволюции половой сферы и даже наличия половых органов, ни внешних, ни внутренних. Задержка потомства в теле материнского организма возникает как простейший из способов размножения, возможных на базе оогамии, то есть неподвижности и увеличения размера женской гаметы (яйцеклетки) в сравнении с подвижностью и малым размером мужской гаметы (сперматозоида). Наоборот, выведение яйцеклеток из тела во внешнюю среду, появление сложнейших структур половой системы высокоразвитых многоклеточных организмов, образование яйцевых оболочек и накопление желтка в яйце необходимы для того, чтобы избежать живорождения.
3) Живорождение (или яйцеживорождение) само по себе невыгодно с эволюционной точки зрения, поскольку базово означает производство небольшого количества потомков, сниженные возможности для естественного отбора и соответствующее ограничение биологического разнообразия таксона. Кроме того, при живорождении действию естественного отбора организмов по отношению к факторам внешней среды оказываются подвержены только постэмбриональные или даже только имагинальные стадии, а не целиком все стадии онтогенеза, как при нормальной яйцекладке.
4) Причинами реверсий живорождения являются различные аберрации репродуктивной системы, изменяющие ее предковое состояние и побочно приводящие к невозможности нормальной яйцекладки: а) переходы от наружного оплодотворения к внутреннему, когда у организма еще отсутствуют специализированные половые протоки (яйцеводы) с придаточными железами и сперматеками; б) нарушения в хорошо развитой половой яйцекладной системе: утрата имагинальных половых органов (вследствие педогенеза, неотении, педоморфоза), ларвальный мейоз, изменение места оплодотворения с эктодермальных частей половой системы на внутригонадное.
5) Яйцеживорождение, а в отдельных случаях также и живорождение, представляют собой обратимые состояния репродуктивной стратегии, от которых возможен эволюционный возврат к яйцекладке.
6) В жизненном цикле высших растений, а также водорослей и грибов, отсутствуют явления, гомологичные живорождению и яйцекладке животных. Растительная (в широком смысле) оогамета – яйцеклетка не выводится во внешнюю среду, а прорастает на материнском организме (гаметофите) и дает начало спорофиту. Семена растений не гомологичны яйцу, поскольку представляют собой сложные многоклеточные образования, состоящие из тканей разного онтогенетического происхождения. В связи с этими особенностями, растительные организмы логичнее называть спорородящими и/или семяродящими, а не живородящими.
Автор надеется, что предлагаемые в настоящей статье подходы будут способствовать целенаправленным исследованиям в области эволюции живорождения и выявлению конкретных путей трансформации одного способа отрождения потомства в другой, без обращения к затейливым статистическим подсчетам, вычислениям “энергетики размножения” или “инвестирования в потомство”, столь популярным в современной биологии. Для понимания причин и следствий живорождения необходимо немодное в нынешнее время детальное знание анатомии, эмбриологии и репродуктивного поведения животных. Именно такой информации остро не хватает, особенно по насекомым и многим группам морских беспозвоночных животных. Этот пробел невозможно заполнить данными смежных с классической зоологией дисциплин – ни экологией, ни биохимией, ни генетикой, ни молекулярной биологией, ни тем более математикой или физикой.
Список литературы
Аристотель. О возникновении животных. М.–Л.: АН СССР, 1940. 250 с.
Батыгина Т.Б., Брагина Е.А., Ересковский А.В., Островский А.Н. Живорождение у растений и животных: беспозвоночных и низших хордовых. Санкт-Петербург: СПбГУ, 2006. 132 с.
Беклемишев В.Н. Основы сравнительной анатомии беспозвоночных. Т. 2. М.: Наука, 1964. 446 с.
Бэр К.М. Об открытии профессором Вагнером бесполого размножения личинок, о дополнительных наблюдениях по этому предмету г. Ганина и педогенезисе вообще // Приложение к X тому Зап. Имп. Акад. наук, 1866. Т. 1. С. 1–77. [Немецкий оригинал: Baer K.H. Ueber Prof. Nic. Wagners’s Entdeckung von Larvae die sich fortpflanzen. Herr Ganin’s verwandte und ergänzende Beobachtungen und über dei Paedogenese überhaupt // Bulletin de l’Académie Impériale des Sciences de St. Petersburg. 1866. Bd. 9. S. 64–137.].
Вагнер Н.П. Самопроизвольное размножение гусениц у насекомых. Казань: Казанский университет, 1862. 50 с.
Гаврилов И.А. О причинах и следствиях яйцеживорождения у гемиптероидных насекомых (Insecta: Paraneoptera) // Мат. отч. науч. сес. ЗИН РАН. Санкт-Петербург, 2009. С. 12–13.
Гаврилов-Зимин И.А. Морфологические, цитогенетические и онтогенетические основы систематики псевдококцид (Homoptera: Coccinea: Pseudococсidae) Палеарктики: Дис. … докт. биол. наук. СПб: ЗИН РАН, 2017. 673 с.
Десницкий А.Г. Вольвокс как модель для изучения гибели клеток и старения // Онтогенез. 2021. Т. 52. № 4. С. 295–304.
Ежиков И.И. О ранних эмбриональных стадиях и их связи с типами постэмбрионального развития у насекомых // ДАН СССР. 1940. Т. 28. № 6. С. 574–576.
Ересковский А.В. Сравнительная эмбриология губок (Porifera). СПб.: Изд-во СПбГУ, 2005. 304 с.
Захваткин А.А. Сравнительная эмбриология низших беспозвоночных. М.: Советская наука, 1949. 395 с.
Захваткин Ю.А. Факультативное живорождение у Chrysomela sanquinolenta L. (Coleoptera: Chrysomelidae) // Науч. докл. высш. школы. Биол. науки. 1966. № 3. С. 27–31.
Захваткин Ю.А. Эмбриология и систематика жуков-листоедов (Coleoptera, Chrysomelidae) // Энтомол. обозр. 1970. Т. 49. № 2. С. 347–354.
Зоология беспозвоночных в двух томах / Ред. В. Вестхайде, Р. Ригер. М.: КМК, 2008. 935 с. [Немецкий оригинал: Westheide W., Rieger R. Spezielle Zoologie. Teil 1: Einzeller und Wirbellose Tiere. Teil 2. Wirbel und Schädeltiere. Heidelberg Berlin: Spektrum Akademischer Verlag, 2004. 922+712 s.].
Иванова-Казас О.М. Сравнительная эмбриология беспозвоночных животных. Часть 1. Простейшие и низшие многоклеточные. Новосибирск: Наука, 1975. 372 с.
Иванова-Казас О.М. Сравнительная эмбриология беспозвоночных животных. Часть 2. Трохофорные, щупальцевые, щетинкочелюстные, погонофоры. М.: Наука, 1977. 312 с.
Иванова-Казас О.М. Сравнительная эмбриология беспозвоночных животных. Часть 3. Иглокожие и полухордовые. М.: Наука, 1978а. 166 с.
Иванова-Казас О.М. Сравнительная эмбриология беспозвоночных животных. Часть 4. Низшие хордовые. М.: Наука, 1978б. 166 с.
Иванова-Казас О.М. Сравнительная эмбриология беспозвоночных животных. Часть 5. Членистоногие (онихофоры, тихоходки, пентастомиды, пантоподы, трилобиты, хелицеровые и ракообразные). М.: Наука, 1979. 224 с.
Иванова-Казас О.М. Сравнительная эмбриология беспозвоночных животных. Часть 6. Неполноусые. М.: Наука, 1981. 207 с.
Иванова-Казас О.М. Эволюционная эмбриология животных. СПб.: Наука, 1995. 565 с.
Иванова-Казас О.М. Неотения как особый модус эволюции. 1. Неотения у низших Metazoa, полихет и моллюсков // Зоол. журн. 1997. Т. 76. № 11. С. 1244–1255.
Касьянов В.Л. Репродуктивная стратегия морских двустворчаиых моллюсков и иглокожих. Л.: Наука, 1989. 179 с.
Клюге Н.Ю. Систематика насекомых и принципы кладоэндезиса. В двух томах. М.: КМК, 2020. 518 + 531 с.
Малахов В.В. Происхождение билатерально-симметричных животных (Bilateria) // Журн. общ. биол. 2004. Т. 64. № 5. С. 371–388.
Наумов Н.П., Карташев Н.Н. Зоология позвоночных. Часть 2. М.: Высшая школа, 1979. 272 с.
Нидхэм Дж. История эмбриологии. М.: Госиздат, 1947. 842 с.
Островский А.Н. Эволюция полового размножения мшанок отряда Cheilostomata (Bryozoa: Gymnolaemata). СПб.: Изд-во СПбГУ, 2009. 404 с.
Плавильщиков Н.Н. Очерки по истории зоологии. М.: Наркомпрос РСФСР, 1941. 296 с.
Смирнов А.В. Педоморфные признаки в строении голотурий и роль педоморфоза в возникновении и эволюции класса Holothuroidea // Морфогенез: гетерохронии, гетеротопии и аллометрия (Серия “Геобиологические процессы в прошлом”) / Ред. С.В. Рожнов. 2014. М.: ПИН РАН. С. 178–209.
Трапезникова И.В., Гаврилов И.А. О яйцеживорождении у мучнистых червецов (Homoptera: Coccinea: Pseudococcidae) // Тр. ЗИН РАН. 2008. Т. 312. С. 43–53.
Шатров А.Б. К вопросу об основах онтогенеза членистоногих (Arthropoda) и его морфологическом выражении // Энтомол. обозр. 2003. Т. 82. № 2. С. 511–525.
Ananthakrisnan T.N., Dhileepan K. Thrips-fungus association with special reference to the sporophagous Bactrothrips idolomorphus (Karny) (Tubulifera: Thysanoptera) // Proc. Indian Acad. Sci. (Animal Sci.). 1984. V. 93. № 3. P. 243–249. https://doi.org/10.1007/BF03186284
Bagnell R.S. On Thysanoptera from the Seychelles Islands and Rodriques // Ann. Magaz. Nat. Hist. 1921. V. 9. № 7. P. 257–293.
Blackburn D.G. Structure, function, and evolution of the oviducts of squamate reptiles, with special reference to viviparity and placentation // J. Exp. Zool. 1998. V. 282. P. 560–617.
Blackburn D.G. Evolution of vertebrate viviparity and specializations for fetal nutrition: a quantitative and qualitative analysis // J. Morphol. 2015. V. 276. № 8. P. 961–90.
Blackburn D.G., Evans H.E. Why are there no viviparous birds? // Am. Nat. 1986. V. 128. № 2. P. 165–190.
Blackburn D.G., Starck J.M. Morphological specializations for fetal maintenance in viviparous vertebrates: an introduction and historical retrospective // J. Morphol. 2015. V. 276. P. E1–E16.
Blackburn D.G., Stewart J.R. Morphological research on amniote eggs and embryos: an introduction and historical retrospective // J. Morphol. 2021. V. 282. P. 1024–1046.
Blackman R.L. Reproduction, cytogenetics and development // Aphids, their biology, natural enemies and control / Eds A.K. Minks, P. Harrewijn. Amsterdam: Elsevier, 1987. V. 2A. P. 163–195.
Bontems Ch. Localization of spermatozoa inside viviparous and oviparous females of Chrysomelinae // Biology of Chrysomelidae / Eds P. Jolivet, E. Petitpierre, T.H. Hsiao. Dordrecht: Kluwer Acad. Publishers, 1988. P. 299–316.
Bournier A. L’embryogenèse de Caudothrips buffai Karny [Thys. Tubulifera] // Ann. de la Société Entomol. de France (N.S.). 1966. V. 2. № 2. P. 415–435.
Buchner P. Endosymbiosestudien an Schildläusen. 6. Die nicht in Symbiose lebende Gattung Apiomorpha und ihre ungewöhniche Embryonalentwicklung // Zeitschrift für Morphologie und Ökologie der Tiere. 1957. B. 46. S. 481–528.
Buchner P. Endosymbiosestudien an Schildläusen. 7. Weitere Beiträge zur Kenntnis der Stictococcinensymbiose // Zeitschrift für Morphologie und Ökologie der Tiere. 1963. B. 52. S. 401–458.
Buchner P. Endosymbiosis of animals with plant microorganisms. New York: John Wiley & Sons, 1965. 909 p.
Buckley D. Evolution of viviparity in salamanders (Amphibia, Caudata) // eLS. 2012. P. 1–5. https://doi.org/10.1002/9780470015902.a0022851
Carayon J. Trois espèces africaines de Physopleurella (Hémipt. Anthocoridae) dont l’une présente un nouveau cas de viviparité pseudoplacentaire // Bull. du Muséum Nat. d’Histoire Naturelle. 1956. V. 28. № 1. P. 102–110.
Carayon J. La viviparite chez les Heteropteres // Verhandlungen Internat. Congress Entomol. (Wien). 1961. V. 1. P. 711–714.
Carayon J. Traumatic insemination and paragenital system // Uzinger R.L. Monograph of Cimicidae (Hemiptera–Heteroptera). Baltimore: The Thomas Say Foundation, 1966. P. 81–166.
Cobben R.H. Evolutionary trends in Heteroptera. Part I. Eggs, architecture of the shell, gross embryology and eclosion. Wageningen: Pudoc, 1968. 477 p.
Danzig E.M., Gavrilov-Zimin I.A. Palaearctic mealybugs (Homoptera: Coccinea: Pseudococcidae). Part 1. Subfamily Phenacoccinae. St. Petersburg: ZIN RAS, 2014. 678 p. (Fauna of Russia and neighbouring countries. New series, № 148. Insecta: Hemiptera: Arthroidignatha).
Dhileepan K., Ananthakrisnan T.N. Ovarian polymorphism in relation to reproductive diversity and associated histological and histochemical attributes in some sporophagous tubuliferan thysanoptera // Proc. Indian Acad. Sci. (Anim. Sci.). 1987. V. 96. № 1. P. 1–13. https://doi.org/10.1007/BF03179386
Dunn C.W., Giribet G., Edgecombe G.D., Hejnol A. Animal phylogeny and its evolutionary implications // Annu. Rev. Ecol. Evol. Syst. 2014. V. 45. P. 371–395. https://doi.org/10.1146/annurev-ecolsys-120213-091627
Edmunds G.F. Ovoviviparous mayflies of the genus Callibaetis (Ephemeroptera: Baetidae) // Entomol. News. 1945. V. 56. P. 169–171.
Eichler W. Parthenogenese und Ovoviviparie als Entwicklungseigentümlichkeiten bei Läusen und Federlingen // Tierärztliche Umschau. 1946. V. 1. № 1. P. 10.
Eitel M., Guidi L., Hadrys H., Balsamo M., Schierwater B. New insights into Placozoan sexual reproduction and development // PLoS One. 2011. V. 6. № 5. P. e19639. https://doi.org/10.1371/journal.pone.0019639
Fernando W. The early embryology of viviparous Psocid // Quart. J. Microsc. Sci. 1934. V. 77. P. 99–120.
Garstang W. The theory of recapitulation – a critical restatement of the biogenetic law // Linn. Soc. London (Zool.). 1922. V. 35. P. 81–101.
Gavrilov-Zimin I.A. Ontogenesis, morphology and higher classification of archaecococcids (Homoptera: Coccinea: Orthezioidea) // Zoosystematica Rossica (Suppl. 2). 2018. 260 p. https://doi.org/10.31610/zsr/2018.supl.2.1
Gavrilov-Zimin I.A. Egg retention, viviparity and ovoviviparity in Paraneoptera // Comp. Cytogen. 2021. V. 15. № 3. P. 239–252. https://doi.org/10.3897/CompCytogen.v15.i3.70216
Gavrilov-Zimin I.A., Danzig E.M. Taxonomic position of the genus Puto Signoret (Homoptera: Coccinea: Pseudococcidae) and separation of higher taxa in Coccinea // Zoosystematica Rossica. 2012. V. 22. № 1. P. 97–111. https://doi.org/10.31610/zsr/2012.21.1.97
Gavrilov-Zimin I.A., Stekolshchikov A.V., Gautam D.C. General trends of chromosomal evolution in Aphidococca (Insecta, Homoptera, Aphidinea + Coccinea) // Comp. Cytogen. 2015. V. 9. № 3. P. 335–422. https://doi.org/10.3897/CompCytogen.v9i3.4930
Gavrilov-Zimin I.A., Grozeva S.M., Gapon D.A. et al. Introduction to the study of chromosomal and reproductive patterns in Paraneoptera // Comp. Cytogen. 2021. V. 15. № 3. P. 217–238. https://doi.org/10.3897/compcytogen.v15.i3.69718
Giard A. Sur la progenése // Bull. Scientifique de la France et de la Belgique. 1887. V. 18. P. 25.
Golub N.V., Nokkala S. Chromosome numbers of two sucking louse species (Insecta, Phthiraptera, Anoplura) // Hereditas. 2004. V. 141. P. 94–96. https://doi.org/10.1111/j.1601-5223.2004.01859.x
Griffith O.W., Blackburn D.G., Brandley M.C. et al. Ancestral state reconstructions require biological evidence to test evolutionary hypotheses: a case study examining the evolution of reproductive mode in squamate reptiles // J. Experim. Zool. (Mol. Dev. Evol.). 2015. V. B. P. 1–11. https://doi.org/10.1002/jez.b.22614
Hagan H.R. The embryogeny of the polyctenid, Hesperoctenes fumarius Westwood, with reference to viviparity in insects // J. Morphol. 1931. V. 51. P. 1–117.
Hagan H.R. Embryology of viviparous insects. New York: The Ronald Press Company, 1951. 472 p.
Hansen B. Brood-protection in deep-sea holothurian Oneirophanta mutabilis Théel // Nature. 1968. V. 217. P. 1062–1063.
Harker J.E. The role of parthenogenesis in the biology of two species of mayfly (Ephemeroptera) // Freshwater Biol. 1997. V. 37. P. 287–297.
Hathaway C.R. Verificação da viviparidade em Thysanoptera // Memórias do Instituto Oswaldo Cruz. 1938. V. 33. P. 357–358.
Hendler G. Adaptational significance of the patterns of ophiuroid development // Am. Zool. 1975. V. 15. № 3. P. 691–715.
Hille Ris Lambers D. An apparently unrecorded mode of reproduction in Aphididae // Proc. of 8th Internat. Congr. of Entomol., Stockholm, 1950. P. 235.
Hood J.D. Some further new Thysanoptera from Panama // Proc. Biol. Soc. Washington. 1934. V. 47. P. 57–82.
Israelsson O., Budd G.E. Eggs and embryos in Xenoturbella (phylum uncertain) are not ingested prey // Dev. Gen. Evol. 2005. V. 215. № 7. P. 358–363. https://doi.org/10.1007/s00427-005-0485-x
Iwan D. Ovoviviparity in tenebrionid beetles of the melanocratoid Platinotina (Coleoptera: Tenebrionidae: Platynotini) from Madagascar, with notes on the viviparous beetles // Ann. Zoologici (Warzsawa). 2000. V. 50. № 1. P. 15–25.
Jentsch S. Ovoviviparie bei einer einheimischen Copeognathenart (Hyperetes guestphalicus) // Zoologicher Anzeiger. 1936. V. 116. P. 287–289.
John O. Fakultative Viviparität bei Thysanopteren // Entomologische Mitteilungen. 1923. V. 12. № 5–6. P. 227–232.
Kardong K.V. Vertebrates: comparative anatomy, function, evolution. New York: McGraw-Hill, 2012. 794 p.
Kathirithamby J. Review of the Order Strepsiptera // Syst. Entomol. 1989. V. 14. P. 41–92. https://doi.org/10.1111/j.1365-3113.1989.tb00265.x
Kathirithamby J., Hrabar M., Delgado J.A. et al. We do not select, nor are we choosy: reproductive biology of Strepsiptera (Insecta) // Biol. J. Linn. Soc. 2015. V. 116. № 1. P. 221–238. https://doi.org/10.1111/bij.12585
Klapow L.A. Ovoviviparity in the genus Excirolana (Crustacea: Isopoda) // J. Zool. 1970. V. 162. P. 359–369. https://doi.org/10.1111/j.1469-7998.1970.tb01271.x
Kluge N.J. A new subgenus Oculogaster subgen. n. for viviparous representatives of Procloeon s. l., with discussion about status of the generic name Austrocloeon Barnard 1932 and the species name africanum Esben-Petersen 1913 [Cloeon] (Ephemeroptera, Baetidae) // Zootaxa. V. 4107. № 4. P. 491–516.
Kocarek P. A case of viviparity in a tropical non-parasitizing earwig (Dermaptera Spongiphoridae) // Tropical Zool. 2009. V. 22. P. 237–241.
Köhler F., von Rintelen T., Meyer A., Glaubrecht M. Multiple origin of viviparity in Southeast Asian gastropods (Cerithioidea: Pachychilidae) and its evolutionary implications // Evolution. 2004. V. 58 (10). P. 2215–2226.
Kollmann J. Das Ueberwintern von europäischen Frosch- und Tritonlarven und die Umwandlung des mexikanischen Axolotl // Verhandl. Natur. Ges. Basel. 1884. B. 7. S. 387–398.
Lee G.-H., Li H. Ovoviviparity and morphology of the female reproductive system in Palumbina (Lepidoptera, Gelechiidae, Thiotrichinae) // J. Asia-Pacific Entomol. 2018. V. 21. № 3. P. 772–777.
Lester S.M. Ultrastructure of adult gonads and development and structure of the larva of Rhabdopleura normani (Hemichordata: Pterobranchia) // Acta Zoologica (Stockh.). 1988. V. 69. № 2. P. 95–109.
Levin T.C., King N. Evidence for sex and recombination in the choanoflagellate Salpingoeca rosetta // Curr. Biol. 2013. V. 23. № 21. P. 2176–2180.
Lister M. Letter concerning a kind of viviparous fly // Philosophical Transactions. 1671. V. 6. № 72. P. 2170–2171.
Marini M., Campadelli G. Ootaxonomy of Goniini (Diptera Tachinidae) with microtype eggs // Bollettino di Zoologia. 1994. V. 61. № 3. P. 271–283. https://doi.org/10.1080/11250009409355894
Mayer G., Franke F.A., Treffkorn S. et al. Onychophora // Evolutionary developmental biology of invertebrates 3: Ecdysozoa I: Non-Tetraconata / Ed. A. Wanninger. Wien: Springer-Verlag, 2015. P. 53–98. https://doi.org/10.1007/978-3-7091-1865-8_4
Meier R., Kotrba M., Ferrar P. Ovoviviparity and viviparity in the Diptera // Biol. Rev. 1999. V. 74. P. 199–258.
Mockford E.L. Life history studies on some Florida insects of the genus Archipsocus // Bull. Florida Museum Nat. Hist. 1957. V. 1. № 5. P. 253–274.
Morse C., Neboiss A. Triplectides of Australia (Insecta: Trichoptera: Leptoceridae) // Mem. Nat. Museum of Victoria. 1982. V. 44. № 2. P. 61–98.
Nagrale S.M. Life history of mycophagous thrips Elaphrothrips procer (Schmutz) (Thysanoptera: Phlaeothripidae) // Sci. Res. Report. 2012. V. 2. № 3. P. 256–259.
Nakata S., Maa T.C. A review of the parasitic earwings (Dermaptera: Arixeniina; Hemimerina) // Pacific Insects. 1974. V. 16. № 2–3. P. 307–374.
Nielsen C. Entoproct life-cycles and the entoproct/ectoproct relationship // Ophelia. 1971. V. 9. №. 2. P. 209–341.
Nozaki H., Yamada T.K., Takahashi F. et al. New “missing link” genus of the colonial volvocine green algae gives insights into the evolution of oogamy // BMC Evol. Biol. 2014. V. 14. № 37. P. 1–10. https://doi.org/10.1186/1471-2148-14-37
Ostrovsky A.N., Lidgard S., Gordon D.P. et al. Matrotrophy and placentation in invertebrates: a new paradigm // Biol. Rev. 2016. V. 91. P. 673–711. https://dx.doi.org/10.1111%2Fbrv.12189
Peinert M., Wipfler B., Jetschke et al. Traumatic insemination and female counter-adaptation in Strepsiptera (Insecta) // Sci. Rep. 2016. V. 6 (25052). P. 1–10. https://doi.org/10.1038/srep25052
Pyron R.A., Burbrink F.T. Early origin of viviparity and multiple reversions to oviparity in squamate reptiles // Ecol. Lett. 2013. V. 17. P. 13–21.
Ramirez-Llodra E., Reid W.D.K., Billett D.S.M. Long-term changes in reproductive patterns of the holothurian Oneirophanta mutabilis from the Porcupine Abyssal Plain // Marine Biol. 2005. V. 146. P. 683–693. https://doi.org/10.1007/s00227-004-1470-z
Richard C. Contribution à l'étude morphologique et biologique des Stictococcinae (Hom. Coccoidea) // Ann. de la Société Entomol. de France. 1971. V. 7. P. 571–609.
Riesgo A., Novo M., Sharma P.P. et al. Inferring the ancestral sexuality and reproduction condition in sponges (Porifera) // Zoologica Scripta. 2013. V. 43. P. 101–117.
Roth L.M., Willis E.R. An analysis of oviparity and viviparity in the Blattaria // Trans. Am. Entomol. Soc. 1957. V. 83. № 4. P. 221–238.
Sellier R. La viviparite chez les insectes // Année Biologique (Paris). 1955. V. 31. P. 525–545.
Stebbing A.R.D. Aspects of the reproduction and life cycle of Rhabdopleura compacta (Hemichordata) // Marine Biol. 1970. V. 5. № 3. P. 205–212.
Taylor P.D., Waeschenbach A. Phylogeny and diversification of bryozoans // Palaeontology. 2015. V. 58. P. 585–599. https://doi.org/10.1111/pala.12170
Tompa A.S. Oviparity, egg retention and ovoviviparity in Pulmonates // J. Moll. Stud. 1979. V. 45. № 2. P. 155–160.https://doi.org/10.1093/oxfordjournals.mollus.a065489
Trauth S.E., Sever D.M., Semlitsch R.D. Cloacal anatomy of paedomorphic female Ambystoma talpoideum (Caudata: Ambystomatidae), with comments on intermorph mating and sperm storage // Canadian J. Zool. 1994. V. 72. P. 2147–2157.
Treffkorn S., Hernández-Lagos O.Y., Mayer G. Evidence for cell turnover as the mechanism responsible for the transport of embryos towards the vagina in viviparous onychophorans (velvet worms) // Front. Zool. 2019. V. 16. № 16. P. 1–16. https://doi.org/10.1186/s12983-019-0317-x
Uichanco L.B. Studies on the embryogeny and postnatal development of the Aphididae with special reference to the history of the “symbiotic organ” or “mycetom” // Philippine J. Sci. 1924. V. 24. P. 143–247.
Viswanathan T.R., Ananthakrishnan T.N. On the partial ovoviviparity in Tiarothrips subramanii (Ramk.); (Thysanoptera: Insecta) // Curr. Sci. 1973. V. 42. № 18. P. 649–650.
Vreysen M.J.B., Seck M.T., Sall B., Bouyer J. Tsetse flies: their biology and control using area-wide integrated pest management approaches // J. Invert. Pathol. 2013. V. 112. Suppl. 1. P. S15–S25. https://doi.org/10.1016/j.jip.2012.07.026
Wahl G.M., Sparkes T.C. Egg dispersal in the acanthocephalan Acanthocephalus dirus: field data // J. Parasitol. 2012. V. 98. № 4. P. 894–896.
Wake M.H. Evolution of oviductal gestation in amphibians // J. Experim. Zool. 1993. V. 266. P. 394–413.
Warburg M.R. Pre- and post-parturial aspects of scorpion reproduction: a review // Eur. J. Entomol. 2012. V. 109. P. 139–146.
Washburn S.L. Tools and human evolution // Sci. American. 1960. V. 203. № 3. P. 63–75.
Webb J.K., Shine R., Christian K.A. The adaptive significance of reptilian viviparity in the tropics: testing the maternal manipulation hypothesis // Evolution. 2006. V. 60. P. 115–122.
Westblad E. Xenoturbella bocki n. g., n. sp., a peculiar, primitive turbellarian type // Arkiv för Zoologi. 1949. V. 1. P. 3–29.
Willey A. The anatomy and development of Peripatus novae-britanniae / Zoological results based on material from New Britain, New Guinea, Loyalty Islands and elsewhere collected during the years 1895, 1896 and 1897. Part 1. Cambridge: Univ. Press, 1898. P. 1–52.
Wong S.K., Thornton I.W.B. The internal morphology of the reproductive systems of some psocid species // Proc. Entomol. Soc. London (A). 1968. V. 43. № 1–3. P. 1–12.
Wourms J.P. Reproduction and development of chondrichthyan fishes // Am. Zoologist. 1977. V. 17. P. 379–410.
Wourms J.P. The challenges of piscine viviparity // Israel J. Zool. 1994. V. 40. P. 551–568.
Zilberman B., Pires-Silva C.M., Moreira I.E. et al. State of knowledge of viviparity in Staphylinidae and the evolutionary significance of this phenomenon in Corotoca Schiødte, 1853 // Papéis Avulsos De Zool. 2019. V. 59 (e20195919). P. 1–10. https://doi.org/10.11606/1807-0205/2019.59.19
Дополнительные материалы отсутствуют.
Инструменты
Успехи современной биологии