

Успехи физиологических наук, 2019, T. 50, № 2, стр. 93-104
Методические и методологические принципы исследования когнитивных способностей крыс
Н. Р. Григорьев 1, Т. А. Баталова 1, *, Г. Е. Чербикова 1
1 Федеральное государственное образовательное учреждение высшего образования
“Амурская государственная медицинская академия” Министерства здравоохранения Российской Федерации
675006 Благовещенск, Россия
* E-mail: batalova_ta@mail.ru
Поступила в редакцию 20.01.2018
Аннотация
В обзоре рассматривается современное состояние методических и методологических способов в исследовании когнитивного поведения у животных. Освещаются подходы к проблеме в историческом плане, представлены собственные методы исследования познавательного поведения. Предлагается количественная оценка индивидуального уровня когнитивных способностей крыс при решении ими стандартной когнитивной задачи в проблемной закономерно изменяющейся среде. Прослеживается процесс формирования в мозге крыс когнитивной, или дорожной, карты, постулированной Толменом в 1948 г., существование которой подтверждено нейрофизиологически Нобелевскими лауреатами по физиологии 2014 г.
МЕТОДОЛОГИЧЕСКИЕ ПРИНЦИПЫ ЭКСПЕРИМЕНТАЛЬНОГО ИССЛЕДОВАНИЯ КОГНИТИВНЫХ СПОСОБНОСТЕЙ ЖИВОТНЫХ
Время изучения физиологии высших функций мозга, редуцированных до условных и безусловных рефлексов, прошло, о чем свидетельствуют современные достижения нейроанатомии, нейрогистологии, нейрофизиологии и экспериментальной психологии. Исследованиями в области физиологии высшей нервной деятельности (поведения) животных [25] установлено, что сложные условные рефлексы формируются на базе врожденных безусловных рефлексов. Когда ввиду каких-либо причин условный рефлекс не достигает результата, создается нештатная ситуация возникает поисковая активность [3], или поведение поиска выхода из этой проблемной ситуации как у человека, так и у высших животных. Аналогично всем другим формам поведения, оно включает в себя как аффективные, так и когнитивные компоненты. Авторы теории поисковой активности В.С. Ротенберг и В.В. Аршавский [27] изучали аффективную составляющую поисковой активности. Однако для успешного и результативного поиска необходим достаточный для этого процесса уровень когнитивных способностей [19]. Исследованию функциональной организации поисковой активности и оценке когнитивных способностей у крыс были посвящены все наши ранние работы [8–14]. Невозможность быстро найти правильное разумное решения в высоко технологичных сферах деятельности человека приводит к катастрофе. Решение когнитивных задач животными в проблемных ситуациях упраздняет споры о присутствии или отсутствии мышления у животных. Животные мыслят, когда находят новые рациональные способы удовлетворения своих потребностей. Крысы способны усваивать правила поведения, оперировать образами и, как показывают наши опыты понимать причинно-следственные связи [19].
Экспериментальные психологи давно считают крыс интеллектуально одаренными животными. Об этом свидетельствуют их коллективный разум, поразительно успешное противостояние всяческим способам истребления и поразительная выживаемость вида в веках.
В физиологии поведения передовым и прогрессивным концептуальным подходом настоящего времени является теория функциональных систем П.К. Анохина [2], которая пришла на смену рефлекторной теории. В философском аспекте это совпало с появлением парадигмы физиологии активности [6], сменившей парадигму реактивности.
Теория условных рефлексов на современном этапе развития нейрофизиологии не решает проблемы когнитивного научения и такого же поведения, поскольку для его осуществления необходима рассудочная деятельность мозга и другое, не реактивное, а активное поведение. “Когда обезьяна строит свою вышку, чтобы достать плод – это “условным рефлексом” назвать нельзя. Это есть случай образования знания, уловление нормальной связи вещей!” [25]. Неизменная обстановка и стабильная среда не располагают к предвидению, в таких условиях требуется только память, или временная связь. В реальной жизни обстановка всегда выходит за рамки стабильности. Внешняя среда постоянно изменяется и довольно быстро: каждую минуту, секунду, даже ее десятые доли, и изолироваться от этих изменений невозможно. “Все течет, все изменяется. Нельзя в одну и ту же реку войти дважды!” – утверждал Гераклит Эфесский.
Нами создана гексагональная проблемная камера (ГПК) с закономерно изменяющейся структурой среды, которая переводит реактивное оборонительное поведение в поведение активного поиска, что позволяет измерять когнитивные способности у крыс [8, 9, 12, 16]. Во всех устройствах, способах изучения когнитивных способностей и условных рефлексов обстановка была строго статична, включая и лабиринты Э. Толмена [52]. Наши устройство и способ базируются на закономерных и постоянных изменениях среды ГПК, где крысе запрещены повторные выходы в одну и ту же дверку и необходим постоянный поиск новой неблокированной дверки в свободном поведении.
Для животных и человека когнитивные способности необходимы и востребованы только в проблемных ситуациях, поскольку во врожденных и приобретенных формах поведения в них нет никакой надобности. В этих случаях необходима только сформированная программа и алгоритм для ее выполнения, так, как это происходит в ЭВМ и потому их называют не мыслящими, а вычислительными. При решении возникающих проблем необходим прогноз и вынесение функции вперед в будущее на отрезке времени, когда необходимы предвидение и поиск наиболее вероятных результативных способов удовлетворения потребности. В закономерно изменяющейся среде ГПК крысе необходима рассудочная деятельность мозга для принятия правильного решения при выборе направления побежки, формирования плана и программы действий [40]. При такой активности крысы в камере постоянно необходимо то, что называлось ранее психонервным научением [5], моделью потребного будущего [6], опережающим отражением действительности [2], вероятностным прогнозом [32], моделью проблемной среды [33], когда мозг крысы осуществляет постоянный поиск и принимает правильные решения. Отсюда следует, что когнитивное поведение необходимо изучать не в статичной и неизменной среде, как при выработке условных рефлексов, а в динамично и закономерно изменяющейся, когда память не просто фиксирует прошлое, а направлена вперед в будущее.
КРАТКИЙ ИСТОРИЧЕСКИЙ ОБЗОР И КРИТИЧЕСКИЕ ЗАМЕЧАНИЯ К МЕТОДАМ ЭКСПЕРИМЕНТАЛЬНОГО ИЗУЧЕНИЯ КОГНИТИВНОГО ПОВЕДЕНИЯ ЖИВОТНЫХ
В физиологии исследования когнитивного поведения использовались только хорошо известные методики, разработанные бихевиористами в первой половине 20-ого века в Соединенных Штатах Америки. Устройства и лабиринты различной сложности для изучения пищедобывательного или оборонительного поведения, которые никоим образом не соответствуют тому, что животные в них стремятся решить какую-либо проблему – когнитивную задачу.
Известен проблемный ящик Торндайка [23] для исследования инструментального научения “методом проб и ошибок”. Внутрь закрытого подвесной дверкой ящика помещается голодная кошка. Рядом с ящиком находится пища, которая хорошо видна через решетчатую стенку. Исследуя все внутри ящика, кошка случайно обнаруживает, а не находит, свисающую с потолка веревочку, потянув за которую, она открывает подвесную дверку ящика и выходит из него. Именно обнаруживает, поскольку она ничего не искала. Когда ищут, то всегда знают и понимают, что ищут и что надо найти. Это только в сказках бывает: “Пойди туда – не знаю куда, найди то – не знаю что”. Ищущий найдет потому, что точно знает, что он ищет. Со временем этот процесс освобождения кошки из заточения ускоряется научением путем “проб и ошибок”. Так был открыт закон упражнения и закрепления стереотипа, закон эффекта реактивного поведения.
В проблемной камере Скиннера для изучения когнитивного оперантного поведения [23] с автоматической подачей пищевых таблеток крыса при исследовательском поведении натыкается на рычаг или педаль и, случайно нажав на них, получает пищевое подкрепление. Со временем этот пищевой рефлекс закрепляется, ускоряется и совершенствуется. Такое поведение крысы называют оперантным, где операция – это однообразное нажатие лапкой на педаль для получения пищи (оперантный рефлекс). От классического условного рефлекса отличается только тем, что отсутствует начальное действие условного раздражителя.
Эдвард Толмен изучал у крыс поиск выхода из лабиринта различной сложности с повторяющейся Т-образной структурой каналов с одним входом и двумя выходами, где один из выходов всегда ведет в тупик, а другой в продолжение лабиринта и к выходу. Усложнение устройства достигалось увеличением числа этих элементов. Такое научение он связывал с созданием в мозге крыс когнитивной карты. Именно так он назвал план и программу действий и выбора направления движения на каждой Т-образной развилке лабиринта. Критерием научения служило время прохождения крысой лабиринта в направлении пищевой приманки. Задача для крыс в экспериментах Э. Толмена была простой, одноэкстремальной. Ставил ее перед животным и экстенсивно усложнял увеличением числа Т-образных элементов сам экспериментатор [52].
Однако, несмотря на увеличение числа элементов системы лабиринта, вероятность суммарного случайного выбора правильного решения крысой всегда оставалась величиной постоянной равной 0.5 или 50%, как и в одном Т-образном элементе лабиринта.
Широко известно высказывание К. Маркса: “В любой науке ровно столько научного, сколько в ней математики”. В данном контексте математика не подтверждает участия рассудочной деятельности мозга в поиске выхода в таком лабиринте, что свидетельствует о машинальных действиях крыс наугад. Закрепляется только ловкость и быстродействие их выполнения в течение нескольких месяцев ежедневных тренировок прохождения этого лабиринта.
Э. Толмен был первым исследователем когнитивного поведения крыс, который отверг сенсорно-моторную или реактивную природу поиска правильного маршрута и преобразовал простую бихевиористскую формулу поведения S → R в S → O → R, где S – стимул, раздражитель, О – организм, R – реакция. В дальнейшем Э. Толмен заменит в этой формуле понятие “организм” на понятие “когнитивная карта мозга”, существование которой сейчас не только признано физиологами поведения и экспериментальными психологами, но и широко используется в политике и в экономике. В настоящее время у нас ее называют “дорожной картой”, то есть так же, как назвал ее Нобелевский лауреат Джон О’Киф. Однако, по нашему мнению, называть карту “дорожной” не совсем правильно, так как такое название указывает на ее использование только для решения пространственных задач на местности. Вызывает недоумение, когда, говоря о плане или способе решения социальных, экономических и политических задач, а называют это “дорожной картой”.
Такие популярные методики как восьмилучевой лабиринт и водный тест Морриса используются для изучения только пространственной памяти, а потому здесь не приводятся. Не упоминаем также статичные открытые поля и крестообразные лабиринты, поскольку память – не единственный компонент когнитивных способностей. Кроме памяти, когнитивную деятельность мозга детерминируют сознание, внимание, планирование, прогнозирование, принятие решений, понимание причинно следственных отношений.
Все вышеперечисленные методики контролируют только 2 компонента когнитивной деятельности мозга – память и научение, умение. Нет самого главного составляющего когнитивного процесса – опознания, знания – семантического содержания задачи и управления на основе вероятностного прогнозирования. Согласованность памяти, умения и знания подтверждается клиническими наблюдениями. Нарушение когнитивных функций мозга человека включает триаду симптомов: агнозию, апраксию и амнезию.
Физиологию поведения в зарубежных научных исследованиях с давних пор называют экспериментальной психологией, которая долгое время у нас находилась под запретом, поскольку выходила за рамки рефлекторной теории И.П. Павлова об условных рефлексах. Что создало изоляцию от международного научного сообщества, которая на многие десятилетия прервала развитие физиологии и психологии в международном масштабе.
Однако физиология поведения советского времени имела значительные достижения и успехи, как в методологических, так и в концептуальных подходах, вопреки жесткой критике и запретам. Существенным и значительным достижением были физиология активности Н.А. Бернштейна и его “модель потребного будущего” [6], которая предшествует действию, устанавливает связь от прошлого к наиболее вероятному будущему – цели. Под моделью потребного будущего явно угадывается “аппарат акцептор результатов действия” из теории функциональных систем (ТФС).
Системный подход – прогрессивное продолжение теории условных рефлексов, где ответное действие завершается результатом. В рефлекторной теории инициатор поведения – раздражитель. ТФС заменяет раздражитель потребностью, которая в гипоталамусе и коре трансформируется в соответствующую ей мотивацию. ТФС рефлекторную дугу замыкает в кольцо с обратной связью, что превращает ее в функциональную систему, где первым элементом и системообразующим фактором стал результат, который достигается как и в условном рефлексе всегда со 100% вероятностью. ТФС П.К. Анохина – прогрессивное продолжение теории условных рефлексов, но все осталось в рамках сенсорно-моторного и сенсорно-секреторного реактивного поведения, от которого отрекся еще в сороковые годы 20-ого века Эдвард Толмен. “Гипотезы” Ю. Конорского [20] также никак не вписываются в условные рефлексы, поскольку гипотеза является научным предположением или доказательством использования, соответствующей ей определенной методологии.
В рефлекторной теории всем действиям должен предшествовать раздражитель, без которого не может быть ни условного, ни безусловного рефлексов и временной связи. Это сформированный И.П. Павловым руководящий философский принцип строго детерминизма. А поэтому рефлекторная теория не приемлет таких понятий и концептов Беритова И.С. как “психонервное научение” и “образная память” которые были предложены автором уже в 1964 [5] до появления психобихевиоризма и психофизиологии, поскольку исключали первопричину – действие условного или безусловного раздражителя. По И.С. Беритову психонервная активность высших животных и человека преобладает над всеми другими формами обучения и является определяющим фактором поведения. Обучение поиску пищевого подкрепления у собак происходило с одного урока и носило характер одномоментной формы научения, в отличие от многократных повторений при выработке условного рефлекса. Такой однократный опыт легко воспроизводился через месяц и более длительное время.
Отдаленными от условных рефлексов были “экстраполяции”, а тем более “элементарная рассудочная деятельность” Л.В. Крушинского [22]. Все эти научные достижения прошлого века имеют между собой много общего. В современной физиологии поведения эти феномены не что иное, как проявления когнитивных способностей в различных формах поведения и под разными названиями. В современных нейрофизиологических подходах все эти концепции, несмотря на различные названия, имеют общую сущность, обязательным моментом которой является нейрофизиологический знаменатель и механизм – когнитивная карта мозга. В настоящее время она заменила собой такие понятия как “образ”, “гипотезы”, “экстраполяции”, “модель потребного будущего”, “аппарат акцептор результатов действия”, мозговую модель проблемной среды [33] при решении мозгом животных и человека когнитивных задач.
Наша методика экспериментальных исследований когнитивных способностей крыс в свободном поведении поиска интегрирует все эти модели поведения и в то же самое время не присоединяется ни к одной из них. В действиях крыс в проблемной камере есть “гипотезы”, вероятностный прогноз и модель потребного будущего, экстраполяция, психонервный образ, акцептор результатов действия, элементарная рассудочная деятельность. Есть и то, чего не было во всех других методиках, – решение стандартной когнитивной задачи при закономерном информационном изменении среды, где целенаправленные действия могли быть рациональными правильными или, напротив, неразумными, иррациональными, ошибочными, с количественной оценкой результатов на основе вероятностного прогноза.
Общность всех этих достижений прошлой экспериментальной психологии и физиологии поведения заключается в том, что нейрофизиологический механизм этих форм поведения единый – формирование в процессе научения когнитивной карты мозга в гиппокампе и парагипокампальной извилине, открытие которой отмечено Нобелевской премией по физиологии и медицине за 2014 г. Наша модель опытов когнитивного научения и формирования дорожной, или когнитивной, карты подтверждает это открытие реальным поведенческим соответствием гексагонального нейрофизиологического клеточного паттерна в парагипокампальной извилине и функциональной организацией клеток, составляющих эту архитектуру.
Устройство и способ изучения когнитивных способностей крыс в проблемной камере
Известна зависимость темпов и уровня развития науки от новых применяемых методов исследования, высказанная старейшиной физиологов мира: “Часто говорят, и недаром, что наука движется толчками, в зависимости от успехов, делаемых методикой. С каждым шагом методики вперед мы как бы поднимаемся ступенью выше, с которой открывается нам более широкий горизонт с невидимыми ранее предметами” [28].
Рис. 1.
Установка для изучения когнитивных способностей крыс. Включает в себя ГКП с наклоненными внутрь стенками в виде усеченной гексагональной пирамиды с прозрачной крышкой с люком – 1, электростимулятор – 2, монитор с компьютером и регистратором – 3, открытая дверка – 4, заблокированная дверка – 5, столешница – 6, светлые точки над дверками – светодиоды, сигнализирующие ошибки и правильные выходы – 7, электродный пол камеры – 8. Стенки с дверками для выходов из камеры пронумерованы от 1 до 6.
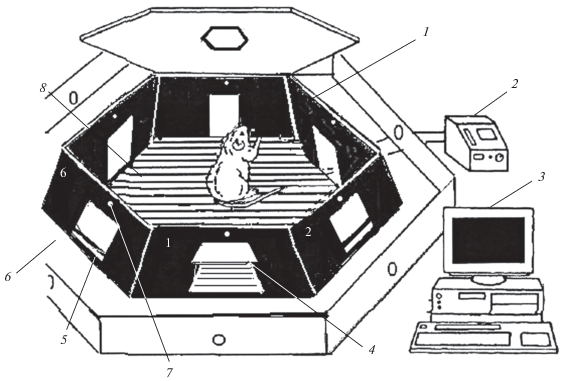
ГПК (рис. 1 ) [9, 10, 16, 41] устанавливают на гексагональный стол с ограждением по периметру. В центре стола выпилено гексагональное отверстие по размерам камеры, на края которого она устанавливается таким образом, чтобы все испражнения тестируемых крыс проваливались через электродную решетку пола в таз, расположенный под столом. Тем самым гигиенические условия в камере близки к идеальным. После тестирования каждой крысы решетка электродного пола протирается ватным тампоном, смоченным спиртом. На рисунке нижняя часть установки не изображена. Камеру закрывают сверху прозрачной крышкой. Над установкой располагается WEB-камера для регистрации всех событий на видео DVR Player 365, 2008-A. Крыса, помещенная в камеру, полностью предоставлена себе и совершает все действия самостоятельно без внешнего воздействия на нее экспериментатора.
Установка в целом (рис. 1 ) включает в себя довольно много элементов, но, говоря современным информационным языком, все компоненты установки являются интерфейсами и гаджетами для удобства наблюдения и регистрации поведения свободного поиска выхода.
Методика подготовки животных к тестированию и сам процесс тестирования неоднократно подробно описаны во всех наших публикациях. Тестирование состоит из шести поисковых циклов, или 6 помещений крысы в камеру, пока без ошибок или с ошибками не будут сделаны каждым тестируемым животным 6 выходов в пространство, окружающее камеру. После выхода из камеры использованная дверка блокируется. Передвижение крысы от центра камеры к выходам из нее называют побежками, в американском и английском варианте их называют экскурсиями [41].
Единственным вмешательством экспериментатора в поведенческий эксперимент является помещение животного в камеру перед каждым новым поведенческим циклом. Животное свободно ищет и выбирает маршрут и направление каждой побежки и тем самым создает себе задачу последовательности и чередования блокирования, т.е. проблему, как не попасть на блокированную дверку и найти оставшиеся незаблокированные дверки. Крыса должна отказаться от безуспешных попыток открыть блокированные дверки и выбрать новое направление побежки и так до последней, которая завершит поисковый цикл. При всех блокированных дверках животных никогда не тестируют, чтобы не вызвать “состояние выученной беспомощности” [49], или отказа от поиска [48], который появляется у большинства животных. Человеку в подобной ситуации также свойственна поведенческая депрессия, которая в крайних случаях может закончиться смертельным исходом при невозможности решить задачу. Поисковая активность является обратной позитивной стороной “медали” в отличие от негативной стороны “выученной беспомощности”, что прекрасно иллюстрирует философский закон единства и борьбы противоположностей. Поисковая активность – такое функциональное состояние организма, когда одержимый им человек или животное демонстрирует “удивительную принципиальную ненасыщаемость в поиске”, что позволяет определять ее уже как потребность в “самом процессе постоянного изменения” и развития [3, 26, 27]. Все вместе это указывает на то, что нет раздельного существования психики и биологии, есть неразрывное диалектическое единство психобиологии.
Когнитивные способности крыс измеряют уровнями вероятности события, где мерой являются величины от 0 до 1.0 или от 0 до 100% согласно теории. Случайный выбор правильного решения закономерно уменьшается, а сложность решения задачи постоянно растет ввиду того, что использованные для выхода дверки блокируют. Такое тестирование представляет иерархию от высоковероятных событий от 100% до маловероятных событий – 16.6%. Так формируется вероятностная структура среды, где крысам необходим прогноз будущих действий, основанный на пространственной памяти пройденного пути в целенаправленном поведении.
Вероятность случайного выбора правильного решения в экспериментах Толмена всегда оставалась величиной постоянной, равной 50% как в начале, так и в конце лабиринта, сколько бы не было таких Т-образных развилок. В нашем способе вероятность случайного выбора и уровень вероятности события не являются постоянной величиной. В первом поисковом цикле вероятность ошибки равна 0. Во втором цикле вероятность совершения ошибки равна 1/6 или (16.6%); в третьем цикле – 2/6 или (33.3%); в четвертом цикле – 3/6 или (50%); в пятом – 4/6, или (66.3%); в шестом – 5/6, или (83.3%). Последний шестой цикл завершает тестирование особи, после чего можно рассчитать средний уровень величины когнитивного показателя. Оценку когнитивных способностей по когнитивному показателю (КП) производили двумя способами: 1. Расчет средней величины без учета вероятности случайного выбора. 2. Расчет средней величины КП с учетом вероятности случайного выбора.
Первый способ исчисления величины среднего КП: сумму всех безошибочных и ошибочных побежек принимают за 100%, правильных побежек только 6 и их принимают за х. Вот пример такого расчета КП, взятый из реального тестирования. Сумма всех побежек с правильными и неправильными выборами направления равна 17, что составляет 100%, из них правильных может быть только 6. 17–100, 6 – х, 600/17 = 35.29%, то есть КП ниже 50%, что свидетельствует об отсутствии познавательных способностей.
Второй способ вычисления величины среднего КП с учетом изменения вероятности случайного выбора устраняет неточность расчета КП при первом способе и позволяет более объективно и достоверно оценить когнитивные способности тестируемого животного. В первом цикле любая побежка эффективна, вероятность всегда равна 100%. Во втором цикле вероятность сделать ошибку равна только 16.6%, в третьем 33.3%. В рубежном четвертом цикле вероятность сделать ошибку равна 50%, а в пятом цикле вероятность сделать ошибочный выбор увеличивается до 66.3%. В шестом цикле она достигает 83.3%, и тестирование прекращается, так как наступает безвыходная ситуация.
В том же опыте используют те же 17 побежек, но теперь среднюю величину КП рассчитывают с учетом номеров поискового цикла при переходе высоковероятных событий к событиям маловероятным. Те же 6 правильных побежек и 11 ошибочных, т.е. общая сумма = 17. Чаще всего ошибки совершают на 5 и 6 циклах при низкой вероятности. В данном случае именно так. На 5 цикле 5 ошибок, на 6 цикле 6. Цена 1 ошибки на 5 цикле = = 66.6%, на 6 цикле 83.3%, вычисляем и находим среднюю величину КП. Общая сумма шести безошибочных побежек равна 600%. В 5 и 6 циклах стоимости ошибок соответствует 66.6% и 83.3%, умножаем первую на 5, а вторую величину на 6 ошибок, суммируем. Итоговую величину делим на 17 – общее число побежек и получаем точный КП, равный 84.2%: 600 + 66.6 × 5 + 83.3 × 6/12 = = 84.2%. Это указывает на относительно высокий уровень когнитивных способностей, который оказался при точном расчете выше среднего. Общая формула на все варианты оценки КП:
КП = 600% + N2х 16.6% +N3x 33.3% + N4x 50% + + N5x 66.3% + N6x 83.3%, где N2, N3, N4, N5, N6 количество ошибок и номер соответствующего поискового цикла, 600% – цена 6 правильных побежек. Ценность имеющейся в мозге каждой крысы необходимой информации (осведомленности) оценивается изменением вероятности достижения цели [18].
КП величиной до 50% указывает, что выбор случаен и поведение поиска неуправляемо. КП, превышающий 50%, указывает на тенденцию к обучению рациональному поиску правильного пути, знаменует начало приобретения опыта и процесса формирования когнитивной карты. 100% уровень КП полностью исключает вероятность случайного выбора и указывает на высокий уровень когнитивных способностей, наличие рассудочной деятельности у крыс и окончательно сформированную когнитивную карту. Такая оценка вероятностного прогноза в проблемной камере соответствует тонкому измерению пространственного интеллекта крыс. Большинство исследователей поведения крыс относят их к классу интеллектуально одаренных животных, что математически достоверно подтверждают эти опыты. С ошибками или без ошибок, но выход из камеры для тестируемой крысы в конечном итоге обеспечен всегда, что формирует и сохраняет эту приобретенную поисковую активность в общей структуре поведения на длительное время. После длительных перерывов в тестированиях в несколько месяцев крысы демонстрируют приобретенное поведение активного поиска без включения аверсивной стимуляции [10].
АНАЛИЗ И ОБСУЖДЕНИЕ ПСИХОФИЗИОЛОГИЧЕСКИХ МЕХАНИЗМОВ ФОРМИРОВАНИЯ КОГНИТИВНОГО ПОВЕДЕНИЯ КРЫС В ПРОБЛЕМНЫХ СИТУАЦИЯХ
Электростимуляция лап крыс в камере используется только для формирования мотивации активного избегания в начальной стадии выработки этого условного рефлекса. После его упрочения он легко трансформируется в поисковое поведение и необходимость электростимуляции отпадает. Крысы после помещения в камеру сами активно и самопроизвольно работают: ищут из нее выход. Примечателен тот факт, что у крыс никогда не отмечается развития утомления или насыщаемости этой поисковой активностью, независимо от того, что она будет безупречно успешной и результативной или с частыми ошибочными побежками. Такое явление мы называем экспериментально сформированной поисковой активностью в ГПК. Экспериментальные крысы долгое время сохраняют и актуализируют эту форму приобретенного поискового поведения. Такое поведение – это сформированный план и программа всех действий, последовательности и чередования во времени, которые приведут крысу к правильному решению и оптимальному результату, но только не у всех тестируемых крыс. Динамика и закономерность закрытия использованных для выхода дверок, как психологическое содержание, должна быть понята тестируемой крысой, о чем будет свидетельствовать правильный выбор пути, ведущий к цели более коротким и эффективным путем. Закономерность всех изменений в ГПК, понятная экспериментатору, должна стать такой же понятной подопытной крысе: нельзя ходить в одну и ту же дверку дважды, для каждой следующей побежки необходимо находить другой выход. На то, что этот принцип усвоен животным, указывают случаи, когда отмечается 100% уровень безошибочных побежек во всех поисковых циклах. Такой результат демонстрируют не более 12–15% тестируемых крыс [41].
Процесс познания, который управляет поведением при всевозможных, но закономерных изменениях внешней среды и обстоятельств, включает в себя выбор и оценку альтернативных выходов из проблемной камеры. Такое субъективное состояние крыс возможно при условии, когда мозг тестируемого животного имеет достаточный объем кратковременной и долговременной памяти, усваивает закономерность событий, происходящих при изменении проблемной среды, что можно считать реализацией задачи пространственного научения и формированием когнитивной карты.
Для такой высшей нервной деятельности необходимы ключевые структуры головного мозга: передний мозг и его префронтальная кора [7], гипоталамус как мотивационный центр, ретикулярная формация ствола мозга [24], гиппокамп и парагиппокампальная извилина, как навигационная система мозга крысы (интерфейс) – создатель и носитель когнитивной карты [46, 38 ] .
Для осуществления результативного поведения поиска выхода из камеры постоянно востребованы все три основных компонента психики: 1. Гнозис – процесс познания закономерностей изменений в проблемной камере. 2. Праксис – умение выполнять целенаправленные побежки и открывать неблокированную дверку головой или лапками, переход к неблокированным дверкам. 3. Мнезис – необходимый объем кратковременной и долговременной памяти. А все это вместе взятое есть психика, существование которой не признается многими физиологами даже у интеллектуально одаренных животных.
В прикладном аспекте экспериментальные исследования когнитивного поведения на крысах имеют большое практическое значение для клинической нейропсихологии, понимания механизмов когнитивных процессов и, возможно, будут способствовать разработке способов коррекции когнитивных нарушений у человека. Если аффективные расстройства психики относительно излечимы, то медицина когнитивных расстройств остается долгие века практически беспомощной при травмах мозга, старческом маразме, болезни Альцгеймера и других психических болезнях, включая сюда наркомании и алкоголизм [7].
ГПК и предложенный нами метод использованы для контроля развивающейся пространственной памяти [53], в фармакологии поведения для точной и тонкой проверки эффективности когнотропных (ноотропных) препаратов [4, 14, 17], а также для изучения влияния минерального состава пищи на когнитивные способности [28, 39], при экспериментальном стрессе [15]. Были получены данные о том, что познавательная деятельность мозга существенно отличается при различных потребностях и соответствующих им пищевой, питьевой и оборонительной мотивациях [13]. Методика позволила дифференцировать исследовательское поведение в открытом поле и крестообразном лабиринте от поведения поиска [11].
Функциональное состояние крыс, которое обеспечивает результативное поведение в сложной проблемной ситуации, обуславливает переключающая функция эмоций по П.В. Симонову [29, 31]. Оборонительное поведение крыс, где стимулом являлся аверсивный раздражитель, легко трансформируется в активное поисковое поведение выхода из проблемной ситуации. В этом случае детерминантой поведения становятся, согласно формуле П.В. Симонова, не избегание или избавление, а мотивация и положительные эмоции от безошибочных правильно и точно выполняемых действий: Э = – П × (Ин – Ис), где Э – эмоции, П – потребность избежать действия аверсивного раздражителя, Ин – информация необходимая – знание закономерности блокирования использованных для выхода дверок, Ис – имеющиеся знания и понимание этих закономерностей. При этом Ис – это и есть сознание, в том понимании П.В. Симонова, что сознание – это знание вместе со всеми (в данном случае у экспериментатора и тестируемой крысы, когда имеет место 100% соответствие правильных действий). “Потребность познания ведет свое происхождение от универсальной потребности в информации, изначально присущей всему живому, наряду с потребностью вещества и энергии. Удовлетворение любой потребности требует информации о путях и способах достижения цели. Вместе с тем существует потребность в информации, как стремление к новому, ранее неизвестному, безотносительно к его прагматическому значению в смысле каких-либо биологических и социальных нужд. Филогенетические предпосылки этой потребности обнаруживаются уже у животных” [30].
Во время приобретения и накопления опыта (тестирований) происходит увеличение прагматической информации в мозге крысы – Ис, а потому разница (Ин – Ис) становится отрицательной, а эмоции при успешном и результативном поиске в проблемной камере положительными. Разница (Ин – Ис) есть оценка вероятности удовлетворения потребности. Именно она детерминирует результативность поиска в проблемной камере, что полностью соответствует формуле применительно к данной ситуации [50]. Когнитивная, или познавательная информационная, часть поведения поиска имеет обязательное эмоциональное сопровождение, которое форсирует и стабилизирует интенсивность действий. В наших экспериментах это поступательное стремление всех тестируемых крыс к 100% вероятности достижения результата.
Человеку и животным свойственно врожденное стремление к совершенствованию, приобретению опыта, которому нет предела и насыщения [48]. У экспериментальных крыс поисковая активность остается практически неизменной и постоянно присутствующей в общей структуре всех других специфических форм поведения.
В сравнительном аспекте уровень когнитивных способностей одной крысы может быть оценен уровнем когнитивных способностей другой крысы. Как в шахматных турнирах рейтинг чемпиона проверяется уровнем рейтинга нового претендента. У крыс чисто рейтинговая оценка уровня когнитивных способностей заключается в том, что в экспериментальные и контрольные тестируемые группы подбираются животные одного возраста, веса, пола. Все животные имеют одинаковые условия содержания, одинаковое число сочетаний, одинаковую подготовку и число тестирований, и время проведения опытов. Различными будут только уровни когнитивных способностей. У человека 75% спектра интеллектуальных способностей мозга принадлежит, прежде всего, умению решать пространственные задачи [1]. Скорее всего, это так же важно и для высших животных, если только не в большей степени. Общепризнанная интеллектуальная игра в шахматы являет собой пример постоянного решения навигационных пространственных задач в динамично изменяющейся среде шахматной доски. Обязательным моментом всех действий шахматиста является вероятностный прогноз своих действий и будущих ответных актов противника. Только правильное передвижение фигур и предвидение шагов противника ведут к победе, неправильные передвижения приводят к поражению. Поведение крыс в проблемной камере на примитивном уровне моделирует такую пространственную игру в ГПК. Каждая безошибочная побежка в динамично и закономерно изменяющейся среде основана на вероятностном прогнозе предстоящих действий, точность которых обеспечивает только достаточный уровень когнитивных способностей и сформированная когнитивная карта в гексагональной координатной сети медиальной энторинальной коры парагиппокампальной извилины.
КОГНИТИВНЫЕ СПОСОБНОСТИ И КОГНИТИВНАЯ КАРТА В СОВРЕМЕННЫХ НЕЙРОФИЗИОЛОГИЧЕСКИХ ИССЛЕДОВАНИЯХ
Первым обнаружил клетки чувства пространственного местонахождения Джон O'Киф в 1971 г., которые он назвал картой места. Он использовал методику регистрации залповой электрической активности нейронов гиппокампа у крыс в свободном поведении в трехлучевом лабиринте небольших размеров [45–47]. Тем самым нейрофизиологически был подтвержден постулат Эдварда Толмена о существовании когнитивной карты в гиппокампе при свободном поведении у крыс. Затем ученики Д. O'Кифа Мэй-Бритт и Эдвард Мозеры в свободном поведении у крыс обнаружили клетки места и клетки координационной сети – навигационной системы в парагиппокампальной извилине. За эти открытия всем трем авторам была присуждена Нобелевская премия по физиологии и медицине за 2014 г. Эти нейрофизиологические открытия показали, как представлена на клеточном уровне когнитивная карта мозга в гиппокампе и парагиппокампальной извилине.
Дальнейшие исследования показали, что в функциональный состав карты места входят клетки пространственной навигации и кодирования направления при свободном движении c широкой памятью окружающего пространства. Это дало возможность назвать интеграцию этих клеток когнитивной картой мозга, как ее называл Э. Толмен [52]. Расширение исследований авторов Нобелевской премии Джона О’Киф, Мэй-Бритт и Юджина Мозер привело к новым достижениям и открытиям в этой области. Рядом с гиппокампом в медиальной энторинальной коре парагиппокампальной извилины выявлена самостоятельная и независимая от клеток места новая интеграция нервных элементов, которая имела свои особые функции [42, 43]. Клетки интегрировались в гексагональную архитектуру, функциональная организация которой была названа координатной сеткой [51]. Клеточная система координат необходима для точной навигации и управления движением в заданном направлении. Вместе с гиппокампальными клетками места энторинальные клетки функционально активны при поиске и выборе правильного пути, решения пространственных навигационных задач и формирования когнитивной карты для целенаправленного передвижения [37, 42]. Навигация связана с чувством расстояния и управлением движениями, основанными на знании предыдущих положений. Определение той или иной части пространства обеспечивают многие функционально специфические типы клеток координатной сетки. Главные клетки направления и краевые клетки в средней энторинальной коре формируют когнитивную карту внешнего пространства [35]. Эксперименты с новорожденными крысятами подтвердили предположение Иммануила Канта о врожденности навигационных способностей у животных [34]. Гексагональная координатная сетка имеет дорзо-вентральный градиент размерности, обеспечивающий измерение линейных расстояний от 30 см до 1 метра и углового измерения окружающего пространства со стандартным размером угла в 60° [44].
Фиксацию в памяти пространственных закономерностей изменений при поиске правильного пути обеспечивают входящее в эту гексагональную координатную сетку подразделение различных по функции клеток.
Рис. 2.
Справа схема головного мозга крысы с изображением гиппокампа и парагиппокампальной извилины. Левая сторона - гексагональная структура координатной сетки. Полный пространственный поведенческий аналог с центральным входным отверстием и шестью выходными дверками ГПК. Интернет ресурс [22].

Рис. 2 заимствован из публикаций Д. O'Киф, М.-Б. Мозер и Э. Мозер, где изображены опорные точки клеток координатной сетки слева и мозг крысы с парагиппокампальной извилиной энторинальной коры (рис. 2 ). Все вместе клетки образуют обобщенную гексагональную сеть, обозначенную на рисунке черной жирной линией, соединяющей 6 опорных точек. Эта нейрофизиологическая когнитивная карта полностью соответствует гексагональной структуре проблемной камеры и расположению ее 6 выходных дверок. Седьмая опорная точка соответствует центральному входному отверстию люка камеры, откуда начинаются побежки (рис.1 ). Трассы возбуждения клеток в парагиппокампальной извилине в экспериментах М.-Б. и Э. Мозер соотносятся с побежками крыс в проблемной камере от центра к выбранной дверке со стандартным углом отклонения в 60° (рис. 2 ). Именно так выглядела бы нейрональная картина реальных побежек крыс в проблемной камере при безошибочных 100% успешных побежках. Внешнее идеальное соответствие заключается в том, что интегральная система включает в себя все те клетки, которые необходимы для научения правильной навигации крыс в проблемной камере. В экспериментах М.-Б. и Э. Мозер выявлены нейроны, действующие как компас, когда голова животного устанавливается в определенном направлении при секторе угла в 60°. Структура гексагональной сети в парагиппокампальной извилине мозга включает в себя клетки, активирующиеся при столкновениях со стенками (в нашей методике – это удар головой в блокированную дверку камеры) [43]. Краевые клетки отображают геометрию боковых стенок камеры, главные клетки управляют поведением, выполняют интегративную и комбинаторную функцию [37].
Важно отметить, что гиппокампальные нейроны кодируют потоки информации не только о настоящем и прошлом, но и о предполагаемом будущем местонахождении. Такое будущее отражается в возбуждении нейронов места, в котором с высокой вероятностью крыса будет находиться после принятия решения к действию при выполнении задачи поиска пищевой приманки [34]. Наличие нейронов с такой эпизодической памятью, обращенной в будущее, убедительно свидетельствует о наличии модели потребного будущего, или вероятностном прогнозировании на нейрональном уровне. Предвидение и вынесение функции вперед на отрезке времени и есть управление, когда установлена непрерывная связь настоящего с прошлым и неизбежным будущим на основе прошлого опыта [36], что подтверждает крылатое выражение “Управлять значит предвидеть”.
Наше устройство и способ не только соответствуют по форме гексагональной координатной сетке в парагиппокампальной извилине, но и качественному функциональному составу клеток. Побежки крыс из центра к дверкам камеры имеют шестикратную вращательную симметрию со стандартным масштабом геометрии угла в 60° при полном пространственном обзоре камеры изнутри в 360° [37, 38]. Когнитивная карта маршрутов побежек в камере состоит из налагающихся друг на друга двух равносторонних треугольников при 100% уровне когнитивных способностей. Свободное поведение крыс поиска выхода при 100% результативности на видеосъемке опыта невероятно и абсолютно идентично гексагональной когнитивной карте мозга в парагиппокампальной извилине. Это указывает на полное совпадение структурно-функциональной нейрональной единицы – когнитивной карты и ее внешнего навигационного паттерна. Этот факт подтверждает реальность существования когнитивной карты вместо временной связи в коре двух рефлекторных дуг и справедливость гипотезы Э. Толмена и окончательную победу физиологии активности в психофизиологии.
Такое совпадение нейрофизиологической картины возбужденных клеток когнитивной карты мозга с реальным поведенческим паттерном не может быть случайным при достижении 100% эффективности поведения генерализованного поиска.
ЗАКЛЮЧЕНИЕ
Открытия Нобелевских лауреатов М.-Б. и Э. Мозер изменяют понимание (парадигму) того, как интеграции специализированных нейронов обеспечивают высшие сенсорные функции мозга – восприятие и память. Клетки чувства места и навигационная система мозга в гиппокампе и парагиппокампальной извилине обслуживают когнитивные функции мозга в целом. Предложенные нами устройство и способ тестирования подтверждают на поведенческом уровне существование гексагональной структуры нейрональной сети у крыс. Методика тестирования в ГПК позволяет дать количественную оценку уровня когнитивных способностей, необходимых для решения сложных навигационных задач. Вероятностный прогноз закономерных изменений среды обеспечивает результативный поиск выхода из камеры. Внутренние нейрофизиологические паттерны координатной сети нейронов соотносятся с внешними действиями крыс в гексагональной среде проблемной камеры, которые контролируют процесс формирования когнитивной карты от начала и до завершения при 100% результате. Эксперименты указывают на то, что упорядоченная гексагональная координатная сеть в парагиппокампальной извилине является обязательным нейрофизиологическим атрибутом для навигации в закономерно и динамично изменяющейся среде. Вероятностное прогнозирование обеспечивает планирование будущих действий, решение логических задач и формирование программы. Такая интегративная работа мозга в целом немыслима без потребности, мотивации и эмоций - основных инициаторов и участников всех видов и форм поведения, включая успешную навигацию в гексагональной проблемной камере.
Список литературы
Айзенк Г. Интеллект: новый взгляд // Вопросы психологии. 1995. № 1. С. 111–129.
Анохин П.К. Избранные труды. Философские аспекты теории функциональной системы. М.: Наука, 1978. 399 с.
Аршавский В.В., Ротенберг В.С. Поисковая активность и ее влияние на экспериментальную и клиническую патологию // Журн. высш. нервн. деят-ти. 1976. Т. 26. № 2. С. 424–428.
Баталова Т.А. Психобиологические особенности при комплексной коррекции у крыс: автореф. дис. … д-ра биол. наук. Благовещенск, 2011. 39 с.
Беритов И.С. Вопросы высшей нервной деятельности и ее развития // Избранные труды. М.Л.: Наука, 1964. Т. 3. 479 с.
Бернштейн Н.А. Физиология движений и активности. М.: Наука, 1966. 349 с.
Гольдберг Э. Управляющий мозг: Лобные доли, лидерство и цивилизация. М.: Смысл, 2003. 335 с.
Григорьев Н.Р. Автор. свид-во СССР № 1776385 // Бюл. изобр. 1992. № 43. 1–8 с.
Григорьев Н.Р. Метод исследования поисковой активности и отказа от поиска в эксперименте у крыс // Журн. высш. нервн. деят-ти. 1996. Т. 46. № 2. С. 400–405.
Григорьев Н.Р. Функциональная организация поисковой активности при пищевом и оборонительном поведении // Журн. высш. нервн. деят-ти. 1998. Т. 48. № 1. С. 75–83.
Григорьев Н.Р., Кириченко Е.Ф., Темпер Ю.Б., Чербикова Г.Е. Динамика интегративных параметров ориентировочно-исследовательского и поискового поведения у крыс // Журн. высш. нервн. деят-ти. 1998. Т. 48. № 5. С. 868–876.
Григорьев Н.Р. Функциональная организация поисковой активности (экспериментальное исследование): автореф. … дис. д-ра мед. наук. Чита, 1998. 41 с.
Григорьев Н.Р., Артемчук С.Ф., Чербикова Г.Е. Параметры поисковой активности в структуре пищевого, оборонительного и питьевого поведения (сравнительный анализ) // Журн. высш. нервн. деят-ти. 2001. Т. 50. № 3. С. 383–385.
Григорьев Н.Р., Тиханов В.И., Пластинин М.Л. Поисковая активность и ее фармакологическая регуляция изотиорбамином // Дальневост. мед. журн. 2001. № 3. С. 37–41.
Григорьев Н.Р., Сергиевич А.А. Динамика показателей оборонительного поведения у крыс при психоэмоциональном стрессе на фоне применения целита // Фундам. исслед. 2012. № 5–1. С. 189–192.
Григорьев Н.Р., Чербикова Г.Е., Юрьев Е.Ю. Пат. РФ № 2432902 // Бюл. изобр. 2011. № 31.
Григорьев Н.Р., Ли-Тингли, Лишао-Юань. Когнитивные способности крыс в оперантном поведении активного избегания в различные этапы онтогенеза и при воздействии некоторых препаратов // Журн. высш. нервн. деят-ти. 2014. Т. 64. № 3. С. 314–323.
Григорьев Н.Р. Пат. РФ № 2604048 // Бюл. изобр. 2016. № 34.
Григорьев Н.Р. Физиологические истоки интеллекта животных и человека. LAP Academic Publishing. 2017. 133 с.
Конорски Ю. Интегративная деятельность мозга. М.: Мир, 1970. 412 с.
Коржова В. GPS в нашем мозге: Нобелевская премия по физиологии и медицине 2014. https://biomolecula.ru/articles/gps-v-nashem-mozge-nobelevskaia-premiia-po-fiziologii-i-meditsine-2014.
Крушинский Л.В. Биологические основы рассудочной деятельности. М.: МГУ, 1986. 270 с.
Мак-Фарленд Д. Поведение животных. Психобиология, этология и эволюция. М.: Мир, 1988. 518 с.
Мэгун Г. Бодрствующий мозг. М.: Мир, 1965. 210 с.
Павлов И.П. Двадцатилетний опыт объективного изучения высшей нервной деятельности (поведения) животных. М.: Наука, 1973. 659 с.
Ротенберг В.С., Аршавский А.А. Стресс и поисковая активность // Вопросы философии. 1980. № 4. С. 117–127.
Ротенберг В.С., Аршавский В.В. Поисковая активность и адаптация // М.: Наука, 1984. 192 с.
Сергиевич А.А., Григорьев Н.Р. Поведенческие параметры у животных из различных типологических групп при влиянии минерального фактора // Вестн. новых мед. технологий. 2012. Т. 19. № 2. С. 217–221.
Симонов П.В. Потребностно-информационная организация интегративной деятельности мозга // Журн. высш. нервн. деят-ти. 1979. Т. 29. № 3. С. 467–482.
Симонов П.В. Темперамент. Характер. Личность. М.: Наука, 1984. 159 с.
Симонов П.В. Лекции о работе головного мозга. Потребностно-информационная теория высшей нервной деятельности. М.: Ин-т психологии РАН, 1998. 96 с.
Фейгенберг И.М. Вероятностное прогнозирование в деятельности человека и поведении животных. М.: Ньюдиамед, 2010. 179 с.
Шамис А.Л. Поведение. Восприятие. Мышление. Проблемы создания искусственного интеллекта. М.: Научная и учебная литература УРСС, 2004. 223 с.
Bjerknes T.L., Witter M.P., Moser E.I., Moser M.B. Development of the spatial representation system in the rat // Science. 2010. V. 328. P. 1576–1580.
Bjerknes T.L., Moser E.I., Moser M.B. Representation of geometric borders in the developing rat // Neuron. 2014. V. 82. № 1. P. 71–78.
Ferbinteanu J., Shapiro M.L. Prospective and Retrospective Memory Coding in the Hippocampus // Neuron. 2003. V. 40. № 6. P. 1227–1239.
Fyhn M., Molden S., Witter M.P., Moser E.I., Moser M.B. Spatial representation in the entorhinal cortex // Science. 2004. V. 305. P. 1258–1264.
Fyhn M., Hafting T., Treves A. at al. Hippocampal remapping and grid realignment in entorhinal cortex // 2007. Nature. V. 446. P. 190–194.
Golokhvast K.S., Sergievich A.A., Grigorev N.R. Geophagy (Rock eating). Experimental stress and cognitive ideosyncrasy // Asian Pacific J. of Tropical Biomed. 2014. V. 4. № 1. P. 930–934.
Grigor’ev N.R., Cherbikova G.E. Experimental research model for the search of activity and cognitive abilities in rats // Intern. J. Psychophysiology. 2008. V. 69. № 3. P. 250.
Grigor’ev N.R., Batalova T.A., Kirichenko E.F., Cherbikova G.E., Sergievich A.A. Typological Features in the Behavior of Rats // Neuroscience and Behavioral Physiology. 2008. V. 38. № 6. P. 597–603.
Hafting T., Fyhn M., Molden S., Moser M.B., Moser E.I. Microstructure ofaspatial map in the entorhinal cortex // Nature. 2005. V. 436. P. 801–806.
Hafting T., Fyhn M., Bonnevie T., Moser M.B., Moser E.I. Hippocampus-independent phase precession in entorhinal grid cells // Nature. 2008. V. 453. P. 1248–1252.
Kjelstrup K.B., Solstad T., Brun V.H. Finite scale of spatial representation in the hippocampus // Science. 2008. V. 32. P. 1140–1143.
O'Keefe J., Dostrovsky J. The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely moving rat // Brain res. 1971. V. 34. № 1. P. 171–175.
O'Keefe J. Place units in the hippocampus of the freely moving rat // Experimental neurology. 1976. V. 51. № 1. P. 78–109.
O’Keefe J., Nadel L. The hippocampus cognitive map.The hippocampus spatial map. Preliminary evidence from unit activity in the freely-moving rat // Brain Res. 1978. V. 34. P. 171–175.
Rotenberg V.S. Search activity concept: an experimental background // Tel-Aviv University. Actyvitas Nervosa Superior. 2009. V. 51. № 1. P. 12–44.
Seligman M.E.P. Helplessness: On depression, development and death. San Francisco. Freeman. 1975. 275 p.
Simonov P.V. Brain Mechanisms of Emotion. From: Conceptual Advances in Russian Neuroscience. London: Gordon and Breach Science Pablishers. 1999. P. 113–127.
Solstad T., Boccara C.N., Kropff E. et al. Representation of geometric borders in the entorhinal cortex // Science. 2008. V. 322. P. 1865–1868.
Tolman E.C. Cognitive maps in rats and men // Psychol. Rev. 1948. V. 55. P. 189–208.
Xu R.X., Grigoryev N.R., Li-Tingli et al. Development of hexagonal maze procedure for evoluating memory in rats // Biomed. Rep. 2013. № 1. P. 134–138.
Дополнительные материалы отсутствуют.
Инструменты
Успехи физиологических наук