

Успехи физиологических наук, 2021, T. 52, № 1, стр. 77-89
Дискретность и континуальность информации в сознании
М. Е. Баклушев a, *, Г. А. Иваницкий b, **
a Национальный исследовательский университет “Высшая школа экономики”, Институт когнитивных нейронаук
Москва, Россия
b Институт высшей нервной деятельности и нейрофизиологии РАН
Москва, Россия
* E-mail: baklushevm@mail.ru
** E-mail: geivanit@mail.ru
Поступила в редакцию 04.07.2020
После доработки 10.08.2020
Принята к публикации 03.09.2020
Аннотация
В статье рассматривается проблема организации информации в сознании человека, в частности, принципиальная возможность выделения в массиве воспринимаемой и обрабатываемой информации каких-либо структурных единиц, возможность выделения наименьшей, “элементарной” единицы информации применительно к сознанию. Рассматривается процесс восприятия как процесс, в ходе которого получаемая от рецепторов информация последовательно обобщается, сравнивается с накопленным ранее опытом и становится материалом для формирования понятий высокого уровня абстракции. Рассматривается процесс “понимания” понятий, как процесс обратный их формированию – процесс, в ходе которого столкновение сознания с ранее усвоенным понятием приводит к повторному развертыванию многочисленных образов и ассоциаций, которые ранее стали материалом для его формирования. Рассматривается проблема выделения в массиве информации ключевых характеристик, наиболее значимых ассоциативных связей применительно к нормальной психике и патологии шизофренического спектра.
1. ВСТУПЛЕНИЕ
Главная задача, стоящая перед нейрофизиологией, – понять, как физиологические процессы, протекающие в мозге, соотносятся с нашей субъективной реальностью. Основными методами изучения деятельности мозга на сегодняшний день являются ЭЭГ, МЭГ и фМРТ. В конкретных экспериментах, как правило, происходит анализ какого-либо достаточно сложного когнитивного процесса этими методами, то есть, рассматривается вопрос о том, как изменяются, например, показатели ЭЭГ при осуществлении испытуемым той или иной задачи. При этом достаточно сложный когнитивный процесс соотносится с достаточно сложными изменениями, наблюдаемыми в ЭЭГ или фМРТ, часто называемыми словом “паттерн”. Изучение паттернов тех или иных когнитивных процессов сделало возможным так называемое “чтение мозга” [3]. Стало возможно распознать по паттернам ЭЭГ, какой именно когнитивный процесс протекает в мозге в данный момент. И, тем не менее, физиологический смысл наблюдаемых паттернов, конкретный механизм, обеспечивающий решение тех или иных когнитивных задач остается неясен.
В этой связи представляется интересным и полезным рассмотрение когнитивной деятельности с точки зрения наиболее элементарных “когнитивных актов”, простых операций, которые производит наш мозг с информацией для обеспечения всего многообразия интеллектуальной деятельности.
Понятие информации и информационного процесса достаточно давно используются для описания соотношения между психическим и физиологическим. Подразумевается, что субъективная реальность (психика) и объективная реальность (физиология) являются двумя проявлениями, аспектами информационного процесса. Таким образом, для решения психофизиологической проблемы полезным становится рассмотрение операций, производимых мозгом в отношении информации. Однако, перед тем как рассматривать операции, целесообразно прояснить и конкретизировать само понятие информации применительно к нейрофизиологии и психике.
В вычислительной технике информация является измеримой величиной, ее количество измеряется в битах. За битом стоит “0” или “1” цифрового кода, проведение или отсутствие проведения электрического сигнала. Применительно к человеческому мозгу и сознанию выделение такой “элементарной единицы” информации сопряжено с рядом трудностей. Прежде всего, проблематично выделение “элементарной единицы” информации применительно к нашей субъективной реальности. Поиск такой “элементарной единицы” порождает вопрос о том, насколько вообще можно разделить информацию, содержащуюся и возникающую в нашем сознании на какие-то фрагменты.
С одной стороны, очевидно, что мы выделяем отдельные понятия, образы и категории понятий, отдельные признаки предметов и понятий – в этом плане информации в нашем сознании присуще определенная дискретность. С другой стороны, попытка определения любого, даже очень простого понятия (ботинок, еда, шар и т.д.) приводит к необходимости задействования множества других понятий и, в конечном итоге, всего накопленного индивидом опыта. Мы определяем ботинок как “обувь”, как “изделие из кожи”, связываем его с понятиями “ходьба”, “ноги”, “одежда”. Эти понятия в свою очередь могут быть определены лишь через другие понятия. Таким образом, каждое понятие “тянет за собой” цепочки ассоциаций, связывающие его со всем остальным когнитивным опытом. В этом плане, информация в нашем сознании неделима и предстает в виде целостного когнитивного опыта, где все понятия ассоциативно связаны и фактически неотделимы друг от друга.
Каким же образом сочетается возможность оперирования отдельными понятиями и невозможность их понимания без привлечения всего когнитивного опыта?
Для того, чтобы ответить на этот вопрос, мы рассмотрим два процесса – процесс формирования понятий в ходе восприятия реальности и процесс раскрытия сути понятий при их представлении и использовании в контексте, который мы в данной статье будем называть “пониманием” понятий.
2. ФОРМИРОВАНИЕ ПОНЯТИЙ
Понятия являются результатом обобщения информации, поступающей от органов чувств в течение жизни. Поэтому мы начнем рассмотрение их формирования с процесса восприятия. Мы рассмотрим этот процесс на примере зрительного анализатора.
Восприятие
Понятия имеют смысл, а информация, поступающая к рецепторам, сама по себе бессмысленна. Осмысленной она становится тогда, когда сравнивается с уже существующим человеческим опытом. Именно тогда человек осознает эту информацию, определяет свое к ней отношение или, иначе говоря, значение этой информации для себя. Этот процесс осознания информации описан в теории информационного синтеза [2]. Смысл этой теории в том, что всякая новая информация, поступающая в мозг, сравнивается с информацией, хранящейся в памяти, и субъективное ощущение от увиденного или услышанного является результатом этого сравнения, или, иначе говоря, “синтеза” новой информации с уже имеющимся опытом.
Очевидно, что для того, чтобы что-то с чем-то сравнивать или “синтезировать”, в информационном потоке, поступающем к рецепторам, должны быть выделены некоторые фрагменты. Фрагменты информационного потока могут соотноситься с фрагментами информации, хранящейся в памяти. Таким образом, необходимой становится дискретизация информационного потока, разделение информации воспринимаемой человеком на составные части.
Дискретизация информации при восприятии визуальной сцены
Этот процесс дискретизации действительно имеет место и начинается еще до поступления информации в кору, уже на уровне сетчатки. Основой для него является разница в физических характеристиках частей визуальной сцены. В визуальной сцене, наблюдаемой человеком, изначально присутствуют пятна разного цвета, разной освещенности, разной формы. Именно на контрастировании, усилении этой разницы построена обработка зрительной информации в сетчатке, латеральном коленчатом теле и первичной зрительной коре [4] Обеспечивается он наличием клеток, избирательно реагирующих на освещение центра и периферии, а также механизмом латерального торможения. Благодаря этому происходит контрастирование изображения по цвету, что создает предпосылки для выделения краев объектов, то есть разделения целостной визуальной сцены на составные части. В первичной зрительной коре происходит не только дальнейшее контрастирование изображения по цвету, но и распознавание пространственной ориентации зрительных объектов относительно вертикальной и горизонтальной оси [73, 78].
“Сравнение” с памятью
Начальный процесс восприятия зрительной сцены достаточно хорошо описан в литературе. Он представляет собой ретинотопическое проецирование изображения в первичную зрительную кору с контрастированием цветовых пятен. Дальнейшая “судьба” визуальной информации в современной литературе описана не столь четко. Большинство исследований по этой теме посвящено избирательной активности тех или иных областей коры при распознавании тех или иных геометрических форм и конкретных визуальных объектов [55, 71]. Особенно хорошо описан процесс распознавания человеческих лиц в веретенообразной извилине [80].
Само слово “распознавание”, используемое при описании этих информационных процессов, как раз и подразумевает сравнение новой информации с уже имеющейся. Человек видит новое, ранее не виденное лицо, но распознает его именно как лицо, то есть ранее знакомый объект. Происходит вышеописанный информационный синтез. Дискретизации информации уже не происходит, наоборот, распознаются и сравниваются с данными памяти не отдельные цветовые пятна, а их сочетания (например, лица). Возникает вопрос, каким образом, по какому принципу происходит объединение цветовых пятен в характерные объекты, которые распознаются и откуда в памяти берется информация, необходимая для распознавания – изначальный образ лица, например. Ответ на этот вопрос дает теория прототипов.
Теория прототипов Rosch. Коннекционизм
Распознавание самых разных объектов становится возможным благодаря наличию в памяти их прототипов, обобщенных образов, используемых для сравнения. По мнению Rosch, формирование прототипов начинается с момента начала восприятия и продолжается всю жизнь. В основе этого процесса – усвоение нейронными сетями человека определенных статистических закономерностей. В реальном мире цветовые пятна предъявляются человеку не хаотично, а образуют друг с другом характерные сочетания. Так, одним из наиболее ранних объектов реальности, который обучается распознавать ребенок, является лицо, причем конкретное лицо матери. Это становится возможным благодаря многократному предъявлению характерного сочетания цветовых пятен (глаза, нос, рот). Именно эти характерные, повторяющиеся комбинации признаков становятся прототипами [67].
В свою очередь прототипы тоже образуют между собой устойчивые сочетания, которые также “запоминаются” и становятся прототипами более высокого уровня обобщения. Так, “лицо”, как правило, предъявляется человеку вместе с руками, ногами и другими атрибутами человеческого тела. “Запоминая” эту характерную комбинацию признаков, ребенок формирует прототип “человека”, который в дальнейшем будет дополняться все новыми характерными признаками (не обязательно визуальной модальности). Собственно прототипы с появлением речи и возможности соотносить их со словами и становятся понятиями.
Такой принцип обучения нейронных сетей выделять часто встречающиеся сочетания признаков хорошо укладывается в концепцию коннекционизма. Это один из подходов к изучению когнитивных процессов, который опирается на моделирование когнитивных процессов с помощью искусственных нейронных сетей. Сам процесс обучения нейронных сетей (искусственных или человеческих) выделять характерные или часто встречающиеся комбинации признаков называется статистическим обучением. В настоящее время благодаря принципам статистического обучения смоделированы многие процессы, присущие человеческому мышлению. В частности, показана возможность их обучения выделять слова в человеческой речи [17], классифицировать изображения животных и мебели [63].
Отметим, что с позиции коннекционизма, память, в которой “хранятся” прототипы для сравнения и распознавания – не какое-то абстрактное отдельное место в коре, а свойство самих нейронных сетей, участвующих в восприятии и обучающихся выделять в большом потоке информации специфические комбинации признаков. Распознавание этих комбинаций приводит к активации нейронных сетей более высокого уровня, распознаванию характерных “комбинаций из комбинаций” нижележащего уровня. Возникает иерарахическая система, настроенная на выявление в постоянно поступающей к рецепторам информации все более глубоких закономерностей. Рассмотрим эту систему подробнее.
Информационные преобразования при образовании понятий. Объемлющие характеристики
При формировании понятий или при распознавании визуальных объектов с изначальной сенсорной информацией происходит 2 изменения:
1. Значительный объем информации о физических характеристиках объекта утрачивается. Прототип “стула” или “человеческого лица” не содержит в себе информации о конкретных параметрах того или другого. Для распознавания объекта оказывается достаточным наличия небольшого набора ключевых характеристик. Лицо может быть распознано даже на схематичном изображении. Этот очевидный факт имеет конкретное нейрофизиологическое подтверждение. Показано, что нейроны, участвующие в распознавании лиц, чувствительны к расстояниям между различными частями лица, такими как глаза, рот, брови, волосы [80]. Эти расстояния, характерные пропорции, и выступают в качестве ключевых признаков, позволяющих распознать лицо именно как лицо.
2. Утрата изначальной сенсорной информации “компенсируется” появлением у понятия семантической нагрузки. Распознавание того или иного объекта открывает доступ к огромному многообразию его свойств, о которых человек узнал в течение всей жизни. Узнавание лица или восприятие имени конкретного знакомого человека сразу же актуализирует значительный объем информации, характеризующей роль этого человека в жизни того, кто в данный конкретный момент услышал его имя или узнал его лицо.
Проиллюстрировать это уменьшение объема информации при “повышении ее качества” можно с позиции гипотезы объемлющих характеристик, предложенной Сергиным [6]. Согласно этой гипотезе первый слой нейронов, участвующих в акте восприятия, первый слой нейронов затылочной коры, ретинотопически отображает изображение, попадающее на сетчатку. Второй слой нейронов, участвующий в акте восприятия, обладает более широким рецептивным полем, чем первый и отображает уже не само изображение, а специфические паттерны активации первого слоя. Эти специфические паттерны активации и является объемлющими характеристиками. Объемлющая характеристика возникает благодаря редукции части объема информации (информации о каждом пикселе изображения) и выделении ключевых особенностей (например, область одного цвета). Последующие слои нейронов обладают все более широкими рецепторными полями. Они анализируют сочетания объемлющих характеристик нижележащего слоя, выделяя в них ключевые элементы и формируя объемлющую характеристику для более высокого нейронного слоя. Таким образом, формируется иерархия объемлющих характеристик, морфологической основой которой является иерархия нейронных слоев с все более широкими рецепторными полями. В информационном же плане формирование иерархии объемлющих характеристик как раз и позволяет совершить редукцию объема перцептивной информации за счет выделения осмысленных ее фрагментов (в конечном итоге, узнавания лиц, предметов и т.д.)
Понятие объемлющей характеристики, во многом, позволяет решить сформулированную в названии статьи проблему дискретности, континуальности и “элементарной единицы” информации, по крайней мере, применительно к процессу восприятия.
Первоначальное восприятие визуальной сцены осуществляется дискретно, коль скоро оно осуществляется дискретными элементами – клетками сетчатки, нейронами первичной зрительной коры. Однако каждый последующий уровень обработки зрительной информации подразумевает обобщение информации предыдущего уровня в виде объемлющей характеристики. Отметим здесь, что объемлющая характеристика уже не разделима ни на какие составляющие элементы. Так, узнавая человеческое лицо, мы узнаем именно характерное сочетание элементов, а не сами элементы, не простую их сумму. В этом смысле, можно сказать, что для каждого уровня восприятия элементарной единицей информации является объемлющая характеристика, которая, в свою очередь, является результатом обобщения объемлющих характеристик предыдущего уровня, но ни в коем случае не включает их в виде составных частей. Поэтому и вопрос о выделении элементарной единицы информации должен решаться применительно к каждому конкретному уровню восприятия отдельно.
Абстрактные понятия
Теория прототипов хорошо описывает формирование элементарных понятий на основании обобщения непосредственного сенсорного опыта. По мере своего развития ребенок начинает осваивать все больше абстрактных понятий, которые не имеют непосредственной связи с перцептуальным опытом и являются, своего рода, производными от системы конкретных понятий. Существует несколько теорий относительно того, как в нашем сознании формируются абстрактные понятия.
Barsalou рассматривает формирование абстрактных понятий как процесс моделирования внутренних состояний [11]. Согласно этой теории, абстрактные понятия возникают как обобщение конкретных и соотнесение их с внутренними субъективными ощущениями и переживаниями человека. Lakoff и Johnson рассматривают формирование абстрактных понятий через метафору, то есть отчуждения свойства от объекта [41]. Для описания эмоциональных состояний и иных фрагментов когнитивного опыта используется, своего рода, мышление по аналогии. Абстрактные понятия и перцептуально-недоступные процессы описываются по аналогии с конкретными, доступными непосредственному восприятию. Эта точка зрения находит определенное отражение в языке. Мы можем привести множество примеров слов, происходящих от конкретных понятий, но ассоциируемых с абстрактными. Таковы, например, слова, образованные от характеристик пространства – “повышение” (например, цен), “унижение”, “правота”, “левый” (в значении “фальшивый”, “некачественный”). Glenberg и Kaschak рассматривают формирование абстрактных понятий через развитие моторных навыков, овладение сложными алгоритмами действий [27]. В пользу этой теории говорит тот факт, что при понимании человеком слов, обозначающих абстрактные понятия, происходит активация моторной коры [28].
Общей идеей всех этих подходов является своеобразное “заземление” (в англоязычной литературе термин “grounding”) абстрактных понятий на конкретных, доступных непосредственному восприятию [62]. Мозг обобщает информацию, доступную непосредственному восприятию, выделяет ее ключевые особенности, универсальные для ряда конкретных объектов. Эта информация, в свою очередь, может стать материалом для дальнейшего обобщения с выделением еще более абстрактных и универсальных свойств.
Вновь мы сталкиваемся с уже знакомым нам на примере восприятия зрительной информации последовательным обобщением информации с уменьшением объема данных, но повышением “качества” информации за счет появления смысловой составляющей. Мы можем сказать, что процесс обобщения информации и выделения в ней ключевых характеристик не заканчивается распознаванием конкретных образов или формированием прототипов. Сами эти образы в течение жизни становятся материалом для последующего обобщения. В течение всей жизни человек копит и обобщает информацию о внешнем и внутреннем мире, формируя понятия все более высокого уровня обобщения, абстракции.
Понятие объемлющей характеристики вполне вписывается в существующие представления о формировании абстрактных понятий и может быть перенесено из области непосредственного восприятия в область дальнейшего многоуровневого обобщения изначальной перцептивной информации.
Вопрос о дискретности или континуальности информации в сфере абстрактного мышления также может быть решен с позиции объемлющих характеристик. Становится очевидным, что всякое понятие, каким бы широким оно не было по своему значению, не разложимо ни на какие составные части. В этом легко убедиться, пытаясь, например, “объяснить” понятие или посмотреть его значение в словаре. Такое “объяснение” никогда не будет исчерпывающим, никогда не передаст всю массу субъективных ощущений и ассоциаций, которые понятие вызывает у конкретного человека. В то же время эти ассоциации нельзя рассматривать как составные элементы понятия. Понятие и есть “элементарная единица информации” для своего уровня обобщения. Ассоциации и образы, в результате обобщения которых оно сформировалось, также являются элементарными единицами информации, но уже для другого уровня обобщения.
Роль гиппокампа в формировании понятий.
Общей идеей формирования простых прототипов (стол, стул) и высоко абстрактных понятий, таких как, например, “причина” и “следствие” является обнаружение в изначально-бессмысленном сенсорном потоке закономерностей, повторяющихся сочетаний признаков. Таким образом, логично предположить наличие механизма, обеспечивающего постоянное сличение различных эпизодов памяти (даже существенно разнесенных во времени) и выявление в них закономерностей. Такая роль в последнее время чаще всего отводится гиппокампу и его многочисленным связям с корой [32, 37, 38, 48, 53, 68]. Согласно представлениям ученых, гиппокамп обеспечивает быстрое запоминание отдельных эпизодов (эпизодическую память) посредством изменения функциональных связей внутри гиппокампа, а также связей гиппокампа с корой, в то время как небольшие изменения коротких кортико-кортикальных связей посредством статистического обучения обеспечивают медленное усвоение закономерностей, встречающихся в этих эпизодах, то есть, семантическую память. Таким образом, гиппокамп обеспечивает разграничение отдельных эпизодов памяти, а кора обеспечивает обнаружение в этих разрозненных эпизодах общих закономерностей. Функциональные “петли”, существующие между гиппокампом и корой, являются материальным субстратом этого процесса. Подтверждением этих теорий являются исследования пациентов с повреждением гиппокампа. Показано, что у таких пациентов нарушается способность к установлению новых семантических связей [39, 40]. Интересным фактом, выявленным при детальном изучении гиппокампа и его роли в обобщении опыта и формировании общих семантических представлений, стала его функциональная неоднородность. Показано, что дорзальные отделы гиппокампа вовлечены в формирование более детализированных конкретных представлений, а передние – более абстрактных [43, 60].
3. ИСПОЛЬЗОВАНИЕ ПОНЯТИЙ
Итак, определенным уровнем обобщения информации нашим мозгом являются понятия. Использование понятий подразумевает возможность актуализации набора ассоциаций, связанных с понятием. Какие же нейрофизиологические механизмы могут обеспечивать эту феноменологическую данность?
Для ответа на этот вопрос проводились многочисленные фМРТ-исследования активности мозга при использовании тех или иных понятий. В качестве “использования” применялось восприятие понятий на слух, их представление, чтение. Данные, полученные в ходе этих исследований, можно обобщить в нескольких пунктах:
1. Мысли о каком-либо объекте активируют те же области мозга, которые активируются при его восприятии [16, 46, 75].
2. Чем более модально специфично то или иное понятие, тем больше выражена эта закономерность [47]. Аналогичная закономерность наблюдается для понятий, ассоциированных с действиями, то есть с моторной корой. Показано, что паттерны активации, возникающие при употреблении глаголов “собирать”, “пинать”, “лизать” включают те области моторной коры, которые связаны с соответствующими частями тела (руками, ногами, губами соответственно) [61].
3. Чем более разнообразным является контекст употребления того или иного понятия в языке, чем оно более полисемантично, тем шире паттерн активации, наблюдаемый при его использовании [65, 52 ]
4. Паттерн активации, возникающий при использовании понятия, зависит от индивидуально-личностных условий его формирования. Показано, что при предъявлении картинок с изображением инструментов и назывании этих инструментов у правшей активируются области коры в левом полушарии, а у левшей – в правом. При этом области активации при предъявлении изображений животных не зависят от ведущей руки [33]. Показана аналогичная зависимость паттернов активации от ведущей руки при чтении глаголов “писать” и “бросать” [77]. У профессиональных музыкантов предъявление изображений музыкальных инструментов вызывает активацию областей слуховой ассоциативной коры [30]. Чтение предложений, описывающих действия во время игры в хоккей, вызывает активацию премоторных областей коры у профессиональных хоккеистов [13]. Показана связь паттернов активации при использовании понятий с теми модальностями, в которых тот или иной человек предпочитают получать информацию (например, визуально или на слух) [35, 36].
5. Паттерн активации, возникающий при использовании понятия, зависит от непосредственного контекста его употребления. Например, в зависимости от контекста, лимон может быть охарактеризован и как “желтый”, и как “круглый”, и как “кислый”. В фМРТ исследованиях показано, что паттерн активации при оперировании тем или иным понятием зависит от того, какие его свойства актуализирует контекст. Особенно хорошо это проявляется применительно к понятиям, чьи свойства (как у лимона) хорошо описываются в различных модальностях [31, 51, 59, 72, 66, 74, 76 ].
Общим выводом из этих экспериментальных данных является положение о том, что использование понятия приводит к активации тех нейронных сетей, которые изначально в ходе онтогенеза участвовали в его формировании [8, 11, 20]. Понятие выступает в качестве своего рода ярлыка, позволяющего быстро получить доступ к большому массиву нужной информации [44]. Причем в ходе онтогенеза по мере использования понятия в разных контекстах происходит постоянное изменение смысла понятий, постоянное переобучение нейронных сетей, репрезентирующих понятия. Это сближает и делает фактически неотделимыми понятие и контекст, в котором оно возникает и существует [10, 22, 23].
Мы можем рассматривать возникновение специфических паттернов активации при использовании тех или иных понятий как процесс обратный самому формированию понятий. При формировании понятий происходит процесс последовательного обобщения сенсорных характеристик, “сжатие” большого объема сенсорной информации в понятие, несущее смысл. При употреблении же понятия происходит как бы разворачивание в обратном порядке породивших его сенсорных характеристик, “припоминание” изначального сенсорного или сенсомоторного опыта. Этому “припоминанию” соответствует активация соответствующих областей коры, изначально связанных с восприятием, наблюдаемая в экспериментах. Понятие в сжатом виде содержит “историю” своего формирования – последовательность объемлющих характеристик, нейронный субстрат которых вновь активируется при использовании понятия.
Этот процесс на феноменологическом уровне может быть соотнесен с такими явлениями, как представление и понимание. При произнесении какого-либо слова, названия, например, “Красная площадь”, мы почти непроизвольно визуально представляем данный объект, то есть актуализируется наш прежний перцептивный опыт, послуживший материалом для формирования понятия. Отдавая себе отчет в том, что слово “понимание” само по себе многозначно и используется в современной литературе в разных смыслах, мы все же считаем, что процесс развертывания объемлющих характеристик при употреблении понятия можно назвать и пониманием. Всякое слово, даже малознакомое, при произнесении порождает набор ассоциаций, отсылающих нас к контексту, в котором мы его усвоили. Мы можем не знать, что такое синхрофазотрон, но смутно “понимать”, что он относится к физике, возможно, к физике элементарных частиц. Эти ассоциации, актуализирующие наш прежний опыт взаимодействия с понятием, и будут нашим его пониманием, пусть и на чисто поверхностном уровне. По мере получения новой информации, связанной с понятием, оно начинает порождать у нас все больше ассоциаций, контекст его употребления расширяется (расширяется и соответствующий паттерн активации нейронных сетей), а наше понимание понятия становится полнее и глубже.
4. ОБОБЩЕНИЕ И “ПОНИМАНИЕ” В НОРМЕ И ПАТОЛОГИИ. ВЫБОР ЗНАЧИМЫХ ХАРАКТЕРИСТИК
Таким образом, всякая новая информация, воспринимаемая человеком, порождает возбуждение нейронных сетей, которое можно описать двумя процессами:
1. Процесс обобщения (например, последовательное обобщение зрительной информации от визуального изображения к распознаванию конкретного образа). Этот процесс можно описать как распространение возбуждения “вверх” по сенсорным сетям с последовательным выделением объемлющих характеристик все более высокого уровня.
2. Процесс раскрытия содержания образа или понятия (понимания). Этот процесс сопровождается возникновением в коре паттернов активации, сходных с теми, которые возникают при восприятии. При этом он может запускаться не изображением, а, например, произнесением слова. То есть происходит процесс, в каком-то смысле, обратный обобщению – информация распространяется “вниз” по сенсорным путям, актуализируются ранее накопленные перцептивные данные.
И для одного, и для другого процесса важно выделение ключевых характеристик. Для распознавания лица не нужен весь массив сенсорных данных, достаточно наличия ключевых его частей и характерных пропорций. Понимание понятия также не требует актуализации всех его возможных свойств, усвоенных в ходе жизни. Правильное понимание и использование понятия в нужном контексте происходит благодаря лишь нескольким наиболее значимым его свойствам или характеристикам. Таким образом, информация, привлекаемая для понимания понятий или извлекаемая из изначальных сенсорных данных, неравнозначна по своей значимости.
Говорить о значимости информации неправильно без учета контекста. Что значимо и важно в одном контексте, может оказаться несущественно в другом. Именно поэтому паттерны активации, наблюдаемые при использовании человеком одних и тех же понятий, различаются в зависимости от контекста.
В подавляющем большинстве случаев контекстом является наша повседневная жизнь, а наиболее значимыми свойствами понятий оказываются те, которые связывают понятие с функцией объекта, который оно обозначает. То есть, например, для таких понятий как “ложка”, “чашка”, “вилка” наиболее значимыми будут ассоциации с их назначением при приеме пищи (а не с их формой, материалом, цветом). Очевидно, что именно эти связи чаще всего используются человеком и, таким образом, “закрепляются” при обучении корковых нейронных сетей. При этом в ситуации специфического контекста (например, задача выбрать предметы круглой формы) будут актуализированы другие свойства данных предметов и, соответственно, другие ассоциации обозначающих их понятий.
Отсутствие привязки понятия к текущему контексту породило бы ситуацию, в которой его употребление порождала бы актуализацию почти бесконечного объема данных, накопленных человеком в течение жизни, актуализации самых различных воспоминаний, малозначительных эпизодов, как-либо связанных с понятием. Это, в свою очередь, привело бы либо к снижению продуктивности ассоциативного процесса и, в конечном итоге, всего процесса мышления, либо к “выхватыванию” из огромного массива данных ассоциаций по случайному принципу, что при правильном с сенсорной точки зрения восприятии привело бы к искаженному пониманию целостной реальности.
Шизофрения
Именно такая ситуация имеет место при шизофрении. В исследованиях мышления пациентов, страдающих этой болезнью, показано, что ведущим нарушением мышления, во-многом определяющим всю многообразную клинику болезни, является нарушение способности пациентов разграничить существенные и малосущественные ассоциации, существенные и малосущественные признаки предметов и понятий [1, 5].
В отечественной школе патодиагностики созданы многочисленные экспериментальные методики, хорошо иллюстрирующие эту ключевую особенность больных. Так, например, одной из таких методик является сравнение понятий. Пациенту дается задание ответить на вопрос, что общего и в чем разница между двумя понятиями. При этом исследователь называет пары понятий, постепенно переходя от легко сравнимых (например, остров и полуостров; река и озеро) к менее сравнимым (например, лодка и ложка) и к условно несравнимым (например, ботинок и карандаш; еж и молоко). Задача по поиску общих признаков у карандаша и ботинка вполне может быть решена здоровым, как за счет повышения уровня обобщения (то и другое – предметы человеческого быта), так и за счет актуализации малозначительных (латентных) признаков – “то и другое оставляет след”. Однако, для здорового человека такая актуализация требует довольно значительного и целенаправленного когнитивного усилия (возможно, с перебором всех возможных признаков карандаша и ботинка). Понятия потому и являются “несравнимыми”, что автоматически вызывают весьма разные ассоциации (ботинок – обувь; карандаш – средство для письма или рисования). Иначе обстоит дело у больных шизофренией. Показано, что они значительно чаще здоровых и, главное, гораздо легче, чем здоровые актуализируют, привлекают к решению той или иной задачи латентные признаки предметов и понятий. Легкость, с которой пациенты сравнивают трудносравнимые понятия, выполняют иные экспериментально-психологические задания, давая оригинальные и формально правильные ответы, особенно диссонирует с характерной для больных несостоятельностью при решении многих повседневных задач. Причина этой несостоятельности та же, пациентам приходится постоянно сталкиваться с информационной избыточностью, вызванной их неспособностью жестко подчинить собственный ассоциативной процесс требованиям контекста ситуации.
Это феноменологическое наблюдение имеет ряд параллелей с данными инструментальных и биохимических исследований. Из наиболее ранних работ отметим исследования вызванных потенциалов (ВП) больных шизофренией [7], в ходе которых были обнаружены аномалии поздних волн ВП, физиологическая роль которых как раз состоит в оценке биологической значимости стимула. Интересные результаты получены также при сопоставлении данных о биологической роли дофамина [14, 24, 25], его дисбаланса при шизофрении и клиники болезни. Сопоставление этих данных позволило Капуру сформулировать свою теорию нарушенной салиентности [34]. Под салиентностью в англоязычной литературе понимают и саму по себе значимость (от аглийского “salience” – значимость), и способность мозга присваивать значимость той или иной информации, оценивать значимость информации в контексте. Вслед за советскими психологами, Капур, теперь уже с позиций исследования нейромедиатров, заявляет о том, что ведущим нарушением при шизофрении является неспособность больных правильно (то есть, в зависимости от контекста) оценить значимость той или иной информации, правильно оценить значимость различных ассоциаций, порождаемых тем или иным словом или событием.
Сеть салиентности
Растущий интерес к теме салиентности, оценки значимости стимулов и информации в широком смысле, заставил исследователей фМРТ говорить о функциональной сети салиентности. Анатомически к ней относят орбитальную фронтоинсулярную кору и дорзальную часть передней цингулярной коры в их тесной функциональной связи с подкорковыми лимбическими структурами [69]. В многочисленных исследованиях показана их активация в ответ на стимулы, обладающие высокой биологической значимостью [12, 15, 19, 21, 58, 70]. Большинство нервных узлов сети салиентности представляют собой подкорковые структуры, связанные с эмоциями, регуляцией гомеостаза и “центром удовольствия” [49, 54]. Сеть салиентности находится в тесном функциональном взаимодействии с центральной исполнительной сетью и базовой сетью покоя. Встречаясь со значимыми стимулами, она активирует центральную исполнительную сеть и деактивирует базовую сеть покоя, тем самым заставляя мозг выходить из состояния покоя и предпринимать те или иные действия в ответ на значимые стимулы [50]. Очевидно, что, описывая сеть салиентности, ученые говорят скорее о ее роли в выявлении и обработки стимулов высокой биологической значимости, нежели о ее роли в выделении главного и второстепенного в том или ином контексте. Имеет ли первая и вторая задачи один и тот же физиологический механизм, связанный с одними и теми же мозговыми структурами, еще предстоит выяснить. Существуют довольно многочисленные данные, указывающие на патологию сети салиентности при шизофрении [9, 26, 29, 42, 56, 57, 79]. Эти данные косвенно свидетельствуют о том, что описанная сеть салиентности имеет отношение и к выбору наиболее значимых ассоциаций.
Поиск структур мозга, активно вовлеченных в оценку значимости информации и связанный с этим процессом выбор критерия категоризации не ограничился изучением сети салиентности в норме и патологии. Интересные данные были получены благодаря ингибирующей электростимуляции различных областей. Было показано, что ингибирующая электростимуляция левой префронтальной коры приводит к затруднению классифицировать объекты по определенному признаку (например, как “круглые” или “красные”), тогда как способность к классификации на основании множества сложных свойств (например, “приспособления для хранения жидкости”) не исчезает [18, 45]. Эти данные, наряду с данными о сети салиентности, также указывают на преимущественное вовлечение лобной коры в оценку значимости информации в зависимости от контекста задачи. Причем интересным фактом является то, что более элементарный критерий категоризации (цвет или форма) оказывается более уязвим к ингибирующему воздействию, чем более сложный (применение). Очевидно, это связано с тем, что более элементарные и более модально специфичные критерии категоризации более жестко увязаны с определенным материальным субстратом, опосредующим первоначальное восприятие формы и цвета. Более сложные критерии категоризации являются продуктом длительного обучения нейронных сетей в течение жизни, подразумевают понимание смысла, роли предмета и, видимо, опосредуются более широкими нейронными сетями, не связанными с какими-либо конкретными мозговыми структурами и потому неуязвимыми к прямому ингибирующему воздействию.
4. РЕЗЮМЕ
1. Парадокс дискретности – континуальности информации при восприятии и мышлении может быть разрешен при рассмотрении разных уровней обобщения информации, организованных иерархически: от простого ретинотопического отражения физических характеристик стимула к понятиям высокого уровня абстрагирования. Каждому такому уровню присуща своя элементарная и неделимая единица информации, которая, в свою очередь, является объемлющей характеристикой (по В.Я. Сергину), результатом обобщения информации нижележащего уровня. При этом она не может быть разделена ни в функциональном плане (на уровне паттернов активации коры), ни в феноменологическом на информационные характеристики более низкого уровня (человек воспринимает лицо, визуальный объект или понятие именно как некую целостность, неразделимую на части).
2. Всякая новая информация, воспринимаемая человеком, порождает возбуждение нейронных сетей, которое можно описать двумя процессами: процессом обобщения, подразумевающим распространение возбуждения “вверх” по сенсорным сетям с последовательным выделением объемлющих характеристик все более высокого уровня, и процессом понимания. Этот процесс сопровождается возникновением в коре паттернов активации, сходных с теми, которые возникают при восприятии, и может рассматриваться как процесс, обратный обобщению – информация распространяется в функциональном смысле “вниз” по сенсорным путям. Эти процессы, по видимости, протекают практически одновременно – мы узнаем объект, понимаем его категориальную принадлежность и в тоже время “припоминаем” конкретные свойства объектов такого рода, знания которых накоплены нами в течение жизни. Так, увидев разъяренного льва, мы с одной стороны, распознаем его (обобщаем информацию), а с другой стороны припоминаем ассоциации, порождаемые у нас образом этого животного (например, то что львы опасны) – см. рис. 1. Заметим, что сам по себе конкретный лев информативен только тем, что указывает нам на свою принадлежность ко львам. Из непосредственного наблюдения мы не можем получить никакой информации, кроме этой. Прочая информация (о львах как о классе объектов) актуализируется только после соответствующего распознавания образа и его категориальной принадлежности. Но именно эта информация о львах, как о классе объектов, и важна для нашего спасения.
Рис. 1.
“Восприятие и понимание”. В процессе восприятия семантически нейтральная информация о физических характеристиках стимула обобщается, сравнивается с ранее накопленным опытом и приводит к смысловому распознаванию объекта (в данном случае – льва). Одновременно в памяти актуализируются ключевые свойства данного класса объектов (львов), в частности их потенциальная опасность, что и определяет последующее поведение индивида относительно объекта.
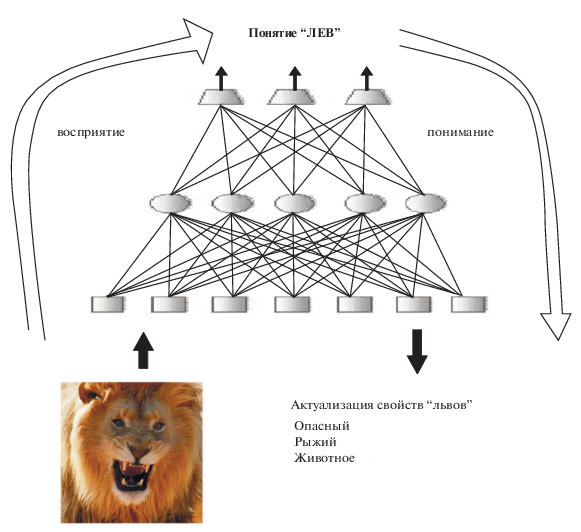
3. Эффективность обоих процессов (обобщения и понимания) обеспечивается выделением в объеме перцептивных данных или данных памяти ключевых характеристик. Именно это специфическое выделение в информационном потоке главного позволяет быстро и эффективно транслировать информацию вверх при процессе обобщения и вниз в процессе понимания, привлекая для осуществления этих процессов ограниченный набор наиболее важных ассоциаций. Это выделение главного, во многом, опосредовано дофамином. Примером нарушения этой фундаментальной особенности восприятия и мышления является шизофрения. Эффективность процесса обобщения и понимания у таких пациентов снижается за счет задействования гораздо большего объема данных. В тоже время пациенты, страдающие шизофренией, часто демонстрируют хорошую способность к решению нестандартных, творческих задач, как раз из-за того, что привлекают к решению задачи больший объем данных.
Список литературы
Зейгарник Б.В. Патопсихология. Издательство Московского университета. Москва. 1986. 234 с.
Иваницкий А.М. Мозговая основа субъективных переживаний: гипотеза информационного синтеза // Журн. высш. нервн. деят. 1996. Т. 46. № 2. С. 241.
Иваницкий А.М. “Чтение мозга”: достижение, перспективы и этические проблемы // Журн. высш. нерв. деят. 2012. Т. 62. № 2. С. 133–142.
Орлов Р.С., Ноздрачев А.Д. Нормальная физиология: Учебник., 2009. 688 с
Поляков Ю.Ф. Патология познавательной деятельности при шизофрении. М., 1974. 86 с
Сергин В.Я. Перцептивное связывание сенсорных событий: гипотеза объемлющих характеристик // Журн. высш. нерв. деят. 2002. 52(6): 645–655
Стрелец В.Б. Исследование биоэлектрической активности головного мозга больных шизофренией по показателям вызванных потенциалов // Невропатология и психиатрия. 1968. Т. 68. № 1. С. 82–89.
Allport DA. Distributed memory, modular subsystems and dysphasia. In: Newman SK, Epstein R, editors. Current Perspectives in Dysphasia. Churchill Livingstone; Edinburgh: 1985. P. 207–244
Alonso-Solís A., Vives-Gilabert Y., Grasa E., Portella M.J., Rabella M., Sauras RB, Roldán A., Núñez-Marín F., Gómez-Ansón B., Pérez V., Alvarez E., Corripio I. Resting-state functional connectivity alterations in the default network of schizophrenia patients with persistent auditory verbal hallucinations // Schizophr. Res. 2015. V. 161. № 2–3. P. 261–268.
Altmann G.T.M. The Ascent of Babel: An exploration of language, mind, and understanding. University Press; Oxford, England: 1997.
Barsalou W.L. Perceptual symbol systems // Behavioral and Brain Sciences. 1999. V. 22(04). P. 557–660.
Bartels A., Zeki S. The neural correlates of maternal and romantic love // Neuroimage. 2004. V. 21. P. 1155–1166
Beilock S.L., Lyons I.M., Mattarella-Micke A., Nusbaum H.C., Small S.L. Sports experience changes the neural processing of action language // Proc. Natl. Acad. Sci. USA. 2008. V. 105(36). P. 13269-73.)
Berridge K.C., Robinson T.E. What is the role of dopamine in reward: hedonic impact, reward learning, or incentive salience? // Brain Res. Rev. 1998. V. 28. P. 309–369.
Blood A.J., Zatorre R.J. Intensely pleasurable responses to music correlate with activity in brain regions implicated in reward and emotion // Proc. Natl. Acad. Sci. USA. 2001. V. 98. P. 11818 –11823
Chao L.L., Martin A. Cortical representations of perception, naming and knowing about color // J. Cogn. Neurosci. 1999. 11. 25–35.
Christiansen M.H., Allen. J., Seidenberg M. Learning to segment speech using multiple cues: a connectionist model // Language and Cognitive Processes. 1998. V. 12(2/3). P. 221–268.
Chrysikou E.G., Hamilton R.H., Coslett H.B., Datta A., Bikson M., Thompson-Schill S.L. Noninvasive transcranial direct current stimulation over the left prefrontal cortex facilitates cognitive flexibility in tool use // Cogn. Neurosci. 2013. V. 4(2). P. 81–9.
Craig A.D. How do you feel? Interoception: the sense of the physiological condition of the body // Nat. Rev. Neurosci. 2002. V. 3. P. 655–666.
Damasio A.R. The brain binds entities and events by multiregional activation from convergence zones // Neural Computation. 1989. V. 1. P. 123–132.
Eisenberger N.I., Lieberman M.D., Williams K.D. Does rejection hurt? An FMRI study of social exclusion // Science. 2003. V. 302. P. 290–292.
Elman J.L. Finding structure in time // Cognitive Science. 1990. V. 14. P. 179–211.
Elman J.L. Learning and development in neural networks: the importance of starting small // Cognition. 1993. V. 48(1). P. 71–99.
Fibiger H.C., Phillips A.G. Reward, motivation, cognition: psychobiology of mesotelencephalic dopamine systems. Handbook of Physiology, Section 1: The Nervous System V. 4: Intrinsic Regulatory Systems of the Brain. Edited by Bloom F.E. New York, Oxford University Press. 1986. P. 647–675.
Fibiger H.C., Phillips A.G. Mesocorticolimbic dopamine systems and reward // Ann. N.Y. Acad. Sci. 1988. V. 537. P. 206–215.
Gaebler A.J., Mathiak K., Koten J.W.Jr., König A.A., Koush Y., Weyer D., Depner C., Matentzoglu S., Edgar J.C., Willmes K., Zvyagintsev M. Auditory mismatch impairments are characterized by core neural dysfunctions in schizophrenia // Brain. 2015. V. 138. № 5. P. 1410–1423.
Glenberg M.A., Kaschak P.M. Grounding language in action // Psychonomic Bulletin & Review. 2002. V. 9(3). P. 558–565.
Glenberg M.A., Sato M., Cattaneo L., Riggio L., Palumbo D., Buccino G. Processing abstract language modulates motor system activity // The Quarterly J. Experimental Psychology. 2008. V. 61(6). P. 905–919.
Heckers S. Neuropathology of schizophrenia: cortex, thalamus, basal ganglia, and the neurotransmitter-specific projection systems // Schizophr. Bull. 1997. V. 23. № 3. P. 403–421.
Hoenig K., Müller C., Herrnberger B., Sim E.J., Spitzer M., Ehret G., Kiefer M. Neuroplasticity of semantic representations for musical instruments in professional musicians // Neuroimage. 2011. V. 56(3). P. 1714–1725.
Hoenig K., Sim E.J., Bochev V., Herrnberger B., Kiefer M. Conceptual flexibility in the human brain: dynamic recruitment of semantic maps from visual, motor, and motion-related areas // J. Cogn. Neurosci. 2008. V. 20(10). P. 1799–814.
Howard M.W., Shankar K.H., Jagadisan U.K. Constructing semantic representations from a gradually changing representation of temporal context // Top. Cogn. Sci. 2011. V. 3. P. 48–73.
Kan I.P., Kable J.W., Van Scoyoc A., Chatterjee A., Thompson-Schill S.L. Fractionating the left frontal response to tools: dissociable effects of motor experience and lexical competition // J. Cogn. Neurosci. 2006. V. 18(2). P. 267–77.
Kapur S. Psychosis as a state of aberrant salience: a framework linking biology, phenomenology, and pharmacology in schizophrenia // Am. J. Psychiatry. 2003. V. 160. № 1. P. 13–23.
Kirby J., Moore P., Schofield N. Verbal and visual learning styles // Contemporary Educational Psychology. 1988. V. 13. P. 169–184.
Kraemer D.J., Rosenberg L.M., Thompson-Schill S.L. The neural correlates of visual and verbal cognitive styles // J. Neurosci. 2009. V. 29(12). P. 3792–3798.
Kumaran D., Hassabis D., McClelland J.L. What learning systems do intelligent agents need? complementary learning systems theory updated // Trends Cogn. Sci. 2016. V. 20. P. 512–534.
Kumaran D., McClelland J.L. Generalization through the recurrent interaction of episodic memories: a model of the hippocampal system // Psychol. Rev. 2012. V. 119. P. 573–616.
Kurczek J., Brown-Schmidt S., Duff M. Hippocampal contributions to language: evidence of referential processing deficits in amnesia // J. Exp. Psychol. Gen. 2013. V. 142(4). P. 1346–54.
Kurczek J., Duff M.C. Cohesion, coherence, and declarative memory: Discourse patterns in individuals with hippocampal amnesia // Aphasiology. 2011. V. 25(6–7). P. 700–712.
Lakoff G., Johnson M. (1980). Metaphors we live by, vol. 111. London: Chicago
Lee S.K., Chun J.W., Lee J.S., Park H.J., Jung Y.C., Seok J.H., Kim J.J. Abnormal neural processing during emotional salience attribution of affective asymmetry in patients with schizophrenia // PLoS One. 2014. V. 9. № 3. P. e90792.
Long L.L., Bunce J.G., Chrobak J.J. Theta variation and spatiotemporal scaling along the septotemporal axis of the hippocampus // Front. Syst. Neurosci. 2015. V. 9. P. 37.
Lupyan G. Linguistically modulated perception and cognition: the label-feedback hypothesis // Front. Psychol. 2012. V. 3. P. 54.
Lupyan G., Mirman D., Hamilton R., Thompson-Schill S.L. Categorization is modulated by transcranial direct current stimulation over left prefrontal cortex // Cognition. 2012. V. 124(1). P. 36–49.
Martin A., Haxby J.V., Lalonde F.M., Wiggs C.L., Ungerleider L.G., Discrete cortical regions associated with knowledge of color and knowledge of action // Science. 1995. V. 379. P. 649–652.
Martin A. Functional neuroimaging of semantic memory. In Functional Neuroimaging of Semantic Memory. Edited by Cabaza R., Kingstone A., Cambridge, Massachusetts: MIT Press 2001. 153–186.
McClelland J.L., McNaughton B.L., O’Reilly R.C. Why there are complementary learning systems in the hippocampus and neocortex: insights from the successes and failures of connectionist models of learning and memory // Psychol. Rev. 1995. V. 102(3). P. 419–457.
Menon V., Levitin D.J. The rewards of music listening: response and physiological connectivity of the mesolimbic system // Neuroimage. 2005. V. 28. P. 175–184.
Menon V. Large-scale brain networks and psychopathology: a unifying triple network model // Trends Cogn. Sci. 2011. V. 15. P. 483–506.
Mummery C.J., Patterson K., Hodges J.R., Price C.J. Functional neuroanatomy of the semantic system: divisible by what? // J. Cogn. Neurosci. 1998. V. 10(6). P. 766–77.
Musz E., Thompson-Schill S.L. Semantic variability predicts neural variability of object concepts // Neuropsychologia. 2015. V. 76. P. 41–51.
Norman K.A., O’Reilly R.C. Modeling hippocampal and neocortical contributions to recognition memory: A complementary-learning-systems approach // Psychol. Rev. 2002. V. 110. P. 611–646.
Ongur D., Price J.L. The organization of networks within the orbital and medial prefrontal cortex of rats, monkeys, and humans // Cereb. Cortex. 2000. V. 10. P. 206–219.
Op de Beeck H., Vogels R. Spatial sensitivity of Macaque inferior temporal neurons // J. Comp. Neurol. 2000. V. 426. P. 505–518.
Palaniyappan L., Liddle P.F. Does the salience network play a cardinal role in psychosis? An emerging hypothesis of insular dysfunction // J. Psychiatry Neurosci. 2012. V. 37. № 1. P. 17–27
Palaniyappan L., Simmonite M., White T.P., Liddle E.B., Liddle P.F. Neural primacy of the salience processing system in schizophrenia // Neuron. 2013. V. 79. P. 814–828.
Peyron R., Laurent B., Garcia-Larrea L. Functional imaging of brain responses to pain. A review and meta-analysis // Neurophysiol. Clin. 2000. V. 30. P. 263–288.
Phillips J.A., Noppeney U., Humphreys G.W., Price C.J. Can segregation within the semantic system account for category-specific deficits? Brain. 2002. V. 125(Pt 9). P. 2067–2080.
Poppenk J., Evensmoen H.R., Moscovitch M., Nadel L. Long-axis specialization of the human hippocampus // Trends Cogn. Sci. 2013. V. 17(5). P. 230–240.
Pulvermüller F., Härle M., Hummel F. Walking or talking?: Behavioral and neurophysiological correlates of action verb processing* 1 // Brain and Language. 2001. V. 78(2). p. 143–168.
Pulvermüller F. How neurons make meaning: brain mechanisms for embodied and abstract-symbolic semantics // Trends Cogn. Sci. 2013. V. 17(9). P. 458–70.
Quinn P.S., Johnson M.H. The emergence of category representations in infants: A connectionist analysis // J. Experimental Child Psychology. 1997. V. 66. P. 236–263.
Quinn P.S., Johnson M.H. Global before basic category representations in connectionist networks and 2-month-old infants // Infancy. 2000. V. 1. P. 31–46.
Rodríguez-Ferreiro J., Gennari S.P., Davies R., Cuetos F. Neural correlates of abstract verb processing // J. Cogn. Neurosci. 2011. V. 23(1). P. 106–18.
Rogers T.T., Hocking J., Mechelli A., Patterson K., Price C. Fusiform activation to animals is driven by the process, not the stimulus // J. Cogn. Neurosci. 2005. V. 17(3). P. 434–45.
Rosch E., Mervis C.B., Gray W.D., Johnson D.M., Boyes-Braem P. Basic objects in natural categories // Cogn. Psychol. 1976. V. 8. P. 382–439.
Schapiro A.C., Turk-Browne N.B., Botvinick M.M., Norman K.A. Complementary learning systems within the hippocampus: a neural network modelling approach to reconciling episodic memory with statistical learning // Philos. Trans R. Soc. Lond. B. Biol. Sci. 2017. V. 5. P. 372(1711).
Seeley W.W., Menon V., Schatzberg A.F., Keller J., Glover G.H., Kenna H., Reiss A.L., Greicius M.D. Dissociable Intrinsic Connectivity Networks for Salience Processing and Executive Control // The J. Neuroscience. 2007. V. 27. № 9. P. 2349–2356.
Singer T., Seymour B., O’Doherty J., Kaube H., Dolan R.J., Frith C.D. Empathy for pain involves the affective but not sensory components of pain // Science. 2004. V. 303. P. 1157–1162.
Tanaka K. Inferotemporal cortex and object vision // Annu. Rev. Neurosci. 1996. V. 19. P. 109–139.
Thompson-Schill S.L., Aguirre G.K., D’Esposito M., Farah M.J. A neural basis for category and modality specificity of semantic knowledge // Neuropsychologia. 1999. V. 37(6). P. 671–6.
Treisman A. Properties, parts, and objects. In Boff, K.R., Kaufmann, L. & Thomas, J.P. (Eds), Handbook of human perception and performance (1stedn. 1986. V. 2. pp. 37.1–35.70.
Van Dam W.O., van Dijk M., Bekkering H., Rueschemeyer S.A. Flexibility in embodied lexical-semantic representations // Hum. Brain Mapp. 2012. V. 33(10). P. 2322–33.
Wiggs C.L., Weisberg J., Martin A. A Neural correlates of episodic and semantic memory retrieval // Neuropsychologia. 1999. V. 37. P. 103–118.
Willems R.M., Francken J.C. Embodied cognition: taking the next step // Front. Psychol. 2012. V. 3. P. 582.
Willems R.M., Hagoort P., Casasanto D. Body-specific representations of action verbs: neural evidence from right- and left-handers // Psychol. Sci. 2010. V. 21(1). P. 67–74.
Wolfe J.M. New York: John Wiley & Sons, Guided Search 2.0: A revised model of visual search // Psychonomic Bulletin and Review. 1999. V. 1(2). P. 202–238.
Wotruba D., Heekeren K., Michels L., Buechler R., Simon J.J., Theodoridou A., Kollias S., Rössler W., Kaiser S. Symptom dimensions are associated with reward processing in unmedicated persons at risk for psychosis // Front. Behav. Neurosci. 2014 V. 18. № 8. P. 382–389.
Yamane S., Kaji S., Kawano K., What facial features activate face neurons in the inferotemporal cortex of the monkey? // Exp. Brain Res. 1988. V. 73. P. 209–214.
Дополнительные материалы отсутствуют.
Инструменты
Успехи физиологических наук