

Успехи физиологических наук, 2022, T. 53, № 1, стр. 34-51
Динамика паттернов электрической активности мозга при дезадаптационных нарушениях
О. Е. Дик *
Институт физиологии им. И.П. Павлова РАН
199034 Санкт-Петербург, Россия
* E-mail: dickviola@gmail.com
Поступила в редакцию 20.08.2021
После доработки 29.10.2021
Принята к публикации 12.11.2021
- EDN: LXJZRA
- DOI: 10.31857/S0301179822010040
Аннотация
Обзор посвящен применению методов вейвлетного, мультифрактального и рекуррентного анализа к исследованию изменений, происходящих в паттернах электрической активности мозга человека, регистрируемой в виде электроэнцефалограммы при дезадаптационных нарушениях, связанных с тревожно-фобическими расстройствами, паническими атаками и умеренными когнитивными нарушениями. Показана возможность применения этих методов для выявления объективных показателей коррекции психогенной боли при тревожно-фобических состояниях и улучшения функционального состояния нервной системы после стимулирующего воздействия, направленного на активацию функциональных связей мозга у лиц с паническими атаками и умеренными когнитивными нарушениями.
ВВЕДЕНИЕ
В настоящее время в развитых странах отмечается значительный рост дезадаптационных нарушений, связанный с чрезмерным стрессом, оказывающим неблагоприятное воздействие на жизнь людей в мегаполисах [34, 40, 72, 73]. К таким невротическим заболеваниям, обусловленным эмоциональными перегрузками, относятся тревожно-фобические расстройства и панические атаки. При тревожно-фобических расстройствах характерными симптомами являются повышенная тревожность, нарушение сна, различные фобии и психогенная боль, означающая жалобы на абдоминальную, вертеброгенную или головную боль, не подтверждаемую медицинскими исследованиями [34]. Паническая атака представляет собой комплекс симптомов, характеризующийся пароксизмальным страхом, часто сопровождающимся чувством неминуемой гибели, или тревогой и ощущением внутреннего напряжения [40, 73, 74]. Возникновение данного патологического состояния может быть связано с нарушениями взаимоотношений симпатической и парасимпатической систем [35], а также с изменениями уровня катехоламинов [69], серотонина [77] или гаммо-аминомасляной кислоты [52].
В последнее десятилетие также наблюдается рост когнитивных расстройств, что оказывается одной из наиболее актуальных медико-социальных проблем [11, 14, 16]. Сосудистые заболевания головного мозга являются частой причиной когнитивных расстройств у лиц старшего возраста [1, 31]. При хронической цереброваскулярной недостаточности состояние когнитивных функций коррелирует с выраженностью поражения головного мозга [29]. Проявления когнитивных нарушений могут протекать от минимальной дисфункции до деменции [65]. Высокий уровень тревожности, стрессовые расстройства повышают риск развития когнитивных нарушений [30, 36, 51]. Значительная распространенность этих нарушений (более 40%) среди неврологических пациентов обуславливает необходимость поиска способов их коррекции [31].
Боль психогенной природы, панические атаки и когнитивные нарушения сопровождаются нарушением функциональных связей между корой и подкорковыми структурами на различных уровнях (таламическом, гипоталамическом или стволовом) [22]. Это должно отражаться в изменении электрической активности мозга, по сравнению с ЭЭГ здорового мозга.
Анализ изменений в характеристиках паттернов электроэнцефалограммы (ЭЭГ) мозга человека при возникновении различных патологических состояний центральной нервной системы возможен с помощью современных математических методов, позволяющих оценивать не только частотные, но и мультифрактальные, и рекуррентные показатели ЭЭГ. Например, в работах [5, 21, 38, 39, 41, 50, 56, 57, 61, 63, 64, 66] показана возможность применения методов вейвлетного и мультифрактального и рекуррентного анализа для оценки таких изменений.
Важно отметить, что при лечении невротических заболеваний, связанных с возникновением психогенной боли или панической атаки, клиницисты отмечают значительные трудности [34]. Актуальность проблемы преодоления этих трудностей связана с недостаточной эффективностью и трудностью выбора главного направления терапии: одни исследователи отдают предпочтение фармакотерапии [62], а другие ведущую роль отводят психофизиологическим методам лечения [59]. Поэтому наряду с разработкой новых безопасных лекарственных препаратов в настоящее время актуальной является разработка эффективных и безопасных немедикаментозных методов терапии болевых симптомов и панических атак. Один из таких методов для снятия психогенной боли разработан в Научно-практическом центре реабилитации им. Г.А. Альбрехта [12, 53]. Другой метод для лечения фобических неврозов разработан в Институте мозга человека им. Н.П. Бехтеревой РАН [27].
Первый метод основан на сосредоточении пациента с закрытыми глазами на месте локализации боли. Считается, что болевые ощущения по ассоциативным системам конвергируются в цветовое отражение, возникающее в головном мозге при закрытых глазах [26, 53]. Другими словами, цветовые образы отражают болевую симптоматику и наличие психоэмоционального напряжения [13]. При этом психотерапевтическое воздействие, снижающее это напряжение, сопровождается достоверным увеличением цветов коротковолновой части спектра [26]. Таким образом, согласно этой методике, пациенты оценивают изменения интенсивности боли и отслеживают сопутствующую этим изменениям трансформацию цветовых пятен от цветов длинноволновой части спектра к коротковолновой на протяжении сеанса психорелаксации. Показано, что у некоторых пациентов снятие боли происходит к двадцатой минуте третьего сеанса, а другим требуется значительно большее количество сеансов [53].
Для оценки эффективности психотерапевтического воздействия для снятия болевого синдрома у пациентов с тревожно-фобическими расстройствами в работах [4, 41, 42] применены методы вейвлетного и мультифрактального анализов фрагментов ЭЭГ во время ощущения пациентами психогенной боли и во время ее снятия с помощью психорелаксационной методики.
Разработанный в Институте мозга человека им. Н.П. Бехтеревой РАН метод для лечения панических атак связан с активацией артифициальных (искусственных) стабильных функциональных связей мозга человека [17]. В основе метода лежит внутримозговой феномен долговременной памяти, представляющий особый вид функциональных связей мозга, формирующихся в условиях активации подкорковых структур мозга [28]. Такие направленные воздействия играют важную роль для коррекции психофизиологической деятельности человека при различных заболеваниях органического и функционального генеза, а также для здоровых лиц в условиях эмоциональных перегрузок [18, 19]. Способность восстанавливать утраченные резервы психических процессов позволяет использовать этот метод в ситуациях, когда фармакотерапия имеет значительные побочные действия на организм [18–20].
Известно, что восстановление функционального состояния мозга происходит у больных с отчетливой реакцией усвоения ритмической фотостимуляции в исходной электроэнцефалограмме (ЭЭГ) [23]. При этом выраженность перестроек электрической активности мозга в соответствии с частотой световых импульсов, определяется возбудимостью нейронов коры головного мозга [32, 48]. В связи с этим для объективного подтверждения положительной динамики функционального состояния в процессе лечения методом активации функциональных связей можно использовать анализ реактивных паттернов ЭЭГ как ответа мозга на разные частоты световых импульсов до и после курса лечения. Такой анализ к оценке изменения степени невротизации у пациентов с неврологическими нарушениями в виде панических атак после лечебных воздействий, направленных на формирование функциональных связей мозга во время ритмической фотостимуляции проведен в работах [7, 46, 71]. Для оценки эффективности лечения панических атак в этих работах применены методы вейвлетного анализа и метода анализа совместных рекуррентностей ЭЭГ и светового сигнала.
В работах [8, 47, 71] рассмотрена динамика паттернов ЭЭГ мозга человека при умеренных когнитивных нарушениях до и после сеансов активации функциональных связей мозга и определены динамические характеристики, которые могут служить объективными показателями улучшения функционального состояния нервной системы после этих сеансов. Для этого использован метод анализа совместных рекуррентностей ритмических световых сигналов и ответов мозга на эти сигналы и дополнительный показатель, такой как коэффициент взаимной корреляции между вероятностями рекуррентностей светового сигнала и ответа мозга на определенные частоты.
ИСПОЛЬЗУЕМЫЕ МЕТОДЫ ИССЛЕДОВАНИЯ
Вейвлетный анализ паттернов ЭЭГ
Вейвлетный анализ паттернов ЭЭГ означает построение и анализ локальных и глобальных вейвлетных спектров и широко применяется при изучении электроэнцефалограмм мозга [10, 37–39, 50, 68].
Под локальным вейвлетным спектром понимается результат вейвлет-преобразования сигнала x(t):
Под глобальным вейвлетным спектром понимается интеграл локального вейвлетного спектра по времени [t1, t2]
т.е. интегральное распределение энергии вейвлетного спектра сигнала по частотам [50].Таким образом, в отличие от быстрого преобразования Фурье, вейвлетный анализ позволяет на основании локального вейвлетного спектра или его проекции, по осям которой отложены время и частота, определять частотно-временную структуру сигнала, т.е. получать информацию об изменении частотных характеристик сигнала во времени, а на основании глобальных вейвлетных спектров выявлять различия в значениях энергии паттернов в разные моменты времени.
Вейвлетный анализ паттернов ЭЭГ при ритмической фотостимуляции
При анализе реактивных паттернов ЭЭГ в ответ на ритмическую фотостимуляцию вейвлетный анализ имеет свои особенности, подробно изложенные в работах [6, 44, 45].
Ритмический фотостимул аппроксимируется в этих работах последовательностью k гауссовых импульсов, следующих друг за другом с заданной частотой fC:
Вейвлетное преобразование такого сигнала p(t) с использованием базового вейвлета Морле определяется функцией:
Реакцию усвоения частоты фотостимуляции в паттернах ЭЭГ можно оценить по величине коэффициента усвоения (kR) в исследуемом частотном диапазоне [fС – Δf, fС + Δf], где Δf = 0.5 Гц, а также по нарастанию энергии вейвлетного спектра непосредственно во время фотостимуляции [44]. Коэффициент усвоения ритма (kR) определяется как отношение максимумов глобальных вейвлетных спектров во время и до действия фотостимула [4]:
Значение коэффициента kR < 1 означает меньшее значение максимума глобального вейвлетного спектра во время фотостимуляции, по сравнению с максимумом спектра до фотостимуляции, т.е. отсутствие нарастания энергии вейвлетного спектра при действии определенного светового сигнала и, соответственно, отсутствие усвоения заданной частоты [39].
Мультифрактальный анализ паттернов ЭЭГ
Метод мультифрактального анализа основан на поиске локальных максимумов вейвлетного спектра и определения скорости уменьшения вейвлет-коэффициентов при уменьшении частоты f [33, 60].
Для каждого значения f находится множество L(f) линий локальных максимумов модулей вейвлет-коэффициентов, т.е. линий, для которых выполняется следующее условие:
Затем вдоль каждой линии вычисляются частичные функции Z(q, f) как сумма q степеней максимумов модулей вейвлет-коэффициентов вдоль каждой линии, так называемые обобщенные статистические суммы:
Далее вычисляется скейлинговая экспонента τ(q):
Если зависимость τ(q) будет линейной, то значение экспоненты Гельдера $h(q) = {\text{const}}$. В этом случае анализируемый сигнал будет монофрактальным. Если же зависимость τ(q) нелинейна, то $h(q) = {{d\tau (q)} \mathord{\left/ {\vphantom {{d\tau (q)} {dq}}} \right. \kern-0em} {dq}} \ne {\text{const}}$. В этом случае сигнал оказывается мультифрактальным с большим числом экспонент Гельдера [15].
Распределение экспонент Гельдера называется спектром сингулярности:
При анализе спектра сингулярности важными оказываются ширина этого спектра ∆h, характеризующая степень мультифрактальности сигнала, и положение спектра сингулярности D(h) на оси значений h, т.к. это положение дает информацию о степени коррелированности последовательных значений сигнала. Величины h < 0.5 соответствуют антикоррелированной динамике, а величины h > 0.5 – коррелированной динамике [15]. Коррелированность последовательных значений сигнала означает, что с большей вероятностью за большим значением сигнала следует большее, и наоборот. В случае, если динамика является одновременно и коррелированной, и антикоррелированной, спектр сингулярности находится в интервале значений 0 < h < 1 [75, 76].
Метод анализа совместных рекуррентностей ЭЭГ и светового сигнала
Метод анализа совместных рекуррентностей ЭЭГ и светового сигнала основан на оценке совместных рекуррентных диаграмм паттерна ЭЭГ и светового сигнала и использован в работах [2, 5, 6, 47, 48].
Совместная рекуррентная диаграмма представляет собой графическое представление матрицы
Совместная рекуррентность с точностью до ε-ошибки определяется как возврат состояния yj фазовой траектории сигнала ЭЭГ к состоянию yi и одновременный возврат состояния zj фазовой траектории светового сигнала к состоянию zi [58].
Фазовые траектории состояний z(t) и y(t) получаются из исходных сигналов x(t) и p(t) методом временных задержек [70]:
Оптимальная временная задержка d находится, как правило, на основании поиска первого минимума функции взаимной информации [49], а оптимальная размерность вложения m определяется методом поиска минимума ближайших ложных соседей [54]. Величину ε-ошибки обычно выбирают равной 1% от величины стандартного отклонения анализируемого фрагмента ЭЭГ [57].
Реакцию усвоения ритма в паттернах ЭЭГ методом анализа совместных рекуррентностей можно оценить по следующим показателям совместных рекуррентных диаграмм [58]:
1) общее число рекуррентных точек RR в рекуррентной диаграмме:
2) отношение числа рекуррентных точек, составляющих диагональные структуры, к общему числу рекуррентных точек (так называемый показатель детерминизма):
3) коэффициент взаимной корреляции между вероятностями рекуррентностей ${{P}_{1}}(\tau )$ и ${{P}_{2}}(\tau )$ светового сигнала и ответа мозга на определенные частоты [67]:
Длинные диагонали выявляют аналогичную эволюцию во времени в двух колебательных процессах, а короткие диагонали показывают уменьшение сходства динамики процессов [58]. Поэтому рекуррентная диаграмма для двух периодических сигналов содержит длинные диагонали, находящиеся друг от друга на расстоянии, равном периоду колебаний, и величина показателя детерминизма, определяющего число рекуррентных точек, лежащих на диагональных линиях, DET = 1. Для двух полностью стохастических сигналов совместная рекуррентная диаграмма не содержит длинных диагоналей, а имеет практически только изолированные точки, при этом величина показателя детерминизма DET близка к нулю. Таким образом, значение показателя DET, определяющего процент рекуррентных точек, расположенных на диагональных линиях совместной рекуррентной диаграммы, связано со сходством динамического поведения двух колебательных процессов.
Совпадение максимумов вероятностей рекуррентностей ${{P}_{1}}(\tau )$ и ${{P}_{2}}(\tau )$ для двух фазовых траекторий z(t) и y(t) при одинаковых временах kt, где k – целое число, означает наличие фазовой синхронизации двух исходных сигналов x(t) и p(t) [55]. В этом случае значение коэффициента взаимной корреляции CPR близко к 1. Напротив, если два сигнала не находятся в фазовой синхронизации, максимумы вероятностей рекуррентностей возникают неодновременно, и значение CPR близко к 0. Так, для полностью синхронизованных сигналов значение CPR = 1, а для полностью стохастических сигналов значение CPR близко к нулю.
АНАЛИЗ ДИНАМИКИ ПАТТЕРНОВ ЭЭГ ПРИ ТРЕВОЖНО-ФОБИЧЕСКИХ РАССТРОЙСТВАХ
Вейвлетные спектры паттернов ЭЭГ при тревожно-фобических расстройствах
В работах [4, 42, 43] показано, что в состоянии концентрации внимания на болевом ощущении ЭЭГ некоторых пациентов содержит альфа и тета ритм, ЭЭГ других – только тета ритм. В связи с этим пациенты с тревожно-фобическими расстройствами распределены в этих работах по двум группам. Во время психорелаксации уровень тета-активности значительно уменьшался у 45% пациентов первой группы, а у 55% этой группы тета-активность сохранялась. При этом улучшение психологического состояния не всегда коррелировало с уменьшением тета-активности. Полное устранение болевого ощущения наблюдалось только у тех лиц, у которых цветовые пятна трансформировались в голубой цвет (61% пациентов первой группы). У 39% больных этой группы во время концентрации на болевом ощущении наблюдались преимущественно цвета длинноволновой части спектра (красный, желтый), и эти цвета сохранялись во время психорелаксации [4, 42]. Для 19% пациентов второй группы при психорелаксации ахроматический черный цвет изменялся на длинноволновой красный, у 81% пациентов трансформация цветовых пятен не наблюдалась. Оба случая не сопровождались снятием болевого ощущения [4, 42]. На основании вейвлетного анализа были оценены локальные и глобальные вейвлетные спектры фрагментов ЭЭГ до, во время и после психотерапевтического воздействия.
Рассмотрим примеры этих локальных и глобальных вейвлетных спектров ЭЭГ при тревожно-фобических расстройствах (рис. 1). При ощущении психогенной боли пациентом из первой группы с тревожно-фобическими расстройствами в глобальном вейвлетом спектре наблюдается два максимума на частотах альфа- и тета-диапазонов (Emax = 5 × 105 и Emax = 5.1 × 105) (рис. 1а, 1д).
Рис. 1.
Примеры локальных и глобальных вейвлетных спектров ЭЭГ человека с тревожно-фобическими расстройствами из первой и второй группы.

Во время психорелаксации значения максимумов глобальной энергии уменьшаются до величины Emax = 4.8 × 104 на частоте альфа-диапазона и до Emax = 1.9 × 104 на частоте тета-диапазона (рис. 1б, 1е). Для пациента с тревожно-фобическими расстройствами из второй группы единственный максимум на частотах тета-диапазона Emax = 10 × 104 (рис. 1в, 1ж) уменьшается во время психорелаксации до величины Emax = 2.48 × 104 (рис. 1г, 1з).
Отметим, что у всех пациентов первой и второй группы во время психорелаксации энергия вейвлетного спектра уменьшалась приблизительно в десять раз, однако не у всех пациентов во время сеансов психорелаксации происходило устранение психогенной боли. Это означает, что использование параметра Emax вейвлетного спектра не позволяет надежно оценить эффективность психорелаксационной методики по устранению психогенной боли у пациентов с тревожно-фобическими расстройствами.
Мультифрактальные характеристики паттернов ЭЭГ при тревожно-фобических расстройствах
В работах [4, 42] продемонстрирована возможность оценки эффективности психорелаксационной методики по устранению психогенной боли у пациентов с тревожно-фобическими расстройствами на основании анализа мультифрактальных характеристик фрагментов ЭЭГ до и во время сеансов психорелаксации.
На рис. 2 представлены усредненные спектры сингулярностей во время ощущения психогенной боли и во время психорелаксации у пациентов с тревожно-фобическими расстройствами из первой и второй группы.
Рис. 2.
Примеры спектров сингулярностей у пациентов с тревожно-фобическими расстройствами во время ощущения психогенной боли (кривые отмечены красным) и во время релаксации (голубым).

Ширина спектра сингулярностей для первой группы меньше, чем для второй как во время ощущения психогенной боли, так и во время психорелаксации (рис. 2а, 2б, 2в). Во время сеанса психорелаксации значение ширины спектра сингулярности для первой группы увеличивается (рис. 2а, 2б), однако в обоих случаях максимальная величина экспоненты Гельдера hmax < 0.5, что означает, что спектры сингулярности для первой группы соответствуют антикоррелированной динамике последовательных значений ЭЭГ. При этом для 61% пациентов первой группы ширина спектра сингулярности значимо возрастает во время психорелаксации, а у 39% лиц изменяется незначительно (рис. 2а, 2б соответственно). Это совпадает с полным устранением болевого ощущения у 61% пациентов исследуемой группы. Таким образом, устранение болевого синдрома кореллирует с уменьшением степени антикоррелированности последовательных значений ЭЭГ.
Для второй группы степень мультифрактальности последовательных значений ЭЭГ значительно превышает эту величину для первой группы (hmax = = 0.98 ± 0.09, ∆h = 0.95 ± 0.08, во время сеанса психорелаксации (рис. 2в). Спектры сингулярностей соответствуют как коррелированной (h > 0.5), так и антикоррелированной (h < 0.5) динамике. Это коррелирует с отсутствием устранения болевого ощущения у больных из второй группы.
Обобщенная информация о средних значениях ширины спектра сингулярности (∆h) и максимума глобального вейвлетного спектра (Emax) для двух групп исследуемых пациентов, а также данные об устранении или отсутствии устранения болевого ощущения представлена в табл. 1.
Таблица1.
Сравнение средних значений ∆h и Emax для двух групп с тревожно-фобическими расстройствами и данных об устранении или отсутствии устранения болевого ощущения во время психорелаксации
| Физиологическое состояние | Максимум глобального спектра, Emax × 104 | Ширина спектра сингулярности, ∆h | Устранение болевого ощущения |
|---|---|---|---|
| Первая группа: | |||
| Во время боли | 61.5 ± 5.8 | 0.28 ± 0.03 | |
| Во время психорелаксации | 7.7 ± 0.8 | 0.35 ± 0.03 | нет (39% ) |
| Во время психорелаксации | 5.1 ± 0.5 | 0.52 ± 0.06 | да (61%) |
| Вторая группа: | |||
| Во время боли | 4.5 ± 0.3 | 1.22 ± 0.11 | |
| Во время психорелаксации | 0.62 ± 0.05 | 1.03 ± 0.10 | нет (100%) |
Данные табл. 1 показывают, что в первой группе устранение болевого ощущения при психорелаксации наблюдается у 61% пациентов группы и коррелирует с возрастанием ширины спектра сингулярности до значений, близких к 0.5. В ЭЭГ 39% пациентов этой группы ширина спектра сингулярности возрастает незначительно во время психорелаксации, и болевое ощущение сохраняется. Для второй группы характерно отсутствие устранения болевого ощущения при психорелаксации у 100% лиц, сохранение коррелированной динамики (h > 0.5) последовательных значений ЭЭГ и высокая степень мультифрактальности. Следовательно, устранение болевых симптомов во время психорелаксации у лиц с тревожно-фобическими расстройствами характерно только для пациентов первой группы.
Таким образом, механизмы, лежащие в основе коррекции психогенной боли при тревожно-фобических состояниях, связаны с изменением динамики последовательных значений ЭЭГ и степенью их мультифрактальности. Рассмотренный подход позволяет количественно оценить эффективность психорелаксационной методики, применяемой для устранения психогенной боли у конкретных пациентов. Отметим, что уменьшение степени антикоррелированности последовательных значений ЭЭГ означает уменьшение случайности “up-down patterns”, и, соответственно, уменьшение неустойчивости нейродинамических процессов мозга, что, в свою очередь, соответствует оптимизации корково-подкорковых отношений у 61% тестируемых пациентов первой группы.
АНАЛИЗ ДИНАМИКИ ПАТТЕРНОВ ЭЭГ ПРИ ПАНИЧЕСКИХ АТАКАХ
Вейвлетные спектры паттернов ЭЭГ при панических атаках
В работах [7, 46] показано, что в фоновой ЭЭГ 69% больных с паническими атаками до сеансов активации функциональных связей мозга преобладает высокоамплитудная активность тета-диапазона, а в ЭЭГ 31% больных наблюдается низкоамплитудная полиморфная активность в дельта-, тета- и альфа-диапазонах. Сеансы активации функциональных связей мозга приводят к значительному уменьшению амплитуды тета-активности или к исчезновению полиморфной активности и усилению активности в альфа-диапазоне [46].
Реактивные паттерны ЭЭГ во время фотостимуляции у большинства пациентов характеризуются асимметрией ответов затылочных долей мозга на фотостимул. Это проявляется в различных значениях максимумов локальных вейвлетных спектров паттернов ЭЭГ, регистрируемых в О1 и О2 затылочных отведениях. Примеры локальных вейвлетных спектров паттернов ЭЭГ в этих отведениях представлены на рис 3. Максимум локального вейвлетного спектра паттерна ЭЭГ в отведении О2 и в данном примере в 1.5 раза превышает максимум спектра в отведении О1.
Рис. 3.
Уменьшение ассимметрии вейвлетных спектров паттернов ЭЭГ после сеансов активации функциональных связей мозга. Локальные вейвлетные спектры паттернов ЭЭГ: до этих сеансов в отведениях О1 (а) и О2 (в) и после них в отведениях О1 (б) и О2 (г).

Глобальные вейвлетные спектры паттернов ЭЭГ, построенные в работах [7, 46], в узких частотных диапазонах вблизи частоты фотостимуляции, до начала действия фотостимула и во время фотостимуляции, показали значительно большее увеличение ответа мозга на фотостимул частоты 20 Гц до проводимых сеансов активации функциональных связей мозга и очень слабое повышение после проводимых сеансов.
В табл. 2 приведены средние значения коэффициентов усвоения (kR) для реактивных паттернов ЭЭГ до и после сеансов активации функциональных связей мозга.
Таблица 2.
Усредненные значения коэффициентов усвоения (kR) ритма заданной частоты
| До активации функциональных связей мозга | После активации функциональных связей мозга | |||||
|---|---|---|---|---|---|---|
| f (Гц) | О1 | О2 | p | О1 | О2 | p |
| Коэффициент усвоения (kR) ритма заданной частоты | ||||||
| 6 | <1 | <1 | 0.094 | <1 | <1 | 0.085 |
| 8 | <1 | 1.1 ± 0.1 | 0.087 | <1 | <1 | 0.075 |
| 12 | 1.9 ± 0.2 | 2.7 ± 0.2 | 0.078 | <1 | <1 | 0.068 |
| 14 | 5.4 ± 0.5 | 122 ± 18 | 0.039 | 2.1 ± 02 | 3.5 ± 0.3 | 0.081 |
| 18 | 35 ± 3.7 | 147 ± 15 | 0.045 | 11 ± 1.2 | 17 ± 1.8 | 0.074 |
| 20 | 22 ± 1.9 | 101 ± 11 | 0.038 | 5.5 ± 0.5 | 7.1 ± 0.7 | 0.081 |
До сеансов активации функциональных связей мозга для пациентов с паническими атаками значение коэффициента усвоения ритма kR < 1 для частоты тета-диапазона, что означает отсутствие нарастания энергии паттерна ЭЭГ во время фотостимуляции и отсутствие усвоения ритма данного диапазона. Незначительное усвоение выявляется для частот альфа-диапазона (kR = 1.9 ± 0.2 и kR = 2.7 ± 0.2 для 12 Гц и kR = 1.1 ± 0.1 для 8 Гц). Значительное усвоение определяется для частот бета-диапазона, например, kR = 22 ± 1.9 и kR = 101 ± 11 для 20 Гц. При этом отмечается практически пятикратное превышение значений коэффициента усвоения ритма kR для отведения О2, по сравнению со значениями для отведения О2. Таким образом, данные табл. 2 демонстрируют статистически значимые различия в средних значениях коэффициента kR, вычисленных для затылочных отведений О1 и О2 (p < 0.05), что свидетельствует об асимметрии реакции усвоения ритма бета-диапазона у большинства тестируемых пациентов.
После сеансов активации функциональных связей мозга асимметрия ответов затылочных долей мозга становится статистически незначимой (p > 0.05), а значения коэффициента усвоения ритма частот альфа-диапазона kR < 1, что означает отсутствие усвоения ритма альфа-диапазона. Усвоение ритма бета-диапазона значительно уменьшается (kR = 11 ± 1.2 для отведения О1 и kR = = 17 ± 1.8 для отведения О2 для частоты 18 Гц).
В работах [7, 46] динамика изменения усвоения ритма в паттернах ЭЭГ у пациентов с паническими атаками после сеансов активации функциональных связей мозга подтверждена также изменением одновременных рекуррентностей в совместных рекуррентных диаграммах этих паттернов и ритмических световых сигналов определенных частот.
Анализ совместных рекуррентностей паттернов ЭЭГ и ритмических световых сигналов при панических атаках
Примеры совместных рекуррентных диаграмм фрагментов ЭЭГ и ритмического светового сигнала частоты 20 Гц до и после сеансов активации функциональных связей мозга представлены на рис. 4б и 4г соответственно. Соответствующие паттерны ЭЭГ во время фотостимуляции данной частотой показаны на рис. 4а и 4в синей линией, а фотостимул – красной линией.
Рис. 4.
Примеры паттернов ЭЭГ во время фотостимуляции частотой 20 Гц до (а) и после активации функциональных связей мозга (в) (отведение О2). ЭЭГ – синяя кривая, фотостимул – красная кривая. б, г – совместные рекуррентные диаграммы паттернов и световых сигналов.

Левая диаграмма (рис. 4б) имеет рекуррентные структуры, cодержащие длинные диагональные линии, что свидетельствует о возникновении одновременных рекуррентностей в паттерне ЭЭГ и световом сигнале заданной частоты. Во время нарастания амплитуды ответа мозга на фотостимуляцию предложенной частоты (интервал значений nL от 600 до 1800) число одновременных рекуррентностей возрастает, что отражается в увеличении длины диагональных линий в рекуррентной диаграмме. В отличие от этого, правая диаграмма (рис. 4г) содержит только короткие диагональные линии, что свидетельствует о слабой совместной рекуррентности в данном световом сигнале и анализируемом паттерне ЭЭГ.
В табл. 3 приведены усредненные значения изменения общего числа рекуррентных точек (RR) в рекуррентной диаграмме для реактивных паттернов ЭЭГ до и после сеансов активации функциональных связей мозга. Данные табл. 3 свидетельствуют об уменьшении числа рекуррентных точек RR после сеансов активации функциональных связей мозга для частот бета-диапазона. Для частоты 18 Гц значение RR = 0.13 ± 0.01 (для отведения О1) и RR = 0.15 ± 0.01 (для отведения О2) до этих сеансов и RR = 0.03 ± 0.003 и RR = 0.02 ± 0.002 после них соответственно. Для частот альфа- и тета-диапазонов характерно отсутствие статистически значимых изменений динамики показателей рекуррентных диаграмм во время действия фотостимула до и после сеансов активации функциональных связей мозга.
Таблица 3.
Усредненные значения общего числа реккурентных точек (RR) в совместных рекуррентных диаграммах паттернов ЭЭГ и световых сигналов
| До активации функциональных связей мозга | После активации функциональных связей мозга | |||||
|---|---|---|---|---|---|---|
| f (Гц) | О1 | О2 | p | О1 | О2 | p |
| 6 | 0.05 ± 0.005 | 0.04 ± 0.004 | 0.095 | 0.06 ± 0.006 | 0.03 ± 0.003 | 0.086 |
| 8 | 0.03 ± 0.003 | 0.05 ± 0.005 | 0.089 | 0.02 ± 0.002 | 0.04 ± 0.004 | 0.085 |
| 12 | 0.08 ± 0.008 | 0.07 ± 0.007 | 0.090 | 0.05 ± 0.005 | 0.06 ± 0.006 | 0.094 |
| 14 | 0.11 ± 0.01 | 0.13 ± 0.01 | 0.078 | 0.02 ± 0.002 | 0.03 ± 0.003 | 0.096 |
| 18 | 0.13 ± 0.01 | 0.15 ± 0.01 | 0.075 | 0.03 ± 0.003 | 0.02 ± 0.002 | 0.096 |
| 20 | 0.12 ± 0.01 | 0.17 ± 0.01 | 0.037 | 0.04 ± 0.003 | 0.02 ±0.002 | 0.089 |
Таким образом, и метод вейвлетного анализа, и метод анализа совместных реккурентностей указывают на то, что для пациентов с паническими атаками отмечается значительная реакция усвоения ритма частот именно бета-диапазона и отсутствие реакции усвоения ритма частот тета- и альфа-диапазона или незначительная реакция усвоения некоторых частот альфа-диапазона. Активация артифициальных стабильных функциональных связей мозга приводит к уменьшению значений коэффициента усвоения и уменьшению общего числа рекуррентных точек для частот возбуждения бета-диапазона. Другими словами, сеансы активации функциональных связей мозга вызывают значительное снижение усвоения ритма бета-дипазона.
АНАЛИЗ ДИНАМИКИ ПАТТЕРНОВ ЭЭГ ПРИ УМЕРЕННЫХ КОГНИТИВНЫХ НАРУШЕНИЯХ
Вейвлетные спектры паттернов ЭЭГ при умеренных когнитивных нарушениях
Примеры вейвлетных спектров реактивных паттернов ЭЭГ в ответ на фотостимуляцию частотой 10 Гц, полученных до и после сеансов активации функциональных связей мозга, представлены на рис. 5. Для паттерна ЭЭГ, зарегистрированного до этих сеансов, характерно следующее. Локальный вейвлетный спектр демонстрирует наличие максимумов на частотах анализируемого диапазона 10 ± 0.5 Гц в интервале времени, предшествующем времени фотостимуляции (рис. 5б). Нормированные интегральные распределения энергий вейвлетных спектров ${{E(t)} \mathord{\left/ {\vphantom {{E(t)} {{{E}_{{\max }}}(t)}}} \right. \kern-0em} {{{E}_{{\max }}}(t)}}$ паттерна ЭЭГ и светового сигнала (сплошная и штрихпунктирная линии соответственно) показывают отсутствие нарастания энергии вейвлетного спектра ЭЭГ во время фотостимуляции (рис. 5в). Глобальный вейвлетный спектр E(f) этого паттерна во время фотостимуляции также имеет значения меньшие, чем до фотостимуляции (рис. 5а). Соответственно, значение коэффициента усвоения ритма предложенной частоты kR < 1. В отличие от этого, после сеансов активации функциональных связей мозга нарастание энергии вейвлетного спектра ЭЭГ происходит непосредственно в период действия фотостимула (рис. 5е), поэтому максимумы энергии локального вейвлетного спектра находятся в интервале времени, соответствующем времени действия стимула (рис. 5д). Отношение максимумов глобальных вейвлетных спектров во время и до действия фотостимула определяет значение коэффициента усвоения ритма kR > 1 (рис. 5г). Таким образом, в анализируемых паттернах ЭЭГ, соответствующих состоянию до активации функциональных связей мозга, усвоение частоты 10 Гц отсутствует, а после сеансов активации наблюдается реакция усвоения этой частоты.
Рис. 5.
Oтсутствие реакции усвоения частоты 10 Гц в паттерне ЭЭГ (отведение О1) до сеансов активации функциональных связей мозга (а–в) и возникновение реакции усвоения после сеансов (г–е). Глобальные вейвлетные спектры (а, г) до фотостимуляции (сплошная линия), во время фотостимуляции (штрихпунктирная линия). Нормированные интегральные распределения вейвлетных спектров ЭЭГ и световых стимулов (сплошные и штрихпунктирные линии (в, е). Начало и конец фотостимуляции показаны вертикальными стрелками

Для реактивных паттернов ЭЭГ в ответ на фотостимуляцию частотой 6 Гц, полученных до и после лечебных сеансов, характерно наличие реакции усвоения (рис. 6). Об этом свидетельствует превышение максимумов глобальных вейвлетных спектров во время действия фотостимула, по сравнению с величинами этих максимумов до фотостимуляции, kR > 1 в обоих случаях (рис. 6а, 6г), а также нарастание энергии вейвлетных спектров ЭЭГ именно во время фотостимуляции (рис. 6б–6е).
Рис. 6.
Наличие реакции усвоения частоты 6 Гц в паттерне ЭЭГ (отведение О2) до сеансов активации функциональных связей мозга (а–в) и после сеансов (г–е).
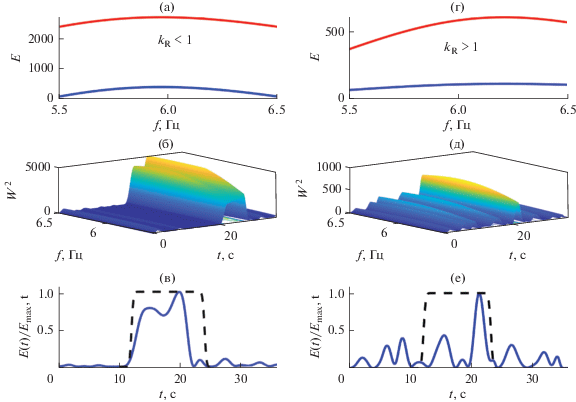
Динамика изменения усвоения ритма в паттернах ЭЭГ у пациентов с умеренными когнитивными нарушениями после проводимых сеансов подтверждена в работах [8, 46, 47, 71] также изменением одновременных рекуррентностей в совместных рекуррентных диаграммах ЭЭГ и ритмических световых сигналов определенных частот.
Анализ совместных рекуррентностей паттернов ЭЭГ и ритмических световых сигналов при умеренных когнитивных нарушениях
На рис. 7а, 7г представлены фотостимул частотой 10 Гц (красной линией) и фрагменты паттерна ЭЭГ длительностью 3 с во время фотостимуляции (синей линией). Совместные рекуррентные диаграммы для светового сигнала и фрагментов ответов мозга до и после сеансов активации функциональных связей приведены на рис. 7б, 7д соответственно. Совместная рекуррентная диаграмма, полученная до сеансов (рис. 7б) содержит очень короткие диагональные линии, локализующиеся в окрестности всегда существующей линии, проходящей под углом в 45°. Это соответствует близкому к нулю значению показателя DET и свидетельствует об отсутствии сходства динамики данного светового сигнала и анализируемого ответа.
Рис. 7.
Фрагменты ЭЭГ длительностью 3 с во время фотостимуляции (синяя линия) и световой стимул частотой 10 Гц (красная линия) до и после сеансов активации функциональных связей мозга (а, г). Совместные рекуррентные диаграммы (б, д). Вероятности ${{P}_{1}}(\tau )$ и ${{P}_{2}}(\tau )$ рекуррентностей для ЭЭГ (синяя кривая) и светового сигнала (красная кривая) (в, е).

В отличие от этого, совместная рекуррентная диаграмма после сеансов (рис. 7д) имеет рекуррентные структуры, cодержащие длинные диагональные линии. Этой диаграмме соответствует значение показателя DET близкое к 0.9, что указывает на значительное сходство динамики сигнала возбуждения и отклика мозга. Стрелка на рис. 7 указывает на увеличенный фрагмент совместной рекуррентной диаграммы, построенной для интервала времени, равного 1 с, для того, чтобы показать детально диагональные линии, равноотстоящие друг от друга на время, равное периоду колебаний светового сигнала.
Рис. 7в иллюстрирует отсутствие совпадения максимумов вероятностей рекуррентностей ${{P}_{1}}(\tau )$ и ${{P}_{2}}(\tau )$ для фазовых траекторий светового сигнала и ответа мозга, а рис. 7е – наличие такого совпадения в моменты времени kt, где k – целое число, t = 0.1 с (соответствует периоду колебаний светового сигнала частотой 10 Гц). Совпадение положений максимумов вероятностей рекуррентностей ${{P}_{1}}(\tau )$ и ${{P}_{2}}(\tau )$ означает возникновение одновременных рекуррентностей в анализируемом фрагменте ЭЭГ и световом сигнале заданной частоты. В этом случае значение коэффициента взаимной корреляции CPR = 0.61 (рис. 7е). В случае отсутствия совпадения максимумов вероятностей рекуррентностей ${{P}_{1}}(\tau )$ и ${{P}_{2}}(\tau )$ для фазовых траекторий светового сигнала и ответа мозга значение коэффициента взаимной корреляции CPR = 0.01 (рис. 7в). Таким образом, рис. 7 представляет пример возникновения фазовой синхронизации во фрагменте ЭЭГ и световом стимуле частотой 10 Гц после примененных воздействий.
На рис. 8а, 8г представлены фрагменты ЭЭГ длительностью 6 с во время фотостимуляции и фотостимул частотой 6 Гц до и после сеансов активации функциональных связей мозга. Совместные рекуррентные диаграммы для сигнала светового возбуждения и фрагментов отклика мозга до и после этих сеансов показаны на рис. 8б, 8г. Совместная рекуррентная диаграмма, полученная до сеансов (рис. 8б), содержит длинные диагональные линии, соответствующие близкому к 0.95 значению показателя DET и свидетельствующие о значительном сходстве динамики сигнала возбуждения и ответа мозга. Положения максимумов вероятностей рекуррентностей ${{P}_{1}}(\tau )$ и ${{P}_{2}}(\tau )$ для фазовых траекторий светового сигнала и ответа мозга при этом совпадают (рис. 8в) в моменты времени kt, где t = 0.17 с, что соответствует периоду колебаний сигнала фотостимуляции частотой 6 Гц. Значение коэффициента взаимной корреляции CPR = 0.69 (рис. 8в). Это свидетельствует о наличии фазовой синхронизации в анализируемом фрагменте ЭЭГ и световом сигнале заданной частоты.
Рис. 8.
Фрагменты ЭЭГ длительностью 6 с во время фотостимуляции и световой стимул частотой 6 Гц до и после сеансов активации функциональных связей мозга (а, г). Совместные рекуррентные диаграммы этих сигналов (б, д) и вероятности ${{P}_{1}}(\tau )$ и ${{P}_{2}}(\tau )$ рекуррентностей (в, е).
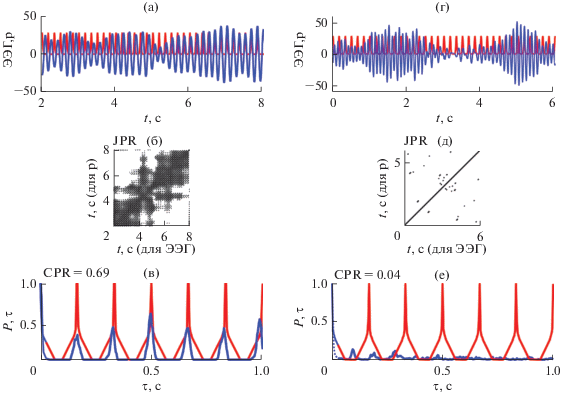
Для фрагмента ЭЭГ, полученного после сеансов активации функциональных связей мозга и светового сигнала частотой 6 Гц (рис. 8г), совместная рекуррентная диаграмма содержит только изолированные точки (рис. 8д), значение коэффициента взаимной корреляции CPR = 0.004 (рис. 8е). Это говорит о том, что в анализируемом фрагменте фазовая синхронизация между сигналом возбуждения и откликом отсутствует. Таким образом, рис. 8 представляет пример отсутствия фазовой синхронизации во фрагменте паттерна ЭЭГ и световом стимуле частотой 6 Гц после проведенных сеансов активации функциональных связей мозга.
Средние значения показателей реакции усвоения до и после сеансов активации функциональных связей мозга приведены в табл. 4. В связи с отсутствием статистически значимых отличий в значениях показателей для разных затылочных локусов ЭЭГ, в табл. 4 приводятся результаты, полученные для затылочного локуса O1.
Таблица 4.
Усредненные значения коэффициентов усвоения (kR) ритма заданной частоты, показателя детерминизма (DET) в совместных рекуррентных диаграммах паттернов ЭЭГ и световых сигналов и показателя фазовой синхронизации (CPR), значения критерия Фишера (F) и уровня значимости критерия (p)
| f (Гц) | До сеансов активации функциональных связей мозга | После сеансов активации функциональных связей мозга | F | p |
|---|---|---|---|---|
| Коэффициент усвоения ритма заданной частоты (kR) | ||||
| 6 | 43 ± 3.7 | 7 ± 0.6 | 467 | 0.005 |
| 10 | <1 | 28 ± 2.6 | 609 | 0.0004 |
| 16 | <1 | <1 | 2.6 | 0.87 |
| Показатель детерминизма (DET) | ||||
| 6 | 0.87 ± 0.07 | 0.51 ± 0.05 | 197 | 0.01 |
| 10 | 0.53 ± 0.06 | 0.97 ± 0.09 | 208 | 0.01 |
| 16 | 0.67 ± 0.07 | 0.56 ± 0.05 | 3.9 | 0.75 |
| Коэффициент взаимной корреляции между вероятностями рекуррентностей (CPR) | ||||
| 6 | 0.49 ± 0.05 | 0.08 ± 0.009 | 401 | 0.003 |
| 10 | 0.05 ± 0.005 | 0.64 ± 0.06 | 377 | 0.002 |
| 16 | 0.06 ± 0.006 | 0.05 ± 0.006 | 2.8 | 0.91 |
Данные табл. 4 показывают, что усвоение частоты фотостимула 16 Гц отсутствует как до, так и после проводимых сеансов (значение коэффициента усвоения ритма kR < 1, процент рекуррентных точек, расположенных на диагональных линиях DET < 0.7, коэффициент взаимной корреляции между вероятностями рекуррентностей светового сигнала и ответа мозга CPR < 0.1, значение F-критерия Фишера F < Fcrit = 4.1 и уровня значимости этого критерия p > 0.05 для всех рассмотренных показателей).
Для частоты светового возбуждения 6 Гц до проводимых сеансов среднее значение коэффициента усвоения ритма kR = 43 ± 3.7, процент рекуррентных точек, расположенных на диагональных линиях DET = 0.87 ± 0.07, среднее значение коэффициента взаимной корреляции между вероятностями рекуррентностей светового сигнала и ответа мозга CPR = 0.49 ± 0.05, а после сеансов средние значения этих показателей уменьшаются до kR = 7 ± 0.6, DET = 0.51 ± 0.05 и CPR = 0.08 ± 0.009. Попарное сравнение средних значений анализируемых показателей вейвлетных спектров и совместных рекуррентных диаграмм до и после сеансов показывает, что значения статистики Фишера F > Fcrit = 4.1 и соответствуют малым значениям уровня значимости p (p < 0.02). Это свидетельствует о наличии достоверных различий между средними значениями рассмотренных показателей для частоты фотостимула 6 Гц до и после проводимых сеансов.
Для частоты светового возбуждения 10 Гц, наоборот, после проводимых сеансов средние значения коэффициента усвоения ритма, процента рекуррентных точек, расположенных на диагональных линиях и коэффициента взаимной корреляции между вероятностями рекуррентностей светового сигнала и ответа мозга увеличиваются до величин kR = 28 ± 2.6, DET = 0.97 ± 0.09 и CPR = 0.64 ± 0.06, по сравнению со средними значениями до этих сеансов kR < 1, DET = 0.53 ± 0.06 и CPR = 0.05 ± 0.005. Значения критерия Фишера F > Fcrit = 4.1 и уровня значимости p < 0.02 демонстрируют наличие достоверных различий между средними значениями рассмотренных показателей для частоты фотостимуляции 10 Гц до и после проводимых сеансов.
Таким образом, оба метода, вейвлетного и реккурентного анализа, указывают на то, что для пациентов с умеренными когнитивными нарушениями после сеансов, направленных на формирование функциональных связей мозга человека, отмечается повышение показателей реакции усвоения ритма частоты возбуждения альфа-диапазона и уменьшается значение этих показателей для частоты возбуждения тета-диапазона.
Уменьшение значений рассмотренных показателей свидетельствует о снижении или даже исчезновении сходства динамического поведения стимула и ответа мозга на него. При этом фазовая синхронизация между откликом мозга и фотостимулом до сеансов наблюдается только для частоты возбуждения тета-диапазона, а после этих сеансов – для частоты возбуждения альфа-диапазона.
Как известно, выраженность перестроек электрической активности мозга в соответствии с частотой, равной или кратной частоте световых импульсов, определяется возбудимостью нейронов коры головного мозга, зависящей от соотношения синхронизирующих и десинхронизирующих процессов [3]. В связи с этим мозг здорового человека или человека с нарушениями функционального состояния центральной нервной системы по-разному реагирует на фотостимуляцию, т.е. по-разному воспроизводит ритмы частот бета-, тета- или альфа-диапазонов [43, 44].
Например, количественные показатели реакции усвоения при нарушениях функционального состояния центральной нервной системы, связанных с сосудистой патологией мозга различной степени выраженности, статистически значимо отличаются от показателей, определенных для здорового человека [5, 6, 43, 44]. По мере нарастания проявлений сосудистой патологии от вегето-сосудистой дистонии к гипертонической болезни и вертебрально-базилярной недостаточности отмечается повышение показателей реакции усвоения частот фотостимуляции [45].
Для здорового человека, как правило, значение коэффициента усвоения ритма меньше единицы или близко к единице, т.е. для него характерна незначительная реакция усвоения ритма световых импульсов в альфа-диапазоне [44]. Расширение диапазона усвоения ритма в сторону тета- или бета-диапазонов и межполушарная асимметрия реакции усвоения ритма связываются с возникновением невротизации пациента, т.е. с повышением уровня его психоэмоциональной возбудимости, и считаются критериями патологии при оценке реакции фотостимуляции [5, 45, 48].
Увеличение показателей реакции усвоения ритма частот тета-диапазона показано в реактивных паттернах ЭЭГ больных с нарушениями сердечного ритма в форме фибрилляции предсердий, причем по мере нарастания проявлений этих нарушений от времени существования мерцательной аритмии, т.е. при переходе от пароксизмальной формы аритмии в постоянную, значение коэффициента усвоения ритма в тета-диапазоне десятикратно возрастает, в то время как в альфа-диапазоне это значение существенно уменьшается, что, вероятно, связано с преобладанием процессов торможения в нервной системе этих пациентов [5, 46].
Отсутствие или незначительность реакции усвоения ритма в диапазоне тета- и бета-частот на фоне слабого усвоения альфа-ритма у здорового человека отражает сбалансированность процессов возбуждения и торможения и преобладание внутренней синхронизации нейронных структур при навязывании внешнего ритма [24, 25]. Повышение показателей реакции усвоения ритмов фотостимуляции связано, как правило, с повышением неустойчивости внутренней синхронизации нейронных ансамблей и усилением их внешней синхронизации [6, 32, 33].
Усиление реакции усвоения в альфа-диапазоне и ослабление в тета-диапазоне указывает на улучшение функционального состояния коры больших полушарий и улучшение ее регуляторных механизмов. Снижение степени невротизации пациентов с невротическими расстройствами в виде панических атак после сеансов, направленных на формирование функциональных связей мозга, отражается в уменьшении количественных показателей реакции усвоения ритмов фотостимуляции бета-диапазона и связано с понижением возбудимости нейронов коры больших полушарий [7, 46].
В случае умеренных когнитивных нарушений повышение количественных показателей реакции усвоения ритма частоты альфа-диапазона, вероятно, связано с повышением возбудимости и лабильности корковых нейронов после проведенных сеансов.
ЗАКЛЮЧЕНИЕ
В настоящей работе продемонстрирована возможность исследования динамики паттернов электрической активности мозга при таких дезадаптационных нарушениях, как тревожно-фобические расстройства, панические атаки и умеренные когнитивные нарушения, с помощью методов вейвлетного, мультифрактального и реккурентного анализа.
Показано, что коррекция психогенной боли при тревожно-фобических состояниях связана с изменением динамики последовательных значений ЭЭГ и снижением степени их мультифрактальности.
Анализ изменений реакции усвоения ритмов фотостимуляции и определение количественных характеристик этой реакции, вычисленных до и после сеансов терапии, связанных c воздействием, направленным на активацию функциональных связей мозга у лиц с паническими атаками и умеренными когнитивными нарушениями, позволяет выявить объективные показатели улучшения функционального состояния нервной системы после этих сеансов. К таким показателям относятся коэффициент усвоения ритма заданной частоты возбуждения, вычисленный на основе вейвлетного анализа реактивных паттернов ЭЭГ, и коэффициент взаимной корреляции между вероятностями рекуррентностей светового сигнала и ответа мозга на определенные частоты, вычисленный на основе анализа совместных рекуррентных диаграмм ритмических световых сигналов и ответов мозга на эти сигналы.
Рассмотренные результаты являются еще одним доказательством высокой чувствительности примененных методов анализа вейвлетных спектров и показателей совместных рекуррентных диаграмм к выявлению динамических изменений, происходящих в реактивных паттернах биоэлектрической активности мозга в ответ стимулирующие воздействия.
Эти результаты могут быть использованы в клинике для достоверного нахождения улучшения функционального состояния головного мозга пациентов с дезадаптационными нарушениями.
Работа выполнена при финансовой поддержке Государственной программы РФ 47 ГП “Научно-технологическое развитие Российской Федерации” (2019–2030), подпрограмма “Фундаментальные научные исследования для долгосрочного развития и обеспечения конкурентоспособности общества и государства” (47_110_ДРиОК).
Список литературы
Антипенко Е.А. Антиооксидантная терапия при дисциркуляторной энцефалопатии // Журн. неврологии и психиатрии им. С.С. Корсакова. 2010. № 110. С. 53.
Дик О.Е., Глазов A.Л. Применение анализа совместных рекуррентностей к оценке фазовой синхронизации физиологических сигналов // Журн. технической физики. 2021. Т. 91. Вып. 12. С. 2045–2058.
Дик О.Е., Святогор И.А., Ишинова В.А., Ноздрачев A.Д. Фрактальные характеристики функционального состояния мозга у пациентов с тревожно-фобическими расстройствами // Физиология человека. 2012. Т. 38. С. 30–36.
Дик O.E. Вейвлет и мультифрактальный анализ реакции мозга человека на ритмическую фотостимуляцию при сосудистой патологии // XVIII Всероссийская научно-техническая конференция с межд. участием Нейроинформатика-2016, 25–29 апреля 2016 НИЯУ МИФИ: Сб. научных трудов в 3-х частях. Ч. 3. М.: НИЯУ МИФИ. 2016. С. 12–20.
Дик О.Е., Ноздрачев А.Д Динамика паттернов электрической активности мозга при нарушениях его функционального состояния // Успехи физиологических наук. 2020. Т. 51. № 2. С. 1–20.
Дик О.Е., Святогор И.А., Дик О.Е. и др. Анализ реактивных паттернов ЭЭГ у лиц с фибрилляцией предсердий // Физиология человека. 2019. Т. 45. С. 49–63.
Дик О.Е., Ноздрачев А.Д. Механизмы изменения динамической сложности паттернов физиологических сигналов (научная монография). СПб.: Изд-во Санкт-Петербургского университета, 2019. 200 с. ISBN 978-5-288.
Дик О.Е., Святогор И.А., Резникова Т.Н. и др. Анализ паттернов ЭЭГ у лиц с паническими атаками // Физиология человека. 2020. Т. 46. № 2.С. 63.
Дик О.Е., Глазов A.Л. Параметры фазовой синхронизации в электроэнцефалографических паттернах как маркеры когнитивных нарушений // Журн. технической физики. 2021. Т. 91. Вып. 4. С. 678–688.
Добеши И. Десять лекций по вейвлетам. Ижевск: НИЦ Регулярная и стохастическая динамика, 2001. 115 с.
Дремин И.М., Иванов О.В., Нечитайло В.А. Вейвлеты и их использование // Успехи физических наук. 2001. Т. 171. С. 465–501.
Дробижев М.Ю., Макух Е.А., Дзантиева А.И. Когнитивные расстройства в общей медицине // Лечащий врач. 2007. № 8. С. 35.
Ишинова В.А., Святогор И.А. Цветовое отражение боли у пациентов с тревожно–фобическими расстройствами // Вестник С.-Пб. гос. Мед. Акад. им Мечникова. СПб. 2009. Т. 31. С. 198–199.
Луговая А.М. Способ коррекции психоэмоционального состояния по методу А.М. Луговой. Патент РФ N 2313282. 2007. 22 с.
Новикова И.А., Соловьев А.Г., Местечко В.В. Особенности эмоциональной сферы лиц пожилого возраста с пограничными психическими расстройствами // Медицинская психология в России: электрон. науч. журн. 2014. № 4. С. 27.
Павлов А.Н., Анищенко В.С. Мультифрактальный анализ сложных сигналов // Успехи физических наук. 2007. Т. 177. С. 859.
Парфенов В.А. Лечение постинсультных когнитивных нарушений // Русский медицинский журнал. 2010. № 16. С. 986.
Резникова Т.Н., Краснов А.А., Селиверстова Н.А. и др. Исследование “внутренней картины болезни” у больных с органической и функциональной патологией ЦНС в процессе лечебных активаций методом артифициальных стабильных функциональных связей мозга человека // Вестник Клинической психологии. 2004. Т. 2. № 1. С. 84.
Резникова Т.Н., Федоряка Д.А., Селиверстова Н.А., Моховикова И.А. Опыт использования сенсорной импульсной стимуляции у больных с паническими атаками // Вестник психотерапии. 2018. № 68. С. 47.
Резникова Т.Н. Селиверстова Н.А. О коррекции психического состояния немедикаментозным способом при рассеянном склерозе // Клиническая и специальная психология // 2019. Т. 8. № 1. С. 177.
Резникова Т.Н., Селиверстова Н.А., Дик О.Е. и др. Оценка психофизиологического состояния у пожилых лиц с умеренными когнитивными нарушениями при сенсорных импульсных стимуляциях // Психическое здоровье. 2020. № 9. С. 12–18.
Русинов В.С., Гриндель О.М., Болдырева Г.Н., Вакар Е.М. Биопотенциалы мозга человека. Математический анализ. М.: Медицина, 1987. 254 с.
Святогор И.А. Классификация ЭЭГ паттернов и их нейрофизиологическая интерпретация при дезадаптационных расстройствах // Биологич. Обратная Связь. 2000. Т. 3. С. 10–19.
Святогор И.А., Резникова Т.Н., Петухова Н.В. Динамика психофизиологического состояния в процессе лечения методом артифициальных стабильных функциональных связей // Биологическая обратная связь. 2001. № 4. С. 24.
Святогор И.А., Гусева Н.Л. ЭЭГ-реакция усвоения ритма в норме и при нарушении функционального состояния центральной нервной системы // Вестник клинической нейрофизиологии. 2014. Т. 1. С. 13–19.
Святогор И.А., Дик О.Е., Ноздрачев А.Д. и др. Анализ изменений ЭЭГ-паттернов в ответ на ритмическую фотостимуляцию при различных нарушениях функционального состояния ЦНС // Физиология человека. 2015. Т. 41. С. 41–49.
Серов Н.В. Световая терапия. Смысл и значение цвета информация–цвет–интеллект. СПб.: Речь. 2002. 160 с.
Смирнов В.М., Резникова Т.Н., Губачев Ю.М. Артифициальные стабильные функциональные связи и нейро-регуляторная коррекция психовегетативных расстройств // Физиол. чел. 1987. Т. 13. № 5. С. 715–722.
Смирнов В.М., Бородкин Ю.С., Резникова Т.Н. Свойство мозга человека в условиях активации подкорковых структур формировать комплекс устойчивых внутримозговых функциональных связей. В кн.: Научные открытия (Сборник кратких описаний). М.-СПб., 1999. Вып. 1. С. 23–26.
Старчина Ю.А. Когнитивные нарушения и их лечение у пациентов с артериальной гипертензией // Consilium medicum. 2009. № 1. С. 1.
Табеева Г.Р. Когнитивные и некогнитивные расстройства у пациентов пожилого возраста, ассоциированные со стрессом // Неврология, нейропсихиатрия, психосоматика. 2015. № 1. С. 87.
Федотчев А.И., Бондарь А.Т., Акоев И.Г. Резонансные ЭЭГ – реакции при ритмических световых воздействиях разной интенсивности и частоты // Журн. ВНД. 2001. Т. 51. № 1. С. 17–23.
Федотчев А.И., Бондарь А.Т., Матрусов С.Г. и др. Использование сигналов обратной связи от эндогенных ритмов пациента для нелекарственной коррекции функциональных расстройств // Успехи физиол. наук. 2006. Т. 37. № 4. С. 82–92.
Arneodo A., Bacry E., Muzy J.F. The thermodynamics of fractals revisited with wavelets // Physica A. 1995. V. 213. P. 232–275.
Bass C., Pevele R., House A. Somatoform disorders: severe psychiatric illnesses neglected by psychiatrists// British J. Psychiatry. 2001. V. 179. C. 11–14.
Batelaan N.M., Van Balkom A.J., Stein D.J. Evidence -based pharmacotherapy of panic disorder: An update // Intern. J. neuropsychopharmacology. 2012. V. 15. № 3. P. 403–415.
Bierman E.J.M., Comijs H.C., Rijmen F. et al. Anxiety symptoms and cognitive performancein later life: results from the longitudinal aging study // Aging Ment Health. 2008. V. 12. № 4. P. 517.
Bosnyakov D., Gabova A., Kuznetsova G. et al. Time–frequency analysis of spike–wave discharges using a modified wavelet transform // J. Neurosci. Methods. 2006. V. 1654. P. 80–88.
Borodina U.V., Aliev R.R. Wavelet spectra of visual evoked potentials; time course of delta, theta, alpha and beta bands // Neurocomputing. 2013. V. 121. P. 551–555.
Bozhokin S.V., Suvorov N.B. Wavelet analysis of transient processes in EEG photic // Biomed. Electron. 2008. V. 3. P. 13–19.
Cosci F. The psychological development of panic disorder: implications for neurobiology and treatment // Rev Bras Psiquiatr. 2012. V. 34. P. 9–31.
Dick O.E., Mochovikova I.A. Multifractal and wavelet analysis of epileptic seizures // Chaos Theory: Modeling, Simulation and Applications, Selected Papers from the 3rd Chaotic Modeling and Simulation Intern. Conference / C.H. Skiadas, I. Dimotikalis and C. Skiadas (Eds). World Scientific Publishing. 2011. P. 159–166.
Dick O.E., Svyatogor I.A. Potentialities of the wavelet and multifractal techniques to evaluate changes in the functional state of the human brain // Neurocomputing. 2012. V. 82. P. 207–215.
Dick O.E. Wavelet analysis of the human brain lability to reproduce the external rhythm // J. Chaotic Modeling and Simulation. 2013. V. 3. P. 387–394.
Dick O.E., Svyatogor I.A. Wavelet and multifractal estimation of the intermittent photic stimulation response in the electroencephalogram of patients with dyscirculatory encephalopathy // Neurocomputing. 2015. V. 165. P. 361–374.
Dick O.E. Nonlinear dynamics of reactive EEG patterns under cerebrovascular and cardiovascular distortions // J. Chaotic Modeling and Simulation. 2017. V. 4. P. 501–512.
Dick O.E. Wavelet and recurrence analysis of EEG patterns of subjects with panic attacks // Studies in Computational Intelligence. 2020. V. 856. P. 172.
Dick O.E., Glazov A.L. Estimation of the synchronization between intermittent photic stimulation and brain response in hypertension disease by the recurrence and synchrosqueezed wavelet transform // Neurocomputing. 2021. V. 455. P. 163–177.
Fedotchev I., Bondar A.T., Akoev I.G. Dynamic characteristics of the human resonance EEG responses to rhythmic photostimulation // Human Physiology. 2000. V. 26. № 2. P. 64–72.
Fraser A.M., Swinney H.L. Independent coordinates for strange attractors from mutual information // Phys. Rev. 1986. V. 33. № 2. P. 1134–1140.
Hramov A.E., Koronovsky A.A., Makarov V.A. et al. Wavelets in neuroscience. Springer Series in Synergetics. Berlin: Springer, 2015. 314 p.
Gallacher R. Early stage dementia // Nursing Standard. 2014. V. 28. № 20. P. 61.
Goddard A.W., Mason G.F., Appel M. et al. Impaired GABA neuronal response to acute benzodiazepine administration in panic disorder // Am. J. Psychiatry. 2004. V. 161. № 12. P. 2186–2193.
Ishinova V.A., Svyatogor I.A., Reznikova T.N. Features of color reflection in psychogenic pain in patients with somatoform disorders during psychotherapeutic treatment // Spanish J. Psychology. 2009. V. 12. P. 715–724.
Kennel M.B., Brown R., Abarbanel H.D. Determining embedding dimension for phase-space reconstruction using a geometrical construction // Phys. Rev. A. 1992. V. 45. № 6. P. 3403.
Kurths J., Romano M.C., Thiel M. et al. Synchronization analysis of coupled noncoherent oscillators // Nonlinear Dynamics. 2006. V. 44. P. 135.
Marwan N., Wessel N., Meyerfeldt U. et al. Recurrence plot based measures of complexity and ist application to heart rate variability data // Physical Review E. 2002. V. 66. P. 26702–26710.
Marwan N., Kurths J. Nonlinear analysis of bivariate data with cross recurrence plots// Physics Letters A. 2002. V. 302. P. 299–307.
Marwan N., Romano M.C., Thiel M. et al. Recurrence plots for the analysis of complex systems // Physics Reports. 2007. V. 438. P. 237–329.
Milrod B., Busch F., LeonA C. et al. A pilot open trial of brief psychodynamic psychotherapy for panic disorder // J. Psychother. Pract. Res. 2001.V. 10. P. 239–245.
Muzy J.F., Bacry E., Arneodo A. Multifractal formalism for fractal signals: the structure–function approach versus the wavelet–transform modulus–maxima method // Phys. Rev. E. 1993. V. 47. P. 875–884.
Natarajan K., Acharya R., Alias F. et al. Nonlinear analysis of EEG signals at different mental states // BioMedical Engineering. 2004. V. 3. P. 7–18.
Nutt D. Anxiety disorders comorbid with depression: panic disorder and agoraphobia / D. Nutt, A. Feeney, S. Argyropolous. Martin Dunitz Ltd. 2002. 110 p.
Popivanov D., Stomonyakov V., Minchev Z. et al. Multifractality of decomposed EEG during imaginary and real visual–motor tracking // Biological Cybernetics. 2006. V. 94. P. 149–156.
Qianli M.A., Xinba N., Jun W. et al. A new measure to characterize multifractality of sleep electroencephalogram // Chinese Science Bulletin. 2006. V. 51. P. 3059–3064.
Qureshi S.U., Long M.E., Bradshaw M.R. et al. Does PTSD impair cognition beyond the effect of trauma? // J. Neuropsychiatry Clin. Neurosci. 2011. V. 23. № 1. P. 16.
Rangaprakash D., Pradhan N. Study of phase synchronization in multichannel seizure EEG using nonlinear recurrence measure // Bioledical Signal Processing and Control. 2014. V. 11. P. 114–112.
Romano M.C., Thiel M., Kurths J. et al. Detection of synchronization for non-phase-coherent and non-stationary data // Europhysics Letters. 2005. V. 71. P. 466.
Senhadji L., Wendling F. Epileptic transient detection: wavelets and time–frequency approaches // Neurophysiol. Clin. 2002. V. 32. P. 175–192.
Sullivan G.M., Coplan J.D., Kent J.M. et al. The noradrenergic system in pathological anxiety: a focus on panic with relevance to generalized anxiety and phobias // Biol. Psychiatry. 1999. V. 46. № 9. P. 1205–1218.
Takens F. Detecting strange attractors in turbulence / In: Dynamical Systems and Turbulence, Lecture Notes in Mathematics / D. Rand, L.S. Young, eds. Berlin: Springer, 1981. V. 898. P. 366–381.
Titov V.E., Dick O.E. Computational evaluation of the effectiveness of therapy method with help recurrent analysis // J. Physics: Conference Series. 2021. V. 1889. P. 42092–42098.
Westenberg H.G., Liebowitz M.R. Overview of panic and social anxiety disorders // J. Clin. Psychiatry. 2004. V. 65. Suppl. 14. P. 22–26.
Wilson K.A., Hayward C. A prospective evaluation of agoraphobia and depression symptoms following panic attacks in a community sample of adolescents // J. Anxiety Disord. 2005. V. l. 19. № 1. P. 87–103.
Wink A.M., Bullmore E., Barnes A. et al. Monofractal and multifractal dynamics of low frequency endogenous brain oscillations in functional MRI // Human Brain Mapping. 2008. V. 29. P. 791–801.
Xu Y., Ma Q.DY., Schmitt D.T. et al. Effects of coarse–graining on the scaling behavior of long–range correlated and anti–correlated signals // Physica A. 2011. V. 390. P. 4057–4072.
Yamamoto M. Fluctuations observed in biological time series signals and their functional significance // Front. Med. Biol. Eng .1991. V. 3. P. 135–137.
Yu S.T., Kim M.K., Kim B. et al. The Effects of 5-HTR1A Polymorphism on Cingulum Connectivity in Patients with Panic Disorder // Psychiatry Investig. 2013. V. 10. № 4. P. 399–406.
Дополнительные материалы отсутствуют.
Инструменты
Успехи физиологических наук