

Вестник РАН, 2021, T. 91, № 3, стр. 273-282
БИОГЕОЦЕНОТИЧЕСКИЕ ПРИНЦИПЫ РЕСТАВРАЦИИ ПАСТБИЩ В ЦЕНТРАЛЬНОАЗИАТСКОЙ ПУСТЫНЕ
З. Ш. Шамсутдинов a, *, Н. З. Шамсутдинов b, **, Н. С. Орловский c, ***, Э. З. Шамсутдинова a, ****
a Федеральный научный центр кормопроизводства и агроэкологии им. В.Р. Вильямса
г. Лобня, Московской области, Россия
b Всероссийский научно-исследовательский институт гидротехники и мелиорации им. А.Н. Костякова,
Москва, Россия
c Институт изучения пустынь им. Дж. Блаустейна Университета Бен Гуриона
Седе Бокер, Израиль
* E-mail: aridland@mail.ru
** E-mail: nariman@vniigim.ru
*** E-mail: nicolai@bgu.ac.il
**** E-mail: darplant@list.ru
Поступила в редакцию 09.09.2020
После доработки 13.01.2021
Принята к публикации 19.01.2021
Аннотация
Возрастающее антропогенное и техногенное воздействие на почвенно-растительный покров и водные ресурсы Центральной Азии резко ускорили деградацию и опустынивание природных пастбищ этого региона, что сопровождалось ухудшением кормовой базы животноводства, а в итоге и качества жизни населения. Теоретическое обоснование методов экономически выгодной экологической реставрации утраченного биоразнообразия и кормовой производительности опустыненных земель – важная научная задача. Для её решения авторы статьи предлагают использовать подходы, базирующиеся на биогеоценотических принципах формирования поливидовых многоярусных пастбищных биогеоценозов, воссоздания оптимальных флороценотических структур путём введения в их состав зональных базовых доминантных видов и жизненных форм кормовых растений. Такие воссозданные биогеоценозы обладают свойством самовозобновления и самоподдержания естественной структурно-функциональной организации. Они имеют и большое природоохранное значение, поскольку способствуют приостановке процессов опустынивания аридных пастбищных земель, прекращению пыльных бурь, восстановлению оптимальных параметров окружающей среды.
Пустынно-пастбищный природный комплекс Центральной Азии объединяет обширную по площади и экологически обособленную часть территорий стран этого региона с его крайне засушливым климатом и экстремальными природными условиями. Естественные аридные пастбища, занимающие огромные пространства, служат основой кормовой базы овцеводства, мясного скотоводства, табунного коневодства и верблюдоводства [1, 2]. Образно говоря, это столовые для животных, где они круглый год питаются, и дом, где проходит вся их жизнь. Здесь овцы, козы, верблюды, лошади рождаются, взрослеют, давая затем человеку возможность получать от них ценные продукты [3].
Следует особо отметить важнейшую фундаментальную особенность природных пастбищ – их способность к постоянному самовозобновлению и самовоспроизводству биологической (кормовой) продукции. Пастбищные биогеоценозы – неисчерпаемый возобновляемый биологический ресурс в отличие от исчерпаемых минеральных.
Значение природных флористически и ценотически полночленных природных пастбищ Центральной Азии выходит далеко за пределы интересов животноводства. Как важнейший компонент биосферы они не только оказываются источником кормов, но и определяют состояние земельных ресурсов, плодородие почв, генетическое и ценотическое биоразнообразие, а также качество среды обитания человека в этом обширном регионе. Однако непрерывный рост численности населения, развитие животноводства, интенсивное горнорудное и ирригационное освоение аридных районов Центрально-Азиатского региона, оказывающих всевозрастающее антропогенное и техногенное воздействие на почвенно-растительный покров и водные ресурсы, без учёта возможных экологических последствий, стали причиной деградации и опустынивания природных пастбищ [4]. Эти негативные процессы вызвали обеднение биоразнообразия, снижение производительности биогеоценозов и, как следствие, ухудшение кормовой базы пастбищного животноводства, а в итоге и качества жизни населения. Подобные тревожные процессы характерны не только для Центральной Азии.
Учёные разных стран сходятся во мнении, что масштабы нынешней деградации окружающей среды и разрушения многих составляющих биоты Земли значительны, причём изменения происходят в “катастрофически короткие сроки” [5, 6], и это требует интенсивного международного сотрудничества при участии научного сообщества. Главные задачи, которые уже сегодня необходимо решать общими усилиями всех стран, – сохранение и поддержание жизнеспособной природной среды обитания человека, а также восстановление деградированных составляющих биоты [7, 8]. Генеральная Ассамблея ООН резолюцией 73/284, принятой на 69-м пленарном заседании 1 марта 2019 г., постановила провозгласить 2021–2030 годы “Десятилетием Организации Объединённых Наций по восстановлению экосистем” и призвала государства – члены ООН содействовать укреплению политической воли, мобилизации имеющихся ресурсов, усилению научных исследований в области восстановления экосистем на глобальном, региональном, национальном и местном уровнях [9].
Биогеоценотические принципы экологической реставрации. Основанное в 1988 г. Общество экологической реставрации (Society for Ecological Restoration), объединяющее учёных разных стран, определяет такого рода реставрацию как деятельность человека по инициированию и ускорению восстановления экосистемы с целью её оздоровления, поддержания целостности и устойчивости, в конечном счёте – возвращения к её исторической эволюционной траектории [10]. В этом контексте задачи экологической реставрации утраченных биоразнообразия и кормовой производительности опустыненных пастбищных земель в аридных районах Центральной Азии предопределяют необходимость биогеоценотического подхода к решению обостряющейся масштабной проблемы [11–13]. Он может быть реализован на основе высева фитоценотически сбалансированных комбинаций (смесей) зонально типичных жизненных форм и доминантных видов кормовых растений климаксовых (стабильных) сообществ – представителей дикорастущей флоры. Методологической основой их подбора служит принцип системности, базирующийся на фундаментальных постулатах биогеоценологии. Согласно им, природные пастбища – это целостные образования с присущей им функциональной взаимосвязью базовых компонентов растительного сообщества, макро- и микрофаунистического комплекса и почвы. При этом сами растительные сообщества рассматриваются в качестве системообразующей структуры биогеоценоза [14–17].
Перечислим биогеоценотические принципы, на которые мы опирались при разработке адаптивных методов экологической реставрации опустыненных пастбищ Центральной Азии.
1. Принцип доминанты зонально типичных жизненных форм растений [16, 18, 19]. Каждой природной зоне присущ свой зональный тип биогеоценоза, носителем которого служат жизненные формы растений. Для аридных условий Центральной Азии характерны две синузии (группы особей одного или сходных видов, образующих ярусы) жизненных форм: весенние мезофильные эфемеры и эфемероиды, использующие весенние осадки, и галоксерофильные виды кустарников родов Haloxylon, Salsola и полукустарничков родов Artemisia, Kochia, Camphorosma, Eurotia и другие виды, использующие водно-минеральные ресурсы большого объёма почвы, обладающие способностью произрастать и господствовать в растительном покрове в сухое время года. При этом в ходе восстановительных работ на пастбищах, конструируемых по типу зонально типичных биогеоценотических структур с использованием доминантных видов, достигается более полное заполнение фундаментальной экологической ниши и оптимизация расходования ресурсов среды. Применительно к условиям аридных зон Центральной Азии это различные комбинации кормовых галоксерофильных кустарников, полукустарничков и эфемероидных трав.
2. Принцип типов адаптивной стратегии растений. Российский эколог, один из основателей учения о морфологии географического ландшафта Л.Г. Раменский [20] выделил три типа стратегии: виоленты (“силовики”), он уподобил их львам растительного покрова, патиенты (“выносливцы”) – верблюдам, эксплеренты (“выполняющие”) – шакалам. Фундаментальные исследования по этому вопросу проведены британским экологом Дж. Граймом [21, 22], который фактически переоткрыл три типа стратегии, ранее сформулированные Л.Г. Раменским [23]. Типы адаптивных стратегий Дж. Грайм предложил обозначать буквенными индексами: конкуренты (виоленты по Раменскому) – С, стресс-толеранты (по Раменскому патиенты) – S и рудералы (по Раменскому эксплеренты) – R [22].
Выявление типов адаптивных стратегий имеет существенное значение для анализа и понимания биогеоценотических механизмов восстановительной сукцессии. Тип растений с виолентными свойствами (С-виды) высококонкурентен, способен благодаря быстрому росту захватывать и длительное время удерживать за собой захваченную территорию, подавляя соперников энергией жизнедеятельности и полнотой использования ресурсов среды. Растения с патиентным типом стратегии (S-виды) исключительно выносливы к крайне жёсткой окружающей среде, способны выживать даже в неоптимальных абиотических и биотических условиях. Третий тип адаптивной стратегии – эксплеренты (R-виды) наделены низкой способностью к конкуренции, но зато могут быстро захватывать освобождающиеся территории. Знание типов адаптивной стратегии растений, участвующих в формировании полночленных кустарниково-полукустарничково-травянистых биогеоценозов, которые зонально типичны для ксеротермических условий пустыни, как нам представляется, имеет ключевое значение в постижении биогеоценотических механизмов ускоренной восстановительной сукцессии на опустыненных землях, лишённых базовой растительности.
3. Принцип дифференциации экологической ниши занимает важное место в теоретической экологии [24]. Понятие экологической ниши в известной мере объясняет, каким образом различные виды могут нормально функционировать в конкретном экотопе [25]. Принцип дифференциации и упаковки экологических ниш реализуется на всех стадиях восстановительной сукцессии за счёт совмещения растений с разным типом развития, распределения корневых систем в почвенной среде, с неодинаковой потребностью в элементах минерального питания и солнечной радиации, разной способностью к усвоению труднодоступных форм почвенной влаги, фосфора и калия, с разной феноритмикой [26, 27]. Участие растений с различающейся экологией и биологией в процессе сукцессионного восстановления зонально типичных кустарниково-полукустарничково-травянистых биогеоценозов может происходить на основе более рациональной упаковки экологических ниш и взаимного дополнения видов. При этом процесс дифференциации ниш должен происходить на основе флуктуационной, сезонной, ярусной, функциональной и сукцессионной взаимодополняемости видов [28, 29].
4. Следующий важный биогеоценотический принцип, имеющий существенное значение для понимания процессов ускоренного восстановления кустарниково-полукустарничково-травянистых пастбищ – адекватность конструкции моделируемых биогеоценозов зональным схемам естественных пустынных биогеоценотических структур. Основным носителем биогеоценотической структуры, по Г.М. Зозулину [19] и В.С. Залетаеву [30], служат зонально типичные жизненные формы растений.
С опорой на эти биогеоценотические принципы нами разработаны адаптивные методы экологической реставрации опустыненных пастбищных земель. В их основе – высев экологически различающихся видов кормовых растений в фитоценотически сбалансированных комбинациях с целью формирования саморегулирующихся и самовозобновляющихся пастбищ.
Адаптивные биогеоценотические методы экологической реставрации опустыненных земель. Для формирования кустарниково-полукустарничково-травянистых осенне-зимних поливидовых пастбищ мы использовали в разном процентном соотношении галоксерофильный кустарник саксаул чёрный (Haloxylon aphyllum (Minkw.) Iljin) – 20%, ксерогалофильные полукустарнички кохию простёртую (Kochia prostrata (L.) Schrad.) – 30% и полынь развесистую (Artemisia diffusa Krasch. ex Poljakov) – 30%, эфемероид мятлик луковичный (Poa bulbosa L.) – 20%. Смесь перечисленных видов кормовых растений высеяли под зиму, в ноябре в урочище Тутли (пустыня Карнабчуль на юге Узбекистана).
На рисунке 1 представлена структура кустарниково-полукустарничковых осенне-зимних пастбищ с чётко выраженными четырьмя синузиями (ярусами). По типу адаптивной стратегии представитель первого яруса саксаул чёрный (Haloxylon aphyllum) относится к виолентным растениям. В возрасте 7 лет он занимает верхний ярус надземной сферы полидоминантного сообщества, достигая высоты 400–450 см при диаметре кроны 320–350 см. Что касается подземной сферы, то его корневая система занимает экологическую нишу в слое почвы 0–1350 см. В горизонтальном направлении корни распространяются на длину от 200 до 450 см. Чаще всего деление главного корня прослеживается на глубине 180–450 см, ниже на всю глубину от корней второго порядка отходят корни длиной 100–200 см с мелкими всасывающими корешками на концах. Вокруг крупных корней на всю глубину наблюдается значительное увлажнение почвы по сравнению с участками, отдалёнными от них. Очевидно, эти корни выделяют воду в сухие слои почвы. Это явление под названием “гидравлический лифт” описано В.Г. Онипченко [16]. Корни достигают уровня капиллярно увлажняемых грунтовыми водами слоёв и распадаются на большое количество тонких поглощающих волосков.
Рис. 1.
Вертикальный профиль кустарниково-полукустарничковых осенне-зимних пастбищ в возрасте 7 лет:
1 – саксаул чёрный (Haloxylon aphyllum), 2 – кохия простёртая (Kochia prostrata), 3 – полынь развесистая (Artemisia diffusa), 4 – мятлик луковичный (Poa bulbosa). Длина профиля 6.5 м
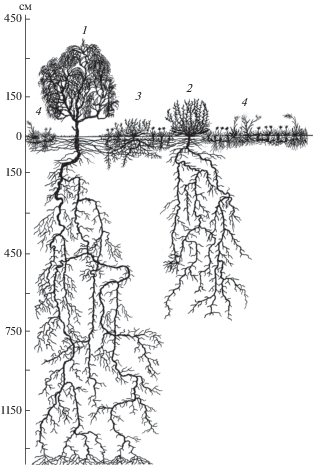
До влажного слоя почвы, залегающего на глубине 700–800 см, корни саксаула идут вниз более или менее отвесно, образуя в рыхлых участках плотных слоёв почвогрунта клубки волосовидных тонких поглощающих корешков. Во влажных слоях, где оптимальная влажность почвогрунта более или менее постоянна за счёт грунтовых вод, корни разветвляются и боковые располагаются в горизонтальном направлении. Здесь очень много белых всасывающих корешков. Пробивая плотные, напоминающие бетон слои грунта, корни утончаются, изгибаются и сплющиваются. Нередко такие слои они пробивают в сплетённом виде. Сплетение, как правило, бывает двойное, иногда тройное.
Вторая синузия представлена кохией простёртой (Kochia prostrata). По жизненной форме это полукустарничек, по экологии – ксерогалофит, по адаптивной стратегии – виолент. В надземной сфере средняя высота этого растения – 65 см. На кусте в среднем 29 генеративных (длиной 56–70 см) и 54 густо облиственных (длиной 12–30 см) вегетативных побега. В подземной сфере корневая система кохии занимает экологическую нишу в толще почвы 0–680 см, в горизонтальном направлении достигают длины 165 см. Первое деление главного корня отмечается в слое 50–90 см. На глубине 350–400 см от корней первого порядка отходят 6–7 тонких (0.3–0.4 см) корней, на которых развиваются многочисленные очень мелкие корешки, а на глубине 680 см присутствует большое количество тонких разветвлений и поглощающих волосков.
Ярус полыни развесистой (Artemisia diffusa) – зонально типичного ксерогалофильного полукустарничка – занимает, по сравнению с саксаулом чёрным и кохией простёртой, небольшой объём экологической ниши в воздушной и почвенной среде. Высота растения в среднем 32 см, ширина куста до 40 см. Корневая система охватывает слой почвы 0–120 см. На глубине 35–40 см главный корень делится на 4 боковых толщиной 0.3–0.4 см, они заканчиваются многочисленными мелкими поглощающими корешками.
Синузия ксерофильных эфемероидов представлена осокой толстостолбиковой (Carex pachystylis J. Gay) и мятликом луковичным (Poa bulbosa L.). К ним примешиваются единичные экземпляры костреца кровельного (Bromus tectorum L.) и костреца дантониевидного (Bromus danthoniae Trin.). Высота мятлика луковичного, как и эфемеровых злаков, не превышает 20 см, а его корневая система мочковатая и характеризуется поверхностным расположением. На 7-м году жизни он сильно разрастается, образуя сплошную и плотную дернину. Ниша, занимаемая эфемероидами в подземной сфере, ограничивается слоем почвы толщиной до 16 см (осока толстостолбиковая) и до 35 см (мятлик луковичный).
Полидоминантные многоярусные осенне-зимние пастбища, имеющие в своём составе кормовые кустарники, полукустарнички, эфемероидные травы, – это сообщество фототрофных растений, объединённых по принципу максимального использования водных ресурсов и элементов минерального питания почвенной среды. Между фототрофами преобладают кооперативные отношения, обеспечивающие более благоприятные условия существования.
В процессе восстановительной сукцессии кустарниково-полукустарничково-травянистых пастбищных сообществ экологические ниши дифференцируются путём ярусного и сезонного взаимодополнения участвующих в ней видов растений. Популяция саксаула чёрного занимает верхний ярус, кохии простёртой – второй, полыни развесистой – третий, а эфемероиды (мятлик луковичный, осока толстостолбиковая) и эфемеры – нижний ярус. Ярусное расположение кроны обеспечивает формирование оптимальной инфраструктуры размещения листового аппарата для эффективного фотосинтеза.
Наряду с ярусным взаимодополнением упаковка ниш в указанных сообществах происходит и по принципу сезонного взаимодополнения фенологически разноритмичных видов растений: саксаула чёрного, кохии простёртой с продолжительностью вегетационного периода 226–242 дня. У полыни развесистой также достаточно длительный период вегетации, но с летней депрессией жизненных процессов. А у мятлика луковичного и осоки толстостолбиковой он короток – с февраля до середины апреля, то есть они используют ресурсы среды, особенно запасы почвенной влаги, до начала отрастания кустарников и полукустарничков. Благодаря разновременной вегетации, выстраиванию своего рода экологической очереди скудные водно-минеральные ресурсы среды используются оптимально. Данные, характеризующие кормовую продуктивность кустарниково-полукустарничково-травянистых осенне-зимних пастбищ в пустыне Карнабчуль, приведены на рисунке 2.
Рис. 2.
Кормовая продуктивность кустарниково-полукустарничково-травянистых осенне-зимних пастбищ в пустыне Карнабчуль:
а – саксаул чёрный (Haloxylon aphyllum), б – кохия простёртая (Kochia prostrata), в – полынь развесистая (Artemisia diffusa), г – мятлик луковичный (Poa bulbosa)
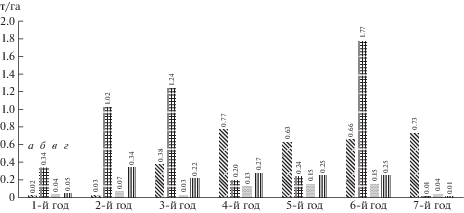
Со 2–3 года вегетации кормовая продуктивность полидоминантных осенне-зимних пастбищ в наших опытах увеличивалась (рис. 3), достигая к середине октября, в зависимости от складывающихся метеорологических условий, 0.6–12.0 т/га сухой кормовой массы. Для сравнения: продуктивность естественных пастбищ в засушливые годы не превышает 0.01 т/га, в благоприятные по увлажнению – 0.5 т/га сухой массы.
Рис. 3.
Фрагмент кустарниково-полукустарничковых пастбищ осенне-зимнего использования в пустыне Карнабчуль

Распределение кормовой продукции со второго года по жизненным формам следующее: кустарник (саксаул чёрный) – 22.4%, полукустарнички (кохия простёртая, полынь развесистая) – 67.6%, травы (мятлик луковичный и эфемеры) – 10.0%. Присутствие зонально-типичного виолентного кустарника и ксерогалофильных полукустарничков обеспечивает формирование самовозобновляющихся, высокопродуктивных, экологически устойчивых пастбищных сообществ. Входящие в их состав эфемероиды с характерной для них поверхностно развитой мочковатой корневой системой образуют достаточно плотный дёрн, тем самым обеспечивая улучшение структуры почвы и снижение её эрозии.
Поскольку данные пастбища состоят из растений, поедаемых в разные сезоны года (саксаул чёрный и полынь развесистая – осенью и зимой; мятлик луковичный и эфемеры – весной; кохия простёртая – во все сезоны), они служат прекрасным выпасом осенью и зимой.
Адаптивные методы формирования пастбищ круглогодового использования. Для формирования поливидовых пастбищ круглогодового использования нами применялись следующие доминантные виды кормовых растений: кустарник саксаул чёрный (Haloxylon aphyllum) – 15%, ксерогалофильный полукустарник солянка малолистная (Halothamnus subaphyllus (C.A. Mey.) Botsch.) – 15%, полукустарнички – кохия простёртая (Kochia prostrata) – 25%, солянка восточная (Salsola orientalis S.G. Gmel.) – 25%, полынь развесистая (Artemisia diffusa) – 20%. Посев производили в конце ноября. С целью изучения строения кустарниково-полукустарничково-травянистых пастбищ был заложен вертикальный разрез участка (рис. 4).
Рис. 4.
Вертикальный профиль кустарниково-полукустарничково-травянистых пастбищ круглогодового использования в пустыне Карнабчуль:
1 – саксаул чёрный (Haloxylon aphyllum), 2 – солянка малолистная (Halothamnus subaphyllus), 3 – кохия простёртая (Kochia prostrata) + солянка восточная (Salsola orientalis), 4 – полынь развесистая (Artemisia diffusa), 5 – мятлик луковичный (Poa bulbosa) + осока толстостолбиковая (Carex pachystylis). Длина профиля 6.2 м
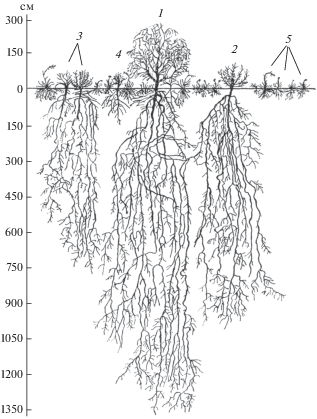
Структура пастбища круглогодового использования состоит из пяти синузий, взаимодополняющих друг друга по эколого-биологическим характеристикам и занимаемым экологическим нишам. Саксаул чёрный, будучи по адаптивной стратегии виолентом, занимает в круглогодовой пастбищной системе доминантную позицию. Средняя высота растений – 320 см, ширина кроны – 250 см. Кроны саксаула чёрного не смыкаются, поэтому надземное пространство на уровне 300–320 см не заполнено. Корневая система растения, мощно развитая и приспособленная для усвоения влаги и элементов минерального питания большого объёма почвогрунта, проникает на глубину более 1300 см. На глубине 150–250 см корни второго порядка делятся на множество корней последующих порядков. На глубинах 150–450 см и ниже, где слои почвы сухие, боковые корни имеют множество поглощающих (всасывающих) корешков. Бетоноподобные плотные слои почвогрунта, встречающиеся на глубине 650–700 см, корни пробивают в утончённом, сплетённом виде. На глубинах 750–900 см корни саксаула чёрного достигают слоёв почвогрунта, увлажняемых грунтовыми водами, поэтому боковые корни обрастают множеством волосовидных.
Вторую синузию образует популяция галоксерофильного кустарника солянки малолистной (Halothamnus subaphyllus). По адаптивной стратегии она совмещает черты виолента и патиента, обладает способностью поддерживать стабильный водный баланс и нормальное состояние оводнённости в экстремально жёстких условиях пустынного климата, что обеспечивает устойчивость к засухе и толерантность к солевому стрессу. Растения имеют среднюю высоту 90 см при ширине кроны 105 см. Кроны не смыкаются, и воздушное пространство на высоте 100–105 см от поверхности земли остаётся незаполненным. В то же время кроны саксаула чёрного, солянки малолистной и полукустарничков (кохия простёртая и солянка восточная), как правило, на разной высоте смыкаются, иногда даже перекрываются, механически не соприкасаясь друг с другом. В подземной сфере корневая система солянки малолистной занимает почвенную нишу толщиной 0–1015 см, она глубоко проникает в почву и довольно разветвлёна. На глубине 60–80 см главный корень делится на несколько корней второго порядка, а они в свою очередь – на множество корней последующих порядков. На глубине 600–750 см наблюдается множество мелких поглощающих корешков. Корни углубляются до зоны увлажнения грунтовыми водами.
Третий ярус полукустарничков – ксерогалофильной кохии простёртой и галоксерофильной солянки восточной (Kochia prostrata + Salsola orientalis). Оба полукустарничка – зонально типичные пустынные виды, кохия простёртая по адаптивной стратегии принадлежит к виолентам, солянка восточная – к патиентам. Стратегический тип растений с патиентными свойствами – это виды, существование которых достигается не энергией жизнедеятельности, а выносливостью к воздействию стрессовых факторов среды. Средняя высота кохии простёртой 65 см, солянки восточной – 70 см, ширина кроны соответственно 55 и 60 см. Подземная часть синузии полукустарничковых растений охватывает довольно обширную экологическую нишу – толщу почвогрунта 0–750 см. Их корневые системы по всему почвенному профилю хорошо разветвлены. Главные корни на глубине 70–120 см распадаются на мелкие корни второго порядка, которые разветвляются на последующие очереди боковых корней с множеством активных поглощающих корешков.
Четвёртая синузия полукустарничков – полынь развесистая (Artemisia diffusa) – в надземной сфере в среднем достигает 36 см высоты при ширине кроны 42 см. Кроны не смыкаются, находясь на расстоянии 75–100 см друг от друга. Подземная часть охватывает небольшую почвенную толщу – 0–100 см. Корневая система хорошо разветвлена по всему почвенному профилю. На глубине 15–20 см главный корень распадается на множество боковых корней второго и последующих порядков, несущих поглощающие корешки.
Пятая синузия травянистых растений – эфемероиды (Carex pachystylis + Poa bulbosa) и эфемеры. Высота надземной части 20–25 см. Подземная слабо развита, корни проникают в слой почвы до 20 см, но основная их масса (до 85%) сосредоточена в слое почвы 0–15 см.
В процессе восстановительной сукцессии многовидовых кустарниково-полукустарничково-травянистых пастбищных сообществ происходит дифференциация и рациональная упаковка экологических ниш путём ярусного взаимодополнения растений. В надземной сфере каждый вид занимает свой ярус в зависимости от максимальной высоты: древовидный кустарник саксаул чёрный достигает 320–400 см, кустарник солянка малолистная – 120–140 см, полукустарнички (кохия простёртая, солянка восточная) – 65–70 см. Это обеспечивает рациональное размещение фотосинтезирующего аппарата растений в воздушном пространстве для эффективного использования солнечной энергии. В подземной сфере растения формируют корневые системы, осваивают различные экологические ниши с целью эффективного использования почвенной влаги и элементов минерального питания.
Наряду с ярусным взаимодополнением в пастбищах круглогодового использования дифференциация происходит и по принципу сезонного взаимодополнения фенологически разноритмичных видов растений: как упоминалось, саксаул чёрный, солянка малолистная, кохия простёртая, солянка восточная длительно вегетирующие (226–242 дня), с конца марта до начала декабря, как правило, до зимних заморозков находятся в зелёном состоянии. А у эфемероидов период вегетации короток: с февраля до середины апреля.
Реставрированные пастбища круглогодового использования, имеющие в своём составе виды кормовых растений различных жизненных форм, характеризуются высокой и устойчивой кормовой производительностью по годам и сезонам (рис. 5). Уже на втором году процесса реставрации она в 2 раза превышает продуктивность природных пастбищ. Максимальной производительности пастбища круглогодового использования достигают на 3–5 годах жизни (3.45–4.39 т/га сухой кормовой массы). В формировании общей кормовой продукции поливидового пастбища значительна роль полукустарничков – кохии простёртой и солянки восточной: первые 2–4 года они способны быстро формировать кормовую массу. А вот у саксаула чёрного созревание происходит значительно медленнее, и на его долю приходится 1/3 часть общей кормовой продукции. Присутствие разносезонных поедаемых полукустарничков в сочетании с кустарником саксаулом чёрным и эфемероидами позволяет использовать поливидовые пастбища на протяжении всего года (рис. 6).
Рис. 5.
Кормовая продуктивность кустарниково-полукустарничково-травянистых пастбищ круглогодового использования в пустыне Карнабчуль:
а – саксаул чёрный (Haloxylon aphyllum), б – солянка малолистная (Halothamnus subaphyllus), в – кохия простёртая (Kochia prostrata), г – солянка восточная (Salsola orientalis), д – полынь развесистая (Artemisia diffusa), е – мятлик луковичный (Poa bulbosa), ж – всего
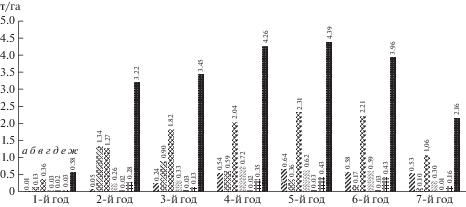
Рис. 6.
Выпас овец на восстановленных кустарниково-полукустарничково-травянистых круглогодовых пастбищах в пустыне Карнабчуль

По жизненным формам кормовая продукция пастбищ распределяется следующим образом: на долю кустарников (саксаул чёрный, солянка малолистная) приходится 28.3%, полукустарничков (кохия простёртая, солянка восточная, полынь развесистая) – 63.6%, трав (мятлик луковичный и эфемеры) – 8.1%. Оценка продуктивности пастбищ на протяжении года показывает: на весенне-летний период приходится 58% получаемого подножного корма, осенне-зимний – 42%. Следует подчеркнуть, что поливидовые пастбища могут служить отличным выпасом для овец во все сезоны года.
٭ ٭ ٭
Воссоздание оптимальных зональных флороценотических структур, введение в их состав базовых доминантных видов и жизненных форм кормовых растений стабильных сообществ – важный способ оптимизации природной среды в центральноазиатской пустыне. В ходе исследований сформированы флористически и ценотически полночленные многоярусные самовозобновляющиеся пастбищные биогеоценозы с характерным набором видов, которые встречаются в естественной эталонной пастбищной растительности. Восстановленные многовидовые, многоярусные кустарниково-полукустарничково-травянистые пастбища осенне-зимнего и круглогодового использования обладают свойством самовозобновления и самоподдержания структурно-функциональной организации. Важно также, что они эффективно выполняют природоохранную функцию в условиях пустынь: приостанавливаются процессы опустынивания аридных земель, прекращаются пыльные бури, восстанавливаются оптимальные параметры окружающей среды. Совокупность позитивных изменений – восстановленное биоразнообразие, оптимизированная экологическая среда, повышенная кормовая производительность некогда опустыненных земель – создают фундаментальные биологические, экологические и научно-организационные предпосылки для устойчивого развития пустынно-пастбищного животноводства как важной составной части жизнеспособного сельского хозяйства центральноазиатского региона.
Список литературы
Нечаева Н.Т., Шамсутдинов З.Ш. Антропогенная динамика пустынных биогеоценозов и пути восстановления их продуктивности // Проблемы антропогенной динамики биогеоценозов / Чтения памяти академика В.Н. Сукачёва / Отв. ред. И.А. Шилов. М.: Наука, 1990. С. 31–53.
Жамбакин Ж.А. Пастбища Казахстана. Технология использования. Алматы: Кайнар, 1995.
Гаевская Л.С. Каракулеводческие пастбища Средней Азии. Ташкент: ФАН, 1971.
Зонн И.С., Куст Г.С., Орловский Н.С., Ши Пей Чжун, Тянь Юй-Чжао. Опустынивание // Пустыни и опустынивание. Энциклопедия. М.: Международные отношения, 2018. С. 421–423.
Novacek M.J., Cleland E.E. The current biodiversity extinction event: Scenarios for mitigation and recovery // Proceedings of the National Academy of Sciences. 2001. V. 98(10). P. 5466–5470.
Harris J.A., Hobbs R.J., Higgs E.S., Aronson J.A. Ecological restoration and climate change // Restoration Ecology. 2006. V. 14(2). P. 170–176.
Керженцев А.С. Функциональная экология. M.: Наука, 2006.
Palmer M.A., Zedler J.B., Falk D.A. Ecological Theory and Restoration Ecology // Foundations of Restoration Ecology / Eds. Falk D.A., Palmer M.A., Zedler J.B. Washington: Island Press. 2016. P. 3–26.
Resolution adopted by the General Assembly on 1 March 2019 73/284. United Nations Decade on Ecosystem Restoration (2021–2030). https://undocs.org/en/A/RES/73/284
The SER International Primer on Ecological Restoration. Society for Ecological Restoration. International Science & Policy Working Group (Version 2: October, 2004). https://www.ctahr.hawaii.edu/littonc/PDFs/ 682_SERPrimer.pdf
Steffen W., Richardson K., Rockström J. et al. Planetary Boundaries: Guiding Human Development on a Changing Planet // Science. 2015. V. 347. Is. 6223. P. 736–747.
Palmer M.A., Ruhl J.B. Aligning Restoration Science and Law to Sustain Ecological Infrastructure for the Future // Frontiers in Ecology and the Environment. 2015. V. 13 (9). P. 512–519.
Suding K.N., Higgs E., Palmer M.A. et al. Seeing the Forest and the Trees: Comprehensive Ecological Restoration // Science. 2015. V. 348. Is. 6235. P. 638–640.
Основы лесной биогеоценологии / Под ред. В.Н. Сукачёва, Н.В. Дылиса. М.: Наука, 1964.
Работнов Т.А. Фитоценология. М.: МГУ, 1983.
Онипченко В.Г. Функциональная фитоценология: синэкология растений. М.: Красанд, 2013.
Миркин Б.М., Наумова Л.Г. Введение в современную науку о растительности. М.: ГЕОС, 2017.
Сукачёв В.Н. Идея развития в фитоценологии // Советская ботаника. 1942. № 1–3. С. 5–17.
Зозулин Г.M. Научные и практические аспекты использования ландшафтов как эталонов природы // Человек и биосфера. Ростов: Изд-во Ростовского университета, 1977. С. 158–167.
Раменский Л.Г. Избранные работы. Проблемы и методы изучения растительного покрова. Л.: Наука, 1971.
Grime J.P. Plants strategies and vegetation processes. New York: John Wiley & Sons, Ltd., 1979.
Grime J.P. Plant strategies, vegetation processes, and ecosystem properties. 2 ed. New York: John Wiley & Sons, Ltd., 2001.
Раменский Л.Г. Введение в комплексное почвенно-геоботаническое исследование. М.: Сельхозгиз, 1938.
Одум Ю. Основы экологии. М.: Мир, 1975.
Hutchinson G.E. Concluding remarks // Cold Spring Harbor Symposia on quantitative Biology. 1957. V. 22. P. 415–427.
Северцов А.С. Соотношение фундаментальной и реализованной экологических ниш // Журнал общей биологии. 2012. № 5. С. 323–333.
Абатуров Б.А. Конкурентное разобщение травяных и древесных растительных сообществ в степной и лесной природных зонах // Успехи современной биологии. 2014. № 5. С. 467–475.
Куркин K.A. Система конструирования луговых травосмесей // Бюл. Моск. об-ва испытателей природы. Отд. Биол. 1983. № 4. С. 3–14.
Миркин Б.М. Теоретические основы современной фитоценологии. М.: Наука, 1985.
Залетаев В.С. Жизнь в пустыне. Географо-биогеоценотические и экологические проблемы. М.: Мысль, 1976.
Дополнительные материалы отсутствуют.