

Вестник РАН, 2022, T. 92, № 9, стр. 869-876
ЭКОЛОГИЧЕСКАЯ ДЕТЕРМИНАЦИЯ ИНДИГЕННЫХ БИФИДОБАКТЕРИЙ КИШЕЧНИКА ЧЕЛОВЕКА
О. В. Бухарин a, *, С. В. Андрющенко a, **, Н. Б. Перунова a, ***, Е. В. Иванова a, ****
a Институт клеточного и внутриклеточного симбиоза УрО РАН
Оренбург, Россия
* E-mail: ofrc@list.ru
** E-mail: rattus000@gmail.com
*** E-mail: perunovanb@gmail.com
**** E-mail: walerewna13@gmail.com
Поступила в редакцию 04.04.2022
После доработки 25.04.2022
Принята к публикации 12.05.2022
- EDN: ZMYWFU
- DOI: 10.31857/S0869587322090055
Аннотация
В обзоре рассматривается экологическая детерминация индигенных (постоянно присутствующих) бифидобактерий толстого кишечника человека. Под экологической детерминацией (от лат. determinere – “определяю”) понимается совокупность природных явлений местообитания (биотопа), определяющих роль индигенных микроорганизмов в микробиоценозе. С использованием симбиотического подхода предпринята попытка выявить экологические условия обитания бифидобактерий и их физиологических эффектов в микросимбиоценозе. Установлены особенности индигенных бифидобактерий по их природе: эволюционно-генетические (филогенетическая удалённость, консервативность генома, метаболическая специализация), биохимические (лизоцимрезистентность, конститутивная ацетатпродукция) и физиологические (микробное распознавание “свой–чужой”, иммунорегуляция), имеющие значение в адаптации (персистенции), обеспечении мутуалистических эффектов и стабильности бифидофлоры в популяции.
Принимая во внимание, что детерминация в классическом понимании основана на объективной причинной обусловленности явлений [1], мы подошли к изучению индигенных бифидобактерий кишечника человека с позиции экологической детерминации. В контексте микробиологии принцип экологической детерминации был выдвинут голландским микробиологом М.В. Бейеринком в начале XX в., а конкретно сформулирован в 1934 г. его соотечественником Л.Б. Бекингом. Исходно он представлялся в виде максимы “всё есть везде”. Затем данный подход был существенно ограничен, и сформировалось понимание, что условия среды обусловливают население биотопа [2], а значит – диапазон их физиологических проявлений. Следствием этого стало существование конечного числа более или менее стабильных зон оптимума адаптации в многомерном пространстве эволюционного ландшафта приспособленности [3]. Подход применим в том числе к микроорганизмам, населяющим пищеварительный тракт [4].
Микросимбиоценоз кишечника по праву относят к одному из наиболее многообразных по составу. На чрезвычайную сложность населяющей человека и животных микрофлоры указывает тот факт, что абсолютное количество бактерий толстого кишечника оказывается сопоставимо с количеством клеток организма хозяина. В последние десятилетия использование современных метагеномных подходов позволило расширить наши представления не только о составе микробиоты, но и о её динамике и экологии, что отражает эволюционную сторону формирования мутуалистических отношений в системе “паразит–хозяин” [5, 6].
Большое биохимическое разнообразие и динамичность содержимого толстого кишечника существенно осложняют поиск наиболее важных зависимостей и взаимосвязей в микробиоценозе, установление влияния тех или иных факторов, определяющих его состав и результат взаимодействия с организмом хозяина [7]. С учётом того, что микробиота продуцирует вещества, оказывающие значительное нейроэндокринное регуляторное действие на организм, кишечная микробиота может считаться полноценным микробным органом, принимающим участие в обеспечении гомеостаза хозяина [8].
Роль индигенных бифидобактерий кишечника человека до сих пор остаётся предметом исследований. Показано, что они выступают одними из немногих надёжных мутуалистов человека, не обладающих патогенными свойствами вне зависимости от состояния организма хозяина [9]. Накоплены фактические материалы, свидетельствующие, что некоторые виды бифидобактерий стабильно обосновались в толстом кишечнике и остаются нашими верными помощниками, участвуя в поддержании гомеостаза [10]. При этом сравнительно небольшая численность бифидобактерий у взрослых лиц и относительно небольшой размер генома не могут служить отражением их функциональной роли в микросимбиоценозе. Таким образом, конкретная экологическая ниша бифидобактерий в микросимбиоценозе кишечника и соответствующие ей главные критерии адаптивности остаются не вполне прояснёнными. В связи с этим мы попытались выявить наиболее значимые характеристики бифидобактерий, определяющие их место в микробиоценозе кишечника человека, и механизмы их персистенции. Признаки были сгруппированы в три категории с учётом имеющегося фактического материала и их природы: эволюционно-генетические, биохимические и физиологические.
Эволюционно-генетические параметры адаптивности бифидобактерий. По современной систематике род Bifidobacterium входит в семейство Bifidobacteriaceae, порядок Bifidobacteriales, подкласс Actinobacteridae, класс Actinobacteria, тип Actinobacteria. Видовая и количественная малочисленность представителей этой таксономической ветви, установленная по результатам микробиомных исследований, указывает на филогенетическую удалённость/изолированность от большинства патогенных и условно-патогенных кишечных бактерий, что затрудняет процесс горизонтального переноса генов патогенности и антибиотикорезистентности между ними [11]. У бифидобактерий к настоящему моменту не выявлено островов патогенности, несмотря на то, что известно несколько случаев их фиксирования (вкупе с иными микроорганизмами) при некоторых инфекционно-воспалительных заболеваниях [10]. Число и разнообразие внутри- и внехромосомных мобильных генетических элементов у бифидобактерий невелико, и особенности их организации мало способствуют их переносу другим микроорганизмам [12, 13].
К настоящему времени опубликовано и описано значительное количество последовательностей геномов бифидобактерий [10, 14]. Тем не менее причины их обитания в кишечнике человека на протяжении всей жизни, их адаптация и выживание в условиях желудочно-кишечного тракта, а также их физиологические эффекты требуют дальнейшего изучения. Бифидобактерии как свободноживущие формы прокариот имеют сравнительно небольшой и консервативный геном, что в свете тенденции к его редукции [15] может отражать устойчивый тренд их эволюции в сторону специализации к конкретному типу биотопов путём фиксации физиологических возможностей. Сравнительный анализ геномов бифидобактерий и типичных представителей облигатно-анаэробного звена кишечного микросимбиоценоза на основании оценки размеров генома и набора генов двухкомпонентных систем позволил нам выполнить следующее ранжирование таксонов: Bifidobacterium spp. (менее 2.5 млн пар нуклеотидов (п.н.), 5–19 белков) < Propionibacterium spp. (2.5–3.5 млн п.н., 6–34 белка) < Prevotella spp. (2.6–3.6 млн п.н., 4–22 белка) < Clostridium spp. (4.1 ± ± 0.06 млн п.н., 14–32 белка) < Peptoclostridium spp. (4.1 ± 0.7 млн п.н., 51 белок) < Bacteroides spp. (5.0–6.26 млн п.н., 50–86 белков) [12].
Подтверждением консервативности генома бифидобактерий служат результаты сравнительного анализа специфичного для Bifidobacterium кластера ортологичных генов – кор-генома, который позволил установить наличие 10 разных филогенетических групп, частично коррелирующих с экологическими нишами. Например, представители групп B. adolescentis (B. catenulatum, B. pseudocatenulatum и B. adolescentis), B. longum (B. breve и B. longum), B. pseudolongum (B. animalis subsp. lactis) и B. bifidum типичны для кишечного тракта человека и коммерчески используются в качестве пробиотических штаммов [16].
Оценка адаптивного потенциала генома микроорганизмов по вычислению абсолютного и относительного показателей “сигнального ценза” – количества генов двухкомпонентных сигнальных систем [17] – показала, что геном бифидобактерий в среднем кодирует больше детерминант сигнальных систем, чем лактобактерии, а по относительной регуляторной эффективности превосходит как лактобактерии, так и бактероиды. Далее мы обратили внимание на известные детерминанты систем межклеточной коммуникации через аутоиндукторы “чувства кворума” и установили, что все секвенированные штаммы несут гены, обеспечивающие оба пути синтеза ключевого предшественника аутоиндуктора-2 – дигидрокси-2,3-пентандиона – как из S-рибозил-L-гомоцистеина (ген luxS), так и из рибулозо-5-фосфата [18]. Гомологов известных детерминант аутоиндукторов семейства гомосеринлактонов, равно как и рецепторов к аутоиндуктору-2, у бифидобактерий не обнаружено. Хотя бифидобактерии образуют медиатор “общебактериального присутствия”, конкретных специфических сигнальных систем межмикробного взаимодействия у них не выявлено. Учитывая исключительное биохимическое разнообразие кишечника как местообитания в сравнении с прочими биотопами тела человека, можно предположить, что данные условия предъявляют особые требования к регуляторно-адаптивному потенциалу генома его обитателей. Из этого напрашивается вывод, что для бифидобактерий характерен выраженный адаптивный потенциал генома [12].
Успехи последних исследований показали, что бифидобактерии эволюционировали совместно со своими хозяевами и многие их физиологические характеристики могут зависеть от места их обитания. Считается, что бифидобактерии претерпели специфическую генетическую и метаболическую адаптацию, чтобы облегчить колонизацию кишечника человека [19]. В частности, анализ геномов бифидобактерий in silico выявил большой арсенал генов, кодирующих ферменты, участвующие в расщеплении сложных углеводов, которые не могут метаболизироваться ферментами хозяина или большинства микроорганизмов кишечной микробиоты [20, 21]. Специфическая генетическая и метаболическая адаптация бифидобактерий обеспечила им колонизацию кишечника человека и определила реализацию их мутуалистических эффектов в качестве индигенного симбионта. Анализ ряда эволюционно-генетических параметров бифидобактерий показал, что свойства их генома отражают достаточно длительную и узкую специализацию прокариот ко вполне определённой экологической нише хозяина – толстому кишечнику.
Биохимические параметры адаптивности бифидобактерий. Высокая трофическая активность вида (как один из признаков экологической детерминации) может быть использована при анализе роли микросимбионтов кишечника, участвующих в процессах пищеварения и обмена веществ в организме хозяина. По генному составу микробиота обладает значительной индивидуальной компонентой, так как две трети генов представлены лишь у 20% людей [22]. Таким образом, можно говорить о том, что хозяин передал часть метаболических функций своей микробиоте [10]. Выживание любого организма в любом биотопе определяется прежде всего приспособлением к наиболее общим физико-химическим факторам, формирующим условия среды конкретного местообитания.
Один из ключевых факторов, определяющих возможность колонизации и персистенции прокариот в биотопах организма хозяина, – их устойчивость к естественному антисептику – лизоциму. Установлено, что у индигенных видов бифидобактерий этот показатель на порядки превышает уровень продукции лизоцима в кишечнике [23]. Устойчивость к нему у бифидобактерий обеспечивается за счёт модификации пептидогликана, а также резистентностью к его неферментативному действию [24, 25]. Высокая устойчивость к лизоциму у бифидобактерий как в кишечнике, так и в материнском молоке выступает фактором отбора индигенных для человека видов Bifidobacterium spp. [26].
Известно, что адгезивная активность бифидобактерий (как один из важных факторов колонизации биотопа) характеризуется вариабельностью (видо- и штаммоспецифичность) как набора, так и структуры отдельных молекулярных детерминант адгезии к клеткам кишечного эпителия и кишечной слизи [27]. Установлено, что только сортаза-зависимые фимбрии (ворсинки) имеются у всех видов индигенных бифидобактерий [28] и оказывают иммуномодулирующий эффект в отношении продукции цитокина ФНОα. Наибольшей вариабельностью обладает ген FimA, кодирующий главную субъединицу пилина в локусе pil2 [29].
Следующий фактор, способствующий колонизации бифидобактерий кишечного биотопа, – наличие у них разнообразных специфически-анаэробных биохимических превращений веществ [30]. Бифидобактерии выявляются главным образом в пристеночной области оксигенируемого кишечника [31], но сохраняют жизнеспособность и метаболическую активность при концентрациях атмосферного кислорода до 15% и более [32] за счёт механизмов антиоксидантной защиты (пероксидаза, NADH-оксидаза, наличие в геноме реуляторного фактора SIR2) [33].
В процессы метаболизма в кишечнике вовлечены самые разнообразные метаболические фенотипы микробиоты, каждый из которых может стать приоритетным в различных биохимических процессах и иметь значение как для организма человека, так и для поддержания микробного сообщества в биотопе [34]. Вместе с тем многие процессы микробной ферментации субстратов в организме человека реализуются комбинированно, так как у микроорганизмов может быть только часть пути метаболизма. Микроорганизмы, объединяясь в ассоциации, методически выполняют метаболические функции на этапе ферментации и утилизации субстратов. Бифидобактерии и бактероиды через “cross-feeding”-взаимодействия последовательно участвуют в расщеплении полисахаридов до моносахаров с образованием конечных субстратов – короткоцепочечных жирных кислот. По сравнению с геномом человека, кодирующим только 17 гликозид-гидролаз, геном бифидобактерий содержит порядка 56 детерминант карбогидраз, ферментирующих олигосахариды до моносахаров [35]. Ацетат, образуемый бифидофлорой, выступает в качестве основного совместного субстрата для производства бутирата и как фактор роста ряда облигатно-анаэробных бактерий [36]. Так, главным конечным метаболитом облигатных анаэробов в кишечнике становится ацетат (анион уксусной кислоты и её растворимых солей) [37].
Образование бифидобактериями ацетата происходит через так называемый “бифидный шунт” фермента фруктозо-6-фосфат-фосфокетолазы (F6PPK). Рост концентрации кислорода в среде также не снижает уровень продукции ацетата. Выявлено, что высокая интенсивность ацетатпродукции может быть обеспечена при наличии у бифидобактерий генетических детерминант мембранного транспорта углеводов [38]. Способность выделять уксусную кислоту у индигенных бифидобактерий носит высококонсервативный, конститутивный характер, являясь итогом их базового пути катаболизма. Известно, что ацетат бифидобактерий, стимулируя противовоспалительную функцию энтероцитов хозяина, способен блокировать всасывание шига-токсина [39] и снижает способность сальмонелл к адгезии и инвазии [40]. В ходе проведённой нами работы показано, что концентрации ацетата, создаваемые индигенными бифидобактериями кишечника (как in vitro, так и in vivo), способны снижать устойчивость к лизоциму нерезидентных для микробиоты человека грампозитивных бактерий, модифицирующих свой пептидогликан путём N-деацетилирования [38]. Таким образом, ацетатпродукция бифидобактерий служит фактором селекции неиндигенной грампозитивной микробиоты. Выявлен механизм персистенции индигенной бифидофлоры через альтернативную модификацию пептидогликана микроорганизмов, где ацетат играет роль ключевого регулятора, определяющего доминантную роль бифидобактерий в кишечном биотопе хозяина, обеспечивая как первичную дискриминацию неиндигенных кишечных ассоциантов через блокирование де-N-ацетилирования их пептидогликана, так и сохранение индигенной грампозитивной микробиоты с О-ацетилированием пептидогликана.
Физиологические эффекты индигенной бифидофлоры в симбиозе. Взаимодействия индигенной бифидофлоры с хозяином и с ассоциативным звеном, попавшим в кишечник, обусловлены ассоциативным симбиозом. Термин “ассоциативный симбиоз”, предложенный профессором МГУ Е.С. Лобаковой, вследствие своей универсальности хорошо прижился в инфекционной симбиологии. Это многокомпонентная система, включающая хозяина в качестве макропартнёра, стабильного доминантного микросимбионта (нормальная, индигенная микрофлора) и минорных ассоциированных микросимбионтов с разнонаправленным действием. Инфекция же – модельная система ассоциативного симбиоза с участием трёх векторов этой композиции: доминанты, ассоцианты, микросимбиоценоз. Первые две группы симбионтов не требуют разъяснения для читателя, а третий термин – микросимбиоценоз – означает “общение” (греч.). Микробы собираются вместе, чтобы “пообщаться” и “принять решение” для определения своей дальнейшей судьбы в биотопе. Не исключено, что отсюда вытекают удивительно верные выводы о возможности бактерий оставаться в биотопе, так как опасности для них нет. В противном случае сигнал будет передан через кишечно-мозговую ось для выработки регулятора гомеостаза – нейрогормона задней доли гипофиза окситоцина – в целях нормализации обстановки [41]. Вот в чём основа сегодняшней сенсации – “микробы управляют нами”. Эти критерии мы использовали в последующей работе в качестве системообразующего фактора микросимбиоценоза (рис. 1).
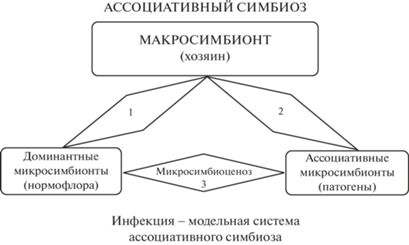
Так что же такое микросимбиоценоз? Это единая динамическая система, состоящая из многовидовых консорциумов, образующих симбиотические связи между собой и макроорганизмом в условиях биокоммуникации с целью создания гомеостаза для жизнедеятельности хозяина и своей собственной. Если признать, что микросимбиоценоз – “пульт управления”, позволяющий регулировать ситуацию в целях сохранения гомеостаза кишечного биотопа, то мы приобретаем нового мощного союзника. Может быть, пора включить применение окситоцина для терапии пациентов с тяжёлой формой и осложнениями новой короновирусной инфекции COVID-19? Возможно, мы просмотрели очередную природоподобную технологию?
Возвращаясь к экологической детерминации индигенных бифидобактерий, приведём ещё один пример очевидной полезности микроорганизмов, непосредственно участвующих в диагностике той микрофлоры, которая находится в кишечном биотопе. Какая она – своя или чужая? На этот вопрос мы постарались ответить с помощью тех же индигенных бифидобактерий с учётом накопленного в работе с ними опыта.
Изучение отношений “паразит–хозяин” позволило сформулировать алгоритм микробного распознавания “свой–чужой” в микросимбиоценозе кишечника человека на основе экспериментально выявленного оппозитного феномена (усиление/подавление) размножения и адаптации микросимбионтов пары “доминант–ассоциант” [6]. Оказавшись фундаментальной находкой, эта пара сразу стала использоваться при отборе бифидобактерий для пробиотических целей. Если к этому добавить, что бифидобактерии, кроме дискриминации чужеродного материала, участвуют в инициальном этапе “сигналинга” – регуляции иммунного гомеостаза хозяина [41], то становится ясно, почему им уделяется так много внимания.
После выявления различий в динамике адаптивного потенциала бифидобактерий таких критериев, как антилизоцимная активность (АЛА), биоплёнкообразование (БПО) и ростовые свойства (РС, КОЕ), стало понятно, что при снижении этих параметров (адаптивного потенциала и РС) исследуемых штаммов вход для них закрыт. Они просто не приживутся в этой среде. Если микробам некомфортно, тормозятся их рост и размножение. Но если штамм “свой” – параметры только растут, отражая благоприятный характер экологической среды и принятие штамма окружением. Этот методический приём с использованием пары “доминант–ассоциант” был нами описан и прошёл испытания в течение ряда лет с положительным результатом.
Метод определения “свой–чужой” получил распространение при отборе перспективных микробных штаммов для создания новых пробиотиков, что и стало основанием для регистрации этих штаммов как в отечественной (Государственная коллекция микроорганизмов нормальной микрофлоры – ГКНМ), так и международной коллекциях. Были предложены новые пробиотические штаммы B. bifidum ICIS-202 (ГКНМ № 1257), B. bifidum ICIS-310 (ГКНМ № 1258), B. bifidum ICIS-643 (ГКНМ № 1259) (депонирование в ГНМК и регистрация в базе данных NСBI BioProject), отмеченные в 2017 г. золотой медалью на Международной выставке-конкурсе “Биоиндустрия” в Санкт-Петербурге (Патенты РФ № 2670054, № 2726653, № 2704423, № 2678123).
Использование антилизоцимной активности и биоплёнкообразования микроорганизмов в качестве биомишени позволило сформировать линейку индикаторных культур микроорганизмов, пригодных для оценки биологической активности новых пробиотиков мишень-направленного действия. Применение данных свойств в связи с новой короновирусной инфекцией COVID-19 в настоящее время очень актуально, поскольку не меньшее значение приобретает проблема биоплёнкообразования микроорганизмов и их роль в развитии воспалительной патологии висцеральных органов и систем организма человека.
В перечень критериев оценки новых пробиотических штаммов была внесена впервые обнаруженная нами антипептидная (антицитокиновая) активность бифидобактерии [10] в отношении про- и противовоспалительных цитокинов – сигнальных медиаторов. С помощью оригинальной методики выявлены перспективные штаммы с выраженным иммунорегуляторным действием, которые могут быть использованы в качестве основы при создании противовоспалительных препаратов нового поколения. Полученные результаты создали предпосылки для введения новой трактовки понимания механизмов реализации воспалительной реакции и начальных этапов адаптивного иммунитета. Установлен прикладной аспект изучения бифидобактерий, которые обладают бивалентным эффектом: с одной стороны, регулируют иммунитет, а с другой – снижают персистентный потенциал патогенов, что можно использовать для создания комбинированных биопрепаратов (синбиотиков, пробиотиков). Это актуально в решении вопросов борьбы с персистирующими и антибиотикорезистентными патогенами.
В других работах противовоспалительные свойства бифидобактерий связывают с наличием у них таких противовоспалительных белков, как ингибитор сериновой протеазы серпин [42], а также внеклеточных макромолекул – экзополисахаридов (EPS), образующих слой гликановой слизи в просвете кишечника [43]. Среди детерминант серин-треониновых протеинкиназ выявлен ген цитокинового рецептора FN3, специфически связывающего фактор некроза опухоли (ФНО) α [44]. Воздействие провоспалительных цитокинов (ФНОα и интерлейкин-6) вызывало у Bifidobacterium longum экспрессию целого ряда генов, продукты которых реализуют противовоспалительный эффект [45]. На интегральном фенотипическом уровне эффект таких механизмов отражает комплексное участие бифидобактерий в регуляции факторов врождённого иммунитета, поддерживая баланс цитокинов и микробицидных веществ в биотопе толстого кишечника человека [10].
Таким образом, можно утверждать, что изучение экологической детерминации значительно расширяет понимание механизмов персистенции в организме человека бесценного помощника – кишечной микробиоты, находящейся на страже его здоровья.
* * *
Рассмотренные характеристики микробиоты представляют совокупность условий, необходимых для экологической детерминации индигенных бифидобактерий в толстом кишечнике человека. Эволюционная надёжность микроорганизмов как симбионтов обеспечивается их генетическими особенностями, которые у бифидобактерий кишечника характеризуются филогенетической удалённостью, консервативностью генома и метаболической специализацией. Филогенетическая удалённость от большинства кишечных бактерий затрудняет процесс приобретения бифидобактериями факторов патогенности и антибиотикорезистентности. Консервативность генома и малое число генов сигнальных систем – свидетельство способности бифидофлоры к более простому адаптивному поведению в микросимбиоценозе и специализации данных микроорганизмов к занимаемому биотопу. Наряду с этим перечисленные выше параметры бифидобактерий штаммоспецифичны и служат индивидуальным маркером штамма (“fingerprint”), внося вклад в понимание адаптивной стратегии прокариот при ассоциативном симбиозе с человеком.
Непосредственное закрепление бактерии в биотопе организма хозяина реализуется, как правило, при наличии соответствующих факторов персистенции. У поверхности слизистой оболочки кишечника бактерии должны обладать выраженной способностью к адгезии и быть устойчивыми как минимум к лизоциму – универсальному природному антисептику. Это объясняет, почему бифидобактерии проявляют выраженную лизоцимрезистентность, а также демонстрируют способность к специфической адгезии к компонентам слизи и поверхности клеток кишечного эпителия. Не исключено, что дополнительным эволюционным свидетельством и фактором такой стабильности выступает биохимическая специализация бифидобактерий, выраженная в способности выделять уксусную кислоту, которая носит конститутивный характер, являясь продуктом их базового пути катаболизма.
Эволюционно длительному сосуществованию симбионтов способствует наличие у них уникальных свойств, имеющих ценность для хозяина, то есть особых мутуалистических эффектов. Понимание роли индигенной бифидофлоры при взаимодействии с хозяином и микросимбиоценозом иллюстрирует ассоциативный симбиоз. Установлено, что микробиота обеспечивает первичный отбор микросимбионтов, и делают это доминантные микроорганизмы – бифидобактерии. Первичная дискриминация чужеродного материала бифидобактериями – инициальный этап сигналинга в регуляции иммунного гомеостаза хозяина. Дальнейшие этапы регуляции осуществляются активацией дендритных клеток непосредственно бифидобактериями и их метаболитами с последующим влиянием на дифференцировку наивных CD4+ T-лимфоцитов в сторону регуляторных лимфоцитов и поддержанием оптимального цитокинового баланса кишечного биотопа человека. Перечисленные конститутивные признаки индигенных видов бифидобактерий детерминируют их в качестве мутуалистически надёжного посредника в микросимбиоценозе кишечника и регулятора гомеостаза (здоровья) хозяина.
Представленный материал расширяет наши знания об условиях выживания и механизмах персистенции индигенных бифидобактерий кишечника человека, раскрывая их экологические особенности и новые физиологические эффекты в организме, что способствует пониманию патогенетической роли бифидофлоры. Практическое значение имеют и выявленные в процессе работы уникальные штаммы бифидобактерий, депонированные в отечественных и международных коллекциях и пригодные для терапевтических целей в качестве эффективных про- и симбиотиков.
Список литературы
Комлев Н.Г. Словарь иностранных слов. М.: Эксмо, 2006.
O’Malley M.A. Everything is everywhere: but the environment selects: ubiquitous distribution and ecological determinism in microbial biogeography // Stud. Hist. Philos. Biol. Biomed. Sci. 2008. V. 39 (3). P. 314–325.
Gavrilets S. Fitness Landscapes and the Origin of Species (MPB-41). Princeton: Princeton University Press, 2004. http://www.jstor.org/stable/j.ctv39x541
Scanlan P.D. Microbial evolution and ecological opportunity in the gut environment // Proc. Biol. Sci. 2019. V. 286 (1915). P. 20191964.
Eloe-Fadrosh E.A., Rasko D.A. The human microbiome: from symbiosis to pathogenesis // Annu. Rev. Med. 2013. V. 64. P. 145–163.
Бухарин О.В., Перунова Н.Б. Микросимбиоценоз. Екатеринбург: Изд-во УрО РАН, 2014.
Yatsunenko T., Rey F.E., Manary M.J. et al. Human gut microbiome viewed across age and geography // Nature. 2012. V. 486 (7402). P. 222–227.
Baquero F., Nombela C. The microbiome as a human organ // Clin. Microbiol. Infect. 2012. V. 4. P. 2–4.
van Reenen C.A., Dicks L.M.T. Horizontal gene transfer amongst probiotic lactic acid bacteria and other intestinal microbiota: what are the possibilities? A review // Arch. Microbiol. 2011. V. 193 (3). P. 157–168.
Бухарин О.В., Перунова Н.Б., Иванова Е.В. Бифидофлора при ассоциативном симбиозе человека. Екатеринбург: Изд-во УрО РАН, 2014.
Hu Y., Yang X., Li J. et al. The Bacterial Mobile Resistome Transfer Network Connecting the Animal and Human Microbiomes // Appl. Environ. Microbiol. 2016. V. 82 (22). P. 6672–6681.
Андрющенко С.В., Иванова Е.В., Перунова Н.Б. и др. Генетическая характеристика адаптивного потенциала бифидобактерий биотопа дистального отдела кишечника человека // Журнал микробиологии, эпидемиологии и иммунобиологии. 2018. № 4. С. 4–11.
Mancino W., Lugli G.A., Sinderen D.V. et al. Mobilome and Resistome Reconstruction from Genomes Belonging to Members of the Bifidobacterium Genus // Microorganisms. 2019. V. 7 (12). P. 638.
Bukharin O.V., Andryuschenko S.V., Perunova N.B. et al. Genome sequence data announcement of Bifidobacterium bifidum strain ICIS-202 isolated from a healthy human intestine stimulating active nitrogen oxide production in macrophages // Data Brief. 2019. V. 27. P. 104761.
Wolf Y.I., Koonin E.V. Genome reduction as the dominant mode of evolution // Bioessays. 2013. V. 9. P. 829–837.
Wong C.B., Odamaki T., Xiao J.Z. Insights into the reason of Human-Residential Bifidobacteria (HRB) being the natural inhabitants of the human gut and their potential health-promoting benefits // FEMS Microbiol. Rev. 2020. V. 44. P. 369–385.
Galperin M.Y., Higdon R., Kolker E. Interplay of heritage and habitat in the distribution of bacterial signal transduction systems // Mol. BioSyst. 2010. V. 6 (4). P. 721–728.
Tavender T.J., Halliday N.M., Hardie K.R., Winzer K. LuxS-independent formation of AI-2 from ribulose-5-phosphate // BMC Microbiol. 2008. V. 8. P. 98.
Rodriguez C.I., Martiny J.B.H. Evolutionary relationships among bifidobacteria and their hosts and environments // BMC Genomics. 2020. V. 21 (1). P. 26.
Duranti S., Longhi G., Ventura M. et al. Exploring the Ecology of Bifidobacteria and Their Genetic Adaptation to the Mammalian Gut // Microorganisms. 2020. V. 9 (1). P. 8.
Sims I.M., Tannock G.W. Galacto- and Fructo-oligosaccharides Utilized for Growth by Cocultures of Bifidobacterial Species Characteristic of the Infant Gut // Appl. Environ. Microbiol. 2020. V. 86 (11). e00214-20.
Tyakht A.V., Kostryukova E.S., Popenko A.S. et al. Human gut microbiota community structures in urban and rural populations in Russia // Nat. Commun. 2013. V. 4. P. 2469.
Rada V., Splichal I., Rockova S. et al. Susceptibility of bifidobacteria to lysozyme as a possible selection criterion for probiotic bifidobacterial strains // Biotechnol. Lett. 2010. V. 32 (3). P. 451–455.
Андрющенко С.В., Перунова Н.Б., Бухарин О.В. Молекулярные механизмы взаимодействия бактерий с лизоцимом и их роль в микросимбиоценозе // Успехи современной биологии. 2015. Т. 135 (5). С. 453–463.
Sakurai T., Hashikura N., Minami J. et al. Tolerance mechanisms of human-residential bifidobacteria against lysozyme // Anaerobe. 2017. V. 47. P. 104–110.
Minami J., Odamaki T., Hashikura N. et al. Lysozyme in breast milk is a selection factor for bifidobacterial colonisation in the infant intestine // Benef. Microbes. 2016. V. 7 (1). P. 53–60.
Martín R., Bottacini F., Egan M. et al. The Infant-Derived Bifidobacterium bifidum Strain CNCM I-4319 Strengthens Gut Functionality // Microorganisms. 2020. V. 8 (9). P. 1313.
Westermann C., Gleinser M., Corr S.C., Riedel C.U. A Critical Evaluation of Bifidobacterial Adhesion to the Host Tissue // Front Microbiol. 2016. V. 7. P. 1220.
Turroni F., Serafini F., Foroni E. et al. Role of sortase-dependent pili of Bifidobacterium bifidum PRL2010 in modulating bacterium-host interactions // Proc. Natl. Acad. Sci. USA. 2013. V. 110 (27). P. 11151–11156.
Takemoto K., Yoshitake I. Limited influence of oxygen on the evolution of chemical diversity in metabolic networks // Metabolites. 2013. V. 3 (4). P. 979–992.
Friedman E.S., Bittinger K., Esipova T.V. et al. Microbes vs. chemistry in the origin of the anaerobic gut lumen // Proc. Natl. Acad. Sci. USA. 2018. V. 115 (16). P. 4170–4175.
Talwalkar A., Kailasapathy K. Metabolic and biochemi-cal responses of probiotic bacteria to oxygen // J. Dairy Sci. 2003. V. 86 (8). P. 2537–2546.
Feng T., Wang J. Oxidative stress tolerance and antioxidant capacity of lactic acid bacteria as probiotic: a systematic review // Gut Microbes. 2020. V. 12 (1). P. 1801944.
Heinken A., Thiele I. Systems biology of host-microbe metabolomics // Wiley Interdiscip. Rev. Syst. Biol. Med. 2015. V. 7 (4). P. 195–219.
Milani C., Lugli G.A., Duranti S. et al. Bifidobacteria exhibit social behavior through carbohydrate resource sharing in the gut // Sci. Rep. 2015. V. 5. P. 15782.
Rivière A., Selak M., Lantin D. et al. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut // Front Microbiol. 2016. V. 7. P. 979.
Rios-Covian D., Cuesta I., Alvarez-Buylla J.R. et al. Bacteroides fragilis metabolises exopolysaccharides produced by bifidobacteria // BMC Microbiol. 2016. V. 16 (1). P. 150.
Бухарин О.В., Андрющенко С.В., Перунова Н.Б., Иванова Е.В. Механизм персистенции индигенных бифидобактерий под действием ацетата в кишечном биотопе человека // Журнал микробиологии, эпидемиологии и иммунобиологии. 2021. № 3. С. 276–282.
Fukuda S., Toh H., Taylor T.D. et al. Acetate-producing bifidobacteria protect the host from enteropathogenic infection via carbohydrate transporters // Gut Microbes. 2012. V. 3 (5). P. 449–454.
Sun Y., O’Riordan M.X. Regulation of bacterial pathogenesis by intestinal short-chain Fatty acids // Adv. Appl. Microbiol. 2013. V. 85. P. 93–118.
Бухарин О.В., Стадников А.А., Перунова Н.Б. Роль окситоцина и микробиоты в регуляции взаимодействий про- и эукариот при инфекции. Екатеринбург: Изд-во УрО РАН, 2019.
Alvarez-Martin P., Fernández M., O’Connell-Motherway M. et al. A conserved two-component signal transduction system controls the response to phosphate starvation in Bifidobacterium breve UCC2003 // Appl. Environ. Microbiol. 2012. V. 78 (15). P. 5258–5269.
Ferrario C., Milani C., Mancabelli L. et al. Modulation of the eps-ome transcription of bifidobacteria through simulation of human intestinal environment // FEMS Microbiol. Ecol. 2016. V. 92 (4). P. fiw056.
Dyakov I.N., Mavletova D.A., Chernyshova I.N. et al. FN3 protein fragment containing two type III fibronectin domains from B. longum GT15 binds to human tumor necrosis factor alpha in vitro // Anaerobe. 2020. V. 65. P. 102247.
Veselovsky V.A., Dyachkova M.S., Menyaylo E.A. et al. Gene Networks Underlying the Resistance of Bifidobacterium longum to Inflammatory Factors // Front Immunol. 2020. V. 11. P. 595877.
Дополнительные материалы отсутствуют.