

Водные ресурсы, 2020, T. 47, № 3, стр. 302-311
Система оценки и прогнозирования биопродуктивности экосистем озерного типа
В. В. Бульон *
Зоологический институт РАН
199034 Санкт-Петербург, Россия
* E-mail: vboulion@mail.ru
Поступила в редакцию 27.02.2019
После доработки 24.09.2019
Принята к публикации 12.11.2019
Аннотация
Разработана система диагностики биологической продуктивности озерных экосистем с входными параметрами: географическая широта, удельный водосбор, среднегодовая температура воздуха, годовая сумма осадков, средняя и максимальная глубина водоема, время пребывания воды в нем, средние за вегетационный сезон значения цветности воды и суточного ассимиляционного числа хлорофилла “a” фитопланктона. На примере некоторых озер и водохранилищ РФ, расположенных в разных географических зонах (от Северо-запада до Дальнего Востока), прогнозируется годовая продукция наземной растительности на водосборной площади и ключевых групп гидробионтов: первичных продуцентов, бактериопланктона и консументов, включая рыб.
ВВЕДЕНИЕ
В условиях дефицита сведений об экологическом состоянии внутренних водоемов разрабатываются все более новые способы их диагностики [15, 20, 22, 23, 40, 47, 50]. Диагностика водоемов, функционирующих в природных условиях или находящихся под антропогенным воздействием, основывается на системе наблюдений, оценки и прогноза ряда гидробиологических показателей. Система диагностики (СД) часто совмещается с моделированием [15, 23], позволяющим реконструировать по имеющимся материалам недостающие биолого-продукционные характеристики водных объектов. Система базируется на знании закономерных связей между биотическими и абиотическими элементами водной среды. Она представляет собой сжатую форму балансовой модели, имитирующей биотический поток энергии в экосистеме, с той лишь разницей, что модель описывает все этапы трансформации вещества и энергии в этом потоке, а в СД на первом плане – входные и целевые характеристики экосистемы, а связывающая их промежуточная информация дана в “спрессованном” виде.
Цель данной работы – на примере некоторых озер и водохранилищ РФ, находящихся в разных климатических зонах, продемонстрировать процедуру диагностики биологической продуктивности водных экосистем, индикаторы которой – годовая продукция ключевых групп гидробионтов: первичных продуцентов, редуцентов и консументов разног порядка, включая рыб.
МАТЕРИАЛ И МЕТОДЫ
Система диагностики биологической продуктивности экосистем озерного типа (рис. 1) основана на балансовой модели биотического потока энергии в водном объекте, находящемся под воздействием окружающей среды – суши и атмосферы [4]. Взаимоотношения между организмами разных трофических уровней представлены в СД связывающими эти уровни переходными коэффициентами и комбинированными уравнениями. Исходные параметры, требуемые для диагностики, легко определяемы либо доступны из справочных источников. В число исходных параметров СД не включено содержание общего фосфора TP в воде, так как его определение технологически трудоемко и затратно. Поток фосфора в водную среду с водосборного бассейна и с атмосферными осадками рассчитывается с использованием доступной информации о среднегодовой температуре воздуха Tem и годовой сумме осадков Pre. В процессе диагностики учитывается то, что не только продукция фитопланктона, но и аллохтонное растворенное органическое вещество (РОВ) – важный источник энергии для гетеротрофных микроорганизмов. Показателем содержания аллохтонного РОВ в экосистеме озерного типа выбрана цветность воды Pt – легко определяемый гидрохимический показатель.
Рис. 1.
Диагностика продукции гидробионтов в экосистемах озерного типа (на примере Рыбинского водохранилища). 1 – входные, 2 – промежуточные, 3 – целевые параметры. Обозначения и единицы измерения в тексте.
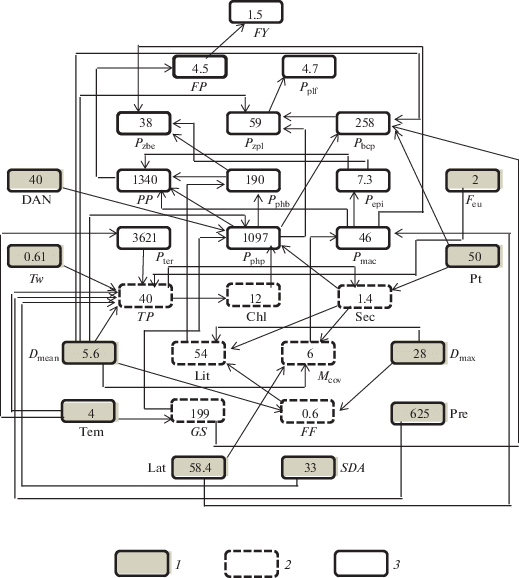
СД прогнозирует следующие показатели трофического статуса водоема в текущих и изменяющихся условиях среды: продукцию фитопланктона Pphp, макрофитов Pmac, микрофитобентоса Pphb, эпифитов Pepi, зоопланктона Pzpl, зообентоса Pzbe, рыб FP и допустимый (без подрыва популяции) вылов рыб FY. Последние два показателя используются для разработки способов рациональной эксплуатации биологических ресурсов водоемов.
Для испытания СД выбраны четыре водохранилища и три озера, расположенных в разных климатических зонах на территории РФ.
Богучанское водохранилище – четвертое (после Иркутского, Братского и Усть-Илимского водохранилищ) в Ангарском каскаде. Его наполнение происходило с 2012 по 2015 г. Площадь зеркала Богучанского водохранилища – 2326, водосборного бассейна – 831 000 км2. Водохранилище расположено на территории Красноярского края и Иркутской области в зоне тайги юго-западной части Средне-Сибирского плоскогорья. Из-за большой удаленности от морей и океанов климат здесь резко континентальный. Зимы холодные, малооблачные, безветренные; лето – в среднем теплое, но короткое. Все водохранилища Ангарского каскада – объекты рыбного хозяйства [28, 29].
Зейское водохранилище заполнялось с 1974 по 1985 г. Оно строилось с целью не только получения электроэнергии, но и для уменьшения ущерба от наводнений, которые часто происходили в пойме рек Зеи и Амура. Водохранилище расположено на территории в границах Амурской области в верхнем течении р. Зеи. Площадь зеркала – 2419, водосборного бассейна – 83 800 км2. Зейское водохранилище по величине — на третьем месте в России и на четвертом в мире (после Красноярского, Братского и Асуанского). Климат в районе водохранилища резко континентальный. Зима продолжительная и морозная. Среднегодовая температура воздуха отрицательная. Водохранилище расположено в зоне вечной мерзлоты [10].
Бурейское водохранилище (Амурское море) на р. Бурее (левый приток Амура) расположено на территории Амурской области и Хабаровского края. Его наполнение началось в апреле 2003 г. и завершилось летом 2009 г. Площадь зеркала – 750 км2 при площади водосборного бассейна 70 700 км2. В зоне водохранилища муссонный климат имеет некоторые черты резко континентального климата. Зимы здесь суровые, малоснежные. Только июль можно назвать летним месяцем [9].
Озеро Ханка расположено на границе Приморского края (Россия) и провинции Хэйлунцзян (Китай). Это самый крупный пресноводный водоем на Дальнем Востоке. Площадь его зеркала 3030, водосбора 18 400 км2. В озеро впадают 24 реки, вытекает р. Сунгача – приток Уссури (бассейн Амура). На озере организован российско-китайский Ханкайский заповедник. Дно озера устлано лёссовым грунтом, что в условиях мелководности озера и частых ветров является причиной высокой мутности воды. Климат здесь имеет ярко выраженный муссонный характер. Зимы холодные, бесснежные, солнечные. Летом ветер с востока и юго-востока приносит влажный воздух и обильные дожди [32].
Онежское – второе по величине (после Ладожского) озеро Европы. В него впадают 52 реки длиной >10 км и порядка тысячи малых речек и ручьев. Главные притоки – Водла, Шуя и Суна. Вытекает из озера р. Свирь, впадающая в Ладожское озеро. Площадь зеркала Онежского озера 9720 км2. Регион, где находится озеро, относится к атлантико-арктической зоне умеренного пояса. Климат характеризуется как переходный от морского к континентальному [27, 36, 43].
Ладожское озеро – крупнейшее в Европе, расположено в Республике Карелии и Ленинградской области. Водосборная площадь Ладожского озера включает в себя озера Онежское, Ильмень и Сайму. Сток воды происходит через р. Неву, соединяющую озеро с Невской губой Финского залива. Площадь зеркала Ладожского озера 17 870 км2. Климат, как и в районе Онежского озера, колеблется от морского до континентального [31].
Рыбинское водохранилище образовано в конце 1930-х–начале 1940-х гг. и охватывает части территорий Тверской и Вологодской областей. Площадь зеркала водохранилища 4580 км2. По размерам оно второе среди водохранилищ на Волге (после Куйбышевского) [38, 54].
Основные географические, морфометрические и гидрохимические показатели озер приведены в табл. 1.
Таблица 1.
Средние значения входных, промежуточных и целевых параметров для прогнозирования биологической продуктивности некоторых озер и водохранилищ РФ
| Параметры | Богучанское вдхр | Зейское вдхр |
Бурейское вдхр |
оз. Ханка | Рыбинское вдхр | Онежское озеро | Ладожское озеро |
|---|---|---|---|---|---|---|---|
| Входные | |||||||
| Lat, °с.ш. | 58.4 | 54.5 | 50.3 | 45 | 58.4 | 61.5 | 60.6 |
| SDA | 357 | 35 | 94 | 6.1 | 33 | 5.82 | 14.5 |
| Tem, °C | –3.5 | –3.1 | –3.5 | 2 | 4 | 3 | 3.7 |
| Pre, мм/год | 425 | 525 | 690 | 565 | 625 | 630 | 610 |
| Dmean, м | 25 | 38 | 14.5 | 2.5 | 5.6 | 30 | 47 |
| Dmax, м | 75 | 97 | 58 | 6.5 | 28 | 127 | 230 |
| Tw, годы | 0.55 | 4 | 0.38 | 10 | 0.61 | 15.6 | 11.7 |
| Pt, град. | 30 | 75 | 75 | 40 | 50 | 20 | 23 |
| DAN, мкг C/(мг сут) | 40 | 40 | 40 | 70 | 40 | 30 | 40 |
| Feu | 1.5 | 2 | 2 | 3.5 | 2 | 1 | 1.5 |
| Промежуточные | |||||||
| GS, дни | 134 | 137 | 134 | 181 | 199 | 190 | 196 |
| FF, отн. величина | 1 | 1.2 | 0.75 | 1.2 | 0.6 | 0.71 | 0.61 |
| Lit, % | 17 | 9.5 | 34 | 20 | 54 | 30 | 18 |
| Mcov, % | 1.8 | 2.7 | 7.8 | 6.6 | 6 | 2.6 | 1.2 |
| TP, мкг/л | 26 | 14 | 11 | 27 | 40 | 9 | 21 |
| Chl, мкг/л | 7 | 3.1 | 2.3 | 7.6 | 12 | 1.7 | 5.3 |
| Sec, м | 2 | 1.9 | 2 | 0.29 | 1.4 | 3.3 | 2.3 |
| Целевые, ккал/(м2 год) | |||||||
| Pter | 1805 | 1879 | 1805 | 3049 | 3621 | 3327 | 3531 |
| Pphp | 586 | 248 | 199 | 224 | 1097 | 253 | 762 |
| Pmac | 13 | 28 | 115 | 131 | 46 | 13 | 6 |
| Pepi | 2 | 4.4 | 18 | 21 | 7.3 | 2.1 | 1 |
| Pphb | 32 | 7.3 | 21 | 14 | 190 | 24 | 44 |
| PP | 633 | 288 | 354 | 389 | 1340 | 293 | 812 |
| Pbcp | 186 | 318 | 140 | 57 | 258 | 129 | 298 |
| Pzpl | 39 | 39 | 20 | 11 | 58 | 21 | 56 |
| Pzbe | 6.5 | 4.4 | 16 | 21 | 38 | 4.8 | 6.6 |
| Pplf | 3.1 | 3.1 | 1.6 | 0.87 | 4.7 | 1.7 | 4.5 |
| FP | 2.1 | 2 | 1.7 | 1.5 | 4.5 | 1.2 | 2.9 |
| FY | 0.7 | 0.67 | 0.57 | 0.5 | 1.5 | 0.4 | 1 |
Для диагностики биологической продуктивности перечисленных водоемов используются следующие входные параметры – независимые переменные (табл. 1): Lat – географическая широта, °с.ш.; SDA – удельный водосбор; Tem – среднегодовая температура воздуха, °С; Pre – годовая сумма осадков, мм/год; Dmean и Dmax – средняя и максимальная глубины водоема, м; Tw – время пребывания воды в водоеме, годы; Pt – цветность воды в водоеме, град.; DAN – суточное ассимиляционное число хлорофилла a фитопланктона; Feu – фактор эвтрофирования водоема, при отсутствии антропогенной нагрузки Feu = 1.
Промежуточные параметры – следующие ниже зависимые переменные.
Pter = 4 (3000/(1 + EXP(1.315 – 0.119 Tem))) – продукция наземной растительности на водосборе, ккал/(м2 год) [21, 46]. Параметр Pter используется как промежуточный для расчета TP, но он также целевой, характеризующий продуктивность системы водосбор–озеро.
FF = 3Dmean/Dmax – фактор формы котловины водоема.
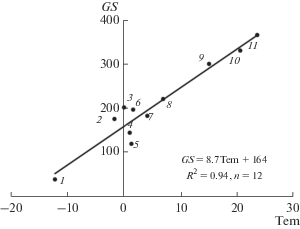
GS = 8.71 Tem + 164 – длительность вегетационного сезона, дни (рис. 2).
Рис. 2.
Зависимость длительности вегетационного сезона (GS, дни) от среднегодовой температуры воздуха (Tem, °C). Озеро на о. Хейса в архипелаге Земля Франца-Иосифа (1), Зейское (2) и Бурейское (3) водохранилища, озера Кривое в Северной Карелии (4) и Зеленецкое на Кольском п-ве (5), Богучанское водохранилище (6), озера Красное на Карельском перешейке (7), Нарочь в Беларуси (8), Инкити в Грузии (9), Киннерет в Израиле (10), Джордж в Уганде и Кодаи в Индии (11). По [9, 10, 25, 26, 51, 53].
TP = 1000 ((SDA × 0.1 × 0.002 × 0.02 Pter × × 2(Tem‑-12)/10 × Feu + 0.00003 Pre) (1 – R) Tw)/Dmean – содержание общего фосфора в воде, мкг/л. Коэффициенты: 0.1 – отношение массы органического углерода к ее калорийности; 0.002 – соотношение P : C в растительном материале [41]; 0.02 – средняя для планеты доля фосфора, выносимого в водоемы из наземных экосистем с природным ландшафтом [18]; 0.00003 – среднее содержание TP в атмосферных осадках, г/л [33]; R = 5/(5 + Dmean/Tw) – коэффициент седиментации фосфора [45].
Sec = 10 ^(1.26 – 0.31 × LOG10 (Pt) – 0.36 × × LOG10 (TP)) – прозрачность воды в водоеме, м [48].
Lit = (1 – ((Dmax – 2 Sec)/(Dmax + 2 Sec × × EXP(3 – FF 1.5))) 0.5/FF)100 – отношение площади литоральной зоны к площади водоема, % [45].
Mcov = IF (Lat < 55) THEN (56.5 (Sec/Dmean)) ELSE (23.6 (Sec/Dmean)) – степень зарастания водоема макрофитами, % [5]. В этом логическом уравнении зафиксировано допущение, что на высоких широтах степень зарастания ниже, чем на низких широтах, из-за смены мягких грунтов на каменистые субстраты.
Chl = 0.103 × TP 1.29 – содержание хлорофилла а в планктоне, мкг/л [6].
Целевые параметры – следующие ниже зависимые переменные.
Pphp = 0.8 (IF (2 Sec > Dmean) THEN (DAN × Chl × × Dmean (1 – Dmean/(4 Sec))GS/100) ELSE (DAN × × Chl × Sec × GS/100)) – продукция фитопланктона. Это логическое уравнение учитывает тот факт, что в отдельных водоемах потенциальная глубина фотического слоя может превышать среднюю глубину [45].
Pmac = 10 ^ (2.21+ 1.08 LOG10 (Mcov) – 0.49 × × (90/(90-Lat))) – продукция макрофитов, ккал/(м2 год) [5].
Pepi = 0.16Pmac – продукция эпифитов, ккал/(м2 год) [5].
Pphb = (0.32 Pphp × Lit)/100 – продукция микрофитобентоса, ккал/(м2 год) [5].
PP = Pmac + Pepi + Pphb + Pphp – суммарная первичная продукция, ккал/(м2 год).
Pbcp = 0.2 Pphp + 10 (Pt/12) 0.0025 × 0.33 Dmean × × GS – продукция бактериопланктона с учетом потребленного им аллохтонного РОВ, ккал/ (м2 год). Коэффициент 0.2 означает, что ≈20% продукции фитопланктона трансформируется в бактериальную продукцию; 10 – калорийность органического углерода, ккал/г; Pt / 12 – концентрация аллохтонного РОВ, мг C/л; 0.0025 – константа скорости утилизации аллохтонного РОВ бактериопланктоном, сут–1 [3]; 0.33 – эффективность роста бактериопланктона [14].
Pzpl = 0.59 × 0.16 (Pphp (1 – 0.2/0.33 – 0.1/ (0.3Dmean)) + Pbcp) – продукция зоопланктона, ккал/(м2год); коэффициент 0.59 – отношение продукции планктонного сообщества к продукции нехищного зоопланктона; 0.16 – эффективность роста нехищного зоопланктона [12]; 0.2 – доля чистой продукции фитопланктона, трансформированная в бактериальную продукцию; 0.33 – эффективность роста бактериопланктона; 0.1 – скорость седиментации фитопланктона, м/сут [11]; 0.3 – скорость оборота биомассы фитопланктона, сут–1 [17].
Pzbe = 0.16(0.6Pmac + 0.73Pepi + 0.73Pphb + + 0.1Pphp/(0.3Dmean)) – продукция зообентоса, ккал/(м2 год), где коэффициенты 0.16 – эффективность роста зообентоса [1]; 0.6 и 0.73 – доля продукции макрофитов, эпифитов и микрофитобентоса, потребленная зообентосом, с поправкой на ассимиляцию продукции этих автотрофов бактериобентосом; 0.1 – скорость седиментации фитопланктона, м/сут; 0.3 – скорость оборота биомассы фитопланктона, сут –1. Формула рассчитана на основе модельного алгоритма [4].
Pplf = 0.08Pzpl – продукция планктоноядных рыб, ккал/(м2 год) [4].
FP = 0.16 (0.5 Pplf + 0.5 × 0.08 Pzbe) + 0.5 Pplf + + 0.5 × 0.08 Pzbe – продукция рыбного сообщества, ккал/(м2 год), где коэффициенты 0.16 – эффективность роста хищных рыб [42]; 0.5 – доля продукции планктоноядных и бентосоядных рыб, потребленная хищными рыбами, а также утраченная в результате вылова или естественной смертности; 0.08 – эффективность роса бентосоядных рыб [42].
FY = FP/3 – вылов рыб без ущерба популяции, ккал/(м2 год) [4].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Значения входных параметров и спрогнозированных на их основе промежуточных и целевых параметров биологической продуктивности водоемов представлены в табл. 1. Для расчета годовой величины продукции фитопланктона важно знать продолжительность вегетационного сезона GS. Ранее предлагалось рассматривать GS как функцию географической широты Lat [44, 45]. Однако эту функцию нельзя считать универсальной для всех географических зон, так как она игнорирует подчиненность GS меридиональной зональности. Проанализировав данные некоторых сайтов и литературных источников, автор пришел к выводу, что значения GS тесно связаны со среднегодовой температурой воздуха Tem (рис. 2).
Верификация показаний СД проводилась по данным гидробиологических исследований Онежского озера (рис. 3) и Рыбинского водохранилища (рис. 4). Установлено сходство прогнозируемых величин продукции гидробионтов с результатами натурных наблюдений.
Рис. 3.
Сравнение результатов прогноза с эмпирическими данными [27, 30, 35, 37] для Онежского озера: 1 – продукция фитопланктона; 2 – бактериопланктона; 3 – макрофитов; 4 – зоопланктона; 5 – зообентоса; 6 – вылов рыбы, ккал/(м2 год). Сплошная линия – линия регрессии, пунктирная – линия симметрии (x = y).
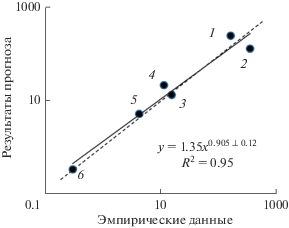
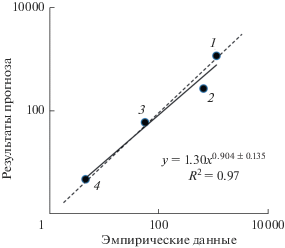
Основным маркером трофического статуса водоемов принято считать содержание Chl в планктоне. СД прогнозирует содержание Chl для экосистемы, функционирующей в условиях, близких к природным. Если содержание Chl, установленное эмпирическим путем, превышает прогнозируемую величину, это свидетельствует об эвтрофировании водоема. В этом случае подбирается соответствующая поправка (в виде коэффициента эвтрофирования Feu) в уравнение, описывающее зависимость TP от Pter и Pre, чтобы приблизить прогнозируемое значение Chl к эмпирическому. Так как в СД входные, промежуточные и целевые параметры взаимосвязаны, корректировка Chl приведет к изменению всех остальных показателей трофического состояния экосистемы.
СД позволяет ретроспективно прогнозировать трофическое состояние водоема, если бы он находился внутри водосбора с природным ландшафтом и не подвергался бы антропогенной биогенной нагрузке. Приняв Feu = 1, находим, что продукция фитопланктона в отсутствие эвтрофирования составляла бы в рассматриваемых здесь водоемах (кроме Онежского озера, практически не подвергающегося эвтрофированию) от 90 (оз. Ханка) до 590 ккал/(м2 год) (Рыбинское водохранилище), что в 1.4–2 раза меньше, чем на текущем этапе функционирования экосистем (табл. 2).
Таблица 2.
Продукция фитопланктона и суммарная первичная продукция в водоемах озерного типа (ккал/(м2 год)) на текущем этапе и реконструкция этих показателей в отсутствие процесса эвтрофирования. Коэффициенты эвтрофирования Feu указаны в табл. 1. Результаты анализа СД
| Водоем | Продукция фитопланктона | Суммарная первичная продукция | ||
|---|---|---|---|---|
| на современном этапе | в отсутствие эвтрофирования | на современном этапе | в отсутствие эвтрофирования | |
| Богучанское вдхр | 586 | 404 | 633 | 446 |
| Зейское вдхр | 248 | 140 | 288 | 186 |
| Бурейское вдхр | 199 | 109 | 354 | 294 |
| оз. Ханка | 224 | 89 | 389 | 318 |
| Рыбинское вдхр | 1097 | 593 | 1340 | 776 |
| Ладожское озеро | 762 | 546 | 813 | 588 |
Исходя из значений TP, Pt, DAN и GS, чистая продукция фитопланктона меняется от ≈200 в олиготрофном Бурейском водохранилище до 1100 ккал/(м2 год) в мезотрофном с чертами эвтрофии Рыбинском водохранилище (табл. 1). Фитопланктон – основной продуцент органического вещества в Богучанском, Зейском, Рыбинском водохранилищах и озерах Онежском и Ладожском. На его долю приходится 82–94% PP суммарной первичной продукции. Вклад фитопланктона в PP суммарную первичную продукцию значительно меньше в Бурейском водохранилище и оз. Ханка (55–60%), но возрастает роль макрофитов, эпифитов и микрофитобентоса.
Отношение Pbcp к Pphp меняется от 23 (в Рыбинском водохранилище) до 70% (в Бурейском водохранилище). В Зейском водохранилище, отличающемся высокой цветностью воды (Pt = 75 град.) Pbcp может даже превышать Pphp. Большая величина Pbcp/Pphp свидетельствует о том, что бактериопланктоном ассимилируется не только автохтонное, но и в значительной мере – аллохтонное РОВ. Вычислительные эксперименты, выполненные с использованием СД, показали, что продукция бактерий за счет ассимиляции аллохтонного РОВ составляет от 15 (Рыбинское водохранилище) до 70 и 85% (Бурейское и Зейское водохранилища) суммарной продукции бактерий (табл. 3).
Таблица 3.
Средние значения продукции бактериопланктона за счет ассимиляции аллохтонного РОВ (Pbcp[all], ккал/(м2 год)), ее доли в общей продукции бактериопланктона (Pbcp[all]/Pbcp), продукции нехищного зоопланктона за счет потребления фитопланктона (Phzo[php], ккал/(м2год)) и ее доли в общей продукции нехищного зоопланктона (Phzo[php]/Phzo)
| Водоем | Pbcp[all] | Pbcp[all]/Pbcp | Phzo[php] | Phzo[php]/Phzo |
|---|---|---|---|---|
| Богучанское вдхр | 69 | 0.37 | 36 | 0.55 |
| Зейское вдхр | 269 | 0.85 | 15 | 0.23 |
| Бурейское вдхр | 100 | 0.71 | 12 | 0.35 |
| оз. Ханка | 12 | 0.19 | 10 | 0.51 |
| Рыбинское вдхр | 38 | 0.15 | 59 | 0.59 |
| Онежское озеро | 78 | 0.61 | 15 | 0.42 |
| Ладожское озеро | 146 | 0.49 | 47 | 0.50 |
Продукция зоопланктона Pzpl – это сумма продукций нехищного Phzo и хищного зоопланктона за вычетом рациона хищного зоопланктона. По результатам СД, вклад фитопланктона в Phzo варьирует от 23 (Зейское водохранилище) до 59% (Рыбинское водохранилище), вклад бактериопланктона – от 41 до 77% (табл. 3).
Pzpl составляет 5–15% Pphp. Повышенные значения Pzpl/Pphp указывают на присутствие аллохтонной компоненты в потоке вещества и энергии в трофической цепи. В среднем для всех анализируемых водоемов Pzpl составляет 8.4 ± 3.6% при медиане 7.3%.
Главный фактор, определяющий площадь литоральной зоны и, следовательно, вклад литоральных сообществ (макрофитов, эпифитов, фитобентоса, зообентоса и бентосоядных рыб) в общую биологическую продуктивность водоема озерного типа, – форма его котловины. По форме котловины все рассматриваемые озера и водохранилища (кроме Зейского) приближаются к водоемам равнинного типа, их литоральная зона занимает от 17 (Богучанское) до 54% (Рыбинское водохранилище) их акваторий. Ложе Зейского водохранилища ближе к каньонному типу, его литоральная зона занимает лишь 9.5% акватории. Отношение Pzbe/PP меняется от 0.8 (Ладожское озеро) до 5.4% (оз. Ханка). Малая величина Pzbe/PP свидетельствует о доминировании в экосистеме потока энергии через планктонное сообщество, большая величина – о доминировании роли бентосного сообщества. Отношение продукции рыбного сообщества к продукции зообентоса меняется от 7% в оз. Ханка до 45% в Зейском водохранилище и Ладожском озере, что означает превалирование в рыбном сообществе в первом случае – бентосоядных рыб, во втором – планктоноядных рыб.
Наиболее сложный и поэтому заслуживающий пристального внимания прогноз касается биологической продуктивности мелководного и огромного по площади лессового оз. Ханка с большим содержанием взвешенных минеральных веществ. Озеро находится на территории с умеренным муссонным климатом, продолжительность вегетационного периода – от 168 до 192 дней [52], что хорошо согласуется с данными СД – 180 дней.
По эмпирически установленному содержанию Chl, равному 7.3 мкг/л [2], легко реконструировать с помощью СД содержание в воде TP (27 мкг/л), а по TP и результатам натурных наблюдений за прозрачностью воды (Sec – 0.15–0.35 м) восстановить среднее значение Pt, отражающее содержание в воде аллохтонного РОВ. По [2], Pphp ≈ 200 ккал/(м2 год). Такая величина Pphp возможна при условии, если среднее за сезон DAN ≈70 мг C/мг. По [24], для оз. Ханка DAN ≈ ≈ 100 мг C/мг. Такая величина DAN сильно завышена, но, как следует из СД, снижение DAN до приемлемой величины сопряжено с удлинением вегетационного сезона. Вполне вероятно, что в оз. Ханка, где фотосинтез протекает и в зимнее время, благодаря отсутствию снежного покрова и прозрачности льда значение GS может быть выше рассчитанного по Tem.
По [2], общий запас рыб в оз. Ханка составляет от 4.25 до 14.5 тыс. т (при площади озера 4070 км2), или 1–3.6 ккал/м2. Принимая годовой P/B-коэффициент для рыб, равный 0.5, получаем, что FP варьирует от 0.5 до 1.8 ккал/(м2 год), что согласуется с результатом СД – 1.5 ккал/(м2 год) (табл. 1). Следовательно, допустимый вылов рыб (без подрыва популяции) может быть оценена величиной ≈0.5 ккал/(м2 год), или ≈5 кг/га. Совпадение эмпирических и спрогнозированных величин FP гарантирует надежность оценки продукции других ключевых групп гидробионтов.
Рыбопродуктивность водоемов (FP и FY) находится в прямой зависимости от Pphp, так как они связаны между собой потоком энергии через зоопланктон, зообентос и бактериопланктон. Эта взаимосвязь имеет большое практическое значение и издавна обсуждается в гидробиологической литературе [7, 13, 39, 49, 51]. В FP трансформируется 0.36–0.87% Pphp и 0.33–0.7% PP.
ЗАКЛЮЧЕНИЕ
Система диагностики биопродуктивности вод озерного типа создана с учетом накопленных знаний о трофических взаимоотношениях между ключевыми группами гидробионтов и о воздействии окружающей среды на функционирование водных экосистем. Она учитывает фундаментальный закон сохранения вещества и энергии и обладает определенным прогностическим потенциалом, который со временем может быть реализован в гидробиологической и рыбохозяйственной практике.
При исследовании озер и, в особенности, водохранилищ обнаруживается большое значение гетеротрофных бактерий как первоисточника энергии для организмов верхних трофических уровней. Бактериопланктон, утилизирующий аллохтонное РОВ, – дополнительный источник энергии для зоопланктона, который, в свою очередь, служит пищевым объектом для планктоноядных рыб. Следовательно, для прогнозирования общей биологической продуктивности и продукции рыбного сообщества следует учитывать продукцию не только автотрофного планктона, но и той части гетеротрофного бактериопланктона, которая специализируется на утилизации РОВ, поступающего в водоем извне. СД-анализ показал, что в продукцию рыбного сообщества трансформируется 0.32–0.62% (в среднем 0.43%) продукции первопищи (понятие “первопища” подразумевает сумму продукции фитопланктона и бактериопланктона за счет ассимиляции им аллохтонного РОВ [34]).
Важный фактор, определяющий продуктивность литоральных сообществ (макрофитов, эпифитов, фитобентоса, зообентоса и бентосоядных рыб), следовательно, экосистемы в целом, – форма котловины водоемов (соотношение средней и максимальной глубин) и глубина фотической зоны.
Список литературы
Алимов А.Ф. Продуктивность сообществ беспозвоночных макробентоса в континентальных водоемах СССР (Обзор) // Гидробиол. журн. 1982. Т. 18. № 2. С. 7–18.
Барабанщиков Е.И. Зоопланктон озера Ханка. Дис. … канд. биол. наук. СПб.: ЗИН РАН, 2004. 23 с.
Бульон В.В. Аллохтонное и автохтонное органическое вещество в трофической цепи озерных экосистем // Тр. ЗИН РАН. 2017. Т. 321. № 2. С. 115–128.
Бульон В.В. Биотический поток вещества и энергии в системе “озеро и его водосбор” // Успехи современной биологии. 2018. Т. 138. № 5. С. 503–513.
Бульон В.В. Вклад основных групп автотрофных организмов в первичную продукцию водоемов // Вод. ресурсы. 2004. Т. 31. № 1. С. 98–108.
Бульон В.В. Новый взгляд на парадигму фосфорного контроля в лимнологии // Успехи современной биологии. 2016. Т. 136. № 3. С. 311–318.
Винберг Г.Г., Ляхнович В.П. Удобрение прудов. М.: Легкая пром-ть, 1965. 271 с.
Герасимов Ю.В. Популяционная динамика рыб Рыбинского водохранилища за период его существования: роль естественных и антропогенных факторов // Тр. ВНИРО. 2015. Т. 156. С. 67–90.
Гидроэкологический мониторинг зоны влияния Бурейского гидроузла / Под ред. С.Е. Сиротского Хабаровск: ИВЭП ДВО РАН, 2007. 273 с.
Гидроэкологический мониторинг зоны влияния Зейского гидроузла / Под ред. С.Е. Сиротского Хабаровск: ИВЭП ДВО РАН, 2010. 354 с.
Даценко Ю.С., Пуклаков В.В., Эдельштейн К.К. Анализ влияния абиотических факторов на развитие фитопланктона в малопроточном стратифицированном водохранилище // Тр. Кар. НЦ РАН. 2017. № 10. С. 73–85.
Иванова М.Б. Продукция планктонных ракообразных. Л.: ЗИН АН СССР, 1985. 222 с.
Ивлев В.С. Интенсивность фотосинтеза и рыбная продукция прудов // Бюл. МОИП. Отд. Биол. 1939. Т. 48 (1). С. 29–34.
Инкина Г.А. Скорость потребления кислорода бактериопланктоном // Экспериментальные и полевые исследования биологических основ продуктивности озер. Л.: ЗИН АН СССР, 1979. С. 103–120.
Калинкина Н.М., Коросов А.В., Сярки М.Т. К созданию экспертной системы Онежского озера: оптимизация мониторинга состояния экосистемы по показателям зоопланктона // Принципы экологии. 2017. № 1. С. 117–132.
Калинкина И.М., Сярки М.Т., Теканова Е.В., Чекрыжева Т.А., Тимакова Т.М., Полякова Т.Н., Рябинкина А.В. Особенности формирования кормовой базы рыб Онежского озера // Органическое вещество и биогенные элементы во внутренних водоемах и морских водах. Петрозаводск: КНЦ РАН, 2012. С. 252–256.
Китаев С.П. Экологические основы биопродуктивности озер разных природных зон. М.: Наука, 1984. 207 с.
Коплан-Дикс И.С., Назаров Г.В., Кузнецов В.К. Роль минеральных удобрений в эвтрофировании вод суши. Л.: Наука, 1985. 182 с.
Копылов А.И., Косолапов Д.Б. Микробиальная “петля” в планктонных сообществах. Ижевск: КнигоГрад, 2011. 332 с.
Костина Н.В. Информационная система REGION: 25 лет работы и практического применения // Изв. Самарского НЦ РАН. 2015. Т. 17. № 4. С. 15–24.
Лит Х. Моделирование первичной продуктивности Земного шара // Экология. 1974. № 2. С. 13–23.
Меншуткин В.В., Руховец Л.А., Филатов Н.Н. Моделирование экосистем пресноводных озер (обзор). 2. Модели экосистем пресноводных озер // Вод. ресурсы. 2014. Т. 41. № 1. С. 24–38.
Меншуткин В.В., Филатов Н.Н. Разработка экспертной системы “озера Карелии” // Водная среда: комплексный подход к изучению, охране и использованию. Петрозаводск: Кар. НЦ РАН, 2008. С. 18–26.
Минеева Н.М., Щур Л.А., Бондаренко Н.А. Функционирование фитопланктона крупных пресноводных систем при разной обеспеченности ресурсами // Гидробиол. журн. 2012. Т. 48. № 3. С. 21–33.
Многолетние изменения биологических сообществ мезотрофного озера в условиях климатических флуктуаций и эвтрофирования / Под ред. И.С. Трифоновой. СПб.: ЛЕМА, 2008. 246 с.
Мосеев Д.С., Сергиенко Л.А., Кузьмина Е.Ю. Новые виды мхов (Bryophyta) для Земли Франца-Иосифа (Российская Арктика) // Новости систематики низших растений. 2018. Т. 52. № 1. С. 195–203.
Онежское озеро. Экологические проблемы / Под ред. Н.Н. Филатова. Петрозаводск: КарНЦ РАН, 1999. 259 с.
Оценка воздействия на окружающую среду Богучанской ГЭС на реке Ангаре. Предварительный вариант материала для ознакомления общественности. Красноярск: ЗАО “Богучанская ГЭС”, 2008. 454 с.
Прогноз качества воды в водохранилище и нижнем бьефе Богучанского водохранилища. Красноярск; Хабаровск: ИЛ СО РАН, 2009. 187 с.
Распопов И.М. Высшая водная растительность литоральной зоны Онежского озера // Литоральная зона Онежского озера. Л.: Наука, 1975. С. 103–123.
Румянцев В.А., Кудерский Л.А. Ладожское озеро: общая характеристика, экологическое состояние // Общество. Среда. Развитие (Terra Humana). 2010. № 1. С. 171–182.
Рыбохозяйственный атлас озера Ханка. Владивосток: ТИНРО – Центр, 2014. 205 с.
Савенко В.С. Фосфор в атмосферных осадках // Вод. ресурсы. 1996. Т. 23. № 2. С. 189−199.
Сорокин Ю.И. Первичная продукция морей и океанов // Общая экология. Биоценология. Гидробиология. Т. 1. М.: Наука, 1973. С. 7–46.
Теканова Е.В. Первичная продукция Онежского озера в современных условиях. Дис. … канд. биол. наук. СПб.: ИВП КарНЦ РАН, 2004. 23 с.
Тимакова Т.М., Сабылина А.В., Полякова Т.Н., Сярки М.Т., Теканова Е.В., Чекрыжева Т.А. Современное состояние экосистемы Онежского озера и тенденция ее изменения за последние десятилетия // Тр. КарНЦ РАН. 2011. № 4. С. 42–49.
Филатов Н.Н., Кухарев В.И., Регеранд Т.И., Лифшиц В.Х. Водные ресурсы Европейского Севера России: итоги и перспективы исследований. Петрозаводск: КарНЦ РАН, 2006. 538 с.
Фортунатов М.А. Физико-географический очерк Рыбинского водохранилища // Тр. Дарвинского гос. заповедника. 1974. Вып. 12. С. 5 – 30.
Цискаришвили Л.П. Взаимоотношение первичной продукции фитопланктона и рыбопродуктивности в олиготрофных и эвтрофных водоемах Грузии // Тр. Международ. симпоз. по проблемам математического моделирования процессов взаимодействия человеческой активности и окружающей среды. 1979. Т. 1. С. 122–128.
Шитиков В.К., Зинченко Т.Д., Головатюк Л.В. Нейросетевые методы оценки качества вод по гидробиологическим показателям // Изв. Самарского НЦ РАН. 2002.Т. 4. № 2. С. 280–289.
Эволюция круговорота фосфора и эвтрофирование природных вод / Под ред. К.Я. Кондратьева, И.С. Коплан-Дикса. Л.: Наука, 1988. 204 с.
Экологическая система Нарочанских озер. Экологическая система Нарочанских озер. Минск: Изд-во “Университетское”, 1985. 303 с.
Экосистема Онежского озера и тенденции ее изменения / Под ред. З.С. Кауфмана. Л.: Наука, 1990. 264 с.
Håkanson L., Boulion V.V. A practical approach to predict the duration of the growing season for European lakes // Ecol. Modelling. 2001. V. 140. P. 243–245.
Håkanson L., Boulion V.V. The Lake Foodweb – modelling predation and abiotic/biotic interactions. Leiden: Backhuys Publ., 2002. 344 p.
Lieth H. Primary production terrestrial ecosystems // Human Ecol. 1972. V. 1. P. 303–332.
Mooij W.M., Trolle D., Jeppesen E. et al. Challenges and opportunities for integrating lake ecosystem modelling approaches // Aquat. Ecol. 2010. V. 44. P. 633–667.
Nürnberg G.K., Shaw H. Productivity of clear and humic lakes: nutrients, phytoplank-379 ton, bacteria // Hydrobiologia. 1998. V. 382. № 1–3. P. 97–112.
Oglesby R.T. Relationships of fish yield to lake phytoplankton standing crop, production, and morphoedaphic factors // J. Fish. Res. Board Canada. 1977. V. 34. № 12. P. 2271–2279.
Olden J.D., Jackson D.A. Illuminating the ‘‘black box’’: a randomization approach for understanding variable contributions in artificial neural networks // Ecol. Modelling. 2002. V. 154. P. 135–150.
The functioning of freshwater ecosystems /Ed. Z. Kajak, A. Hillbricht-Ilkowska. Cambridge: Univ. Press, 1980. 918 p.
www.khanka-lake.ru
www.pogodaiklimat.ru
www.water-rf.ru/Рыбинское водохранилище
Дополнительные материалы отсутствуют.