

Водные ресурсы, 2021, T. 48, № 3, стр. 307-314
Закономерности пространственно-временнóй динамики сообществ хирономид (Chironomidae, Diptera) в Куйбышевском водохранилище
Т. Д. Зинченко a, *, В. К. Шитиков a
a Институт экологии Волжского бассейна РАН
445003 Тольятти, Россия
* E-mail: zinchenko.tdz@yandex.ru
Поступила в редакцию 21.04.2020
После доработки 11.09.2020
Принята к публикации 25.09.2020
Аннотация
Выполнен статистический анализ динамики донных сообществ различных плесов Куйбышевского водохранилища за многолетний период наблюдений – 1966–2016 гг. Представлены результаты сравнения пространственно-временнóго распределения структурных показателей сообществ хирономид (Chironomidae), включающих в себя видовое разнообразие и численность 48 таксонов по данным гидробиологической съемки на девяти крупных участках водохранилища за различные годы. Непараметрический дисперсионный анализ показал высокую статистическую значимость изменчивости видового состава донных сообществ под влиянием временнóго и пространственного факторов. С использованием многомерной ординации и обобщенного прокрустова анализа показана динамика распределения объектов экосистемы (отдельных видов и участков) в координатах двух латентных осей максимальной вариации. Выполнен геометрический анализ многомерных тр-аекторий изменения сообщества гидробионтов на отдельных плесах за различные периоды времени. Выявлены сходные процессы развития сообществ хирономид в продольной череде плесов на бывшем русле и затопленной пойме, тогда как аналогичные структурные характеристики Камского плеса и эвтрофного Черемшанского залива имеют специфический характер. Рассмотрена приоритетная роль хирономид как важнейшей индикаторной группы, необходимой для биотической оценки многолетних (50 лет) изменений экологического состояния крупного водохранилища.
ВВЕДЕНИЕ
Куйбышевское водохранилище – крупнейший водоем Волжского каскада, широко используемый для нужд энергетики, регулирования сезонного водоснабжения, сельскохозяйственной ирригации и рыболовства [4]. Водохранилище – мощный аккумулятор поверхностного стока с большой территории, приемник загрязненных антропогенных сточных вод. Все это обусловило необходимость комплексного мониторинга всех компонентов экосистемы, который проводился постоянно с 1957 по 1992 г.
Наличие больших массивов накопленной информации актуализирует проведение работ по математическому анализу и разработке экспертных прогнозов состояния экосистемы водохранилища при разных сценариях экономического развития территории, антропогенных нагрузок и климатических изменений [1]. Основные результаты математической обработки мониторинговых данных по Куйбышевскому водохранилищу представлены в [5], где подробно анализируются корреляционные связи между гидробиологическими и гидрохимическими характеристиками экосистемы. Ранее в [2, 19] обсуждались выявленные механизмы формирования структуры сообществ зообентоса под влиянием многолетних процессов эвтрофирования водоема. Поскольку рассматриваемая акватория имеет довольно сложную конфигурацию с чередованием озеровидных расширений и сужений, то актуальна проблема выделения областей относительно однородных по всему комплексу рассматриваемых показателей. В целях ее решения в [7] предложена процедура динамического районирования Куйбышевского водохранилища по совокупности абиотических и биотических факторов с использованием методов многомерной статистики и ГИС-технологий.
Одна из основных задач в области водной экологии – анализ динамики экосистем в многолетнем аспекте, в том числе изменение устойчивости и разнообразия биотических сообществ; скорость и направленность структурных деформаций; выявление ведущих факторов, определяющих такие изменения [17]. Однако до последнего времени отсутствовали общепринятые методики моделирования пространственно-временнóго тренда экосистем с достаточно большим числом компонент (таксонов, участков и периодов времени). Предложены различные многомерные методы [13], представляющие динамику экосистем с помощью ординационных диаграмм, на которых точками отображается таксономическая структура сообществ, наблюдаемая на участках в разное время. Методы геометрического анализа траекторий использовались для сравнения параметров состояния экосистемы в различных условиях путем вычисления корреляции Мантеля между соответствующими матрицами дистанций [9]. И лишь в последнее время предложены многомерные авторегрессионные модели и функции совместного распределения популяций, которые могут быть использованы для непосредственного моделирования взаимодействия видов и динамики сообществ [14, 16].
В настоящей работе рассматриваются результаты применения различных из перечисленных выше статистических методов для выявления основных закономерностей многолетней (50 лет) временнóй динамики таксонов Chironomidae как индикаторов экологического состояния Куйбышевского водохранилища [3].
МАТЕРИАЛЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
Для выполнения статистического анализа из всего массива гидробиологических данных по состоянию экосистемы Куйбышевского водохранилища сформирована репрезентативная выборка численностей различных видов Chironomidae (Diptera), относящихся к семи временны́ м точкам за период 1966–2016 гг. Выбор объекта исследования обусловлен многолетними таксономическими и структурными исследованиями ценоза хирономид как индикаторов состояния водоемов [2], адекватно реагирующих на интегральное “качество среды”.
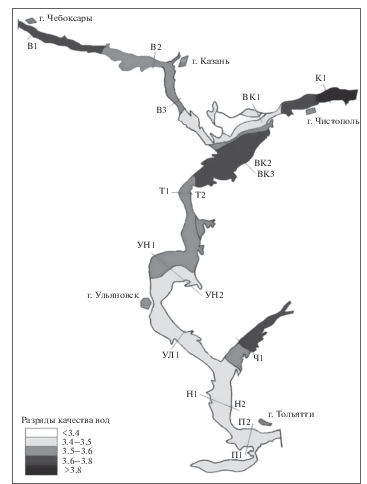
Чтобы обеспечить однородность данных и эквивалентность выборочного усилия, для каждого из девяти участков водохранилища (“плесов”) и каждого из семи временны́ х периодов случайным образом отбиралось по 4 гидробиологических пробы, в которых численность каждого вида усреднялась. Всего обнаружено 48 видов и таксонов хирономид. Основные характеристики плесов и пространственно-временнóе распределение индексов видового разнообразия представлены в табл. 1 и на рис. 1.
Таблица 1.
Распределение средних показателей видового разнообразия ценоза хирономид (Chironomidae) в плесах Куйбышевского водохранилища за разные периоды наблюдений (n – число видов; H – индекс разнообразия Шеннона, бит/экз.; N – численность, экз/м2)
| По периодам наблюдений | По участкам водохранилища | |||||||
|---|---|---|---|---|---|---|---|---|
| годы | n | H | N | плесы | код | n | H | N |
| 1966 | 9.67 | 1.54 | 677 | Волжский | В | 9.14 | 1.71 | 1014 |
| 1975 | 8.22 | 1.22 | 788 | Волго-Камский | ВК | 6.57 | 1.23 | 601 |
| 1984 | 8.00 | 1.17 | 1828 | Камский | К | 6.57 | 1.47 | 350 |
| 1992 | 3.56 | 1.03 | 1028 | Тетюшинский | Т | 7.14 | 1.40 | 662 |
| 2002 | 7.67 | 1.65 | 1360 | Ундорский | УН | 7.71 | 1.61 | 2128 |
| 2009 | 7.44 | 2.01 | 1752 | Ульяновский | УЛ | 5.71 | 1.29 | 1576 |
| 2016 | 4.11 | 1.60 | 481 | Новодевиченский | Н | 8.43 | 1.82 | 1832 |
| Среднее | 6.95 ± 0.43 | 1.46 ± 0.08 | 1131 ± 143 | Приплотинный | П | 6.43 | 1.76 | 1140 |
| Черемшанский залив | Ч | 4.86 | 0.83 | 873 | ||||
Исходную таблицу численностей видов 7 × 9 × 48 предварительно преобразовывали с использованием χ2-трансформации [12], которая позволяет наилучшим образом учесть как удельное влияние таксонов-доминантов с высокой популяционной плотностью, так и несомненную роль в сообществе комплекса редких видов. По формуле Брея–Кёртиса рассчитывалась матрица D 63 × 63 расстояний между каждой парой наблюдений в многомерном пространстве видов.
Статистический анализ включал в себя решение следующих задач:
непараметрический дисперсионный анализ npMANOVA [8], с помощью которого оценивали статистическую значимость изменчивости видового состава донных сообществ под влиянием временнóго и пространственного факторов;
проецирование всего массива данных выполненных наблюдений, представленных матрицей дистанции D, на ординационную плоскость с двумя осями главных координат [13];
обобщенный канонический корреляционный анализ пространства видов [15] и построение на его основе консенсусной прокрустовой конфигурации [11], сравнивающей пространственные распределения таксонов хирономид в изучаемые периоды времени;
геометрический анализ траекторий изменения сообществ СТА (Analysis of Community Trajectories [10]), выполненный по отдельным плесам для различных периодов времени.
Все расчеты выполняли с использованием среды программирования R версии 3.06 и ее дополнительных пакетов vegan, mixOmics, RVAideMemoire, v-egclust и qgraph.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Статистическую значимость влияния пространственно-временны́ х факторов на таксономическую структуру хирономидного комплекса оценивали с использованием непараметрического двухфакторного дисперсионного анализа. Для этого проводили разложение многомерной дисперсии, заключенной в матрице расстояний D, по уровням факторов “Период” и “Плес” с выделением случайных остатков, а р-значения оценивали с использованием алгоритмов рандомизации. Показано, что вариация данных, объясняемая каждым из факторов, статистически значимо превышает случайную ошибку наблюдений: при сравнении временны́ х периодов F = 2.79, р = = 0.002; при группировке по плесам F = 1.61, р = = 0.002.
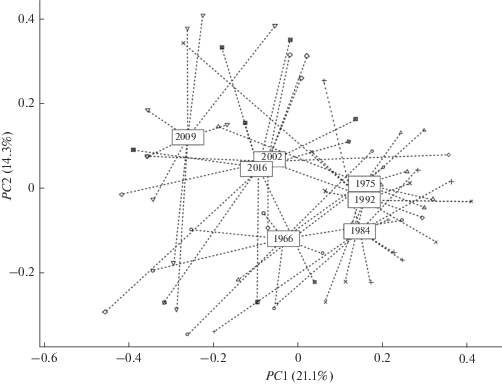
Матрицу дистанций D использовали для оптимального проецирования точек, соответствующих проведенным наблюдениям, из многомерного пространства видов на ординационную плоскость с двумя осями главных координат PC1–PC2 [13]. Расстояния на диаграмме (рис. 2) между центроидами позволяют оценить выраженность изменений среднего уровня видового сходства донных сообществ в течение рассматриваемого временнóго периода, а вариация точек относительно их общего центра тяжести отражает динамику сравнительного β-разнообразия сообществ хирономид, соответствующего каждому плесу. Можно отметить относительную структурную близость ценозов хиромид в 1975–1992 гг. и некоторую специфичность видового состава в 1966 г. и в период исследований в 2002–2009 гг., когда произошла замена реофильного комплекса хирономид фито- и пелофильным ценозом с преобладанием Dicrotendipes nervosus, Polypedilum bicrenatum, Cryptochironomus defectus, Cladotanytarsus mancus и др.
Рис. 2.
Ординационная диаграмма сообщества хирономид в разные периоды времени. Здесь и на рис. 3, 4 по осям абсцисс и ординат – значения первых двух главных координат РС1 и РС2. Показан разброс точек, соответствующих наблюдениям по плесам Куйбышевского водохранилища, относительно проекций центроидов по годам, представленным цифрами в прямоугольниках.
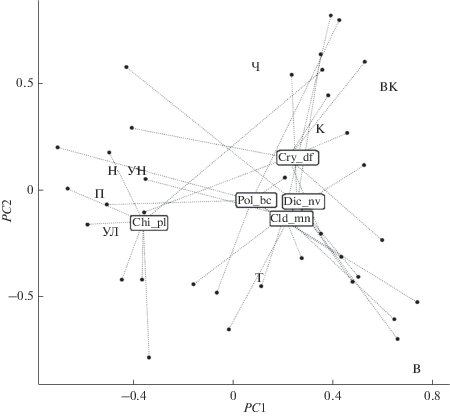
Для проецирования координат отдельных видов хирономид выполняли обобщенный анализ главных компонент с регуляризацией по каждому временнóму периоду, после чего все семь полученных частных ординаций объединяли с использованием прокрустовых преобразований. Диаграмма на рис. 3 показывает, как изменяется “центр тяжести” обилия анализируемых таксонов с течением времени и относительно отдельных плесов. Вариация групп точек относительно каждого их центроида отражает диапазон экологических условий обитания каждого вида, подчеркивая степень их эврибионтности.
Рис. 3.
Проекции центроидов координат отдельных плесов, обозначенных буквенными аббревиатурами из табл. 1, и диаграмма обобщенного прокрустова анализа временнóй динамики основных видов хирономид. Показан разброс точек, соответствующих наблюдениям за различные годы, относительно проекций центроидов видов, представленных кодами в прямоугольниках (Chi_pl – Chironomus plumosus; Cld_mn – Cladotanytarsus mancus; Cry_df – Cryptochironomus defectus; Dic_nv – Dicrotendipes nervosus; Pol_bc – Polypedilum bicrenatum).
Представленные выше алгоритмы ординации основаны на локальных сравнениях комплексов повторных наблюдений на одних и тех же участках, при этом анализ ограничивается их проекциями на две главные оси вариации, которые объясняют в настоящем примере только 35–38% совокупной дисперсии. Альтернативный метод анализа тра-екторий СТА [10] формально рассматривает динамику сообщества как взаимосвязанную последовательность точек в многомерном пространстве, описывающем сходство видовых структур без использования процедур редукции переменных. Ключевой аспект метода – возможность оценки обобщенной геометрической согласованности траекторий, что позволяет количественно анализировать и сравнивать выраженность структурных изменений в каждом изучаемом регионе. К сожалению, метод СТА не позволяет непосредственно визуализировать многомерные траектории, но можно выполнить их приближенное проецирование на плоскость двух главных координат (рис. 4), если характер данных позволяет пренебречь той информацией, которая аккумулирована в остальных осях.
Рис. 4.
Проекция на ординационную плоскость динамических траекторий видовой структуры хирономид по плесам, в легенде – условные обозначения из табл. 1. В узлах ломаных прямых цифрами представлены годы наблюдений.
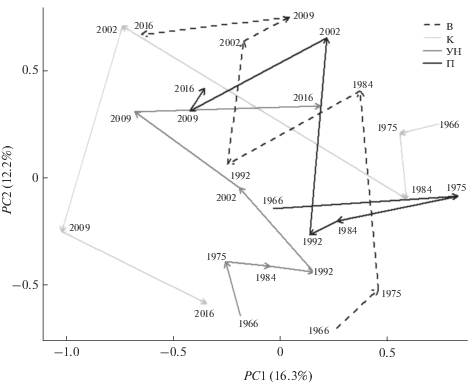
Такие важные геометрические компоненты траектории, как длина и угол поворота отдельных сегментов, скорость изменения и общая направленность, а также близость начальной и конечной точек, могут объяснить сукцессионные циклы сообществ и др. Предложены процедуры, позволяющие вычислить показатели дивергенции/конвергенции для любой пары наблюдаемых сообществ, т.е. рассмотреть, насколько синхронно меняется направленность двух траекторий с течением времени [10]. Для проверки гипотезы о наличии такой монотонной тенденции использовали тест Манна–Кендалла; например, авторами статьи установлено, что динамика изменения сообщества хирономид в Волжском и Ундорском плесах имеет по форме синхронный характер.
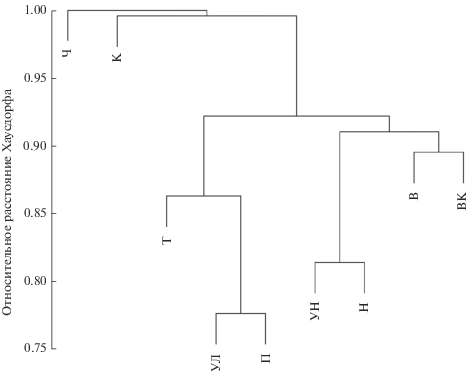
Другая геометрическая характеристика экосистемы – расстояние r Хаусдорфа между всеми парными траекториями. На основе вычисленной матрицы дистанций r с использованием методов кластерного анализа построена дендрограмма (рис. 5), показывающая, что относительно однородные процессы развития сообществ проходили в парах плесов Ундорский и Новодевиченский, Ульяновский и Приплотинный, Волжский и Волго-Камский, тогда как динамика Камского плеса и Черемшанского заливa имеет специфический характер.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Анализ многолетней динамики ценоза хирономид (Сhironomidae, Diptera) с 1966 по 1975 г. на всех плесах Куйбышевского водохранилища показал стабилизацию таксономического состава на фоне общего увеличения численности бентоса (табл. 1). Начиная с 1975 г., который был наиболее жарким и маловодным, наблюдается вспышка развития всего бентоса и при сохранении массовых видов в сообществе хирономид стали регистрироваться малочисленные виды Microchironomus tener, Harnischia fuscimana, Polypedilum scalaenum и др., встречаемость которых была невелика. Дальнейший анализ экологических показателей сообществ хирономид в период эвтрофирования водохранилища (1985–1992 гг.) показал снижение видового разнообразия [3] при одновременном росте общей численности за счет одного-двух доминантных видов (Chironomus plumosus, Procladius ferrugineus). После 1992 г. начался период сукцессионного восстановления экосистемы водохранилища, который продолжается до настоящего времени.
Статистический анализ пространственно-временнóй динамики распределения видов ценоза хирономид позволил провести количественное сравнение выраженности структурных изменений в каждом плесе водохранилища. На фоне значительного стохастического дрейфа, обусловленного многофакторными флуктуациями природно-климатического и антропогенного характера, выявлены выраженные детерминированные тенденции. В частности, на бывшем русле и затопленной пойме самого Куйбышевского водохранилища распределение видового разнообразия и численности сообществ хирономид относительно однородно, в отличие от водных масс участков Камского плеса и Черемшанского залива, где эти показатели существенно ниже (табл. 1). Выделен таксоценоз пелофильных видов, определяющих структурные особенности распределения хирономид. Так, на протяжении всех периодов и во всех районах доминирующими по численности были два таксона – Procladius ferrugineus и Chironomus plumosus. Личинки именно этих видов определяют особенности межгодовых изменений численности и биомассы бентоса водохранилища.
Геометрический анализ траекторий динамики сообществ и использование методов кластеризации позволили выявить относительно сходные процессы развития сообществ хирономид в парах плесов Ундорского и Новодевиченского, Ульяновского и Приплотинного, Волжского и Волго-Камского. Многолетняя динамика структурных характеристик ценоза хирономид Камского плеса и эвтрофного Черемшанского залива имеет специфический характер, что связано со значительной неоднородностью водных масс, особенностями гидрологических условий и гидрохимического режима.
Список литературы
Бухарин О.В., Захаров В.М., Зинченко Т.Д., Немцева Н.В., Розенберг Г.С., Шитиков В.К. Методы биомониторинга для оценки состояния антропогенно-нагруженной равнинной реки // Экология и пром-сть России. 2010. № 11. С. 10–15.
Зинченко Т.Д. Биоиндикационная роль хирономид (Díptera, Chironomidae) в водных экосистемах: проблемы и перспективы // Успехи современной биологии. 2009. Т. 129. № 3. С. 257–270.
Зинченко Т.Д. Многолетнее формирование зообентоса Куйбышевского водохранилища: динамика хирономид (Diptera: Chironomidae) в связи с процессами эвтрофирования // Изв. Самар. НЦ РАН. 2003. Спец. вып. 1. С. 91–101.
Куйбышевское водохранилище (научно-информационный справочник) / Отв. ред. Розенберг Г.С., Выхристюк Л.А. Тольятти: ИЭВБ РАН, 2008. 123 с.
Меншуткин В.В., Паутова В.Н., Номоконова В.Н., Селезнев В.А., Попченко И.И., Зинченко Т.Д., Иватин А.В., Выхристюк Л.А., Выхристюк М.М., Шитиков В.К., Казанцева Т.И. Статистические связи в экосистеме Куйбышевского водохранилища // Гидробиол. журн. 1998. Т. 34. № 5. С. 94–103.
Паутова В.Н., Номоконова В.И. Продуктивность фитопланктона Куйбышевского водохранилища. Тольятти, 1994. 189 с.
Шитиков В.К., Выхристюк Л.А., Паутова В.Н., Зинченко Т.Д. Многофакторное экологическое районирование Куйбышевского водохранилища // Вод. ресурсы. 2007. Т. 34. № 4. С. 481–489.
Anderson M.J. A new method for non-parametric multivariate analysis of variance // Austral. Ecol. 2001. V. 26. P. 32–46.
Clarke K.R., Somerfield P.J., Airoldi L., Warwick R.M. Exploring interactions by second-stage community analyses // J. Experimental Marine Biol. Ecol. 2006. V. 338. P. 179–192.
De Caceres M., Coll L., Legendre P., Allen R.B., Wiser S.K., Fortin M.-J., Condit R., Hubbell S. Trajectory analysis in community ecology // Ecological Monographs. 2019. V. 89. № 2. e01350. 20 p. https://doi.org/10.1002/ecm.1350
Gower J.C. Generalized procrustes analysis // Psychometrika. 1975. V. 40. P. 33–51.
Legendre P., Gallagher E. Ecologically meaningful transformations for ordination of species data // Oecologia. 2001. V. 129. P. 271–280.
Legendre P., Legendre L. Numerical ecology. Amsterdam: Elsevier Sci. BV., 2012. 990 p.
Ovaskainen O., Tikhonov G., Dunson D., Grøtan V., Engen S., Sæther B.-E., Abrego N. How are species interactions structured in species-rich communities? A new method for analysing time-series data // Proc. Royal Society. 2017. Ser. B 284. № 20170768. https://doi.org/10.1098/rspb.2017.0768
Tenenhaus A., Tenenhaus M. Regularized generalized canonical correlation analysis // Psychometrika. 2011. V. 76. P. 257–284.
Thorson J.T., Ianelli J.N., Larsen E.A., Ries L., Scheuerell M.D., Szuwalski C., Zipkin E.F. Joint dynamic species distribution models: a tool for community ordination and spatio-temporal monitoring // Global Ecol. Biogeogr. 2016. V. 25. P. 1144–1158.
Vellend M. The theory of ecological communities. Princeton, New Jersey: Princeton Univ. Press, 2016. 248 p.
Vikhristuk L. Hydrochemical State of the Kuibyshev Reservoir // Water Resour. Development. 1996. V. 12. № 4. P. 547–559.
Zinchenko T.D. Long-term (30 years) dynamics of Chironomidae (Diptera) fauna in the Kuibyshev water reservoir associated with eutrophication processes // Nether. J. Aguat. Ecol. 1992. V. 26. № 2–3. P. 533–542.
Дополнительные материалы отсутствуют.