

Зоологический журнал, 2019, T. 98, № 10, стр. 1112-1136
Эволюция и морфологическая изменчивость щечных зубов кударского пещерного медведя (Ursus kudarensis, Carnivora, Ursidae)
Г. Ф. Барышников a, *, А. Ю. Пузаченко b, **
a Зоологический институт РАН
199034 Санкт-Петербург, Россия
b Институт географии РАН
119017 Москва, Россия
* E-mail: g_baryshnikov@mail.ru
** E-mail: puzak@igras.ru
Поступила в редакцию 20.02.2019
После доработки 15.03.2019
Принята к публикации 20.03.2019
Аннотация
С помощью одномерного и многомерного статистического анализа изучена изменчивость во времени 1370 щечных зубов кударского пещерного медведя (Ursus kudarensis Baryshnikov 1985), собранных в пещерах Кударо 1 и Кударо 3 на Кавказе. Изученный материал включил два хроноподвида – U. k. praekudarensis (середина среднего плейстоцена, около 350 тыс. л. н.) и U. k. kudarensis (поздний плейстоцен, приблизительно 120–40 тыс. л. н.), а также выборку стратиграфического промежуточного положения, обозначенную как переходная форма (конец среднего плейстоцена, около 250 тыс. л. н.). Наше исследование не выявило единого пути эволюционных преобразований щечных зубов U. kudarensis, а показало его своеобразие у премоляров и моляров и даже у отдельных зубов (например, m3) на фоне высокой индивидуальной изменчивости. По размерам зубы U. k. praekudarensis занимают промежуточное положение между более крупными зубами переходной формы и более мелкими зубами U. k. kudarensis. По совокупности морфометрических признаков (размеры, пропорции коронки) переходная форма стоит ближе к U. k. kudarensis, чем к U. k. praekudarensis. В то же время параметры аллометрической изменчивости переходной формы сближают ее с U. k. praekudarensis или же носят оригинальный характер. Расчетная скорость эволюции метрических признаков оказалась различной не только у разных зубов, но и у разных промеров одной зубной коронки, а сами эти изменения демонстрируют разную направленность. Сравнение с основными эволюционными трендами у европейских пещерных медведей показало уникальный, независимый характер эволюции щечных зубов кударского медведя, своеобразие которой выражено в разной степени в верхнем и нижнем рядах.
Изучение древней митохондриальной ДНК пещерных медведей открыло существование нескольких независимых эволюционных линий (Knapp et al., 2009; Stiller et al., 2010). Базальное положение среди них занимает кударский пещерный медведь (Ursus kudarensis Baryshnikov 1985) (Dabney et al., 2013). Первоначально его эволюционная самостоятельность была обоснована признаками зубной морфологии при изучении палеонтологического материала из кударских палеолитических памятников на Кавказе (Baryshnikov, 1998). Позднее она тестировалась морфометрическими данными по пещерным медведям из разных генетических групп и географических районов, которые подтвердили морфологическое своеобразие кударского медведя (Baryshnikov, Puzachenko, 2011, 2017, 2019; Baryshnikov et al., 2018).
Кударский пещерный медведь известен из многих кавказских местонахождений (Барышников, 2007), и его считали эндемичным для региона то тех пор, пока А.В. Шер не передал для изучения древней ДНК таранную кость с р. Адыча в Восточной Сибири, которая показала генетическую близость с U. kudarensis (Sher et al., 2011). Тем самым было не только впервые доказано существование пещерных медведей на севере Сибири, но и стало очевидным, что ареал кударского медведя был в прошлом обширнее, и лишь к позднему плейстоцену площадь его сократилась.
На основе кударского материала были выделены два хроноподвида, сменяющие друг друга во времени: U. k. praekudarensis Baryshnikov 1998 из среднего плейстоцена (слой 5 в пещере Кударо 1) и U. k. kudarensis из позднего плейстоцена (слои 3–4 в пещерах Кударо 1 и Кударо 3). Еще одна выборка обнаружила переходные признаки морфологии щечных зубов (слои 5–8 в пещере Кударо 3) (Baryshnikov, 1998). Мы обозначаем ее как переходную форму, подчеркивая тем самым филогенетическую преемственность между обоими подвидами. Морфологический анализ верхних резцов показал, что переходная форма показывает больше сходства с U. k. kudarensis, чем с U. k. praekudarensis (Baryshnikov et al., 2019).
Все находки кударского медведя в пещерах Кударо 1 и Кударо 3 имеют стратиграфическую привязку, и, таким образом, мы обладаем уникальной возможностью проследить эволюционную изменчивость пещерного медведя на протяжении второй половины плейстоцена и определить темпы и направления выявленных изменений.
В данной статье в фокусе нашего внимания находятся щечные зубы, которые были разделены на три выборки: U. k. praekudarensis, U. k. kudarensis и переходная форма. Задачей исследования было проанализировать их морфометрическую изменчивость с помощью различных методов одномерной и многомерной статистики с целью обнаружения эволюционных трендов и оценки скорости эволюционны преобразований отдельных зубов.
МАТЕРИАЛЫ И МЕТОДЫ
Местонахождения и объем выборки. Пещеры Кударо 1 и Кударо 3 расположены на южном макросклоне Главного Кавказского хребта в пределах Южной Осетии (42.46° с. ш., 43.95° в. д.) на высоте около 1600 м над ур. м. Обе пещеры, расположенные поблизости друг от друга, содержат каменную индустрию раннего и среднего палеолита и были раскопаны под руководством В.П. Любина как археологические стоянки (Любин, 1998).
В пещере Кударо 1 выделены 5 культурных слоев, среди которых палеолитические слои 3–5 содержат зубы кударского медведя. Из них самый нижний, ашельский слой 5 относится к среднему плейстоцену и два выше лежащих, мустьерских слоя 3–4 принадлежат к позднему плейстоцену. Для базальной части пещерных отложений были получены радиотермолюминисцентным методом даты 360 ± 90 тыс. лет назад (RTL-379) для слоя 5в и 350 ± 70 тыс. л. н. (RTL-373) для слоя 5б. Мустьерский слой 3а был датирован радиокарбоновым методом как имеющий возраст 44 150 ± 2400/1850 лет назад (Gr-6079) (Любин, 1998).
В пещере Кударо 3 было выделено 8 слоев, из которых ашельские слои 5–8 относятся к среднему плейстоцену, в то время как мустьерские слои 3–4 лежат в пределах позднего плейстоцена (Любин, 1998). Геоморфологические данные свидетельствуют, что вход в пещеру Кударо 3, расположенный ниже по склону, был вскрыт эрозией приблизительно на 50 000–100 000 лет позднее, чем открылась пещера Кударо 1 (Несмеянов, 1999). Для размытого контакта ашельских и мустьерских слоев получены две RTL-даты: 252 ± ± 51 и 245 ± 49 тыс. л. н. (Любин, 1998). Для медвежьих костей из слоя 3 имеются три AMS радиоуглеродных даты: > 41600 (OxA-19611), 47 900 ± ± 2.500 (OxA-19612) и 47700 ± 1800 лет назад (OxA-19 613) (Baryshnikov, 2011).
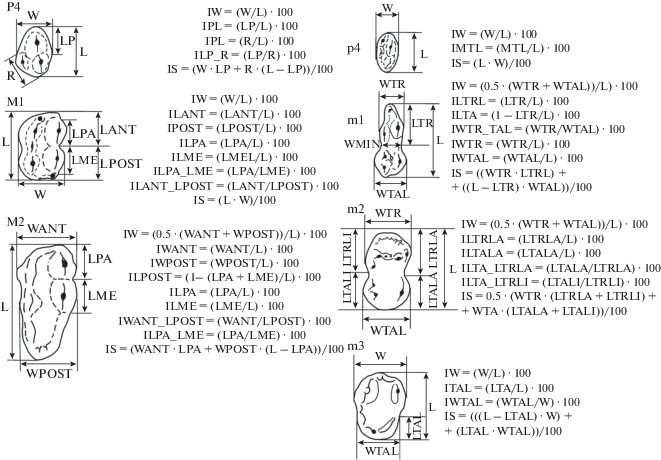
Кударский медведь доминирует в палеонтологическом материале из пещер Кударо 1 и Кударо 3. Всего были исследованы 854 нижних (p4, m1–m3) и 521 верхних (P4, M1, M2) щечных зубов (табл. 1), хранящихся в коллекции Зоологического института РАН (Санкт-Петербург). Схема промеров и формулы индексов даны на рис. 1.
Таблица 1.
Количество изученных зубов кударского пещерного медведя
| Таксон | Местонахождение | Возраст, Ma (по: Baryshnikov, 1998) | Щечные зубы | |||
|---|---|---|---|---|---|---|
| p4 | m1 | m2 | m3 | |||
| U. k. praekudarensis | Кударо 1, 5a-f | > ~ 0.36–0.35 | 100 | 119 | 111 | 131 |
| Переходная форма | Кударо 3, 5-8 | ~0.25–0.24 | 33 | 42 | 43 | 45 |
| U. k. kudarensis | Кударо 1, 3-4; Кударо 3, 3-4 | ~0.12–0.05 | 52 | 54 | 69 | 55 |
| Итого | 185 | 215 | 223 | 231 | ||
| P4 | M1 | M2 | ||||
| U. k. praekudarensis | Кударо 1, 5a-f | > ~ 0.36–0.35 | 116 | 101 | 66 | |
| “переходная форма” | Кударо 3, 5-8 | ~0.25–0.24 | 37 | 53 | 45 | |
| U. k. kudarensis | Кударо 1, 3-4; Кударо 3, 3-4 | ~0.12–0.05 | 28 | 37 | 38 | |
| Итого | 181 | 191 | 149 | |||
Рис. 1.
Схема промеров верхних (P4, M1, M2) и нижних (p4, m1 – m3) щечных зубов кударского медведя (Baryshnikov, 2006) и формулы для расчета индексов. Обозначения промеров: L – наибольшая длина зуба, W – наибольшая ширина зуба, LP – длина паракона P4, LPA – длина паракона M1 и M2, LME – длина метакона, R – наименьшее расстояние между передним краем протокона и задним краем коронки P4, LANT – длина передней части M1, LPOST – длина задней части M1, WANT – наибольшая ширина M2, WPOST – ширина талона M2, LTR – длина тригонида m1, WTR – ширина тригонида m1 и m2, WMIN – ширина m1 в средней части, WTAL – ширина талонида m1, m2 и m3, LTRLA – длина тригонида m2 с лабиальной стороны, LTALA – длина талонида m2 с лабиальной стороны, LTRLI – длина тригонида m2 с лингвальной стороны, LTALI – длина талонида m2 с лингвальной стороны, LTAL – длина талонида m3.
Методы. Методика анализа морфометрической изменчивости зубов кударского медведя была разработана и апробирована ранее на многих объектах (Пузаченко, 2001; Куприянова и др., 2003, Baryshnikov, Puzachenko, 2011, 2017, 2019). В общем случае она включает: подготовку данных (промеров), построение многомерной модели, описывающей основные закономерности (порядок) изменчивости, и анализ полученных результатов. Основная цель подготовки данных – исключение ошибок измерений и заполнение пропущенных данных. Для поиска выбросов использовали визуальный метод – анализ скаттер-диаграмм и тест Граббса (Stefansky, 1972). При анализе пропущенных значений использовали EM-алгоритм (Dempster et al., 1977) и регрессионный анализ. Для приведения измерений зубов к одному масштабу использовали стандартизацию в соответствии с формулой: ${{x}_{{{\text{i}},{\text{rang}}}}}$ = $ = {{\left( {{{x}_{i}} - {{x}_{{{\text{min}}}}}} \right)} \mathord{\left/ {\vphantom {{\left( {{{x}_{i}} - {{x}_{{{\text{min}}}}}} \right)} {\left( {{{x}_{{{\text{max}}}}} - {{x}_{{{\text{min}}}}}} \right)}}} \right. \kern-0em} {\left( {{{x}_{{{\text{max}}}}} - {{x}_{{{\text{min}}}}}} \right)}}$, где xmin и xmax минимальное и максимальное значения промера, соответственно. Построение многомерной модели преследует цель компактного описания изменчивости и выявления основных ее независимых компонент на фоне случайных вариаций (“шума”). Модель представляет собой евклидово пространство с координатами (виртуальными переменными), содержащими основную информацию о варьировании исходных переменных. Каждый объект (зуб) представлен в нем точкой с координатами, определяющими его геометрическое положение относительно всех других объектов с учетом всех исходных измерений. Таким образом, координаты модели задают “морфологическое пространство”, биологическое содержание которого зависит от способа оценки различий между объектами (Пузаченко, 2016). Для построения морфологического пространства мы используем метод неметрического многомерного масштабирования (NMDS) (Davison, Jones, 1983).
В качестве исходных данных для каждого зуба рассчитывались две матрицы “морфологических дистанций”. Первая матрица попарных стандартизированных евклидовых дистанций отражала различия между зубами по размерам. Соответствующая модель морфопространства описывает размерную изменчивость и обозначается как «морфопространство размеров) (МР). Во втором случае морфологические дистанции были получены с использованием ранговой корреляции tau-b Кендалла (Kendall, 1975). Последняя метрика отражает в обобщенной виде различия по форме объектов, а соответствующее ей морфопространство содержит информацию о разнообразии формы черепа (“морфопространство формы”, МФ) (Пузаченко, 2016). Для всех моделей оптимальная размерность (число координат) оценивалось по варьированию показателя “стресса Крускала” (Kruskal, 1964; Куприянова и др., 2003). Анализ моделей включал графическое представление результатов в виде скаттер-диаграм первых координат МР для демонстрации масштаба различий между зубами медведей из групп местонахождений разного геологического возраста (табл. 2), исследование структуры моделей с применением корреляционного анализа, оценку методом одномерного дисперсионного анализа (ANOVA) влияния принадлежности экземпляров зубов к группам на значения координат.
Таблица 2.
Коэффициенты корреляции координат МР (E) и МФ (K) моделей с промерами/индексами нижних щечных зубов и величина объясненной дисперсии координат моделей принадлежностью к группам с разным геологическим возрастом (фактор – “таксон”, %)
| Переменная | p4 | ||||||
|---|---|---|---|---|---|---|---|
| E1 | K1 | ||||||
| L | 0.86 | –0.26 | |||||
| W | 0.88 | 0.51 | |||||
| IW | 0.10 | 0.81 | |||||
| IS | 0.99 | 0.16 | |||||
| Фактор “таксон”, % | – | – | |||||
| m1 | |||||||
| E1 | E2 | K1 | K2 | K3 | |||
| L | 0.89 | –0.41 | –0.29 | 0.26 | –0.22 | ||
| LTR | 0.83 | –0.52 | –0.43 | 0.17 | –0.12 | ||
| WTR | 0.86 | 0.27 | 0.24 | –0.21 | –0.35 | ||
| WTAL | 0.87 | 0.29 | 0.37 | 0.39 | –0.16 | ||
| WMIN | 0.86 | 0.41 | 0.40 | –0.03 | 0.08 | ||
| IW | 0.15 | 0.92 | 0.79 | –0.16 | –0.08 | ||
| IWTR | 0.19 | 0.77 | 0.59 | –0.52 | –0.20 | ||
| IWTAL | 0.09 | 0.84 | 0.79 | 0.21 | 0.06 | ||
| IWTR_TAL | 0.12 | 0.03 | –0.12 | –0.77 | –0.28 | ||
| IS | 0.98 | –0.03 | 0.04 | 0.16 | –0.29 | ||
| Фактор “таксон”, % | – | 30.7 | 38.8 | – | – | ||
| m2 | |||||||
| E1 | E2 | E3 | E4 | K1 | K2 | K3 | |
| L | 0.95 | –0.11 | –0.15 | 0.09 | –0.02 | –0.07 | 0.00 |
| LTRLA | 0.83 | –0.11 | –0.48 | 0.03 | –0.21 | –0.30 | 0.07 |
| LTRLI | 0.74 | –0.13 | –0.11 | 0.58 | –0.02 | 0.10 | 0.43 |
| LTALA | 0.77 | –0.34 | 0.51 | 0.09 | 0.04 | 0.51 | –0.18 |
| LTALI | 0.75 | –0.33 | –0.13 | –0.50 | –0.21 | –0.16 | –0.50 |
| WTR | 0.80 | 0.55 | 0.03 | 0.01 | 0.53 | –0.25 | 0.02 |
| WTA | 0.85 | 0.30 | 0.21 | –0.19 | 0.46 | –0.05 | –0.22 |
| IW | –0.07 | 0.79 | 0.39 | –0.26 | 0.77 | –0.12 | –0.16 |
| ILTALA | 0.06 | –0.35 | 0.86 | 0.03 | 0.08 | 0.78 | –0.26 |
| ILTA_TR_LI | 0.12 | –0.20 | –0.03 | –0.95 | –0.17 | –0.24 | –0.83 |
| IS | 0.99 | 0.10 | 0.00 | –0.03 | 0.20 | –0.09 | –0.09 |
| Фактор “таксон”, % | 14.4 | 33.4 | – | 6.2 | 24.2 | 7.2 | – |
| m3 | |||||||
| E1 | E2 | K1 | K2 | K3 | |||
| L | 0.93 | 0.00 | 0.06 | –0.41 | 0.11 | ||
| LTAL | 0.82 | –0.55 | 0.49 | –0.02 | –0.12 | ||
| W | 0.85 | 0.46 | –0.41 | –0.14 | –0.16 | ||
| WTAL | 0.90 | 0.19 | –0.13 | 0.13 | 0.19 | ||
| IW | –0.41 | 0.53 | –0.56 | 0.46 | –0.37 | ||
| ILTAL | 0.41 | –0.77 | 0.63 | 0.30 | –0.26 | ||
| IWTAL | 0.50 | –0.20 | 0.24 | 0.36 | 0.47 | ||
| IS | 0.97 | 0.20 | –0.12 | –0.24 | 0.07 | ||
| Фактор “таксон”, % | – | – | – | – | – | ||
Оценка масштаба дифференциация групп в морфопространстве может быть получена на основе евклидовых дистанций между групповыми центроидами. Более полное представление о дифференциации дают методы, учитывающие внутригрупповой разброс точек, который определяет величину объема (“ширину”) “морфологической ниши” разных групп, их пересечение. Ширина ниши группы по любой координате пропорциональна энтропии Шеннона (например, Colwell, Futuyma (1971) для экологических ниш) распределения значений координат (Abramov, Puzachenko, 2012). “Ширина” индивидуальной ниши таксона вычисляется как сумма значений энтропии (Hi) (размерность – биты) по всем координатам морфопространства. Для сравнения ниш в пределах одного и того же морфопространства мы предлагаем использовать безразмерную величину (si), характеризующую долю разнообразия в i-той группы (в частности таксона) относительно разнообразия всех групп в данном морфопространстве (Hjnt): ${{s}_{i}} = \frac{{{{2}^{{{{H}_{i}}}}}}}{{{{2}^{{{{H}_{{jnt}}}}}}}} = {{2}^{{\left( {{{H}_{i}} - {{H}_{{jnt}}}} \right)}}}$. При сравнения малых выборок возможна ситуация Hi > Hjnt. В этом случае si приравнивается к 1.
Для оценки пересечения ниш мы используем модифицированный индекс Пианки (Pianka, 1974) в форме (Abramov, Puzachenko, 2012): $O_{{jk}}^{R} = \frac{{\left( {\mathop \prod \nolimits_1^n \mathop \sum \nolimits_1^R {{p}_{{Rj}}}{{p}_{{Rk}}}} \right)}}{{\sqrt {\mathop \prod \nolimits_1^n \left( {\mathop \sum \nolimits_1^R p_{{Rj}}^{2}\mathop \sum \nolimits_1^R p_{{Rk}}^{2}} \right)} }}$, где ${{p}_{{Rj}}}$ и ${{p}_{{Rk}}}$ частоты распределений пары групп j и k на интервале значений n-й координаты морфопространства c числом градаций R. Число градаций определяется для всего диапазона значений координат морфопространства по формуле $R = 1 + {{\lg }_{2}}N$, где N – общий объем выборки всех групп (не только j и k, если число групп больше двух). Величина индекса изменяется от 0 (пересечение ниш отсутствует) до 1 (ниши групп не различаются). При этом для того, чтобы две морфологические ниши не пересекались, необходимо и достаточно отсутствие их пересечения хотя бы относительно одной координаты морфопространства.
Кроме этого, в работе использован тест Манна–Уитни для оценки статистической значимости различий средних значений промеров, индексов и координат морфопространств между разновозрастными группами местонахождений.
Для исследования относительной изменчивости промеров щечных зубов использовали метод многомерной аллометрии (Cock, 1966; Klingenberg, 1996), который позволяет в обобщенной форме (коэффициент многомерной аллометрии, MAC) описать характер изменений промеров при изменении “общих” размеров зуба. MAC определяется как собственное значение конкретного промера для первой главной компоненты (PC1), полученное на основе ковариационной матрицы всех логарифмированных промеров, на ${1 \mathord{\left/ {\vphantom {1 {{{m}^{{0.5}}}}}} \right. \kern-0em} {{{m}^{{0.5}}}}}$ (Jolicoeur, 1963), где m – общее число промеров зуба. Значение MAC больше 1 соответствуют положительной аллометрии, при которой увеличение промера опережает рост общих размеров зуба. Соответственно, если MAC меньше 1, но больше 0 (отрицательная аллометрия), то скорость изменения промера отстает от роста общих размеров. Значения MAC, приблизительно равные 1 или 0, предполагают изометрию или отсутствие аллометрической изменчивости для данного промера, соответственно. Наконец, если MAC меньше 0, то значение промера аллометрически уменьшается при увеличении общих размеров зуба (обратная аллометрия). Доверительный интервал для MAC оценивали по результатам 100 бутстреп-повторностей.
Первая компонента модели для всей выборки (GPC1) описывает общие изменения размеров зубов по отношению к PC1 частных групповых моделей для обоих подвидов и переходной формы. Величина угла в градусах ${{\theta }_{{i,j}}}$ между PC1 двух групп i и j равна: ${\text{t}}{{{\text{g}}}^{{ - 1}}}{{\theta }_{{i,j}}} = ({{{{{180}}^{^\circ }}} \mathord{\left/ {\vphantom {{{{{180}}^{^\circ }}} \pi }} \right. \kern-0em} \pi }){\text{t}}{{{\text{g}}}^{{ - 1}}}\left( {{{b}_{i}} - {{b}_{j}}} \right)$, где b – коэффициенты регрессий PC1 = a + bGPC1. Величина угла между первыми главными компонентами отражает в обобщенном виде различия/сходства между “аллометрическими паттернами” разных групп.
Относительное положение подвидов U. kudarensis и переходной формы (центроиды групп в МР и МФ моделях, ${{\theta }_{{i,j}}}$) иллюстрировано дендрограммами, для построения которых применяли метод невзвешенных попарных средних (UPMGA).
В рамках гипотезы о непрерывной последовательной эволюции между подвидами кударского медведя мы оценили “скорость эволюции” значений отдельных промеров и индексов, выраженную в халденах (Haldane, 1949; Lynch, 1990) на двух временных интервалах: U. k. praekudarensis – переходная форма ~ 110 тыс. лет (t1 – t2) и переходная форма – U. k. kudarensis ~ 130 тыс. лет (t2 – t3). Скорость эволюции морфологического признака x между моментами времени t1 и t2 рассчитывалась по формуле: h = ${{\left| {\left( {\ln {{x}_{2}} - \ln {{x}_{1}}} \right)} \right|} \mathord{\left/ {\vphantom {{\left| {\left( {\ln {{x}_{2}} - \ln {{x}_{1}}} \right)} \right|} {SD\left( {\ln x} \right)}}} \right. \kern-0em} {SD\left( {\ln x} \right)}}$ × × $\left( {{{t}_{2}} - {{t}_{1}}} \right)$, где SD(ln x) – среднее стандартное отклонение логарифмированных значений признака для совокупной выборки. Мы не использовали оценку для времени существования генерации, исходя из предположения о ее постоянстве в пределах вида U. kudarensis.
РЕЗУЛЬТАТЫ
Модели морфопространств и структура изменчивости щечных зубов. Размерность МР и МФ моделей варьирует от 2 до 4 (табл. 2, 3). В случае p4 размерность 1 определяется числом промеров (2) зуба. Независимо от размерности МР моделей их первая координата содержит основную информацию об изменчивости размеров зубов, что отражается в коэффициентах корреляции координаты E1 с самими промерами и индексом “жевательной поверхности” (IS), который пропорционален “общим размерам” зуба. Вторая координата моделей (E2) коррелирует с некоторыми измерениями, характеризующими элементы зуба (тригонид, талонид, метакон, паракон), демонстрирующими частичную или полную независимость от изменчивости общего размера зубной коронки. У нижних щечных зубов и верхнего премоляра (P4) координата содержит информацию о варьировании относительной ширины зуба (индекс IW). Это отражает существенную независимость изменчивости относительной ширины от общих размеров зубов и ее связь с изменчивостью размеров тригонида и/или талонида в нижнем ряду и протокона P4 в верхнем. Соответственно, E2 демонстрирует корреляцию с индексами, характеризующими относительные размеры перечисленных элементов конструкции зуба. В случаях, когда размерность МР модели была больше двух (m2, M2), координаты E3 или E4 были содержательно связаны с компонентами изменчивости абсолютных и относительных размеров талонида и тригонида (m2), паракона и метакона (M2).
Таблица 3.
Коэффициенты корреляции координат МР (обозначения координат E) и МФ (обозначения координат K) моделей с промерами/индексами верхних щечных зубов и величина объясненной дисперсии координат моделей принадлежностью к группам с разным геологическим возрастом (фактор – “таксон”, %)
| Переменная | P4 | |||||
|---|---|---|---|---|---|---|
| E1 | E2 | K1 | K2 | K3 | ||
| L | 0.90 | –0.33 | –0.33 | –0.19 | –0.01 | |
| LP | 0.90 | –0.30 | –0.21 | 0.25 | 0.05 | |
| W | 0.92 | 0.30 | 0.21 | –0.07 | 0.31 | |
| R | 0.90 | 0.36 | 0.34 | –0.08 | –0.07 | |
| IW | 0.31 | 0.81 | 0.69 | 0.11 | 0.46 | |
| ILP | 0.34 | –0.07 | 0.09 | 0.74 | 0.10 | |
| IR | 0.37 | 0.83 | 0.81 | 0.08 | –0.09 | |
| ILP_R | –0.13 | –0.84 | –0.70 | 0.40 | 0.16 | |
| IS | 0.99 | 0.05 | 0.01 | –0.15 | 0.09 | |
| Фактор “таксон”, % | 11.1 | 39.0 | 37.2 | – | 10.5 | |
| M1 | ||||||
| E1 | E2 | K1 | K2 | K3 | K4 | |
| L | 0.93 | –0.13 | –0.23 | –0.26 | –0.18 | 0.11 |
| LANT | 0.88 | 0.08 | 0.02 | –0.25 | –0.29 | 0.22 |
| LPOST | 0.83 | –0.41 | –0.55 | 0.12 | –0.01 | 0.15 |
| LPA | 0.78 | 0.35 | 0.23 | –0.07 | 0.29 | 0.32 |
| LME | 0.68 | 0.52 | 0.23 | 0.22 | –0.07 | –0.35 |
| W | 0.87 | –0.15 | –0.27 | –0.42 | 0.09 | –0.19 |
| IW | 0.09 | –0.07 | –0.13 | –0.34 | 0.43 | –0.51 |
| ILPOST | 0.09 | –0.48 | –0.58 | 0.52 | 0.22 | 0.10 |
| ILPA | –0.05 | 0.59 | 0.55 | 0.20 | 0.56 | 0.29 |
| ILME | 0.00 | 0.71 | 0.46 | 0.47 | 0.07 | –0.50 |
| ILPA_LME | –0.03 | –0.25 | –0.04 | –0.31 | 0.34 | 0.70 |
| ILANT_LPOST | –0.07 | 0.60 | 0.71 | –0.42 | –0.32 | 0.04 |
| IS | 0.94 | –0.14 | –0.26 | –0.36 | –0.04 | –0.05 |
| Фактор “таксон”, % | – | – | – | – | 3.2 | 2.3 |
| M2 | ||||||
| E1 | E2 | E3 | K1 | K2 | K3 | |
| L | 0.90 | 0.02 | –0.17 | 0.50 | –0.02 | –0.55 |
| LPA | 0.46 | –0.55 | 0.69 | –0.47 | 0.33 | –0.39 |
| LME | 0.29 | –0.79 | –0.43 | –0.23 | –0.66 | –0.05 |
| WANT | 0.91 | 0.10 | –0.11 | 0.46 | –0.02 | –0.31 |
| WPOST | 0.69 | 0.51 | 0.17 | 0.56 | 0.43 | 0.20 |
| IW | –0.04 | 0.43 | 0.23 | 0.09 | 0.36 | 0.68 |
| IWPOST | –0.01 | 0.55 | 0.34 | 0.19 | 0.50 | 0.70 |
| ILPA | –0.13 | –0.55 | 0.78 | –0.78 | 0.33 | –0.03 |
| ILME | –0.18 | –0.79 | –0.33 | –0.49 | –0.63 | 0.23 |
| IWANT_POST | –0.04 | –0.52 | –0.29 | –0.27 | –0.52 | –0.50 |
| ILPA_LME | 0.07 | 0.35 | 0.85 | –0.09 | 0.83 | –0.22 |
| ILPOST | 0.19 | 0.84 | –0.25 | 0.79 | 0.21 | –0.13 |
| IS | 0.95 | 0.24 | 0.00 | 0.58 | 0.20 | –0.26 |
| Фактор “таксон”, % | 7.3 | 5.9 | 3.9 | – | 4.0 | – |
В целом размерность МР моделей зависит от числа независимых компонент изменчивости зуба, воспроизводимым данным набором измерений. В общем случае размерность связана с числом “степеней свободы” изменчивости зуба, а набор промеров (= “признаков”), специфически связанных с разными координатами (“степенями свободы”), образует “базис изменчивости”. Обычно промеры зубов кударского медведя демонстрировали корреляцию только с координатой E1, формируя, таким образом, одномерный базис, ограниченный изменчивостью общих размеров зуба. Однако в ряде случаев в изменчивости отдельных структурных элементов зубов обнаруживаются независимые компоненты, расширяющие ее базис. Например, у m2 (табл. 2) наблюдаются независимые компоненты в изменчивости продольных размеров тригонида и талонида с лингвальной и лабиальной сторон зуба, и именно они ответственны за высокую размерность МР модели.
Размерность МФ моделей в большинстве случаев была равна трем, в одном случае – четырем (M1) и еще в одном – единице (p4) (табл. 2, 3). Координаты (К) демонстрируют высокие величины корреляции с индексами, отражающими изменчивость пропорций зубов. В случаях, когда К относительно сильно коррелируют (коэффициент корреляции больше 0.5) с абсолютными значениями промеров, речь может идти о высоком вкладе аллометрической изменчивости в варьирование пропорций зуба (m2, M2). Базис изменчивости формы зубов образуют индекс относительной ширины зуба (IW), некоторые индексы ширины/длины его элементов (индексы ILTALA у m2, ILPA у P4 и M1 и т.д.) и индексы, характеризующие отношения между элементами (например, индексы IWTR_TAL у m1 и ILTA_TR_LI у m2) (табл. 2, 3).
Основная информация о морфометрических различиях щечных зубов между разными группами содержится в варьировании координат МР и МФ моделей. На основании величины объясненной дисперсии (табл. 2, 3) можно сделать вывод, что эти различия не велики. Наибольшие величины дисперсии (> 30%), обусловленные межгрупповыми различиями, получены для координат E2 (m1, m2, P4) и K1 (m1, P4) некоторых зубов. Эти координаты коррелируют с общим индексом ширины зуба (IW) и относительными размерами некоторых его элементов.
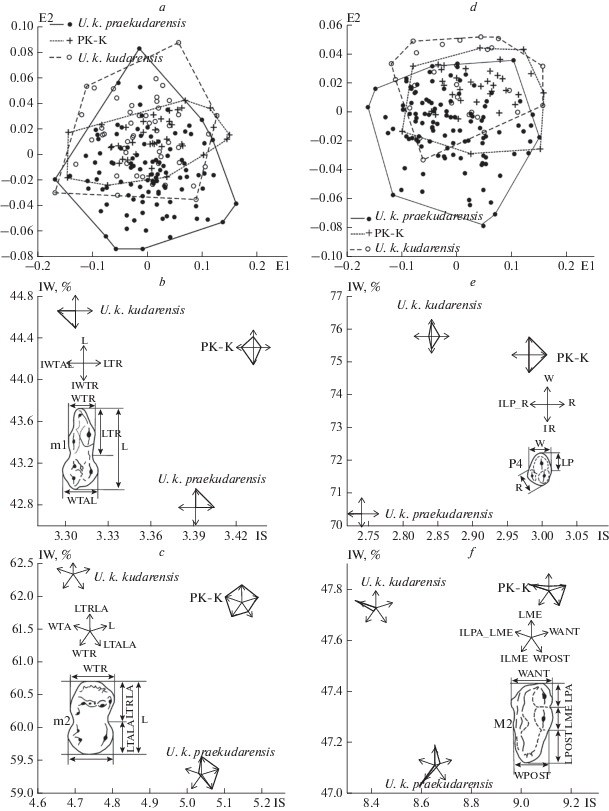
Внутригрупповая изменчивость размеров (координаты МР моделей) перекрывает межгрупповую, и в морфологическом пространстве (рис. 2a, 2b) разновременные группы образуют непрерывное облако точек. В полной мере это относиться к “промежуточной форме” и подвиду U. k. kudarensis.
Рис. 2.
МР модель морфопространества и эволюция m1, m2, P4 и M2. Морфологические пространства: a, d – положение экземпляров m1 и P4 U. k. praekudarensis, переходной формы (PK-K) и U. k. kudarensis в координатах МР модели; b, c, e, f – диаграммы, иллюстрирующие эволюцию m1, m2, P4 и M2 в координатах индекса жевательной поверхности (IS) и индекса относительной ширины зуба (IW); обозначения промеров приведены на рис. 1.
Средние значения промеров зубов для трех выборок кударских медведей приведены в табл. 4. Результаты одномерного анализа (тест Манна–Уитни) дают оценку статистической значимости различий между групповыми средними промеров и отдельных индексов зубов (табл. 5, 6). Результаты теста отражают существенную дифференциацию между средне и позднеплейстоценовыми выборками кударского медведя и значительную дифференциацию переходной формы от U. k. praekudarensis по некоторым промерам и индексам, но не по длине зубов (за исключением P4). Подвид U. k. kudarensis отличается от переходной формы в основном мелкими размерами зубов и средними значениями отдельных промеров m2.
Таблица 4.
Средние значения и стандартизованные ошибки промеров и двух индексов (IS, IS) щечных зубов разновозрастных групп кударских медведей U. k. praekudarensis (PK), “переходной формой” (PK/K) и U. k. kudarensis (K)
| Промеры | PK | PK/K | K | Измерения | PK | PK/K | K |
|---|---|---|---|---|---|---|---|
| P4 | M1 | ||||||
| L | 19.4 ± 0.12 | 19.4 ± 0.19 | 19.0 ± 0.26 | L | 26.1 ± 0.15 | 26.8 ± 0.22 | 26.3 ± 0.21 |
| LP | 12.0 ± 0.10 | 12.5 ± 0.14 | 12.0 ± 0.19 | LANT | 12.6 ± 0.08 | 12.9 ± 0.12 | 12.8 ± 0.10 |
| W | 13.6 ± 0.10 | 14.6 ± 0.17 | 14.4 ± 0.24 | LPOST | 13.3 ± 0.10 | 13.3 ± 0.14 | 13.2 ± 0.13 |
| R | 15.0 ± 0.12 | 16.4 ± 0.19 | 15.7 ± 0.28 | LPA | 9.7 ± 0.07 | 9.8 ± 0.08 | 9.8 ± 0.09 |
| IW | 70.4 ± 0.30 | 75.2 ± 0.45 | 75.8 ± 0.88 | LME | 8.9 ± 0.07 | 9.0 ± 0.10 | 8.9 ± .11 |
| IS | 2.70 ± 0.04 | 2.98 ± 0.06 | 2.80 ± 0.08 | W | 19.1 ± 0.13 | 19.1 ± 0.18 | 19.0 ± 0.16 |
| IW | 73.1 ± 0.26 | 71.2 ± 0.31 | 72.2 ± 0.48 | ||||
| IS | 5.0 ± 0.06 | 5.1 ± 0.09 | 5.0 ± 0.07 | ||||
| M2 | p4 | ||||||
| L | 43.5 ± 0.33 | 44.2 ± 0.43 | 42.5 ± 0.37 | L | 15.1 ± 0.11 | 14.9 ± 0.19 | 15.3 ± 0.14 |
| LPA | 12.8 ± 0.16 | 12.9 ± 0.18 | 12.7 ± 0.21 | W | 9.2 ± 0.08 | 9.4 ± 0.12 | 9.6 ± 0.10 |
| LME | 11.6 ± 0.17 | 10.9 ± 0.20 | 10.7 ± 0.21 | IW | 61.0 ± 0.43 | 63.1 ± 0.57 | 63.2 ± 0.61 |
| WANT | 21.9 ± 0.15 | 22.4 ± 0.19 | 21.7 ± 0.22 | IS | 1.40 ± 0.019 | 1.42 ± 0.03 | 1.47 ± 0.03 |
| WPOST | 19.0 ± 0.18 | 19.9 ± 0.24 | 18.9 ± 0.26 | ||||
| IW | 47.1 ± 0.31 | 47.9 ± 0.35 | 47.7 ± 0.36 | ||||
| IS | 8.6 ± 0.13 | 9.1 ± 0.16 | 8.4 ± 0.15 | ||||
| m1 | m2 | ||||||
| L | 28.5 ± 0.14 | 28.2 ± 0.21 | 27.6 ± 0.21 | L | 29.5 ± 0.17 | 29.5 ± 0.25 | 28.0 ± 0.16 |
| LTR | 18.1 ± 0.12 | 17.7 ± 0.16 | 17.4 ± 0.17 | LTRLA | 17.5 ± 0.14 | 17.5 ± 0.17 | 16.6 ± 0.13 |
| WTR | 10.9 ± 0.07 | 11.1 ± 0.13 | 10.9 ± 0.10 | LTRLI | 15.3 ± 0.11 | 14.8 ± 0.17 | 14.2 ± 0.14 |
| WTAL | 13.5 ± 0.07 | 13.9 ± 0.13 | 13.8 ± 0.11 | LTALA | 11.4 ± 0.09 | 11.2 ± 0.15 | 10.7 ± 0.11 |
| WMIN | 10.8 ± 0.07 | 11.2 ± 0.13 | 11.1 ± 0.11 | LTALI | 13.4 ± 0.12 | 13.1 ± .22 | 12.6 ± 0.15 |
| IW | 42.8 ± 0.18 | 44.3 ± 0.22 | 44.7 ± 0.26 | WTA | 18.2 ± 0.12 | 19.1 ± 0.19 | 18.1 ± 0.13 |
| IS | 3.4 ± 0.04 | 3.4 ± 0.06 | 3.3 ± 0.05 | WTR | 16.8 ± 0.11 | 17.7 ± 0.20 | 16.9 ± 0.11 |
| m3 | IW | 59.4 ± 0.21 | 62.4 ± 0.41 | 62.6 ± 0.32 | |||
| L | 25.2 ± 0.19 | 25.5 ± 0.34 | 25.1 ± 0.28 | IS | 5.0 ± 0.06 | 5.1 ± 0.10 | 4.7 ± 0.06 |
| LTAL | 10.7 ± 0.15 | 10.5 ± 0.25 | 10.8 ± 0.25 | ||||
| W | 18.4 ± 0.11 | 18.9 ± 0.19 | 18.6 ± 0.14 | ||||
| WTAL | 16.6 ± 0.15 | 17.4 ± 0.30 | 16.9 ± 0.22 | ||||
| IW | 73.2 ± 0.39 | 74.2 ± 0.69 | 74.0 ± 0.62 | ||||
| IS | 4.5 ± 0.07 | 4.7 ± 0.12 | 4.5 ± 0.09 | ||||
Таблица 5.
Статистическая значимость различий (p по тесту Манна-Уитни) средних значений промеров и индексов нижних щечных зубов между U. k. praekudarensis (PK), “переходной формой” (PK/K) и U. k. kudarensis (K)
| Переменная | PK–PK/K | PK–K | PK/K–K |
|---|---|---|---|
| p4 | |||
| W | n.s. | 0.005 | n.s. |
| IW | 0.01 | 0.01 | n.s. |
| m1 | |||
| L | n.s. | 0.0009 | 0.03 |
| LTR | n.s. | 0.0006 | n.s |
| WTAL | 0.004 | 0.01 | n.s. |
| WMIN | 0.007 | 0.04 | n.s. |
| IW | <0.0001 | <0.0001 | n.s. |
| ILTR | 0.02 | 0.04 | n.s. |
| ILTA | 0.02 | 0.04 | n.s. |
| IWTR | 0.0006 | 0.004 | n.s. |
| IWTAL | <0.0001 | <0.0001 | n.s. |
| IWTR_TAL | n.s. | 0.001 | 0.04 |
| m2 | |||
| L | n.s. | <0.0001 | <0.0001 |
| LTRLA | n.s. | 0.0004 | 0.003 |
| LTRLI | 0.03 | <0.0001 | 0.02 |
| LTALA | n.s. | <0.0001 | 0.04 |
| LTALI | n.s. | <0.0001 | n.s. |
| WTR | 0.0004 | n.s. | 0.006 |
| WTA | <0.0001 | n.s. | 0.001 |
| IW | <0.0001 | <0.0001 | n.s. |
| IS | n.s. | 0.002 | 0.0007 |
| m3 | |||
| W | 0.03 | n.s. | n.s. |
| WTAL | 0.01 | n.s. | n.s. |
| ILTAL | 0.03 | n.s. | n.s. |
Таблица 6.
Статистическая значимость различий (p по тесту Манна–Уитни) средних значений промеров и индексов верхних щечных зубов между U. k. praekudarensis (PK), “переходной формой” (PK/K) и U. k. kudarensis (K) (приведены промеры и индексы, по которым различается хотя бы одна пара групп при p < 0.05)
| Переменная | PK–PK/K | PK–K | PK/K–K |
|---|---|---|---|
| P4 | |||
| LP | 0.009 | n.s. | n.s. |
| W | <0.001 | 0.003 | n.s. |
| R | <0.001 | 0.03 | 0.04 |
| IW | <0.001 | <0.001 | n.s. |
| ILP | <0.001 | n.s. | n.s. |
| IR | <0.001 | <0.001 | n.s. |
| ILP_R | <0.001 | 0.003 | n.s. |
| ILPOST | <0.001 | n.s. | n.s. |
| IS | <0.001 | n.s. | n.s. |
| M1 | |||
| L | 0.008 | n.s. | n.s. |
| IW | <0.001 | n.s. | n.s. |
| ILPOST | 0.01 | n.s. | 0.04 |
| ILANT_LPOST | 0.03 | n.s. | 0.03 |
| M2 | |||
| L | n.s. | n.s. | 0.01 |
| LME | n.s. | 0.005 | n.s. |
| WANT | 0.03 | n.s. | 0.02 |
| WPOST | 0.008 | n.s. | 0.007 |
| ILME | 0.005 | 0.04 | n.s. |
| ILPA_LME | 0.04 | 0.008 | n.s. |
| ILPOST | 0.006 | n.s. | n.s. |
| IS | 0.02 | n.s. | 0.005 |
Соотношение различий в размерах зубов и их пропорций показано на рис. 2b, 2c, 2e и 2 f. Во всех случаях размеры щечных зубов у предполагаемого предка U. k. praekudarensis были промежуточными между более крупными зубами переходной формы и более мелкими зубами U. k. kudarensis. При этом индекс ширины зубов у них был в среднем ниже по сравнению с переходной формой.
Параметры морфологических ниш U. k. praekudarensis, “промежуточной формы” и U. k. kudarensis в МР и МФ морфопространствах представлены в табл. 7. В среднем в МР моделях для верхних и нижних зубных рядов наибольшие значения индекса si обнаружены у U. k. praekudarensis (см. также рис. 2a, 2d). При этом максимальные значения индекса характерны для верхнего ряда щечных зубов. Относительная ширина морфологической ниши U. k. kudarensis меньше, чем у среднеплейстоценового предкового таксона. Верхний ряд у этого подвида характеризуется минимальным разнообразием размеров зубов. Значения si у переходной формы в среднем ближе к таковым U. k. kudarensis. В нижнем зубном ряду среднее значение индекса (все хроно-группы) изменяется в последовательности m3 > p4 > m1 > m2, а в верхнем ряду – M1 > M2 > P4.
Таблица 7.
Ширина морфологической ниши (Hi, бит), доля от общего объема МР или МФ морфопространств (si) и индекс пересечения ниш $O_{{jk}}^{R}$ U. k. praekudarensis (PK), переходной формы формы (PK/K) и U. k. kudarensis (K)
| Зуб | MР | МФ | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hi/si | $O_{{jk}}^{R}$ | Hi/si | $O_{{jk}}^{R}$ | |||||||||
| PK | PK/K | K | PK–PK/K | PK–K | PK/K–K | PK | PK/K | K | PK–PK/K | PK–K | PK/K–K | |
| p4 | 1.95 | 2.63 | 2.69 | 0.894 | 0.912 | 0.949 | 0.99 | 0.96 | 0.99 | 0.944 | 0.978 | 0.992 |
| 0.57 | 0.92 | 0.96 | 0.99 | 0.98 | 0.99 | |||||||
| m1 | 5.46 | 4.78 | 5.40 | 0.628 | 0.695 | 0.844 | 8.77 | 7.08 | 8.24 | 0.326 | 0.340 | 0.621 |
| 0.90 | 0.56 | 0.86 | 0.99 | 0.31 | 0.68 | |||||||
| m2 | 10. 8 | 10.0 | 10. 1 | 0.844 | 0.490 | 0.461 | 8.85 | 8.84 | 9.14 | 0.371 | 0.550 | 0.644 |
| 0.82 | 0.50 | 0.52 | 0.73 | 0.72 | 0.89 | |||||||
| m3 | 5.34 | 5.54 | 5.07 | 0.840 | 0.783 | 0.800 | 7.61 | 7.14 | 8.11 | 0.740 | 0.861 | 0.718 |
| 0.94 | 1.0 | 0.76 | 0.83 | 0.60 | 1.0 | |||||||
| P4 | 5.32 | 4.58 | 5.03 | 0.517 | 0.635 | 0.720 | 7.37 | 7.09 | 7.93 | 0.260 | 0.421 | 0.617 |
| 0.91 | 0.54 | 0.75 | 0.68 | 0.56 | 1.0 | |||||||
| M1 | 5.18 | 5.20 | 4.66 | 0.892 | 0.951 | 0.814 | 12.23 | 12.13 | 11. 9 | 0.711 | 0.598 | 0.609 |
| 0.99 | 0.99 | 0.69 | 0.92 | 0.85 | 0.74 | |||||||
| M2 | 8.06 | 7.79 | 7.87 | 0.567 | 0.739 | 0.573 | 8.55 | 8.41 | 8.40 | 0.570 | 0.724 | 0.646 |
| 0.86 | 0.72 | 0.76 | 0.83 | 0.76 | 0.75 | |||||||
В МФ морфопространстве высокие значения индекса si типичны как для U. k. praekudarensis, так и для U. k. kudarensis, а разнообразие пропорций зубов переходной формы в этом случае ниже. В нижнем ряду p4 демонстрирует максимальное разнообразие пропорций, а коренные зубы формируют ряд – m3 > m2 > m1. В верхнем ряду разнообразие пропорций M1 выше, чем M2, а P4 (в противоположность p4) характеризуется относительно низкими значениями si.
Средняя величина пересечения морфологических ниш носит противоположную направленность у нижних и верхних зубов. В случае МР моделей зубов нижнего ряда максимальные значения $O_{{jk}}^{R}$ (низкий уровень морфологической дифференциации по размерам зубов) обнаружены в паре U. k. praekudarensis – переходная форма, а минимальные – между U. k. praekudarensis и U. k. kudarensis. В верхнем ряду ситуация носит противоположный характер. В общем, для нижних зубов значения $O_{{jk}}^{R}$ образуют ряд p4 > m3 > > m2 > m1, а для верхних – M1 > M2 ≈ P4.
В моделях МФ пересечение морфологических ниш верхних зубов было выше, чем у нижних. Минимальные значения индекса (максимальная морфологическая дифференциация по пропорциям зубов) отмечены для пары U. k. praekudarensis – переходная форма, а максимальные – для пары переходная форма – U. k. kudarensis. В среднем для нижних зубов значения $O_{{jk}}^{R}$ образуют ряд p4 > m3 > m2 > m1, а для верхних – M1 ≈ M2 > P4.
Величины индекса $O_{{jk}}^{R}$ вместе с координатами центроидов групп медведей с разным геологическим возрастом дают взаимодополняющие друг друга оценки морфологического сходства/различия между ними. На рис. 3 представлена серия дендрограмм, построенных для каждого зуба на основе значений групповых центроидов в МР и МФ морфопространствах (рис. 3a) и соответствующих коэффициентам $O_{{jk}}^{R}$ (рис. 3b). По относительному положению центроидов, переходная форма демонстрирует близость к позднеплейстоценовому U. k. kudarensis, что согласуется с результатами одномерного анализа (табл. 5, 6). Однако по признакам m3 переходная форма занимает в целом обособленное положение относительно обоих подвидов кударского медведя.
Рис. 3.
Дендрограммы, иллюстрирующие морфологическую дифференциацию U. k. praekudarensis (PK), переходной формы (PK-K) и U. k. kudarensis (K): a – на основе взаимоположения групповых центроидов в морфологических пространствах, b – по коэффициентам пересечения морфологических ниш в МР и МФ моделях, c – по величине коэффициентов MAC верхнего зубного ряда, d – по коэффициентам MAC нижнего зубного ряда. Цифры обозначают величину бутстреп-поддержки ветвления дендрограммы в % (100 повторов).
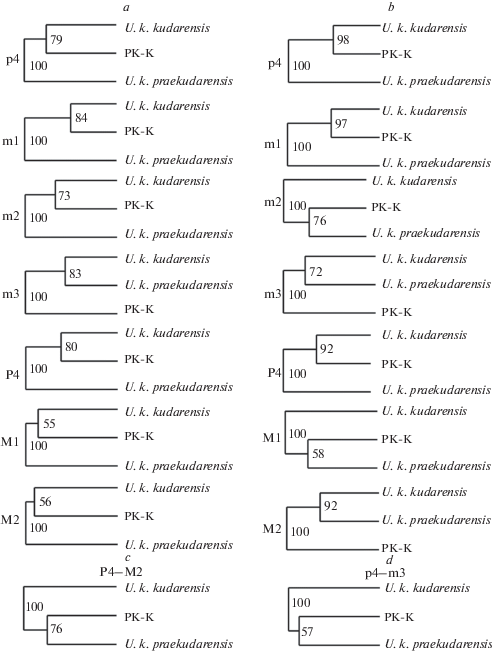
Оценка сходства групп по пересечению морфологических ниш дает менее четкую картину (рис. 3b), что обусловлено в основном разным масштабом внутригрупповой изменчивости (разнообразия) отдельных зубов, а не только относительным положением соответствующих центроидов. В этом случаем меньше половины дендрограмм (p4, P4, m1) демонстрируют сходство переходной формы с U. k. kudarensis. В остальных случаях она оказывается ближе к U. k. praekudarensis (m2, M1) или занимает обособленное положение (m3, M2).
Аллометрическая изменчивость. Аллометрические коэффициенты (MAC) отдельных промеров зубов показывают с какой относительной скоростью они меняются при увеличении/уменьшении общих размеров зуба (табл. 8). В целом, по величинам MAC всех зубов переходная форма оказалась несколько ближе к U. k. praekudarensis, чем к U. k. kudarensis, особенно по аллометрии верхних щечных зубов (рис. 3c, 3d). На этом фоне отдельные зубы демонстрируют специфические аллометрические паттерны прежде всего у переходной формы.
Таблица 8.
Аллометрические коэффициенты (MAC) и их 95% доверительные интервалы (INT) промеров верхних (P4-M2) и нижних щечных (p4-m3) зубов
| Переменная | U. k. praekudarensis | Переходная форма | U. k. kudarensis | |||
|---|---|---|---|---|---|---|
| MAC | INT | MAC | INT | MAC | INT | |
| P4 | ||||||
| L | 0.84 | 0.83–0.85 | 0.91 | 0.90–0.92 | 0.76 | 0.74–0.78 |
| LP | 1.06 | 1.04–1.07 | 0.93 | 0.91–0.96 | 0.99 | 0.97–1.01 |
| W | 0.98 | 0.97–0.99 | 1.10 | 1.09–1.11 | 1.05 | 1.04–1.07 |
| R | 1.10 | 1.09–1.11 | 1.04 | 1.02–1.06 | 1.14 | 1.13–1.16 |
| M1 | ||||||
| L | 0.92 | 0.91–0.93 | 0.95 | 0.93–0.96 | 1.05 | 1.01–1.08 |
| LANT | 0.92 | 0.90–0.93 | 1.02 | 1.01–1.03 | 0.95 | 0.90–1.01 |
| LPOST | 0.96 | 0.94–0.97 | 1.14 | 1.12–1.15 | 0.96 | 0.91–1.02 |
| LPA | 1.05 | 1.04–1.06 | 0.77 | 0.74–0.79 | 0.92 | 0.86–0.97 |
| LME | 1.07 | 1.04–1.10 | 1.03 | 1.00–1.06 | 0.89 | 0.76–1.02 |
| W | 1.06 | 1.05–1.07 | 1.03 | 1.01–1.05 | 0.84 | 0.80–0.88 |
| M2 | ||||||
| L | 0.58 | 0.53–0.64 | 0.59 | 0.54–0.65 | 0.22 | 0.18–0.25 |
| LPA | 0.73 | 0.59–087 | 0.69 | 0.61–0.77 | 1.06 | 0.94–1.18 |
| LME | 1.53 | 1.37–1.69 | 1.68 | 1.53–1.83 | 1.51 | 1.32–1,70 |
| WANT | 0.46 | 0.40–0.51 | 0.53 | 049–0.57 | 0.27 | 0.23–0.31 |
| WPOST | 0.42 | 0.34–0.49 | 0.07 | –0.03–0.16 | –0.01 | –0.08–0.07 |
| p4 | ||||||
| L | 0.94 | 0.93–0.95 | 1.04 | 1.02–1.06 | 0.88 | 0.86–0.90 |
| W | 1.06 | 1.05–1.07 | 0.96 | 0.94–0.97 | 1.10 | 1.08–1.11 |
| m1 | ||||||
| L | 0.87 | 0.86–0.88 | 0.75 | 0.74–0.76 | 0.88 | 0.86–90 |
| LTR | 1.01 | 1.00–1.03 | 0.86 | 0.85–0.88 | 1.05 | 1.02–1.07 |
| WTR | 1.05 | 1.04–1.06 | 1.20 | 1.19–1.21 | 1.05 | 1.03–1.07 |
| WTAL | 0.96 | 0.95–0.97 | 0.93 | 0.92–0.94 | 0.90 | 0.88–0.91 |
| WMIN | 1.09 | 1.08–1.10 | 1.17 | 1.16–1.19 | 1.08 | 1.06–1.11 |
| m2 | ||||||
| L | 0.95 | 0.94–0.96 | 0.87 | 0.87–0.88 | 0.86 | 0.85–087 |
| LTRLA | 1.13 | 1.12–1.14 | 0.82 | 0.80–0.84 | 0.84 | 0.81–0.86 |
| LTRLI | 0.83 | 0.81–0.86 | 0.92 | 0.89–0.95 | 0.97 | 0.93–1.02 |
| LTALA | 0.89 | 0.87–0.90 | 1.04 | 1.02–1.06 | 1.24 | 1.22–1.26 |
| LTALI | 1.26 | 1.24–1.26 | 1.31 | 1.28–1.34 | 1.24 | 1.18–1.30 |
| WTR | 0.89 | 0.87–0.90 | 0.93 | 0.90–0.96 | 0.69 | 0.67–0.71 |
| WTA | 0.97 | 0.96–0.98 | 0.98 | 0.96–1.00 | 0.93 | 0.90–0.95 |
| m3 | ||||||
| L | 0.80 | 0.79–0.81 | 0.79 | 0.77–0.81 | 0.73 | 0.72–0.75 |
| LTAL | 1.54 | 1.53–1.55 | 1.45 | 1.43–1.47 | 1.63 | 1.62–1.64 |
| WTAL | 0.84 | 0.83–0.86 | 0.99 | 0.97–1.0 | 0.81 | 0.50–0.82 |
| W | 0.51 | 0.50–0.52 | 0.54 | 0.52–0.55 | 0.37 | 0.36–0.38 |
Паттерн P4 переходной формы отличается слабой отрицательной аллометрией LP и положительной аллометрией R. При этом ширина P4 увеличивается быстрее длины. Соответствующие паттерны у U. k. praekudarensis и U. k. kudarensis характеризуются тенденцией к относительному увеличению длины паракона (LP) и протокона (R) (положительная аллометрия) на фоне существенной отрицательной аллометрии длины коронки зуба. Аллометрическая изменчивость p4 переходной формы близка к изометрии, в отличие от паттернов U. k. praekudarensis и U. k. kudarensis. У последних длина коронки p4 характеризуется слабой отрицательной аллометрией, а ширина – положительной. Тем не менее индекс ширины зуба (табл. 4) переходной формы практически не отличается от индекса U. k. kudarensis и статистически значимо выше, чем у U. k. praekudarensis. Это объясняется тем, что длина p4 переходной формы несколько меньше, чем у обоих подвидов, а ширина имеет промежуточное значение.
Аллометрические паттерны M1 U. k. praekudarensis и U. k. kudarensis различаются по промерам LPA, LME и W. В первом случае имеет место положительная аллометрия, во втором – отрицательная (табл. 8). Однако это не приводит к статистическим различиям между ними в размерах и основных пропорциях зуба (табл. 5). Паттерн переходной формы отличается сильной отрицательной аллометрией длины паракона и положительной аллометрией длины задней части коронки (LPOST). Паттерны нижнего m1 у U. k. praekudarensis и U. k. kudarensis практически одинаковы и существенно отличаются от аллометрии зуба у переходной формы. Последняя характеризуется сильной отрицательной аллометрией промеров L, LTR и сильной положительной аллометрией ширины тригонида (WTR) и минимальной ширины зуба (WMIN).
Отсутствие аллометрической изменчивости ширины зуба (WANT, WPOST) является особенностью паттерна M2 переходной формы и U. k. kudarensis. В отличие от других зубов для M2 характерны экстремальные значения коэффициентов как положительной (LME), так и отрицательной (L, WANT) аллометрии (табл. 8). В целом аллометрические параметры M2 переходной формы ближе к U. k. praekudarensis. Паттерн нижнего m2 у переходной формы близок к паттерну U. k. kudarensis. Для них характерна существенная отрицательная аллометрия наибольшей длины коронки (L) и положительная аллометрия длины талонида с лабиальной стороны (LTALA), особенно у U. k. kudarensis. У U. k. praekudarensis выражена положительная аллометрия длины тригонида с лабиальной стороны (LTALA), отрицательная – длина с лингвальной стороны (LTRLI). У талонида отмечена противоположная ситуация: отрицательная аллометрия его длины с лабиальной стороны (LTALA) и положительная – с лингвальной стороны зуба (LTALI). Положительная аллометрия промера LTALA наблюдается у переходной формы и у U. k. kudarensis.
Аллометрические паттерны m3 у всех кударских медведей очень сходны и характеризуются сильной отрицательной аллометрией ширины зуба (W) и положительной аллометрией длины талонида (LTALD).
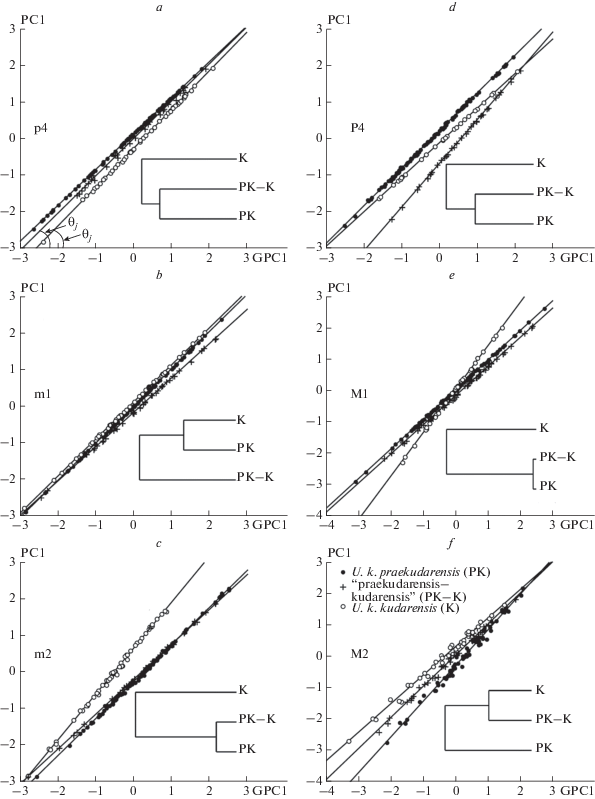
Еще одно представление аллометрических паттернов в форме регрессии частных, групповых главных компонент (PC1) относительно общей главной компоненты (GPC1) приведено на рис. 4. Линии регрессии показывают с какой “скоростью” изменяется общий размер зуба у U. k. praekudarensis, переходной формы и U. k. kudarensis при изменении общих размеров зуба, “генерализованнного” для всех трех таксонов. Различия между PC1 связаны с величинами соответствующих им углов ϴ между линиями регрессий и координатой GPC1. По величине угла ϴ большинства верхних и нижних зубов U. k. praekudarensis и переходная форма демонстрируют подобие аллометрических паттернов, что соответствует общему выводу из анализа MAC. Паттерн переходной формы сближается с таковым U. k. kudarensis в случае M2.
Рис. 4.
Аллометрические паттерны щечных зубов: a–e – аллометрические паттерны p4, m1, m2, P4, M1 и M2 U. k. praekudarensis (PK), переходной формы (PK-K) и U. k. kudarensis (K) и UPGMA дендрограммы, иллюстрирующие их различия по величине угла ϴ между линиями регрессии PC1 и координатой GPC1; объяснения в тексте.
Оценка скорости эволюции. Хотя и с разрывами, выборки зубов кударского медведя из слоев пещер Кударо 1 и Кударо 3 стратиграфически сменяют друг друга, образуя ряд U. k. praekudarensis – переходная форма – U. k. kudarensis (рис. 5). Имеющиеся датировки позволяют приблизительно оценить время существования популяций кударских медведей, остатки которых сохранились в пещерных отложениях, и охарактеризовать темпы преобразования зубного аппарата от середины среднего плейстоцена до позднего плейстоцена. Выше было показано, что основные тренды преобразования параметров щечных зубов в среднем и позднем плейстоцене обусловлены изменениями общих зубных размеров и увеличением относительной ширины коронки. В табл. 9 приведены расчетные величины скорости изменений (h) отдельных промеров и индексов, характеризующих оба тренда.
Рис. 5.
Предполагаемое положение выборок U. k. praekudarensis, переходной формы (PK-K) и U. k. kudarensis (показано стрелками) из пещер Кударо 1 (Kudaro 1) и Кударо 3 (Kudaro 3) относительно временной и кислородно-изотопной шкал (Lisiecki, Raymo, 2005). Высота расположения пещер указана в метрах над уровнем моря (m a. s. l.). t1–t2 (Несмеянов, 1999), t2–t3 – интервалы для расчета скорости эволюции, h (табл. 9).
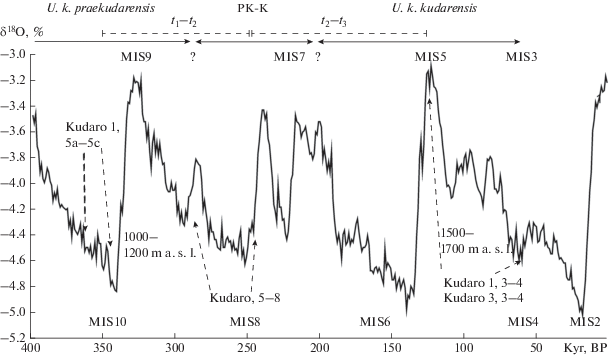
Таблица 9.
Скорость эволюционных изменений (h × 10 тыс. лет) некоторых промеров и индексов щечных зубов в ряду U. k. praekudarensis – переходная форма (MIS 10–MIS 7) и переходная форма – U. k. kudarensis (MIS 7–MIS 3)
| Промер или индекс | MIS 10–MIS 8 | MIS 7–MIS 5 | Промер или индекс | MIS 10–MIS 8 | MIS 7–MIS 5 |
|---|---|---|---|---|---|
| t1– t2 ~ 110 тыс. лет | t2–t3 ~ 130 тыс. лет | t1– t2 ~ 110 тыс. лет | t2 – t3 ~ 130 тыс. лет | ||
| p4 | m1 | ||||
| L | 0.012 | 0.019 | L | 0.021 | 0.030 |
| W | 0.030 | 0.018 | WTAL | 0.049 | 0.008 |
| IW | 0.046 | 0.001 | IW | 0.076 | 0.016 |
| IS | 0.011 | 0.020 | IS | 0.010 | 0.027 |
| m2 | m3 | ||||
| L | 0.006 | 0.068 | L | 0.012 | 0.004 |
| WTAL | 0.052 | 0.057 | WTAL | 0.042 | 0.022 |
| IW | 0.097 | 0.013 | IW | 0.015 | 0.007 |
| IS | 0.016 | 0.061 | IS | 0.026 | 0.014 |
| P4 | M1 | ||||
| L | 0.010 | 0.027 | L | 0.033 | 0.022 |
| W | 0.083 | 0.016 | W | 0.005 | 0.004 |
| IW | 0.120 | 0.012 | IW | 0.064 | 0.028 |
| IS | 0.056 | 0.027 | IS | 0.014 | 0.016 |
| M2 | |||||
| L | 0.024 | 0.046 | |||
| WPOST | 0.047 | 0.042 | |||
| IW | 0.027 | 0.002 | |||
| IS | 0.041 | 0.051 |
Оба премоляра p4 и P4 характеризуются асинхронностью изменений по ширине и длине. В среднем плейстоцене (интервал t1 – t2, рис. 5) изменение ширины коронки было существенно выше по сравнению с последующим периодом (интервал t2 – t3): 0.03 h/10 тыс. лет – 0.018 h/10 тыс. лет (p4) и 0.083 h/10 тыс. лет – 0.016 h/10 тыс. лет (P4). При этом у P4 скорость эволюции была наибольшей по сравнению с остальными щечными зубами и примерно в 2.8 раза выше, чем у p4 (рис. 6a). Вследствие этого уже на уровне переходной формы значения индекса IW и основных параметров премоляров достигли значений, характерных для позднеплейстоценового подвида U. k. kudarensis, а скорость изменения относительной ширины зубов снизилась в 46 (p4) и 10 (P4) раз. На интервале от конца среднего плейстоцена до начала позднего плейстоцена (интервал t2–t3) скорости изменений p4 и P4 по ширине коронки резко снизились, но при этом скорость изменения длины коронки возросла в 1.6 раза (p4) – 1.9 раза (P4). Отметим, что изменения общих размеров p4 и P4 в конце среднего плейстоцена были разнонаправлены – первый несколько увеличился, а второй – уменьшился (табл. 4). На диаграмме первых главных компонент (рис. 6d) для скоростей эволюции L, W, IW и IS (использована ковариационная матрица значений h промеров и индексов p4, P4 для двух интервалов времени) основные эволюционные тренды премоляров выражены относительно PC1.
Рис. 6.
Характеристика скоростей эволюции щечных зубов: a – c: медиана, минимум и максимум скоростей эволюции в халденах общих размеров (S) и ширины (W) щечных зубов (p4, m1–m3, P4, M1, M2). d – f: эволюционные паттерны общих размеров (промеры L, индекс IS) и ширины (промеры W, WPOST, индекс IW) щечных зубов (p4, m1–m3, P4, M1, M2), полученные методом главных компонент на ковариационной матрице скоростей эволюции (h) на интервалах t1–t2, t2–t3 (табл. 9, рис. 5). Стрелками обозначены направления и масштаб изменений паттерна скоростей эволюции.
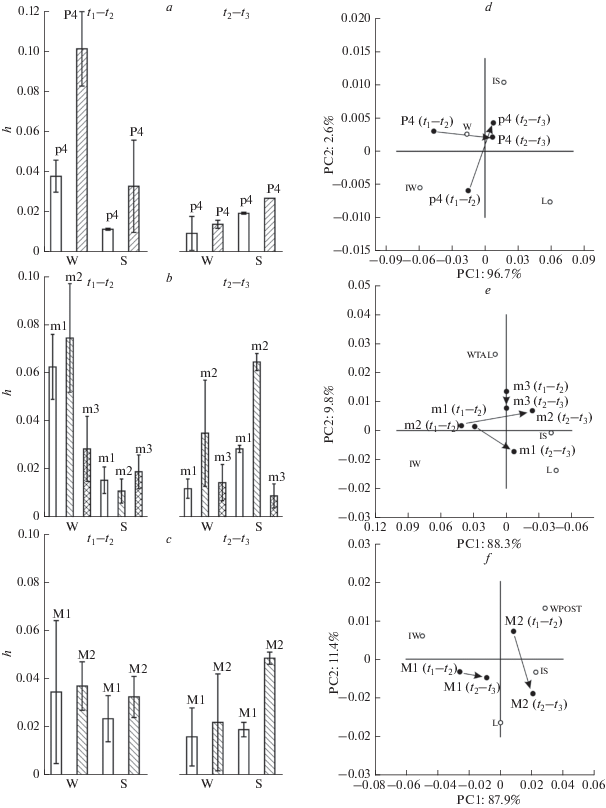
Эволюционные изменения скорости преобразований в целом аналогичные описанным для премоляров обнаружены и у моляров m1 и m2 (рис. 6b, 6e). На интервале t1-t2 изменения ширины коронки у обоих зубов существенно превышали по скорости изменения длины и общих размеров (табл. 9). При этом скорость изменений у m2 была выше, чем у m1. На интервале t2 – t3 наблюдается падение скорости изменений ширины и рост скорости изменения длины и общих размеров зубов. В среднем размеры зубов, особенно m2, у U. k. kudarensis уменьшились по сравнению с U. k. praekudarensis и “переходной формой” (табл. 4). В отличие от m1 и m2, m3 помимо более низких скоростей изменений ширины и общих размеров отличается синхронностью их падения в конце среднего плейстоцена. Отсутствие статистически значимых различий между “переходной формой” и U. k. kudarensis (табл. 5) и устойчивость аллометрического паттерна (табл. 8) подтверждают низкую скорость эволюции m3, что также иллюстрирует рис. 6e.
Скорости изменений моляров M1 и M2 демонстрируют специфические особенности. У M1 изменение ширины зуба практически отсутствует (табл. 4), соответственно оба интервала характеризуются несущественными значениями h (табл. 8). Ширина талона (WPOST) M2 на интервале t1–t2 изменяется с типичной для моляров скоростью (0.042 h/10 тыс. лет), но в отличие от других зубов она падает в конце среднего плейстоцена. На первом этапе средние значения промера растут (табл. 4), а на втором – снижаются примерно с той же скоростью. При этом скорость изменений индекса ширины обоих верхних моляров изменяется также, как и у остальных зубов. На интервале t2–t3 у M1 скорость изменений индекса падает в 2.3 раза, а у M2 – в 13.5 раз. В отличие от всех других зубов, M1 характеризуется высокой скоростью изменений длины коронки (0.033 h/10 тыс. лет) в среднем плейстоцене, которая не возрастает, а снижается в течение второго временного интервала. Длина коронки M1 достигает своих максимальных значений у переходной формы, а затем уменьшается, возвращаясь у U. k. kudarensis к значениям, близким к U. k. praekudarensis. Скорость изменений длины M2 почти в 2 раза выше на втором интервале при той же схеме изменения средних значений L (табл. 4). Статистически значимые изменения параметров верхних моляров были обнаружены (табл. 6) между U. k. praekudarensis и “переходной формой”, между “переходной формой” и U. k. kudarensis. При этом M1 у подвида U. k. praekudarensis имеет размеры и пропорции такие же как у позднеплейстоценового подвида U. k. kudarensis, а по признакам M2 таксоны несколько различаются только по длине метакона. Таким образом, изменения M1 и M2 не характеризуются упорядоченным трендом изменений скоростей эволюции в среднем – позднем плейстоцене (рис. 6f), который выражен у нижних m1 и m2 (рис. 6e).
Согласованность изменчивости щечных зубов. В этом разделе приведены результаты исследования согласованности изменчивости длины коронок щечных зубов верхнего и нижнего рядов. Промер L демонстрирует высокую корреляцию с первой координатой МР моделей (табл. 2 и 3) и, следовательно, его варьирование отражает изменчивость общих размеров зубов.
Методом главных компонент были построены три модели, описывающие изменчивость длины зубов у U. k. praekudarensis (рис. 7a), переходной формы (рис. 7b) и у U. k. kudarensis (рис. 7c). Расчеты велись независимо для каждой выборки на основе ковариационной матрицы логарифмированных медиан промера L для разновозрастных подвыборок из разных слоев пещер Кударо 1 и Кударо 3. Из представленных графиков следует, что отношения между размерами зубов более чем на 95% определяются положением соответствующих точек относительно PC1. Во всех трех выборках кударского медведя наблюдается близкая схема ковариации щечных зубов, в которой на первый план выходят функциональные отношения между верхними (M1, M2) и нижними (m1, m2) молярами, что отражено в положении этих зубов на соответствующих UPGMA дендрограммах (рис. 7). Изменчивость длины m3, видимо, существенно независима от изменений как верхних, так и нижних коренных.
Рис. 7.
Ковариация размеров верхних и нижних щечных зубов и проверка гипотезы о взаимном ограничении изменчивости длины нижних коренных зубов, находящихся в одном ряду: a–c: отношения верхних и нижних щечных зубов в пространстве первых главных компонент (PC1, PC2) у U. k. praekudarensis (a), переходной формы (b) и U. k. kudarensis (c); UPGMA дендрограммы построены по значениям координат точек в пространстве главных компонент; 1 – “сильные” связи между зубами при уровне бутстреп-поддержки на дендрограммы < 55% (100 повторов), 2 – “слабые” связи между зубами при уровне бутстреп–поддержки > 55% (100 повторов); приведена схема окклюзии зубов пещерного медведя; d–f : проверка гипотез о влиянии на длину зуба его относительного положения в ряду нижних (d, e) и верхних (f) щечных зубов в выборках U. k. praekudarensis (pk), переходной формы (pk–k) и U. k. kudarensis (k).
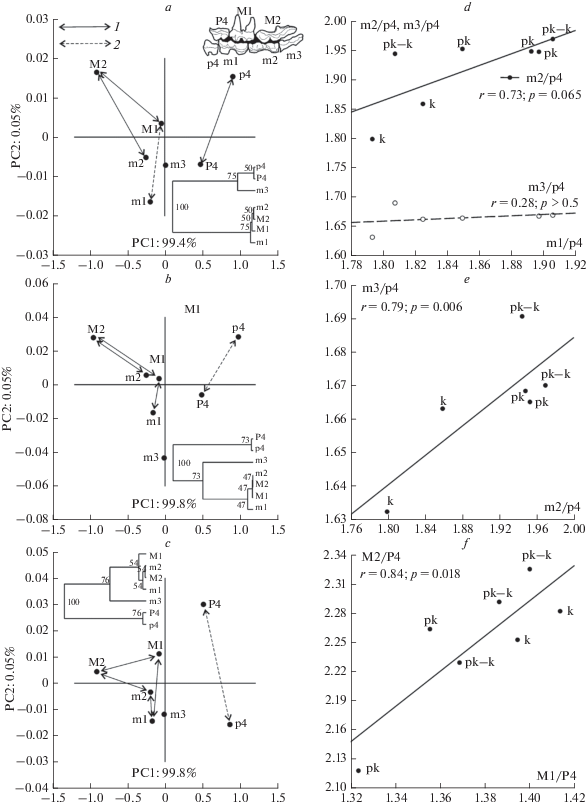
Премоляры p4 и P4 формируют самостоятельный кластер на дендрограммах, демонстрируя относительную независимость изменчивости не только от моляров, но и друг от друга (переходная форма и U. k. kudarensis).
В целом описанные отношения соответствуют схеме окклюзии верхних и нижних зубов у пещерных медведей, приведенной на рис. 7a. Хотя исходных данных (число разновозрастных подвыборок внутри U. k. praekudarensis, переходной формы и U. k. kudarensis) недостаточно для статистически обоснованных заключений, мы не исключаем более высокую согласованность изменчивости размеров m1, m2 и M1, M2 у позднеплейстоценовых U. k. kudarensis по сравнению со среднеплейстоценовыми U. k. praekudarensis.
Далее, мы проверили гипотезу (Kavanagh et al., 2007; Renvoisé et al., 2009; Jernvall, Thesleff, 2012; Asahara et al., 2016) о взаимном ограничении изменчивости длины нижних коренных зубов, находящихся в одном ряду (inhibitory developmental cascade model). Гипотеза основывается на экспериментальных данных, полученных для нижнего зубного ряда грызунов (мышей), и объясняет различия в линейных размерах зуба в зависимости от его положения в ряду относительно m1 как результат ограничений, возникающих в процессе регулирования закладки и роста зубов в онтогенезе (Salazar-Ciudad, Jernvall, 2002). Согласно гипотезе, m1 ингибирует рост m2, а m2 в свою очередь ингибирует рост m3 с “силой”, пропорциональной размеру зубов. Другими словами, чем сильнее m1 влияет на m2, тем слабее m2 влияет на конечные размеры m3 и наоборот. Следовательно, должна существовать положительная регрессия между относительными размерами зубов, которая описывается формулой m2/m1 = a + b(m3/m1). Такая зависимость была подтверждена для многих таксономических групп млекопитающих, но для представителей отряда хищных наклон линии регрессии (определяется параметром b) оказался существенно ниже, чем в большинстве остальных таксонов (Asahara et al., 2016). Проверка гипотезы на наших данных показала отсутствие значимой связи между m2/m1 и m3/m1 (b = 0.25, r = 0.25, p = = 0.5). Связь между линейными размерами зубов обнаружилась только после того, как в модель включили данные по длине p4 (рис. 7d, 7e). Для отношений m1/p4 и m2/p4 была получена искомая регрессия, в которой коэффициент b был приблизительно равен 1 (0.998). Одновременно отсутствовала связь между m1/p4 и m3/p4, что практически исключает влияние размеров p4 на размеры m3 (рис. 8a ). В тоже время существенно более слабая регрессия (b = 0.22, r = 0.79) обнаружена между m2/p4 и m3/p4 (рис. 7e). Этот результат предполагает онтогенетическое влияние m2 на m3.
Так как онтогенез верхних зубов медведей должен регулироваться на основе тех же молекулярно-генетических механизмов, то мы посчитали целесообразным протестировать аналогичной гипотезой верхний зубной ряд. Результаты теста (рис. 7f) подтверждают возможность ограничений варьирования размеров зубов в соответствии с гипотезой ингибирования в рострально-каудальном направлении.
В целом наши результаты указывают на ведущую роль функционального взаимодействия верхнего и нижнего рядов при формировании абсолютных значений размерных характеристик зубов у взрослых кударских медведях. На этом фоне связи между зубами внутри рядов могут быть обусловлены генетико-онтогенетическими регуляторными процессами на стадии формирования и роста коренных зубов.
ОБСУЖДЕНИЕ
Наши результаты не показали единого, “линейного” пути эволюционного преобразования щечных зубов кударского медведя (U. kudarensis), собранных из рыхлых отложений пещер Кударо 1 и Кударо 3. Своеобразием изменчивости во времени характеризуются не только зубы верхнего и нижнего зубного ряда в целом, но и премоляры и моляры и даже отдельные моляры (например, m3). Различия средних значений промеров между U. k. praekudarensis, U. k. kudarensis и переходной формой (табл. 5) выявлены на фоне очень высокой индивидуальной изменчивости (рис. 2a, 2d). При этом переходная форма по многим параметрам, характеризующим размеры зубов, оказалась крупнее и U. k. praekudarensis, и U. k. kudarensis.
Одномерный и многомерный статистический анализ зубных параметров выявил более многочисленные и статистически значимые различия между среднеплейстоценовым подвидом U. k. praekudarensis и переходной формой, чем между переходной формой и позднеплейстоценовым подвидом U. k. kudarensis. В то же время за счет различий в масштабах внутригрупповой изменчивости положение выборки переходной формы в морфологических пространствах относительно выборок двух подвидов не постоянно и различно для разных зубов.
Анализ аллометрической изменчивости отдельных зубов показал близость соответствующих паттернов у U. k. praekudarensis и переходной формы. В некоторых случаях переходная форма демонстрирует оригинальные аллометрические паттерны (рис. 3, 4).
Темпы морфометрических преобразований (скорости эволюции) оказались различными не только у разных зубов (например, у Р4 значительно выше, чем у р4, а у m2 выше, чем у m1), но и отличающимися у промеров одной зубной коронки. Изменения скорости преобразования могут быть и разнонаправленными. При этом скорости преобразований у медведей среднего плейстоцена в среднем были выше, чем у медведей позднего плейстоцена.
Основные эволюционные тренды размерных параметров верхних щечных зубов в группе денингероидно–спелеоидных медведей (без U. rossicus Borissiak 1930) в плейстоцене (Barysnikov, Puzachenko, 2019) характеризуются: P4 – увеличением общих размеров зуба (индекс IS), увеличением отношения размеров паракона к протокону и уменьшением относительной ширины (индекс IW); M1 – увеличением общих размеров зуба, относительным увеличением длины метакона и в целом задней части зуба, уменьшением индекса IW; M2 – увеличением общих размеров зуба, увеличением отношения паракона к метакону (у U. spelaeus Rosenmüller 1794, U. kanivetz ingressus Rabeder, Hofreiter, Nagel et Withalm 2004), относительным удлинением талона (у U. kanivetz kanivetz Vereshchagin 1973) и небольшим увеличением относительной ширины зуба. В нижнем ряду щечных зубов можно выделить следующие тренды (неопубликованные данные): p4 – увеличение индексов IS и IW, m1 – увеличение индекса IS, увеличение длины и ширины тригонида, увеличение ширины талонида и связанным с этими изменениями увеличением индекса IW, m2 – увеличение индекса IS, пропорциональное увеличение длины и ширины тригонида и талонида, m3 – увеличение индекса IS, снижение индекса IW, увеличение длины и ширины талонида. На фоне этих тенденций относительное положение щечных зубов среднеплейстоценового подвида U. k. praekudarensis выглядит следующим образом: P4 – между U. deningeri von Reichenau 1904 и U. spelaeus, M1, M2, p4, m1 – ближе к U. deningeri, чем к спелеоидным медведям, m2, m3 – ближе к спелеоидным медведям (U. s. eremus Rabeder et al. 2004). Таким образом, разные щечные зубы U. k. praekudarensis находились на разных “стадиях” прогрессивной эволюции, аналогичной таковой у пещерных медведей Европы, которую связывают с общей специализацией к потреблению грубой растительной пищи. В рамках общей классификации медведей U. k. praekudarensis располагается на дендрограммах чаще всего между медведем Денингера и крупными европейскими пещерными медведями позднего плейстоцена.
Однако для большинства щечных зубов кударского медведя направление эволюции между средним и поздним плейстоценом существенно отличалось от такового в линии крупных пещерных медведей. Первый верхний премоляр Р4 уже у переходной формы, несильно изменяясь в размерах, приобрел уникальные пропорции, а именно очень высокий индекс IW, превышающий 75% (табл. 4). Для сравнения приведем средние значения индекса у других таксонов: U. etruscus – 70.8, U. deningeri – 71.2, U. spelaeus s. l. – типичная средняя величина 68.6 – 70.3. По значениям этого индекса Р4 кударского медведя из позднего плейстоцена близок к премоляру бурого медведя U. arctos L. 1758 (75.6). М1 кударского медведя уже со среднего плейстоцена характеризуется “доминирование” паракона над метаконом (индекс ILPA_LME), выраженным заметно сильнее, чем у медведя Денингера и больших пещерных медведей. В конце плейстоцена асимметрия размеров передней и задней частей зуба даже усилилась. По величине ILPA_LME наиболее близким к кударскому медведю был малый пещерный медведь (U. rossicus). M2 у U. k. kudarensis по размерам и относительной ширине коронки вполне соответствует средним значением этих показателей в группе больших пещерных медведей.
Эволюция нижнего премоляра p4 от U. k. praekudarensis к U. k. kudarensis соответствует тренду “медведь Денингера–пещерные медведи”. По величине индекса IW, p4 переходной формы близок к таковому U. k. kudarensis, а по общим размерам несколько меньше. Эти параметры премоляра позднеплейстоценового кударского медведя наиболее блики к параметрам U. s. eremus. Эволюция m1 также лежит в общем тренде эволюции пещерных медведей, однако в этом случае при отсутствии изменений в размерах моляр кударских медведей имеет больший индекс ширины коронки. Отметим, что относительная ширина m1 (индекс IW) у медведя Денингера и U. k. praekudarensis находится в пределах значений, типичных для плиоценового вида U. minimus Devéze de Chabriol et Bouillet 1827. Соответствующий индекс у пещерных медведей выше, но он ниже, чем у европейского этрусского медведя (U. etruscus G. Cuvier 1823) и малого пещерного медведя (U. rossicus). Значения индекса у U. k. kudarensis лежат между значениями индекса у больших пещерных медведей (U. k. kanivetz) и у этрусского и малого пещерного медведей. Параметры m2 у U. k. praekudarensis также, как и у медведя Денингера, близки к значениям, характерным для подвидов U. spelaeus eremus, U. s. ladinicus. Изменения этого зуба у переходной формы и U. k. kudarensis выразились в существенном увеличении ширины коронки при некотором сокращении ее длины (табл. 4: индекс IW>62%). По величине IW второго нижнего моляра позднеплейстоценовый кударский медведь находится между этрусским и бурым медведями, но в целом параметры зуба близки к таковым m2 пещерных медведей. Как было показано выше, параметры третьего нижнего моляра m3 кударского медведя на протяжении среднего и позднего плейстоцена изменялись незначительно и не совпадали с трендом на увеличение длины талонида в ряду медведь Денингера – пещерные медведи.
Приведенные сравнения убедительно демонстрируют уникальный, независимый характер эволюции щечных зубов кударского медведя на временном промежутке середина среднего– поздний плейстоцен относительно линии европейских спелеоидных медведей. Во многих случаях позднеплейстоценовый подвид U. k. kudarensis характеризуется относительно более широкими зубными коронками моляров по сравнению с типичными пещерными медведями (U. spelaeus, U. kanivetz). Трудно сказать, в какой мере это может отражать степень адаптации к питанию растительными кормами. Для прояснения этого вопроса потребуются сравнительные исследования изотопного состава костей кударских и пещерных медведей.
Своеобразие зубов поздних кударских медведей выражено в разной степени в верхнем и нижнем рядах. В рамках общей классификации по признакам верхних щечных зубов (Baryshnikov, Puzachenko, 2019), включающей расширенный перечень местонахождений кударского медведя, а также бурого медведя, медведя Денингера, малого и больших пещерных медведей, U. k. kudarensis образует общий кластер с U. k. praekudarensis (P4) или располагается на классификационном дереве в непосредственной близости от него (M1, M2). Аналогичная классификация для нижнего ряда зубов (неопубликованные данные) демонстрирует более сложную картину: p4 – U. k. kudarensis расположен на классификационном дереве внутри группы больших пещерных медведей, между таксонами U. s. eremus и U. s. spelaeus, m1 – U. k. kudarensis расположен между кластером, включающим бурого медведя, и мелким подвидом пещерного медведя (U. s. ladinicus), m2 – U. k. kudarensis расположен внутри группы больших пещерных медведей, между U. s. eremus и U. s. ladinicus, m3 – U. k. kudarensis входит в один кластер с медведем Денингера и U. k. praekudarensis. Неустойчивость классификаций отражает специфику эволюции отдельных зубов, которая “маскирует” филогенетический сигнал.
Количественные оценки скорости изменений морфологических признаков на палеонтологическом материале обычно проблематичны, поскольку в редких случаях можно быть уверенными в прямой генетической преемственности между разновозрастными выборками. В нашем случае гипотеза прямой генетической связи между средне- и позднеплейстоценовыми кударскими медведями не может считаться доказанной, но до получения результатов анализа ископаемой ДНК ее нельзя исключать. В рамках этой гипотезы скорости изменений разных групп параметров одного зуба и разных зубов существенно различаются в среднем и позднем плейстоцене (табл. 9). Если рассматривать переходную форму в качестве реального стратиграфического промежуточного таксона между U. k. praekudarensis и U. k. kudarensis, то следует считать изменения общих размеров зубов обратимыми во времени, поскольку по этим признакам зубы у обоих подвидов меньше, чем зубы переходной формы. При этом некоторые пропорции коронки, по которым различаются подвиды, отмечаются уже у переходной формы.
В литературе имеются сведения о быстрых (в масштабах экологического времени, < 100 лет или < 30 поколений) морфологических изменениях видов грызунов в условиях островной изоляции (Pergams, Ashley, 2001) со скоростями порядка 10–3h на генерацию. Основываясь на данных Б. Куртена по эволюции размеров M1 в линии пещерных медведей, Линч (Lynch, 1990: 730) определил среднюю скорость эволюции в 4.8 × 10–5 (7.5 × 10–6–8.6 × 10–5) на поколение (принято равным 10 лет) или в интервале приблизительно 0.0075 – 0.086 на 10 тыс. лет. Несмотря на некоторые существенные отличия в схеме расчетов, мы полагаем, что эти результаты можно сравнить с нашими оценками скорости эволюции размеров зубов кударского медведя по порядку величин. При таких условиях скорость изменений размеров зубов в среднем – позднем плейстоцене у кударского вида была относительно низкой. Увеличение размеров зубов у переходной формы может быть следствием увеличения общих размеров животных как реакция на суровые условия гляциальной стадии МИС 8 (днепровское оледенение). Последующее уменьшение размеров зубов в позднем плейстоцене, возможно, было обусловлено сочетанием ряда палеогеографических факторов: эмское (микулинское) потепление в начале позднего плейстоцена, сменившееся длительным похолоданием вислинского (валдайского) оледенения в сочетании с более высокогорными условиями в районе кударских пещер по сравнению со средним плейстоценом (рис. 5). Популяция медведей в районе пещер Кударо 1 и Кударо 3, скорее всего, вымирает перед максимумом валдайского оледенения, вероятно, вследствие развития горного оледенения и увеличения стресса при конкуренции за скальные убежища из-за возрастания численности и расселения человека современного анатомического облика.
Аллометрическая изменчивость отражает вариации одних и тех же механизмов генетической и эпигенетической регуляции закладки и роста разных зубов, общих для всех млекопитающих (Jernvall, Thesleff, 2012). Небольшие вариации в экспрессии регуляторных генов могут приводить к заметным изменениям морфологии зуба. При этом не исключены параллелизмы и формирование сходных пропорций зубных коронок на основе разных модификаций онтогенеза. Аллометрическая изменчивость верхнего ряда щечных зубов медведей несет филогенетический сигнал на уровне таксонов видового и даже подвидового уровня (Baryshnikov, Puzachenko, 2019). В нашем случае аллометрический паттерн верхнего и нижнего рядов щечных зубов переходной формы ближе к паттерну U. k. praekudarensis, что можно интерпретировать в качестве косвенного свидетельства об их филогенетической связи. В этом случае формирование на стадии переходной формы некоторых отношений между разными частями зубной коронки, интегрально выраженных, например, в значениях индекса ширины коронки (IW), и типичных для позднеплейстоценового подвида, могло быть реализовано при сохранении исходного аллометрического паттерна U. k. praekudarensis. Элементы этого “базового” паттерна сохраняются у U. k. kudarensis на фоне инноваций, связанных в том числе с уменьшением размеров зубов. Альтернативная гипотеза предполагает парафилию, т. е. независимое происхождение позднеплейстоценовых кударских медведей, обитавших в районе кударских пещер в позднем плейстоцене, от другой среднеплейстоценовой популяции U. kudarensis, в той или иной мере родственной переходной форме, но с несколько иным локальным вариантом аллометрического паттерна.
Механизмы регуляции онтогенеза отдельных зубов определяют и отношения между соседними зубами в пределах зубного ряда (Salazar-Ciudad, Jernvall, 2002; Kavanagh et al., 2007; Renvoisé et al., 2009; Asahara et al., 2016 и др.). Принципы этих отношений, видимо, довольно консервативны, а их варианты у млекопитающих характеризуют таксоны выше родового ранга (семейства и отряды). Семейство медвежьих (Ursidae) в этом отношении характеризуется рядом особенностей (Asahara et al., 2016), которые, вероятно, обусловлены относительно низким взаимодействием между соседними молярами в онтогенезе, что приводит к меньшим различиям в их размерах. В нашей работе мы продемонстрировали вероятность влияния четвертых премоляров на размеры моляров. В онтогенезе премоляры закладываются на месте молочных зубов D4 и d4 и могут непосредственно взаимодействовать (“конкурировать”) с М1 и m1 (Debeljak, 1996). Так или иначе, именно учет варьирования размеров премоляров позволяет описывать существенную часть размерной изменчивости, существующей между молярами. Отметим, что конкретные параметры этих отношений могут содержать филогенетический сигнал, что требует дальнейшего исследования.
Аллометрическая изменчивость и взаимодействия зубов внутри зубных рядов обусловлены особенностями регуляции онтогенеза до завершения роста зубов. У взрослых животных на их изменчивость влияют отношения с соответствующими им зубами противолежащего ряда, возникающие в процессе окклюзии. Мы показали, что этот эпигенетический фактор у кударского медведя играет основную роль в ковариации размеров коронок взрослых животных, на фоне действия которого ковариация внутри каждого ряда практически не различима. При этом относительно независимо варьируют размеры премоляров и третьего нижнего моляра, что соответствует схеме окклюзии верхних и нижних рядов пещерных медведей. У всех рассмотренных выборок кударского медведя обнаружена принципиально одна и та же схема организации функциональных связей верхних и нижних щечных зубов, что может объясняться сохранением пищевой специализации и генетической связи у медведей в среднем–позднем плейстоцене.
Список литературы
Барышников Г.Ф., 2007. Семейство медвежьих (Carnivora, Ursidae) // Фауна России и сопредельных стран. Млекопитающие. Т. 1. Вып. 5. СПб.: Наука. 542 с.
Куприянова И.Ф., Пузаченко А.Ю., Агаджанян А.К., 2003. Временные и пространственные компоненты изменчивости черепа обыкновенной бурозубки, Sorex araneus (Insectivora) // Зоологический журнал. Т. 82. № 7. С. 839–851.
Любин В.П., 1998. Ашельская эпоха на Кавказе // Археологические изыскания. Т. 47. С. 1–192 с.
Несмеянов С.А., 1999. Геоморфологические аспекты палеоэкологии горного палеолита. М.: Научный мир. 392 с.
Пузаченко А.Ю., 2001. Внутрипопуляционная изменчивость черепа обыкновенного слепыша Spalax microphthalmus (Spalacidae, Кodentia). 1. Методика анализа данных, невозрастная изменчивость самцов // Зоологический журнал. Т. 80. № 3. С. 1–15.
Пузаченко А.Ю., 2016. Количественные закономерности морфологического разнообразия черепа млекопитающих // Сборник трудов Зоологического музея МГУ. Т. 54. С. 229–268.
Abramov A.V., Puzachenko A.Y., 2012. Species Co-existence and morphological divergence in West Siberian Mustelids (Carnivora, Mustelidae) // Mammal Study. V. 37. № 3. P. 255–259.
Asahara M., Saito K., Kishida T., Takahashi K., Bessho K., 2016. Unique pattern of dietary adaptation in the dentition of Carnivora: its advantage and developmental origin // Proceedings of the Royal Society B. Biological Sciences. V. 283. № 1832. P. 1–10.
Baryshnikov G., 1998. Cave bears from the Paleolithic of the Greater Caucasus // Quaternary Paleozoology in the Northern Hemisphere. Illinois State Museum Scientific Papers. V. XXVII. Eds. Saunders J.J., Styles B.W., Baryshnikov G.F. Springfield: Illinois State Museum. P. 69–118.
Baryshnikov G., 2006. Morphometrical variability of cheek teeth in cave bears // Scientific Annals, School of Geology, Aristotle University of Thessaloniki. V. 98. P. 81–102.
Baryshnikov G.F., 2011. Pleistocene Felidae (Mammalia, Carnivora) from Paleolithic site in Kudaro caves in the Caucasus // Proceedings of the Zoological Institute RAS. V. 315. № 3. P. 197–226.
Baryshnikov G.F., Gimranov, D.O., Kosintsev P.A., 2019. Variability of the upper incisors in the cave bears (Carnivora, Ursidae) from the Caucasus and Urals // Comptes Rendus Palevol. V. 18. № 2. P. 209–222.
Baryshnikov G., Puzachenko A., 2011. Craniometrical variability in the cave bears (Carnivora, Ursidae): multivariate comparative analysis // Quaternary International. V. 245. P. 350–368.
Baryshnikov G.F., Puzachenko A.Y., 2017. Morphometrical analysis of metacarpal and metatarsal bones of cave bears (Carnivora, Ursidae) // Fossil Imprint. V. 73. № 1–2. P. 7–47.
Baryshnikov G.F., Puzachenko A.Y. 2019. Morphometry of upper cheek teeth of cave bears (Carnivora, Ursidae) // Boreas. https://doi.org/10.1111/bor.12360
Baryshnikov G.F., Puzachenko A.Y., Baryshnikova S.V., 2018. Morphometric analysis of cave bear mandibles (Carnivora, Ursidae) // Revue de Palèobiologie, Genève. V. 37. № 2. P. 379-393.
Cock A.G., 1966. Genetical aspects of metrical growth and form in animals // Quarterly Review of Biology. V. 41. P. 131–190.
Colwell R.K., Futuyma D.J., 1971. On the measurement of niche breadth and Overlap // Ecology. V. 52. № 4. P. 567–576.
Dabney J., Knapp M., Glocke I., Gansauge M.-T., Weihmann A., Nickel B., Valdiosera C., Garcia, N., Pääbo S., Arsuaga J.-L. Meyer M., 2013. Complete mitochondrial genome sequence of a Middle Pleistocene cave bear reconstructed from ultrashort DNA fragments // Proceedings of the National Academy of Sciences of the United States of America. V. 110. P. 15758–15763.
Davison M. L., Jones L.E., 1983. Special issue: multidimensional scaling and its applications // Applied Psychological Measurement. V. 7. P. 373–514.
Debeljak I., 1996. Ontogenetic development of dentition in the cave bear // Geologija. V. 39. P. 13–77.
Dempster A.P., Laird N.M., Rubin D.B., 1977. Maximum Likelihood from Incomplete Data via the EM Algorithm // Journal of the Royal Statistical Society. Ser. B. V. 39. № 1. P. 1–38.
Haldane J.B.S., 1949. Suggestions as to quantitative measurement of rates of evolution // Evolution. V. 3. P. 51–56.
Jernvall J., Thesleff I., 2012. Tooth shape formation and tooth renewal: evolving with the same signals // Development. V. 139. P. 3487–3497.
Jolicoeur P., 1963. Note: the multivariate generalization of the allometry equation // Biometrics. V. 9. P. 497–499.
Kavanagh K.D., Evans A.R., Jernvall J., 2007. Predicting evolutionary patterns of mammalian teeth from development // Nature. V. 449. P. 427–432.
Kendall M. G., 1975. Rank Correlation Methods. London: Charles Griffin and Co. 202 p.
Klingenberg C.P., 1996. Multivariate allometry // Advances in Morphometrics. Eds Marcus L.F., Corti, M., Loy A., Naylor G.J.P., Slice D.E. New York: Plenum Press, P. 23–49.
Knapp M., Rohland N., Weinstock J., Baryshnikov G., Sher A., Doris N., Rabeder G., Pinhasi R., Schmitt H., Hoffreiter M., 2009. Fist DNA sequences of Asian cave bear fossils reveal deep divergences and complex phylogeographic patterns // Molecular Ecology V. 18. № 6. P. 1225–1238.
Kruskal B., 1964. Multidimensional scaling by optimizing goodness of fit to nonmetric hypothesis // Psychometrika. V. 29. P. 1–27.
Lisiecki L.E., Raymo M.E., 2005. A Pliocene-Pleistocene stack of 57 globally distributed benthic δ18O records // Paleoceanography. V. 20. № 1. PA1003. P. 1–17.
Lynch M., 1990. The Rate of Morphological Evolution in Mammals from the Standpoint of the Neutral Expectation // The American Naturalist. V. 136. № 6. P. 727–741.
Pergams O.R.W., Ashley M. V., 2001. Microevolution in island rodents // Genetica. V. 112–113. P. 245–256.
Pianka E.R., 1974. Niche overlap and diffuse competition // Proceedings of the National Academy of Sciences. V. 71. P. 2141–2145.
Renvoisé E., Evans A.R., Jebrane A., Labruère C., Laffont R., Montuire S., 2009. Evolution of mammal tooth patterns: new insights from a developmental prediction model // Evolution. V. 63. № 5. P. 1327–1340.
Salazar-Ciudad I., Jernvall J., 2002. A gene network model accounting for development and evolution of mammalian teeth // Proceedings of the National Academy of Sciences. V. 99. № 12. P. 8116–8120.
Sher A.V., Weinstock J., Baryshnikov G.F., Davydov S.P., Boeskorov G.G., Zazhigin V.S., Nikolsky P.A., 2011. The first records of “spelaeoid” bears in Arctic Siberia // Quaternary Science Reviews. V. 30. № 17–18. P. 2238–2249.
Stefansky W., 1972. Rejecting Outliers in Factorial Designs // Technometrics. V. 14. № 2. P. 469–479.
Stiller M., Baryshnikov G., Bocherens H., Grandal d’Anglade A., Hilpert B., Munzel S.C., Pinhasi, R., Rabeder G., Rosendahl W., Trinkaus E., Hofreiter M., Knapp M., 2010. Withering away – 25 000 years of genetic decline preceded cave bear extinction // Molecular Biology and Evolution. V. 27. № 5. P. 975–978.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал