

Зоологический журнал, 2019, T. 98, № 11, стр. 1205-1212
Три новых вида из группы видов Thinodromus arcuatus, род Thinodromus kraatz 1857 (Coleoptera, Staphylinidae, Oxytelinae), из Восточной области
М. Ю. Гильденков *
Смоленский государственный университет
214000 Смоленск, Россия
* E-mail: mgildenkov@mail.ru
Поступила в редакцию 03.12.2018
После доработки 12.02.2019
Принята к публикации 24.03.2019
Аннотация
Описаны три новых вида рода Thinodromus Kraatz 1857 из группы видов Thinodromus (Amisammus) arcuatus: Thinodromus (Amisammus) diancangensis sp. n. из Юннаня, Thinodromus (Amisammus) gentlus sp. n. из Северной Индии и Непала и Thinodromus (Amisammus) smetanai sp. n. из Тайваня.
К группе видов Thinodromus arcuatus, далее – группе “arcuatus” принадлежат (Гильденков, 2000, 2001; Gildenkov, 2000) следующие виды: Thinodromus (Amisammus) arcuatus (Stephens 1834), Thinodromus (Amisammus) motschulskyi (Hochhuth 1860), Thinodromus (Amisammus) dilaticollis (Eppelsheim 1884), Thinodromus (Amisammus) pilosellus (Eppelsheim 1890) и Thinodromus (Amisammus) japonicus (Cameron 1930). Возможно к ней же относится и Thinodromus (Amisammus) renominatus Herman 1970 из Турции, но самец этого вида до сих пор остается неизвестным и принадлежность его к группе “arcuatus” нельзя считать обоснованной. Все представители группы “arcuatus” отличаются от других видов подрода Amisammus Gozis 1886 наличием во внутреннем мешке эдеагуса двух характерных зубовидных структур, направленных вершинами к дорсальной стороне эдеагуса. Как правило, основания этих структур довольно широко расставлены, а вершины наклоняются к основанию эдеагуса и сходятся, образуя таким образом вершину угла (Гильденков, 2000: С. 919: Рис. 1: 3; Гильденков, 2001: С. 154: Рис. 21 : 1; Gildenkov, 2000: P. 832: Fig. 1: 3). Только у T. japonicus эти склериты располагаются почти параллельно и угла не образуют. Строение и расположение зубовидных структур в мешке эдеагуса служат важными диагностическими признаками. Самые длинные и относительно узкие структуры имеются у T. arcuatus (Гильденков, 2000: С. 919: Рис. 1: 4; Гильденков, 2001: С. 154: Рис. 21 : 2; Gildenkov, 2000: P. 832: Fig. 1: 4); немного более широкие почти такие же длинные структуры у T. motschulskyi (Гильденков, 2000: С. 919: Рис. 1: 5; Гильденков, 2001: С. 154: Рис. 21 : 3; Gildenkov, 2000: P. 832: Fig. 1: 5); у T. pilosellus (Гильденков, 2000: С. 919: Рис. 1: 6; Гильденков, 2001: С. 154: Рис. 21 : 4; Gildenkov, 2000: P. 832: Fig. 1: 6) зубовидные структуры относительно короткие и широкие; у T. dilaticollis эти структуры самые узкие и относительно короткие (Гильденков, 2000: С. 919: Рис. 1: 8; Гильденков, 2001: С. 154: Рис. 21 : 5; Gildenkov, 2000: P. 832: Fig. 1: 8). Сведения о распространении видов из группы “arcuatus” хорошо отражены в последнем каталоге Палеарктики (Schülke, Smetana, 2015). Все указанные выше виды известны только из Палеарктики, причем практически все они распространены на Кавказе и в Турции. Только T. japonicus значительно отдален от других видов группы “arcuatus” на восток – Япония и Южная Корея. Наиболее известным видом в группе является широко распространенный в Европе (Schülke, Smetana, 2015) T. arcuatus, кроме указанных в каталоге регионов для Европы, нам он также известен (Гильденков, 2001; Гильденков и др., 2011) из центральных территорий европейской части России (Московская и Смоленская области) и европейской части Турции (Гильденков, 2009). Распространение этого вида в Северной Африке, Средней Азии и Сибири (Schülke, Smetana, 2015) кажется нам весьма сомнительным и требует подтверждения, так как ранее вид зачастую не отличали от других представителей подрода Amisammus, таких как T. behnei Gildenkov 2000 и T. kurbatovi Gildenkov 2000, описанных сравнительно недавно. В Северной Африке вид могли не отличать от T. mannerheimii Kolenati 1846, достоверные данные о распространении которого в Марокко стали известны также недавно (Гильденков, 2001). T. motschulskyi, кроме указанных в каталоге регионов (Schülke, Smetana, 2015), известен нам (Гильденков, 2009) из азиатской части Турции.
Рис. 1.
Внешний вид дорсально: a – Thinodromus diancangensis sp. n., голотип; b – T. japonicus (Cameron 1930), Япония.

Описание трех новых видов из Восточной биогеографической области представляет интерес для понимания распространения видов группы “arcuatus”. В частности, распространение этих видов заполняет пробел между ареалом T. japonicus и остальных видов группы.
Места хранения материала обозначены следующим образом: cAS – личная коллекция Алеша Сметаны, Оттава, Канада (private collection of Aleš Smetana, Ottava); cGR – личная коллекция Гийома де Ружмона, Оксфорд, Великобритания (private collection of Guillaume de Rougemont, Oxford); cMG – личная коллекция автора, Смоленск, Россия (private collection of Mikhail Gildenkov, Smolensk); cMSch – личная коллекция Михаэля Шульке, Берлин, Германия (private collection of Michael Schülke, Berlin); FMNH – Музей Естествознания Филда, Чикаго, США (Field Museum of Natural History); NHMW – Музей естествознания Вены, Австрия (Naturhistorisches Museum Wien).
Этикетки приведены в оригинальной транскрипции, знак “|” обозначает конец строки. В исследованиях использованы стандартные методы по таксономии насекомых, препарирование, измерения и рисунки выполнены с использованием микроскопа МБС-10, снабженного окуляр-микрометром и мерной сеткой. Препараты гениталий обработаны 10% KOH и зафиксированы в канадском бальзаме или эупарале. В представленных описаниях, где дается отношение длины к ширине для головы, переднеспинки и надкрылий, использованы условные единицы: 7 условных единиц = 0.1 мм, соответственно 1 условная единица составляет около 0.0143 мм. Фотографии получены на Canon EOS 5D Mark III с объективом Canon MP-E 65 mm, использована технология extended focus.
Thinodromus (Amisammus) diancangensis Gildenkov sp. n. (рис. 1a; 3a)
Материал. Голотип ♂, Китай, провинция Юннань “CHINA: Yunnan [CH07-03], Dali Bai Auton Pref., Diancang Shan W Dali, 25°41′49″ N, 100°06′24″ E, 2970 m, sifted at rock edges and under small shrubs, 28.V.2007, M. Schülke” (cMSch).
Паратип: 1♀, Китай, провинция Юннань “CHINA: Yunnan [CH07-03], Dali Bai Auton Pref., Diancang Shan W Dali, 25°41′49″ N, 100°06′24″ E, 2970 m, sifted at rock edges and under small shrubs, 28.V.2007, M. Schülke” (cMG).
Описание. Самец (голотип). Длина тела около 3.1 мм. Голова, переднеспинка и брюшко темно-бурые, надкрылья, ноги и антенны бурые; покровы умеренно блестящие. Тело покрыто довольно короткими светлыми волосками.
Голова поперечная, ее длина от заднего края до переднего края наличника относится к наибольшей ширине примерно как 25 : 37. Виски развиты слабо, глаза большие, выпуклые; видимый сверху диаметр глаза более чем в 3 раза превосходит длину виска (рис. 1a). Пунктировка четкая, крупная и густая, диаметр точек примерно равен диаметру фасетки глаза, расстояние между точками значительно меньше их диаметра. Антенны довольно короткие, 1–7-й членики удлиненные; 8 и 9-й примерно равной длины и ширины; 10-й – слабо поперечный; 11-й членик удлиненный, конусовидный (рис. 1a).
Переднеспинка широкая, ее длина относится к наибольшей ширине примерно как 35 : 42. Пунктировка четкая, крупная и густая, диаметр точек примерно равен диаметру фасетки глаза, расстояние между точками значительно меньше их диаметра (рис. 1a). В основании диска переднеспинки хорошо развито подковообразное вдавление (рис. 1a); в центральной части диска имеются два симметричных хорошо развитых продольных, овальных вдавления; ближе к переднему краю имеется непарное, также хорошо развитое овальное вдавление по медиальной линии (рис. 1a).
Надкрылья довольно широкие, их длина относится к общей ширине примерно как 51 : 59. Поверхность надкрылий четко, крупно и густо пунктирована. Диаметр точек примерно равен диаметру фасетки глаза. Расстояние между точками ззначительно меньше их диаметра, промежутки умеренно блестящие.
Брюшко нежно, мелко и довольно густо пунктировано, диаметр точек во много раз меньше, чем на переднеспинке и надкрыльях, расстояние между точками немного больше их диаметра.
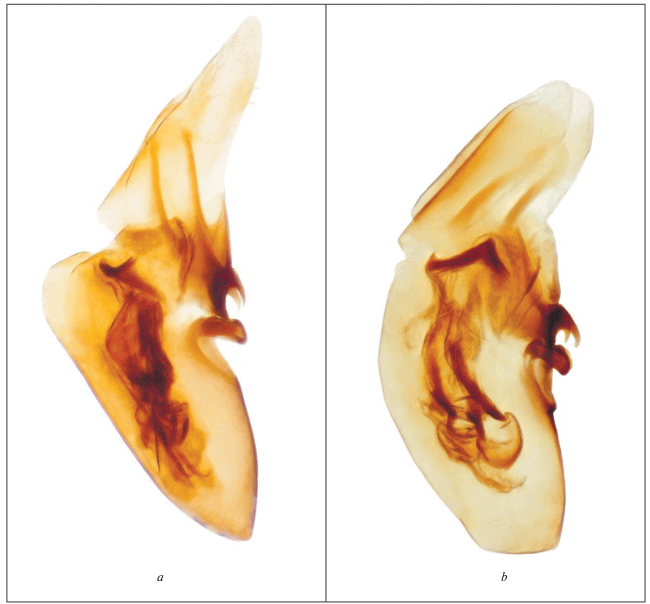
Эдеагус имеет характерное строение (рис. 3a). Парные зубовидные склериты расположены параллельно основанию эдеагуса, очень короткие и довольно широкие, их основания широко расставлены, вершины сильно заостренные, сходятся образуя угол.
Рис. 2.
Внешний вид дорсально: a – Thinodromus gentlus sp. n., голотип; b – T. smetanai sp. n., голотип.

Рис. 3.
Эдеагус латерально: a – Thinodromus diancangensis sp. n., голотип; b – T. japonicus (Cameron 1930), Япония.
Самка (паратип). Имеет сходную с самцом (голотипом) окраску и пропорции тела, половой диморфизм не выражен. Сперматека равно-раздельная.
Дифференциальный диагноз. Размерами, окраской, характером пунктировки на голове и переднеспинке сходен с описанным из Северной Индии T. (Amisammus) vulneratus (Bernhauer 1926), но отличается от него немного более крупной пунктировкой головы и переднеспинки, более короткими и толстыми антеннами, строением эдеагуса. T. vulneratus имеет совершенно иной план строения эдеагуса и относится к группе видов “kiesenwetterii” (Гильденков, 2017, 2018; Gildenkov, 2017). Характером пунктировки сходен с близким T. japonicus (рис. 1b), от которого отличается заметно более крупной пунктировкой головы и переднеспинки; строением эдеагуса, в котором значительно более короткие зубовидные структуры (рис. 3a, 3b), широко расставленные в основании.
Распространение – юго-запад Китая: провинция Юннань.
Этимология. Назван в соответствии с типовым местом.
Thinodromus (Amisammus) gentlus Gildenkov sp. n. (рис. 2a; 4a)
Материал. Голотип ♂, Северная Индия, штат Уттаракханд “N-INDIA: Uttaranchal state ca. 13 km NW Nainital Khairna Bridhe env. 900–1000 m. 13.–17.7.2003 leg. Z. Kejval & M. Trýzna” (NHMW).
Паратипы: 1♂, 4♀♀ Северная Индия, штат Уттаракханд “N-INDIA: Uttaranchal state ca. 13 km NW Nainital Khairna Bridhe env. 900–1000 m. 13.–17.7.2003 leg. Z. Kejval & M. Trýzna” (NHMW; 1♂, 1♀ – cMG); 1♂ Северная Индия, штат Уттаракханд “IND: Uttaranchal Riv. Dukura, Bageshwar Dist. 29°53′38″ N/79°39′45″ E 13.XI.2006 leg. M.A. Jäch (18)” “left tributary of Riv. Gomti at Borgad ca 1 km E Jaisar 1060 m a.s.l.” (NHMW); 1♂ Северная Индия, штат Уттаракханд “Binal Gad 3500 Chakrata Dist. India” “Chakrata Dist. Binal Gad 3500” “Dr. Cameron 15.V.22.” “Trog. lunatus Motsch” “T. (Thinodr.) lunatus Motsch.” “Field Mus. Nat. Hist. 1966 a. Bierig Colln. Acc. Z-13812” (FMNH); 1♀ Непал “NEPAL: Panauti19.III.1981 de Rougemont” (cGR); 1♀ Северная Индия, штат Химачал-Прадеш “INDIA: H.P. Mondi VI.1981” (cGR).
Описание. Самец (голотип). Длина тела около 3.0 мм. Темно-бурый, ноги и основание антенн светло-бурые; покровы довольно блестящие. Тело покрыто длинными светлыми волосками.
Голова поперечная, ее длина от заднего края до переднего края наличника относится к наибольшей ширине примерно как 23 : 37. Виски едва развиты, глаза большие, выпуклые, занимают почти всю боковую сторону головы (рис. 2a). Пунктировка очень нежная, мелкая и довольно редкая, точки не глубокие, их диаметр примерно в 4.5 раза меньше диаметра фасетки глаза, расстояние между точками на темени значительно больше их диаметра, на лбу немного больше их диаметра. Антенны довольно длинные, 1–7-й членики удлиненные; 8-й – примерно равной длины и ширины; 9-й и 10-й – слабо поперечный; 11-й членик удлиненный, конусовидный (рис. 2a).
Переднеспинка довольно широкая, ее длина относится к наибольшей ширине примерно как 31 : 43. Пунктировка очень нежная, мелкая и довольно редкая, точки не глубокие, их диаметр примерно в 4.5 раза меньше диаметра фасетки глаза, расстояние между точками немного больше их диаметра (рис. 2a). В основании диска переднеспинки хорошо развито подковообразное вдавление (рис. 2a); в центральной части диска имеются два симметричных едва развитых овальных вдавления; ближе к переднему краю имеется непарное, также едва развитое овальное вдавление по медиальной линии (рис. 2a).
Надкрылья довольно широкие, их длина относится к общей ширине примерно как 56 : 64. Поверхность надкрылий четко, довольно крупно и густо пунктирована. Диаметр точек примерно равен диаметру фасетки глаза. Расстояние между точками заметно меньше их диаметра, промежутки умеренно блестящие.
Брюшко нежно, очень мелко и редко пунктировано, диаметр точек немного меньше, чем на переднеспинке и голове, расстояние между точками значительно больше их диаметра.
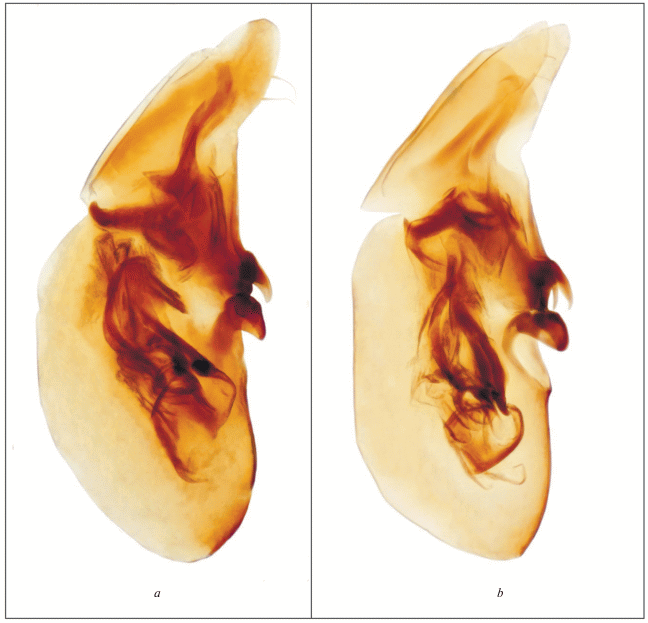
Эдеагус имеет характерное строение (рис. 4a). Парные зубовидные склериты расположены параллельно основанию эдеагуса, короткие и широкие, их основания широко расставлены, вершины слабо заостренные, сходятся образуя угол.
Рис. 4.
Эдеагус латерально: a – Thinodromus gentlus sp. n., голотип; b – T. smetanai sp. n., голотип.
Самка (паратипы). Имеет сходную с самцом окраску и пропорции тела, половой диморфизм не выражен. Сперматека равно-раздельная.
Изменчивость. Паратипы (самцы и самки) имеют идентичные с голотипом окраску, характер опушения и пунктировки поверхности тела. Наблюдается лишь небольшая изменчивость в размерах.
Дифференциальный диагноз. Размерами, окраской, длинными волосками на поверхности тела, характером пунктировки на голове и переднеспинке очень сходен с близким видом T. pilosellus, от которого отличается более четкой пунктировкой на переднеспинке – точки более нежные, но вполне различимые. Также новый вид надежно отличается от T. pilosellus строением эдеагуса. Зубовидные структуры в эдеагусе T. pilosellus довольно короткие, заостренные (Гильденков, 2000: С. 919: Рис. 1: 6; Гильденков, 2001: С. 154: Рис. 21 : 4; Gildenkov, 2000: P. 832: Fig. 1: 6), наклонены вершинами к основанию эдеагуса. Зубовидные структуры у T. pilosellus очень сходны с таковыми у T. japonicus (рис. 3b; 4a), но их основания широко расставлены, а у T. japonicus сближены.
Распространение – Северная Индия, Непал.
Этимология. От латинского “gentle” – “нежный”, название связано с нежной пунктировкой головы и переднеспинки.
Thinodromus (Amisammus) smetanai Gildenkov sp. n. (рис. 2b; 4b)
Материал. Голотип ♂, Тайвань “TAIWAN Kaohsiung Hsien, rd. from Tengchin to Chuyunshan 1400m 25.IV.90 A. Smetana [T23]” (cAS).
Паратипы: 1♂, 1♀, Тайвань “TAIWAN Nantou Hsien, Shuantung 150 m 21.IV.1990 A. Smetana [T14]” (cAS); 1♂, 1♀, Тайвань “TAIWAN Taitung Hsien Foothills of Hsinkangshan nr. Chengkung, 350–400 m 19.VII.93 A. Smetana [T155]” (cAS; 1♂ – cMG).
Описание. Самец (голотип). Длина тела около 3.5 мм. Темно-бурый, ноги и антенны бурые; покровы слабо блестящие. Тело покрыто довольно короткими светлыми волосками.
Голова поперечная, ее длина от заднего края до переднего края наличника относится к наибольшей ширине примерно как 27 : 40. Виски развиты слабо, глаза большие, выпуклые; видимый сверху диаметр глаза более чем в 4 раза превосходит длину виска (рис. 2b). Пунктировка четкая, мелкая и густая, диаметр точек примерно в 3 раза меньше диаметра фасетки глаза, расстояние между точками на темени немного больше их диаметра, вблизи края глаз – заметно меньше их диаметра. Антенны довольно длинные, 1–8-й членики удлиненные; 9-й и 10-й примерно равной длины и ширины; 11-й членик удлиненный, конусовидный (рис. 2b).
Переднеспинка широкая, ее длина относится к наибольшей ширине примерно как 38 : 48. Пунктировка четкая, мелкая и густая, диаметр точек примерно в 3 раза меньше диаметра фасетки глаза, расстояние между точками в центре диска немного больше их диаметра. У внешних углов, при основании, пунктировка более густая, расстояние между точками значительно меньше их диаметра (рис. 2b). В основании диска переднеспинки хорошо развито подковообразное вдавление (рис. 2b); в центральной части диска имеются два симметричных хорошо развитых продольных, овальных вдавления; ближе к переднему краю имеется непарное, также хорошо развитое овальное вдавление по медиальной линии (рис. 2b).
Надкрылья довольно широкие, их длина относится к общей ширине примерно как 66 : 71. Поверхность надкрылий четко, довольно крупно и густо пунктирована. Диаметр точек примерно в 1.5 раза меньше диаметра фасетки глаза. Расстояние между точками едва меньше их диаметра, промежутки умеренно блестящие.
Брюшко нежно, мелко и довольно густо пунктировано, диаметр точек значительно меньше, чем на переднеспинке, расстояние между точками немного больше их диаметра.
Эдеагус имеет характерное строение (рис. 4b). Парные зубовидные склериты наклонены вершинами к основанию эдеагуса, короткие и широкие, их основания широко расставлены, вершины сильно заостренные, сходятся образуя угол.
Самка (паратипы). Имеет сходную с самцом окраску и пропорции тела, половой диморфизм не выражен. Сперматека равно-раздельная.
Изменчивость. Паратипы (самцы и самки) имеют идентичные с голотипом окраску, характер опушения и пунктировки поверхности тела. Наблюдается лишь небольшая изменчивость в размерах.
Дифференциальный диагноз. От сходного окраской и размерами T. japonicus (рис. 1b), хорошо отличается значительно более мелкой пунктировкой головы, переднеспинки и надкрылий (рис. 2b); надёжно отличается более короткими и широкими зубовидными структурами (рис. 3b; 4b) в эдеагусе, основания которых широко расставлены.
Распространение – Тайвань.
Этимология. Назван именем Алеша Сметаны [Aleš Smetana], известного специалиста в области систематики Staphylinidae, собравшего типовую серию данного вида.
БЛАГОДАРНОСТИ
Автор выражает сердечную благодарность своим коллегам и кураторам коллекций, предоставившим материалы для изучения: Aleš Smetana (Оттава); Guillaume de Rougemont (Оксфорд); Michael Schülke (Берлин); Harald Schillhammer (NHMW, Вена); Alfred F. Newton, Margaret K. Thayer and James H. Boone (FMNH, Чикаго). Особую благодарность автор выражает Макарову Кириллу Владимировичу за получение фотографий (Московский педагогический государственный университет, Москва).
Список литературы
Гильденков М.Ю., 2000. Обзор палеарктических видов рода Thinodromus Kraatz (Coleoptera, Staphylinidae). Сообщение 2 // Зоологический журнал. Т. 79. № 8. С. 918–927.
Гильденков М.Ю., 2001. Филогенетические отношения в подсемействе Oxytelinae. Фауна Thinodromus Палеарктики (Coleoptera: Staphylinidae: Oxytelinae). Смоленск: СГПУ. 204 с.
Гильденков М.Ю., 2009. Новые данные о распространении в Палеарктике видов рода Thinodromus и видов рода Carpelimus из подродов Carpelimus s. str., Paratrogophloeus, Bucephalinus (Coleoptera, Staphylinidae) // Известия Смоленского государственного университета. № 2 (6). С. 25–42.
Гильденков М.Ю., 2017. Обзор группы видов Thinodromus lunatus (Coleoptera, Staphylinidae) // Зоологический журнал. Т. 96. № 10. С. 1165–1180.
Гильденков М.Ю., 2018. Шесть новых видов рода Thinodromus Kraatz 1857, близких к Thinodromus (Amisammus) ripicola (Cameron 1941) (Coleoptera, Staphylinidae, Oxytelinae) // Зоологический журнал. Т. 97. № 11. С. 1363–1373.
Гильденков М.Ю., Семионенков О.И., Стародубцева О.А., 2011. Стафилиниды подсемейства Oxytelinae (Coleoptera, Staphylinidae) Смоленской области // Известия Смоленского государственного университета. № 3 (15). С. 261–279.
Gildenkov M.Yu., 2000. A Review of Palaearctic Species of the Genus Thinodromus (Coleoptera, Staphylinidae): Communication 2 // Entomological Review. V. 80. № 7. P. 831–842.
Gildenkov M.Yu., 2017. A Review of the Thinodromus lunatus Species-Group (Coleoptera, Staphylinidae) // Entomological Review. V. 97. № 8. P. 1089–1105.
Schülke M., Smetana A., 2015. Oxytelinae // Löbl I., Löbl D. (edit.) Catalogue of Palaearctic Coleoptera. V. 1 (Hydrophiloidea – Staphylinoidea). Leiden–Boston: Brill. P. 757–795.
Дополнительные материалы отсутствуют.
Инструменты
Зоологический журнал